Effects of Dietary Zearalenone Adsorbent on the Distribution and Expression of LC3 and PCNA in the Uterus of Gilts
HUANG LiBo,1, WANG JinQuan,2, GAO WenBo1, CHEN HongJu1, HOU YanMeng1, YUAN XueJun,3, WANG ChunYang,11Department of Animal Science and Veterinary Medicine, Shandong Agricultural University, Tai’an 271018, Shandong 2Feed Research Institute, Chinese Academy of Agricultural Sciences, Beijing 100081 3Department of Life Sciences, Shandong Agricultural University, Tai’an 271018, Shandong
Abstract 【Objective】 It has been proved that zearalenone (ZEA) can activate estrogen sensitive gene through estrogen receptor in animals, resulting in reproductive toxicity, and affecting endometrial cell growth, oocyte maturation and follicular granulosa cell proliferation. The purpose of this experiment was to investigate the effects of ZEA adsorbent (zeolite + montmorillonite combinations) on distribution and expression of LC3 and PCNA in the uterus of gilts, and to discuss the detoxification effect of the new ZEA adsorbent from the perspective of histochemistry. 【Method】 A total of 48 healthy gilts with a body weight of 30±2.11 kg were randomly divided into 6 groups (n = 8): control group with basal diet, the ZEA group with basal diet + 1.008 mg∙kg-1ZEA, the 0.1% ZEA adsorbent (ZEA 0.1) group with ZEA diet + 1.0 g∙kg-1 new adsorbent, the 0.25% ZEA adsorbent (ZEA 0.25) group with ZEA diet + 2.5 g∙kg-1 new adsorbent, the 0.5% ZEA adsorbent (ZEA 0.1) group with ZEA diet + 5.0 g∙kg-1 new adsorbent, and the ZEA montmorillonite (ZEA+M) group with ZEA diet + 2.5 g∙kg-1 montmorillonite. The preliminary trail period was 7 d, and the trial period was 21 d. 【Result】 ZEA increased the uterine organ index, and adding 0.25% and 0.5% adsorbents in the diet decreased the index of uterine organs obviously. The LC3 and PCNA positive cells were mainly distributed in the glandular epithelial cells, and the LC3 immunoreactivity of lumen epithelial cells were weaker than that of glandular epithelial cells. The immunoreactivity of LC3 in control group was stronger than that of ZEA group. The immunoreactivity and number of positive cells of LC3 in the ZEA new adsorbent group were significantly higher than those in the ZEA group, and there was a dose-dependent trend, but the effect of the ZEA-M group was increased slightly more than that of ZEA group. The results of PCNA in the luminal and glandular epithelium were contrary to that of LC3. The WB and qRT-PCR results also showed that 0.25% and 0.5% new adsorbents could promote the expression of LC3, increase autophagy, and decrease the PCNA immunoreactive reaction. These results indicated that ZEA inhibited the expression of LC3, inhibited autophagy and destroyed the homeostasis of endometrial cells and glandular epithelial cells. However, the new adsorbent could increase the expression of LC3 protein and decrease the expression of PCNA, which could promote autophagy and resist the abnormal proliferation of endometrial cells caused by ZEA. The new adsorbent had a good protective effect on uterine cells through bidirectional regulation.The experimental results provided theoretical basis for the further application of the new adsorbent. 【Conclusion】In this experiment, ZEA induced the uterus proliferation reaction, the new adsorbent (zeolite + montmorillonite combinations) resisted the negative effect of ZEA on the normal physiological function of uterus within a certain limits. As a result, 0.25% and 0.5% doses of this newadsorbents was suitable. Keywords:zearalenone;uterus;LC3;PCNA;gilts;adsorbent
PDF (5193KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 黄丽波, 王金全, 高文博, 陈红菊, 侯衍猛, 袁学军, 王春阳. 玉米赤霉烯酮吸附剂对后备母猪子宫中LC3、PCNA分布和表达的影响. 中国农业科学, 2021, 54(18): 4008-4017 doi:10.3864/j.issn.0578-1752.2021.18.018 HUANG LiBo, WANG JinQuan, GAO WenBo, CHEN HongJu, HOU YanMeng, YUAN XueJun, WANG ChunYang. Effects of Dietary Zearalenone Adsorbent on the Distribution and Expression of LC3 and PCNA in the Uterus of Gilts. Scientia Acricultura Sinica, 2021, 54(18): 4008-4017 doi:10.3864/j.issn.0578-1752.2021.18.018
A1-A4:对照组;B1-B4:ZEA组;C1-C4:ZEA0.1组;D1-D4:ZEA0.25组;E1-E4:ZEA0.5组;F1-F4:ZEA-M组。A3-F3:子宫腔上皮;A4-F4:子宫腺;比例尺:A1-F1:200μm;A2-F2:20μm;A3-F3和A4-F4:10μm。图2同 Fig. 1Distribution of LC3 immunoreactive cells in uterus of gilts
A1-A4: Control; B1-B4:ZEA; C1-C4: ZEA0.1; D1-D4: ZEA0.25; E1-E4:ZEA0.5; F1-F4:ZEA-M. A3-F3: Luminal epithelium; A4-F4: Uterine gland. Bar: A1-F1:200μm; A2-F2:20μm; A3-F3 and A4-F4:10μm. The same as Fig. 2
不同小写字母者表示差异显著(P<0.05),不同大写字母者表示差异极显著 (P<0.01)。n=3 Fig. 3Statistical results of immunoreactivity iod; protein and mRNA expression of LC3 in uterus
Different lowercase letters marked on the column show significant differences at P<0.05, and different capital letters mean significant differences at P<0.01. n=3
不同小写字母者表示差异显著(P<0.05),不同大写字母者表示差异极显著 (P<0.01)。n=3 Fig. 4statistical results of immunoreactivity iod; protein and mRNA expression of PCNA in uterus
Different lowercase letters marked on the column show significant differences at P<0.05; and different capital letters mean significant differences at P<0.01. n=3
GREEN ML, DIEKMAN MA, MALAYER JR, SCHEIDT AB, LONG GG. Effect of prepubertal consumption of Zearalenone on puberty and subsequent reproduction of gilts Journal of Animal Science, 1990, 68(1):171-178. DOI:10.2527/1990.681171xURL [本文引用: 2]
YANG LJ, ZHOUM, HUANG LB, YANG WR, YANG ZB, JIANG SZ, GE JS. Zearalenone-promoted follicle growth through modulation of wnt-1/β-catenin signaling pathway and expression of estrogen receptor genes in ovaries of postweaning piglets Journal of Agricultural and Food Chemistry, 2018, 66(30):7899-7906. DOI:10.1021/acs.jafc.8b02101URL [本文引用: 4]
ZHOUM, YANG LJ, SHAO MH, WANG YX, YANG WR, HUANG LB, ZHOU XM, JIANG SZ, YANG ZB. Effects of zearalenone exposure on the TGF-β1/Smad3 signaling pathway and the expression of proliferation or apoptosis related genes of post-weaning gilts Toxins, 2018, 10(2):49-61. DOI:10.3390/toxins10020049URL [本文引用: 3]
DAI ML, JIANG SZ, YUAN XJ, YANG WR, YANG ZB, HUANG LB. Effects of Zearalenone-diet on expression of ghrelin and PCNA genes in ovaries of post-weaning piglets Animal Reproduction Science, 2016, 168:126-137. DOI:10.1016/j.anireprosci.2016.03.006URL [本文引用: 5]
YOUNG LG, KING GJ. Low concentrations of Zearalenone in diets of mature gilts Journal of Animal Science, 1986, 63(4):1191-1196. DOI:10.2527/jas1986.6341191xURL [本文引用: 1]
MINERVINIF, DELL'AQUILAM E. Zearalenone and reproductive function in farm animals International Journal of Molecular Sciences, 2008, 9(12):2570-2584. DOI:10.3390/ijms9122570URL [本文引用: 1]
RAINEY MR, TUBBS RC, BENNETT LW, COXNM. Prepubertal exposure to dietary zearalenone alters hypothalamo-hypophysial function but does not impair post pubertal reproductive function of gilts Journal of Animal Science, 1990, 68(7):2015-2022. DOI:10.2527/1990.6872015xURL [本文引用: 1]
EDWARDSS, CANTLEY TC, DAY BN. The effects of Zearalenone on reproduction in swine. II. The effect on puberty attainment and postweaning rebreeding performance Theriogenology, 1987, 28(1):51-58. DOI:10.1016/0093-691X(87)90185-3URL [本文引用: 1]
YANG JY, ZHANG YF, WANG YQ, CUIS. Toxic effects of Zearalenone and alpha-Zearalenol on the regulation of steroidogenesis and testosterone production in mouse Leydig cells Toxicology in Vitro, 2007, 21(4):558-565. DOI:10.1016/j.tiv.2006.10.013URL [本文引用: 1]
GAO YS, ZHAOY, ZHANG HF, ZHANG PF, LIUJ, FENG YN, MEN YH, LIL, SHENW, SUN ZY, MIN LJ. Pubertal exposure to low doses of Zearalenone disrupting spermatogenesis through ERα related genetic and epigenetic pathways Toxicology Letters, 2019, 315:31-38. DOI:10.1016/j.toxlet.2019.08.007URL [本文引用: 2]
ABID-ESSEFIS, BAUDRIMONTI, HASSENW, OUANESZ, MOBIO TA, ANANER, CREPPY EE, BACHAH. DNA fragmentation, apoptosis and cell cycle arrest induced by Zearalenone in cultured DOK, Vero and Caco-2 cells: prevention by Vitamin E Toxicology, 2003, 192(2/3):237-248. DOI:10.1016/S0300-483X(03)00329-9URL [本文引用: 2]
ZINEDINEA, SORIANO JM, MOLTO JC, MAñESJ. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin Food and Chemical Toxicology, 2007, 45(1):1-18. DOI:10.1016/j.fct.2006.07.030URL [本文引用: 1]
ITOY, OHTSUBOK. Effects of neonatal administration of zearalenone on the reproductive physiology of female mice Journal of Veterinary Medical Science, 1994, 56(6):1155-1159. DOI:10.1292/jvms.56.1155URL [本文引用: 1]
PARVEENM, ZHUY, KIYAMAR. Expression profiling of the genes responding to Zearalenone and its analogues using estrogen- responsive genes FEBS Letters, 2009, 583(14):2377-2384. DOI:10.1016/j.febslet.2009.06.035URL [本文引用: 1]
ZHENG WL, WANG BJ, SI MX, ZOUH, SONG RL, GU JH, YUANY, LIU XZ, ZHU GQ, BAI JF, BIAN JC, LIU ZP. Zearalenone altered the cytoskeletal structure via ER stress- autophagy-oxidative stress pathway in mouse TM4 Sertoli cells Scientific Reports, 2018, 8(1):3320. DOI:10.1038/s41598-018-21567-8URL [本文引用: 1]
TIEMANNU, TOMEKW, SCHNEIDERF, VANSELOWJ. Effects of the mycotoxins alpha- and beta-zearalenol on regulation of progesterone synthesis in cultured granulose cells from porcine ovaries Reproductive Toxicology, 2003, 17(6):673-681. DOI:10.1016/j.reprotox.2003.07.001URL [本文引用: 1]
WILEY AA, KAUFFOLDJ, WäHNERM, CREAN-HARRISB, MILLERD J, BAGNELLC A, BARTOLF F. Laser microdissection of neonatal porcine endometrium for tissue-specific valuation of relaxin receptor (RXFP1) expression in response to perinatal zearalenone exposure Annals of the New York Academy of Sciences, 2009, 1160:190-191. DOI:10.1111/j.1749-6632.2008.03821.xURL [本文引用: 1]
ABID-ESSEFIS, OUANESZ, HASSENW, BAUDRIMONTI, CREPPYE, BACHAH. Cytotoxicity, inhibition of DNA and protein syntheses and oxidative damage in cultured cells exposed to Zearalenone Toxicology in Vitro, 2004, 18(4):467-474. DOI:10.1016/j.tiv.2003.12.011URL [本文引用: 2]
CHOIS, SHINH, SONGH, LIM HJ. Suppression of autophagic activation in the mouse uterus by estrogen and progesterone The Journal of Endocrinology, 2014, 221(1):39-50. DOI:10.1530/JOE-13-0449URL [本文引用: 2]
WEAVER AC, SEE MT, KIM SW. Protective effect of two yeast based feed additives on pigs chronically exposed to deoxynivalenol and Zearalenone Toxins, 2014, 6(12):3336-3353. DOI:10.3390/toxins6123336URL
ZHANGY, GAOR, LIUM, SHIB, SHANA, CHENGB. Use of modified halloysite nanotubes in the feed reduces the toxic effects of zearalenone on sow reproduction and piglet development Theriogenology, 2015, 83(5):932-941. DOI:10.1016/j.theriogenology.2014.11.027URL [本文引用: 1]
HECKMANN BL, GREEND R. LC3-associated phagocytosis at a glance Journal Cell Science, 2019, 132(5). pii: jcs222984. doi: 10.1242/jcs.222984. [本文引用: 1]
AMÂLINEIC, PÂVÂLEANUI, GRIGORASA, CÂRUNTU ID, GIUSCÂ SE, AVÂDÂNEI ER, LOZNEANUL, BALAN RA. The endometrial regeneration frontiers: from mechanisms to applications in regenerative medicine Romanian Journal of Morphology and Embryology, 2018, 59(2):407-425. [本文引用: 1]
CHOIJ, JOM, LEEE, OH YK, CHOID. The role of autophagy in human endometrium Biology of Reproduction, 2012, 86(3):70. [本文引用: 1]
LIUY, CHANG JA, WANGP, YIN QQ, HUANG WW, LIU CQ, BAI XX, ZHUQ, GAO TZ, ZHOUP. Effects of Saccharomyces cerevisiae on alleviating cytotoxicity of porcine jejunal epithelia cells induced by deoxynivalenol AMB Express, 2019, 9(1):137. DOI:10.1186/s13568-019-0863-9URL [本文引用: 1]
JAKOPOVIĆŽ, HANOUSEK ČIČAK, MRVČIĆJ, PUCIĆI, ČANAKI, JADRANKAF, PLEADINJ, STANZERD, ZJALIĆS, MARKOVK, JAKOPOVIĆŽ., MRVČIĆJ. Properties and fermentation activity of industrial yeasts saccharomyces cerevisiae, S.uvarum, Candida Utilis and Kluyveromyces marxianus exposed to AFB1, OTA and ZEA. Food Technology Biotechnology, 2018, 56(2):208-217. [本文引用: 1]
JIANG SZ, YANG ZB, YANG WR, WANG SJ, WANGY, BROOMHEADJ, JOHNSTON SL, CHIF. Effect on hepatonephric organs, serum metabolites and oxidative stress in post-weaning piglets fed purified Zearalenone-contaminated diets with or without Calibrin-Z Journal of Animal Physiology and Animal Nutrition, 2012, 96(6):1147-1156. DOI:10.1111/jpn.2012.96.issue-6URL [本文引用: 1]
LIUY, QIN XY, LU XF. Crocin improves endometriosis by inhibiting cell proliferation and the release of inflammatory factors Biomedicine & Pharmacotherapy, 2018, 106:1678-1685. DOI:10.1016/j.biopha.2018.07.108URL [本文引用: 1]
 ,1, 王金全
,1, 王金全

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT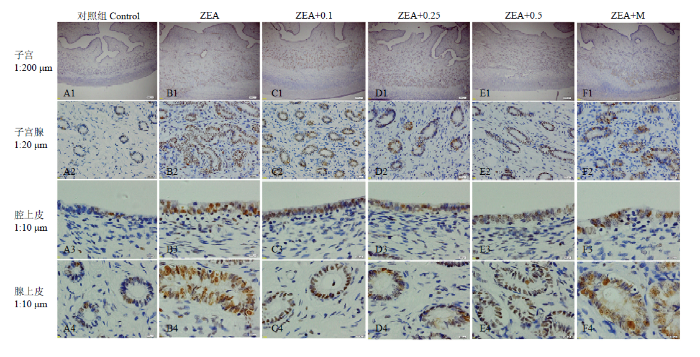 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT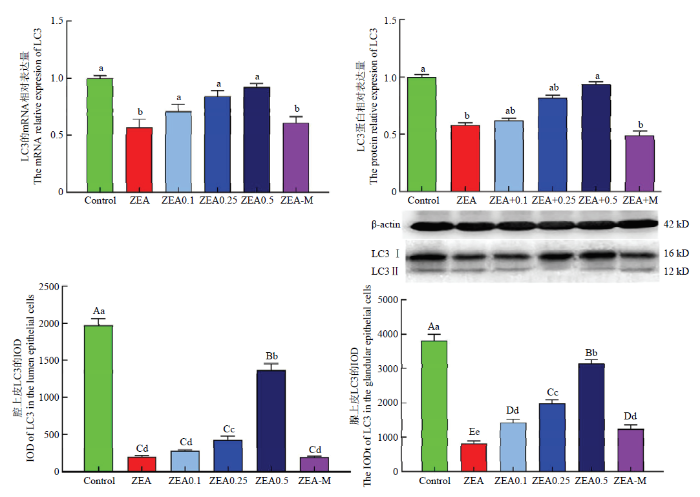 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT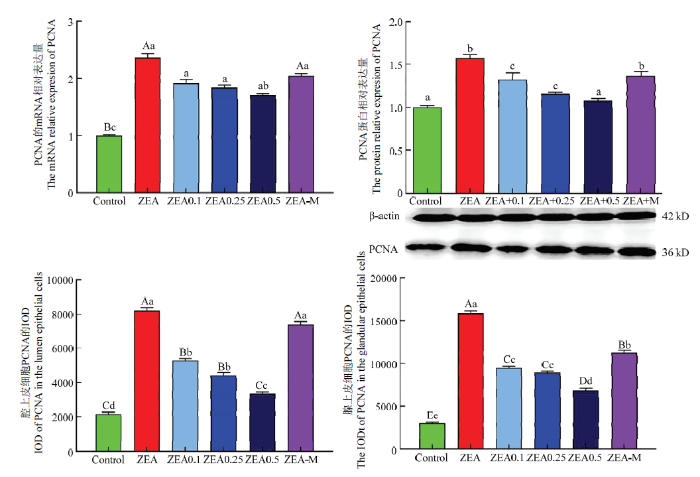 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}