 ,, 刘东尧, 贾绪存, 杨琴, 陈艺博, 董朋飞, 王群,河南农业大学农学院/省部共建小麦玉米作物学国家重点实验室,郑州 450046
,, 刘东尧, 贾绪存, 杨琴, 陈艺博, 董朋飞, 王群,河南农业大学农学院/省部共建小麦玉米作物学国家重点实验室,郑州 450046Maize Tassel Development, Physiological Traits and Yield Under Heat and Drought Stress During Flowering Stage
YAN ZhenHua,, LIU DongYao, JIA XuCun, YANG Qin, CHEN YiBo, DONG PengFei, WANG Qun,College of Agronomy, Henan Agricultural University/National Key Laboratory of Wheat and Maize Crop Science, Zhengzhou 450046通讯作者:
责任编辑: 杨鑫浩
收稿日期:2020-09-3接受日期:2021-01-5
| 基金资助: |
Received:2020-09-3Accepted:2021-01-5
作者简介 About authors
闫振华,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1980KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
闫振华, 刘东尧, 贾绪存, 杨琴, 陈艺博, 董朋飞, 王群. 花期高温干旱对玉米雄穗发育、生理特性和产量影响. 中国农业科学, 2021, 54(17): 3592-3608 doi:10.3864/j.issn.0578-1752.2021.17.004
YAN ZhenHua, LIU DongYao, JIA XuCun, YANG Qin, CHEN YiBo, DONG PengFei, WANG Qun.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】玉米是世界重要的粮食和饲料作物,是我国第一大粮食作物,在保障国家粮食安全中有着不可替代的作用[1]。2019年IPCC报告指出,随着全球气候变暖,未来极端高温天气出现频率将从过去的3.5%增加到4.5%以上,且中国北方未来暖干化趋势越来越明显[2,3]。黄淮海平原是我国重要的夏玉米产区,近年来在玉米生育期内(6—9月)>35℃高温天气出现频率呈逐年上升趋势,尤其以玉米花期(7—8月份)最为明显[4],且高温与干旱时常相伴相随[5],严重影响着玉米结实率和产量。【前人研究进展】研究表明,玉米花期高温、干旱胁迫导致玉米叶片净光合速率和气孔导度下降,叶片中光合产物不同程度降低[6,7,8]。高温、干旱胁迫对玉米生殖生长的影响远远大于对营养生长的影响,对雄配子(花粉粒)生长发育的影响大于雌配子[9]。玉米抽穗至开花期是对高温最敏感的时期,此期高温热害极易引起花粉败育,无法正常开花散粉,致使雌穗不能受精而形成空粒,结实率大幅下降,玉米严重减产[10]。花粉粒的膨大是花药开裂的原初动力之一,高温、干旱胁迫阻碍了花粉粒吸水膨胀过程[11],花药开裂不全或不开裂,花粉发育异常,影响花粉正常生理功能,使产量下降[12]。研究发现,高温胁迫使水稻花药药隔结构解体,绒粘层细胞排列混乱,最终导致花粉败育[13,14]。高温和干旱胁迫均会使玉米雄穗分枝数减少,雄穗主轴长缩短且低于顶叶,形成封闭桶,致使桶内形成无效花粉或花粉量不足[15]。高温、干旱对植株生理特性也有显著影响,逆境使植株体内大量ROS积累,ROS通过激活植株体内的抗氧化系统以调控其对逆境胁迫的响应[16,17,18,19]。研究表明,对花期玉米植株增温处理3—8 d后,植株中ROS积累,超氧化物歧化酶、过氧化物酶和过氧化氢酶活性显著升高,雄穗中可溶性糖和可溶性蛋白含量呈增加趋势[20]。而花期干旱胁迫亦导致玉米叶片抗氧化酶活性、细胞渗透调节物含量不同程度上升,源库平衡被破坏[21]。作物生殖器官对逆境胁迫也会产生反馈效应,当花粉感知高温或干旱信号后,可通过质膜上的钙离子通道、未折叠蛋白反应、活性氧的积累以及含有组蛋白H3的核小体等途径实现对逆境胁迫的响应和反馈[22,23,24,25]。相对单一胁迫而言,复合胁迫加剧了逆境对作物形态和生理特性的影响,并协同降低作物产量和生产力[26];但是,复合胁迫对作物的影响并不是单一胁迫的简单相加效应,且很少能够从单一胁迫中预测出来其影响力度[27,28,29]。【本研究切入点】过去大量研究集中在单一胁迫对玉米穗发育、生长和产量的影响,而对复合胁迫下玉米雄穗生长发育、生理特征及二者复合叠加效应研究较少。当前夏玉米实际生产中,复合胁迫如高温和干旱往往是相伴或相继发生,尤其在黄淮海夏玉米区高温干旱叠加多发生于抽雄至散粉期(7月下旬至8月上中旬),即玉米穗发育和散粉关键时期,二者已成为限制玉米产量形成的2个重要的非生物逆境。因此,研究玉米花期高温干旱复合胁迫对玉米雄穗发育、花粉结构形态及产量形成的影响,对保障夏玉米生长和保证粮食稳定供应具有重要意义。【拟解决的关键问题】本研究通过人工智能温室模拟玉米花期高温、干旱及复合胁迫,采用盆栽试验,研究花期高温、干旱及其复合胁迫对玉米雄穗的形态、结构、生理特性的影响,明确二者复合胁迫的交互效应,以期为夏玉米抗逆稳产提供理论依据和技术支撑。1 材料与方法
1.1 试验设计与材料
试验于2018—2019年在河南农业大学西平试验基地人工智能温室内进行,采用盆栽试验,设置4个处理,分别为CK(对照),T(花期高温),D(花期干旱),TD(花期高温干旱),其中T处理采用人工智能温室控制温度,温室温度比外界对照温度高4℃,且保证白天温度始终处于35℃以上,达到高温胁迫要求,其他条件与对照保持一致;D处理采用透光率95%以上的PO塑料薄膜搭建的旱作棚进行干旱处理,旱作棚顶部与玉米冠层保持2 m间距,保持旱作棚内与对照温度一致,干旱胁迫处理于正式胁迫处理开始前2—3 d进行水分控制预处理,并使用水分测定仪实时监测水分变化,确保在处理当天干旱处理的水分控制在田间最大持水量的60%—65%,且胁迫处理期间实时监测水分状况,根据每盆装土重量计算需要补充的水分量,以保持胁迫处理期间土壤含水量为田间最大持水量的60%—65%;TD处理在人工智能控制温室内进行,保持温室温度比外界对照温度高4℃,白天温度能保持在35℃以上,达到高温胁迫,即与T处理温度控制一致,同时使用水分测定仪实时监测水分,使土壤含水量控制在土壤最大持水量的60%—65%,与D处理的水分控制一致。各胁迫处理均于玉米雄穗露出顶叶2—3 cm时开始进行,处理时间为14 d,之后将盆栽转移至正常大田条件生长直至成熟。供试材料为本课题组前期筛选的热敏感型品种华农138(HN138)和耐热较好的郑单958(ZD958)品种,试验土壤类型为砂姜黑土,取0—20 cm的耕层土壤,风干、过筛、装盆,盆体上口直径为35 cm,下底部直径为25 cm,高30 cm,每盆装土15 kg;土壤基础养分为有机质含量17.8 g·kg-1、碱解氮含量为106.86 mg·kg-1、速效磷含量为52.47 mg·kg-1、速效钾含量为229.03 mg·kg-1。1.2 测定项目与方法
1.2.1 田间小气候监测 利用人工智能温室自动记录系统和小型气象站监测记录处理期间日均温和日最高温度的变化,采用BY-TWS型便携式土壤温湿度速测仪实时测定和记录土壤含水量(图1),保证D处理与TD处理土壤的水分含量的一致性,T处理与TD处理期间平均温度和日最高温一致。图1
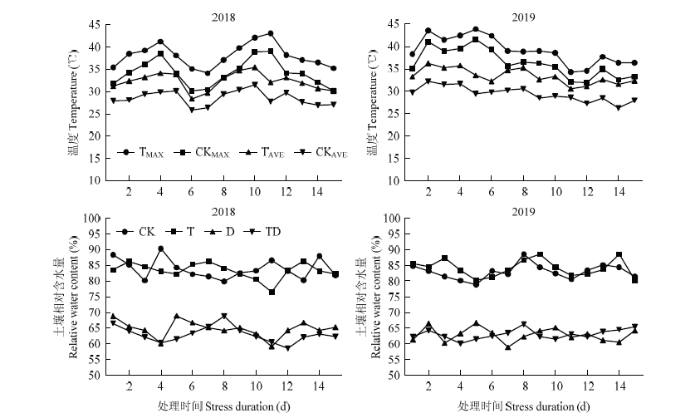 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同胁迫处理下温度和土壤相对含水量的变化
CK:对照;TMAX:高温胁迫日最高温度;CKMAX:对照日最高温度;TAVE:高温胁迫日平均温度;CKAVE:对照日平均温度;T:花期高温胁迫;D:花期干旱胁迫;TD:花期高温干旱复合胁迫。下同
Fig. 1Changes of daily temperature and relative water content under different stress treatments
CK: Control, TMAX: Daily maximum temperature under high temperature stress, CKMAX: Daily maximum temperature under control, TAVE: Daily average temperature under high temperature stress, CKAVE: Daily average temperature under control, T: Flowering high-temperature stress treatment, D: Flowering drought stress treatment, TD: Flowering high-temperature-drought combined stress treatment. The same as below
1.2.2 玉米雄穗形态观察 在雄穗开花后第3天,各处理选代表性植株5株取其雄穗,测量和记录雄穗主轴和分枝的长度。以雄穗最下部的分枝基部为节点,从该节点到雄穗主轴顶端的距离,为雄穗主轴长度;每个分枝基部到顶端的距离为分枝长度,雄穗主轴的最下部到雄穗顶端的小花数量为主轴小花数,每个分枝底部到分枝顶端的小花数量为该分枝小花数,分枝底部到分枝顶端的长度为分枝长度,分枝小花密度=分枝长度/分枝小花数。
1.2.3 花药、花粉粒形态结构观察和花粉活力测定 各胁迫处理期间,采用雄穗套袋方法,在玉米抽雄后第5—6天(玉米雄穗盛花期)取新鲜花药和花粉粒分3份,1份花药采用番红固绿法[30]制作切片,之后在OLYMPUS BX43显微镜下镜检观察,并进行拍照和采集图像。1份新鲜花粉粒用2.5%电镜固定液固定处理,用0.1 mol·L-1磷酸缓冲液洗3次,超纯水冲洗2次,乙醇梯度脱水后浸入叔丁醇保存,使用Quorum K850临界点干燥仪进行干燥,真空喷镀,用日产S3400N型扫描电镜进行观察、拍照。第3份新鲜花粉粒采用TTC染色法[30]测定花粉活力,根据花粉粒染色后的颜色,将花粉活力分为3类:一类是高活力花粉粒,即染色后颜色为深红色或鲜红色的花粉粒;二类是无活力花粉粒,即染色后呈现黄褐色的花粉粒;三类是低活力花粉粒,染色介于深红或鲜红色与黄褐色之间的浅粉或浅红色的花粉粒。
1.2.4 单株散粉量测定 在玉米雄穗小花开放前,各个处理选取长势一致的5株玉米进行雄穗套袋处理,于每天下午16:00收集花粉,去除杂物后过筛、称重,为单日单株散粉量,连续每天取样直至散粉结束。
1.2.5 雄穗生理指标测定 从胁迫处理开始,每2 d取各个处理的雄穗中部3 g左右,用液氮速冻后放置于-80℃超低温冰箱中保存,测定抗氧化酶活性和过氧化产物含量,其中SOD活性采用氮蓝四唑法(NBT)法测定,POD活性采用愈创木酚显色法测定,MDA含量采用硫代巴比妥酸(TBA)法测定,$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量采用羟胺氧化法测定[31]。
1.2.6 考种计产 在玉米成熟时,各处理取10穗测量其穗长、穗粗、秃尖长、穗行数、行粒数和百粒重,并折合14%的水分进行计产。
1.2.7 统计分析 试验数据采用Excel 2010进行数据处理,利用SPSS.26软件进行数据分析,采用GraphPad Prism 8进行作图。
2 结果
2.1 不同胁迫处理下玉米雄穗农艺性状的变化特征
胁迫处理下玉米雄穗农艺性状受胁迫程度表现为高温干旱复合胁迫>干旱胁迫>高温胁迫>对照(表1)。受胁迫影响玉米雄穗主轴长、主轴小花数、分枝数以及分枝小花密度均低于对照,其中D、TD处理与CK间差异显著。T、D和TD胁迫处理下雄穗主轴长比CK平均降低11.86%、23.64%和27.51%,主轴小花数比CK平均降低15.70%、24.87%和32.02%,分枝数和分枝小花密度则比CK分别降低17.31%、33.85%、45.59%和13.56%、27.08%、26.00%。2个品种比较,ZD958雄穗是短轴多分枝类型,主轴长度平均为38.33 cm,分枝数平均为16.32个,而HN138属于长轴少分枝类型,主轴长度平均为44.37 cm,分枝数平均4.40个。在2018高温年份中,T、D和TD处理下HN138的雄穗主轴长与分枝数分别比CK降低20.71%、25.57%、30.30%和14.00%、34.25%和50.00%,胁迫处理与对照差异显著,而ZD958主轴长和雄穗分枝数则比CK分别减少17.57%(T)、31.08%(D)、36.49%(TD)和17.57%(T)、31.08%(D)、36.49%(TD);HN138的T、D和TD处理主轴小花数比CK降低15.23%、22.62%和32.65%,分枝小花密度比CK降低13.01%、28.10%和22.96%,而ZD958各处理主轴小花数和分枝小花密度比CK分别降低8.89%、18.01%、26.43%和14.10%、26.06%、29.05%。各指标除小花密度外,均表现为逆境胁迫下HN138降幅显著高于ZD958,且各胁迫处理与对照间、以及单一胁迫与复合胁迫间差异均达显著水平。Table 1
表1
表1不同胁迫处理对玉米雄穗农艺性状的影响
Table 1
| 年份 Year | 品种 Cultivar | 处理 Treatment | 主轴长 Tassel central branch length (cm) | 主轴小花数 Tassel central branch spikelet number | 分枝数 Tassel branch number | 分枝小花密度 Tassel branch spikelet density (count/cm) |
|---|---|---|---|---|---|---|
| 2018 | ZD958 | CK | 38.40±1.06a | 268.44±21.46a | 14.84±2.04a | 3.14±0.59a |
| T | 38.38±2.70a | 256.42±20.07ab | 12.21±1.60b | 3.02±0.45b | ||
| D | 30.52±1.81b | 227.24±12.65bc | 10.2±1.60bc | 2.73±0.38c | ||
| TD | 28.78±2.75b | 197.47±25.55c | 9.43±1.62c | 2.62±0.23c | ||
| HN138 | CK | 42.78±3.15a | 193.69±10.09a | 4.00±1.26a | 2.44±0.17a | |
| T | 33.92±2.07b | 153.88±12.46ab | 3.44±0.80ab | 2.29±0.32b | ||
| D | 31.84±1.79c | 138.15±9.23bc | 2.63±0.80ab | 1.77±0.28c | ||
| TD | 29.82±1.60c | 130.45±17.58c | 2.00±0.89b | 2.26±0.31b | ||
| 2019 | ZD958 | CK | 38.25±4.64a | 309.60±14.89a | 17.80±1.33a | 4.02±0.14a |
| T | 36.51±2.27a | 268.40±10.03b | 14.80±1.47b | 3.04±0.22b | ||
| D | 30.35±2.26b | 245.60±35.67b | 12.00±1.10c | 2.45±0.15c | ||
| TD | 29.79±1.32b | 198.80±9.66c | 9.60±1.36d | 2.35±0.28c | ||
| HN138 | CK | 45.96±1.28a | 220.40±8.59a | 4.80±1.72a | 3.27±0.48a | |
| T | 35.79±0.57b | 166.60±3.44b | 3.80±0.98a | 2.62±0.18a | ||
| D | 33.18±0.39b | 143.80±4.35c | 3.00±1.10b | 2.33±0.15ab | ||
| TD | 30.99±1.25c | 147.20±9.27d | 2.40±1.02b | 2.01±0.15b | ||
| 变异来源 Source of variation | 年份 Year | NS | NS | NS | NS | |
| 品种 Cultivar | ** | ** | ** | ** | ||
| 处理 treatment | ** | ** | ** | ** | ||
| 年份×品种 Year×Cultivars | NS | NS | NS | NS | ||
| 年份×处理 Year×Treatment | NS | NS | NS | NS | ||
| 品种×处理 Cultivars×Treatment | ** | ** | * | NS | ||
| 年份×品种×处理 Year×Cultivars×Treatment | NS | NS | NS | NS | ||
新窗口打开|下载CSV
2.2 不同胁迫处理下雄穗花粉活力和日散粉量的变化特征
高温、干旱及复合胁迫下玉米雄穗花粉活力显著下降,染色后在显微镜×40倍视野下观察(图2),各处理高活力花粉粒占比依次为CK>T>D>TD,其中CK(图2-A、图2-E)的花粉粒绝大多数呈现高活力的红色或深红色;T(图2-B、图2-F)和D处理(图2-C、图2-G)花粉粒呈现3种形态,一部分高活力的红色,少部分低活力的浅粉红色,一部分无活力褐色;而TD处理(图2-D、图2-H)绝大多数花粉粒染色后呈现无活力的褐色,少数花粉粒呈现低活力的浅粉红色和高活力的红色,且视野中CK中高活力花粉粒的数量明显多于T、D处理,更多于TD处理。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同胁迫处理对雄穗花粉活力的影响
HA:高活力;LA:低活力;NA:无活力;A:CK-ZD958;B:T-ZD958;C:D-ZD958;D:TD-ZD958;E:CK-HN138;F:T-HN138;G:D-HN138;H:TD-HN138(×40)。同一组不同小写字母表示在0.05水平上差异显著
Fig. 2Effects of different stress treatments on pollen activities of tassel
HA: High activity; LA: Low activity; NA: No activity. A: CK-ZD958; B: T-ZD958; C: D-ZD958; D: TD-ZD958; E: CK-HN138; F: T-HN138; G: D-HN138; H: TD-HN138(×40). Values within a group followed by different letters are significantly different at 0.05 probability level
2年数据来看,T、D和TD处理的高活力花粉粒数量分别占总花粉粒的46.00%、29.75%和22.67%,高活力花粉粒比例比CK降低22.00%、38.25%、45.33%(图2),与CK间差异显著,D、TD处理高活力花粉数量显著低于T处理;而T、D和TD处理无活力花粉比CK显著增加10.50%、33.00%和37.00%,D和TD处理无活力花粉比T处理显著高22.83%、26.83%,T与D、T与TD处理间无活力花粉数量差异显著,但D与TD处理间差异不显著。2个品种比较,在T、D和TD处理下,HN138无活力花粉比例分别为36.50%、52.83%和55.33%,而ZD958无活力花粉比例为19.33%、48.67%和54.17%,表现为HN138高温、干旱胁迫下无活力花粉数量远高于ZD958,而ZD958高活力花粉比例为51.66%,32.33%和21.17%,HN138则分别为40.33%、27.17%和24.17%,表现为ZD958高活力花粉比例高于HN138,尤其在2018年的高温年份表现更为突出。
2年试验结果表明玉米雄穗的日散粉量呈单峰曲线(图3),单株日散粉量最大值出现在开始散粉后的4—5 d,散粉持续时间最长为8 d。胁迫处理使散粉持续时间平均缩短1—2 d,其中TD处理缩短2 d,D处理缩短1 d;散粉峰值出现时间提前1—2 d,处理间表现为TD处理早于D处理早于T处理早于CK,其中T、D和TD处理散粉峰值期比CK平均提早了0.5 d、1.5 d和2.5 d。胁迫减少了雄穗每日散粉量,T、D和TD处理的日散粉量较CK降低22.18%、54.75%和67.28%,且处理间差异显著。不同品种比较,二者散粉峰值期对逆境反应差异明显,CK条件下ZD958与HN138的散粉高峰期分别出现在散粉后第4天与第5天;ZD958的散粉峰值期T处理与CK一致均出现在第4天,D处理比CK提早1 d,TD处理则比CK提早2 d;而HN138日散粉峰值期在T、D、TD处理下比CK分别提早1 d、2 d和3 d。正常条件下ZD958的花粉量比HN138平均多20.89%,胁迫显著减少了雄穗散粉量,其中ZD958的日散花粉量在T、D和TD处理下比CK平均降低21.59%、57.18%和68.46%, HN138则比CK平均降低22.77%、52.33%和66.10%,但两品种间差异不显著。
图3
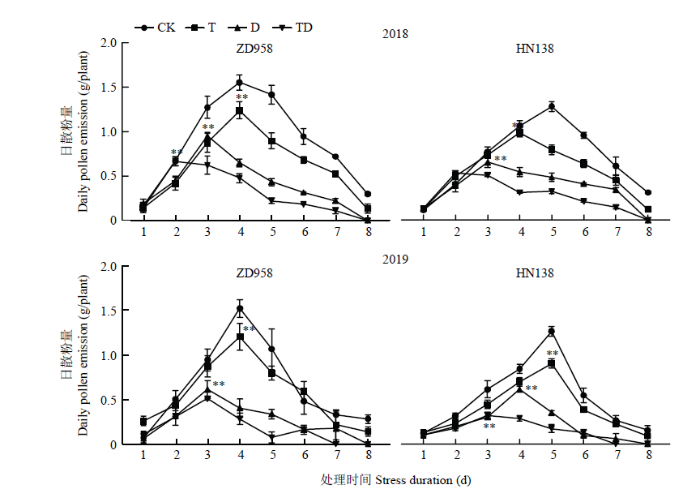 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同胁迫处理对雄穗日散粉量的影响
*,**分别表示不同处理峰值在 0.05 和 0.01 水平差异显著。下同
Fig. 3Effects of different stress treatments on the daily pollen emission of tassel
*,**means the peak values of different treatments are significantly different at 0.05 or 0.01 level. The same as below
2.3 不同胁迫处理下花药结构和花粉粒形态变化
高温、干旱及其复合胁迫影响着雄穗花药结构和花粉粒形态。由图4可以看出,CK的(图4-A1、图4-E1)花药圆润饱满,药隔分明,药室排列整齐,花药壁细胞排列整齐,大小均匀,绒粘层细胞正常,T处理(图4-B1、图4-F1)的花药皱缩变小,药室发生变形,部分花药壁表皮细胞发生畸形,结构排列松散,药隔维管束变小,绒毡层细胞基本正常,D(图4-C1、图4-G1)和TD(图4-D1、图4-H1)处理的花药皱缩变小更为明显,药室变形严重甚至部分断裂,呈现凹陷和褶皱,花药壁表皮细胞出现膨大或缩小,绒毡层细胞退化,药隔维管束变少、变细,花药整体趋近解体。2个品种比较,逆境胁迫下HN138的花药皱缩变小比ZD958更为严重,如T处理下HN138(图4-F1)花药壁出现断裂,而ZD958花药壁尚保持完整;D和TD处理的HN138药室(图4-G1、图4-H1)变形皱缩比ZD958(图4-C1、图4-D1)更为明显,且褶皱多且深,HN138在T和TD处理下(图4-F1、图4-H1)药隔维管束更细,维管束外层薄壁细胞层数的减少和断裂程度亦比ZD958(图4-B1、图4-D1)更明显。图4
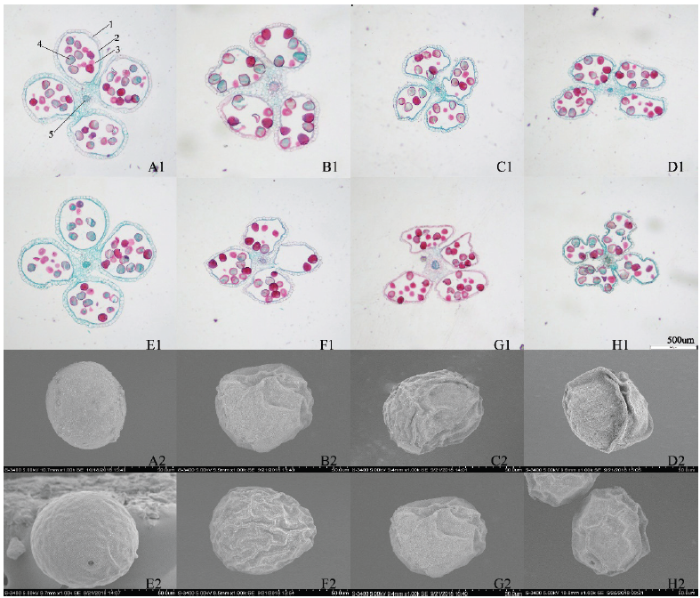 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同胁迫处理对花药结构和花粉粒形态的影响
1:表皮细胞;2:绒毡层细胞;3:药室;4:花粉粒;5:药隔内维管束(×200)。A1:CK-ZD958;B1:T-ZD958;C1:D-ZD958;D1:TD-ZD958;E1:CK-HN138;F1:T-HN138;G1:D-HN138;H1:TD-HN138(×200);A2:CK-ZD958;B2:T-ZD958;C2:D-ZD958;D2:TD-ZD958;E2:CK-HN138;F2:T-HN138;G2:D-HN138;H2:TD-HN138(×1000)
Fig. 4Effects of different stress treatments on anther microstructure and pollen morphology
1: Epiderm cell; 2: Middle layer cell; 3: Anther chamber; 4: Pollen grain; 5: Vascular bundle(×200). A1: CK-ZD958; B1: T-ZD958; C1: D-ZD958; D1: TD-ZD958; E1: CK-HN138; F1: T-HN138; G1: D-HN138; H1: TD-HN138(×200); A2: CK-ZD958; B2: T-ZD958; C2: D-ZD958; D2: TD-ZD958; E2: CK-HN138; F2: T-HN138; G2: D-HN138; H2: TD-HN138(×1000)
电镜扫描观察花粉粒形态发现(图4),CK的(图4-A2、图4-E2)花粉粒外观近似圆形或椭圆,表面光滑且具有清晰且稍微突出的萌发孔;T处理的(图4-B2、图4-F2)花粉表面始现网纹状凸起,但其网纹凸起程度较浅,且褶皱纹路浅且少;D处理(图4-C2、图4-G2)的表面网纹状凸起明显增多,皱缩加剧且褶皱纹路加深,萌发孔轻微凹陷,而TD处理(图4-D2、图4-H2)的表面网纹状更明显,且异常凸起成高脊,皱褶凹陷更深,萌发孔明显皱缩内陷,导致萌发困难。
2.4 不同胁迫处理下雄穗超氧化物歧化酶(SOD)和过氧化物酶(POD)活性的变化
随生育进程推进,各处理玉米雄穗SOD活性呈先升后降的单峰曲线(图5),峰值出现在处理后的第4—8天。胁迫处理初期酶活性增加,显著高于CK,但不同胁迫增加幅度存在差异,T、D和TD处理初期SOD活性平均比CK增加21.91%、24.95%和17.12%,其中D和TD处理的SOD活性最大值比CK显著增加61.21%和72.69%,但峰值之后迅速下降,各处理SOD活性低于对照。2个品种比较,SOD活性达到峰值的日期不同,ZD958品种SOD活性在胁迫后的第6天达到最大,HN138则在第4天达到最大,比ZD958早2 d;T、D和TD处理下ZD958的SOD活性最大值比CK平均高48.85%、82.38%和78.39%, HN138则比CK高61.66%、64.11%和74.99%,且SOD活性峰值在高温年份(2018年)均较正常年份(2019年)提前0—2 d。图5
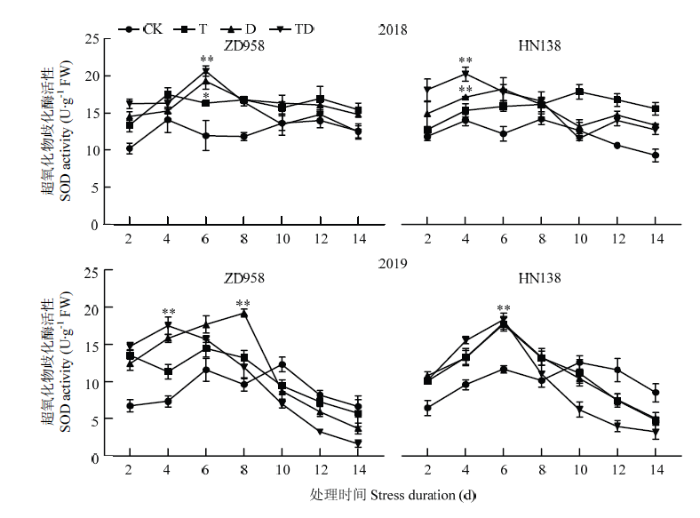 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同胁迫处理对超氧化物歧化酶活性的影响
Fig. 5Effects of different stress treatments on superoxide dismutase activities of tassel
玉米雄穗POD活性与SOD活性变化趋势基本一致(图6),随生育进程呈先上升后下降的单峰曲线,其峰值出现在第6—8天,且胁迫初期的玉米雄穗POD活性显著高于对照,随着胁迫时间延长,8 d后活性明显下降,但依然高于对照。处理间比较,T、D和TD处理的POD活性比CK平均高32.50%、53.37%和67.24%,且与CK差异极显著;且D、TD处理POD活性最大值分别比CK增加108.42%和144.70%,差异极显著。2个品种比较,高温、干旱及复合胁迫下,ZD958品种POD活性最大值出现在处理后的第8天,HN138则出现在处理后的第6—8天;且在T、D和TD处理下ZD958品种的POD活性峰值比CK增加97.21%、130.15%和171.82%,而HN138的POD活性峰值比CK增加40.10%、118.18%和137.23%。
图6
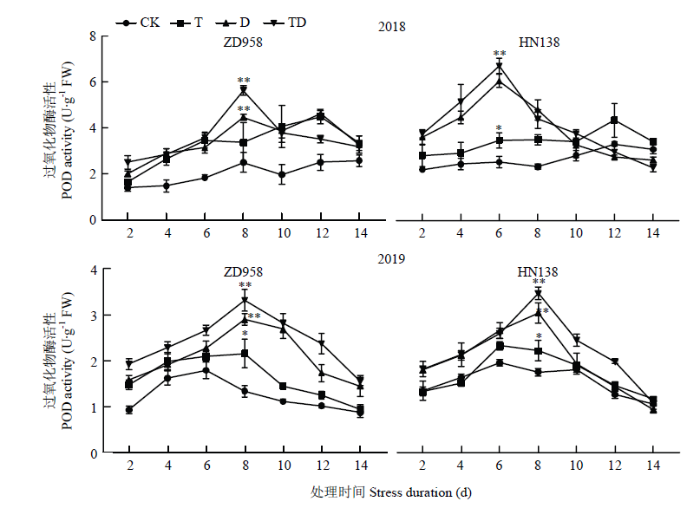 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同胁迫处理对过氧化物酶活性的影响
Fig. 6Effects of different stress treatments on peroxidase activities of tassel
2.5 不同胁迫处理下丙二醛(MDA)含量和超氧阴离子自由基($\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$)含量变化
随着生育进程,各处理MDA含量呈先增后降趋势(图7),且逆境处理下MDA含量最大值出现在处理后6—8 d,之后下降并趋于平缓。高温、干旱及复合胁迫使玉米雄穗MDA含量显著高于对照,其中T、D和TD处理MDA含量比CK平均增加44.18%、64.24%和79.12%。不同胁迫间比较,D、TD处理2 d后,玉米雄穗MDA含量迅速增加,并在6—8 d达到最大值,且两处理MDA含量最大值较CK平均增加94.40%和115.78%;而T处理的MDA含量变化平缓,变化趋势与CK一致,比CK平均增加39.03%。2个品种对逆境胁迫的响应不同,ZD958在TD处理下MDA含量峰值出现在处理后8 d,而HN138的峰值出现在处理后6 d,且在D和T处理下,HN138的MDA含量平均比ZD958高22.61%和16.18%。图7
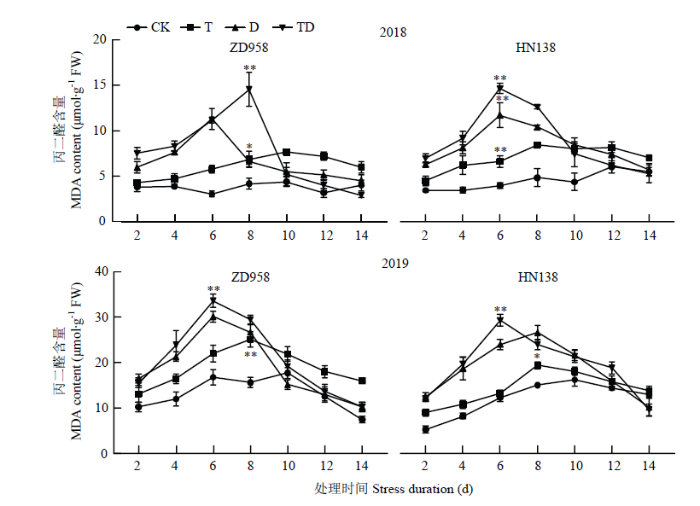 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同胁迫处理对MDA含量的影响
Fig. 7Effect of different stress treatments on MDA content
随生育进程玉米雄穗的$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量与MDA含量变化一致(图8),均呈现先升后降趋势,其最大值出现在处理后的4—8 d。胁迫增加了$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量,T、D和TD处理$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量比CK平均增加22.55%、51.65%和72.29%,差异显著。不同处理在逆境胁迫初期(2—6 d),$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量均急剧增加,但$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量达到峰值时间不同,D和TD处理峰值出现在处理后第4—6天,T处理的峰值出现在处理后第6—8天,D和TD处理的$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量变幅远大于T处理,D和TD处理与CK间$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$最大含量差异极显著。2个品种比较,ZD958的$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量的峰值出现时间较HN138平均滞后1—2 d,且最大值平均比HN138低16.73%。2个品种的$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量峰值出现时间在高温年份(2018年)较正常年份(2019年)平均提前1—2 d。
图8
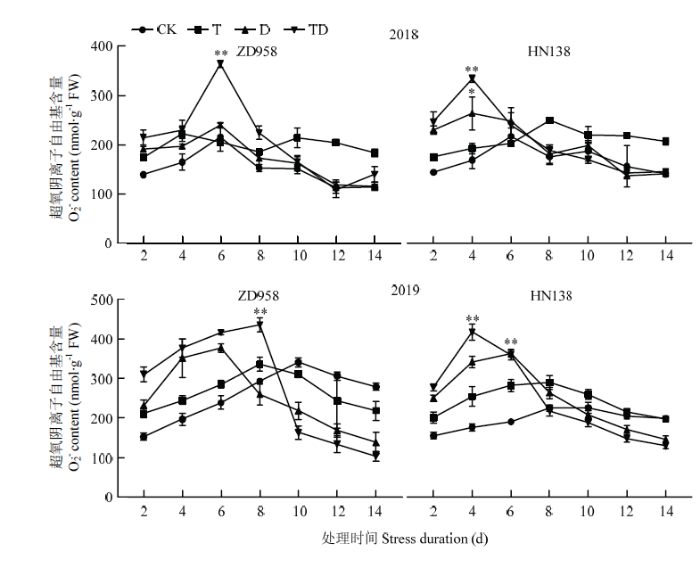 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8不同胁迫处理对超氧阴离子自由基含量的影响
Fig. 8Effects of different stress treatments on superoxide anion radical content
2.6 不同胁迫处理下产量、产量构成因素变化及其相关性
逆境胁迫下玉米穗长、穗粗显著下降,秃尖长显著增加(表2),其中T、D、TD处理穗长和穗粗比CK平均降低8.52%、25.52%、36.07%和2.54%、11.35%、15.34%,秃尖长比CK增加319.95%、410.93%、578.82%。高温、干旱及复合胁迫显著降低了玉米产量、行粒数和千粒重,穗行数受影响较小;T、D和TD处理的产量比CK平均降低18.05%、34.58%和46.24%,行粒数较CK平均减少24.58%、41.80%和52.99%,千粒重比CK平均降低3.01%、6.68%和8.22%,处理间两两差异显著。2个品种比较,高温、干旱及复合胁迫下ZD958的穗长、穗粗、行粒数、产量下降幅度小于HN138,秃尖长显著短于HN138。在T、D、TD处理下,ZD958穗长比CK降低7.05%、25.29%、33.23%,HN138穗长比CK降低9.99%、25.74%、38.92%,HN138秃尖长比ZD958长763.64%、351.39%、71.25%;T和TD处理下ZD958行粒数比HN138多5.34%、1.64%,千粒重分别比HN138高27.38%、24.96%,在T、D、TD处理下ZD958产量较HN138高15.18%、36.14%和40.18%。Table 2
表2
表2不同胁迫处理对产量及其构成要素的影响
Table 2
| 年份 Year | 品种 Cultivars | 处理 Treatment | 穗长 Ear length (cm) | 穗粗 Ear diameter (mm) | 秃尖 Bald tip (mm) | 穗行数 Ear rows | 行粒数 Kernels per row | 千粒重 1000 grain weight (g) | 产量 Yield (kg·hm-2) |
|---|---|---|---|---|---|---|---|---|---|
| 2018 | ZD958 | CK | 15.91±1.12a | 52.12±2.11a | 2.39±2.24c | 17.60±1.20a | 33.40±1.43a | 322.09±5.77a | 12620.21±279.16a |
| T | 14.95±0.65a | 49.66±2.01b | 4.59±0.85c | 17.20±2.04a | 26.70±2.69b | 322.08±5.93a | 9762.19±230.86b | ||
| D | 11.57±1.30b | 45.34±3.05c | 10.01±1.17b | 15.80±2.27a | 17.50±2.91c | 295.49±11.84b | 7700.68±151.23c | ||
| TD | 10.39±0.83c | 42.58±1.75d | 22.64±6.87a | 15.80±1.66a | 15.40±2.24c | 305.45±13.19b | 5809.84±153.80d | ||
| HN138 | CK | 17.48±1.03a | 49.18±1.65a | 6.58±3.48b | 15.40±1.29a | 32.40±2.54a | 261.39±5.15a | 9387.42±170.16a | |
| T | 15.22±0.51b | 49.39±1.04a | 33.99±4.19a | 15.00±1.00a | 24.90±1.97b | 252.25±7.11b | 7677.14±186.88b | ||
| D | 12.92±1.18c | 41.68±3.55b | 32.76±5.23a | 15.20±0.98a | 19.50±1.57c | 240.06±3.50c | 5070.93±139.57c | ||
| TD | 11.11±0.91d | 40.58±2.52b | 30.65±9.76a | 15.00±1.34a | 15.90±1.92d | 239.64±7.38c | 4145.12±179.17d | ||
| 2019 | ZD958 | CK | 15.62±0.32a | 51.41±2.61a | 2.19±1.85a | 16.60±1.56a | 36.00±1.55a | 350.98±12.79a | 12984.97±148.62a |
| T | 14.36±0.25a | 49.04±1.36b | 5.53±0.46a | 15.80±1.40a | 23.90±1.97b | 329.65±16.62ab | 9870.79±182.17b | ||
| D | 11.98±0.97b | 46.24±1.42c | 9.03±0.68b | 14.60±0.92a | 19.20±2.40c | 334.93±18.91a | 9371.21±128.42c | ||
| TD | 10.66±0.52c | 45.69±1.76c | 17.49±1.96c | 14.00±1.26a | 15.00±1.79d | 305.14±19.33b | 8419.67±189.74d | ||
| HN138 | CK | 19.57±1.45a | 49.71±0.89a | 7.43±2.46c | 16.00±1.26a | 29.30±2.72a | 265.86±11.30a | 10153.57±218.83a | |
| T | 18.19±1.72a | 49.08±0.72a | 53.53±16.96b | 15.40±1.28a | 23.00±2.45b | 259.37±6.71b | 9368.37±123.10b | ||
| D | 14.60±1.27b | 46.19±0.75b | 53.24±14.18a | 15.20±1.33a | 19.60±2.46c | 250.63±5.53b | 7469.07±188.68c | ||
| TD | 11.47±1.81c | 42.53±1.90c | 37.92±7.56a | 15.00±1.84a | 15.00±1.41d | 248.98±6.52c | 6005.79±188.81d | ||
| 变异来源 Source of variation | 年份 Year | NS | NS | NS | NS | NS | NS | NS | |
| 品种 Cultivars | ** | ** | ** | ** | * | ** | ** | ||
| 处理 Treatment | ** | ** | ** | ** | ** | ** | ** | ||
| 年份×品种 Year×Cultivars | NS | NS | * | * | * | NS | NS | ||
| 年份×处理 Year×Treatment | NS | NS | NS | NS | * | NS | NS | ||
| 品种×处理 Cultivars×Treatment | ** | ** | ** | NS | * | ** | NS | ||
| 年份×品种×处理 Year×Cultivars×Treatment | NS | NS | NS | NS | * | NS | NS | ||
新窗口打开|下载CSV
2年数据的相关性分析表明(表3),主轴长、主轴小花数、花粉量、花粉活力与产量呈极显著正相关,与穗粒数呈显著正相关,与千粒重呈正相关,但不显著,雄穗分枝数、分枝小花密度与穗粒数、产量呈正相关关系。2个品种在不同年份也存在差异,在高温干旱年份(2018年),ZD958的花粉量、花粉活力与千粒重、产量显著正相关,HN138的花粉量、花粉活力则与产量、穗粒数显著正相关,且HN138分枝数与产量呈显著正相关,ZD958的分枝数与产量呈正相关,但不显著;2019年,ZD958的分枝数与产量、穗粒数、千粒重均显著正相关,HN138的分枝小花密度与产量、穗粒数、千粒重显著正相关。
Table 3
表3
表3不同胁迫处理下产量、产量构成要素与雄穗相关指标的相关分析
Table 3
| 年份 Year | 雄穗相关指标 Tassel-related trait | ZD958 | HN138 | ||||
|---|---|---|---|---|---|---|---|
| 产量 Yield | 穗粒数 Grains per ear | 千粒重 1000 grain weight | 产量 Yield | 穗粒数 Grains per ear | 千粒重 1000 grain weight | ||
| 2018 | 主轴长 Tassel central branch length | 0.7769** | 0.8150** | 0.3344 | 0.7330** | 0.6034* | 0.3704 |
| 主轴小花数 Central branch spikelet number | 0.7283** | 0.7111** | 0.2555 | 0.5745* | 0.5532* | 0.3407 | |
| 分枝数 Tassel branch number | 0.5045 | 0.2377 | 0.4747 | 0.5363* | 0.5928* | 0.3608 | |
| 分枝小花密度Tassel branch spikelet density | 0.4037 | 0.3208 | 0.2157 | 0.0987 | 0.0746 | 0.3723 | |
| 花粉量 Fresh pollen weight | 0.9239** | 0.8321** | 0.5518* | 0.8918** | 0.8639** | 0.6431** | |
| 花粉活力 Pollen activity | 0.8568** | 0.7896 | 0.5887* | 0.6146* | 0.7228** | 0.3597 | |
| 2019 | 主轴长 Central tassel branch length | 0.6070* | 0.5580* | 0.3178 | 0.7899** | 0.6511** | 0.4437 |
| 主轴小花数 Central tassel flower number | 0.7211** | 0.7551** | 0.4336 | 0.7341** | 0.6079* | 0.4898 | |
| 分枝数 Tassel branch number | 0.6441** | 0.5593* | 0.5658* | 0.8242** | 0.7488** | 0.4379 | |
| 分枝小花密度Tassel branch spikelet density | 0.5432* | 0.6943** | 0.3372 | 0.8184** | 0.7597** | 0.5946* | |
| 花粉量 Pollen weight | 0.7325** | 0.7821** | 0.2890 | 0.9314** | 0.7433** | 0.4112 | |
| 花粉活力 Pollen activity | 0.8268** | 0.8279** | 0.4186 | 0.9002** | 0.6874** | 0.4067 | |
新窗口打开|下载CSV
3 讨论
3.1 高温干旱胁迫对雄穗形态特征的影响
逆境胁迫下玉米雄穗的正常发育对花药发育以及花粉量的多少有着至关重要的影响[32]。本研究结果表明,高温胁迫下主轴小花数、分枝小花密度大幅下降,干旱胁迫下雄穗主轴长度、分枝小花密度均显著减少,分枝数减少但差异不显著,而高温干旱胁迫下雄穗主轴长度、分枝数显著降低,同时主轴小花数和分枝小花密度显著减少,表现出既有干旱胁迫导致的雄穗主轴、分枝退化变短,又有高温引发小花数量和分枝小花密度的大幅度减少,且明显表现出了高温干旱胁迫对雄穗生长胁迫的叠加效应,因此导致雄穗花粉总量的骤减,这与前人研究表明结果基本一致[33]。本结果也表明,雄穗主轴长度、分枝小花密度对高温、干旱及复合胁迫均较为敏感,而雄穗分枝数和主轴小花数则对干旱和高温干旱复合胁迫反应更敏感,而受高温影响较小,且二者复合胁迫加剧了分枝小花数和小花密度的减少,呈现出二者的叠加效应。高温敏感型品种HN138具有雄穗主轴长、分枝少的特点,而主轴短、分枝多的品种ZD958对高温、干旱胁迫具有更好的适应性和抗胁迫性。可见玉米雄穗主轴长度、分枝数量及主轴、分枝小花数量可作为判断玉米能否适应花期高温、干旱胁迫的重要指标之一。3.2 高温干旱胁迫对花药结构和花粉活力的影响
玉米花期高温干旱改变了其生殖生长,导致花期不遇,雄穗散粉量减少,花粉活力降低,造成雌穗秃尖加长和籽粒败育增多,产量下降[34,35,36,37]。本研究发现,高温、干旱及其复合胁迫导致玉米散粉期缩短,雄穗散粉盛期提前,且对逆境越敏感的品种(HN138),散粉盛期提早越多,这也是HN138抗高温胁迫能力较ZD958差的原因之一。高温、干旱均可引起花药畸变、花药维管束发育异常导致花粉败育、活力降低[38],但本研究结果发现,两类胁迫引起花药畸变程度和对花粉粒形态影响程度不同,其中高温胁迫引起花药药室轻度形变,花粉粒表面出现网纹状凸起,而干旱胁迫则导致花药药室显著皱缩,且花药维管束显著变小,表皮细胞退化变形,表面网纹状凸起明显增多,皱缩加剧且形成脊,复合胁迫则使花药药室变形严重,花药壁断裂,绒毡层细胞退化,花药趋近解体,萌发孔严重内陷,使萌发和生理功能受到严重影响,表现出干旱胁迫基础上叠加高温胁迫。综合雄穗农艺性状和花粉结构发现,高温、干旱及复合胁迫对花粉活力及败育影响的主导因素不同,高温胁迫下花粉活力下降主要是由于高温影响花粉和花药结构而使活力降低,而干旱和复合胁迫则一方面导致花药、花粉结构畸变加剧,另一方面是雄穗小花数、分枝数显著减少,农艺和结构双重影响导致了干旱及复合胁迫下玉米雄穗花粉量锐减且活力低下。复合胁迫影响虽大于单一胁迫,但并不是二者胁迫相加,而是以干旱胁迫为主,高温胁迫由于对雄穗影响小而被弱化。品种间以分枝数和小花密度较多的品种类型(ZD958)更能适应干旱胁迫和复合胁迫。综合分析,复合胁迫对花粉、花药结构的不利影响大于干旱胁迫大于高温胁迫,但高温、干旱及复合胁迫下玉米雌穗花丝数量、活力及与结实性和产量的关系还有待于进一步深入研究。3.3 高温干旱胁迫对雄穗生理特性的影响
植株体内抗氧化系统和活性氧代谢的平衡变化是植株逆境胁迫的最直接生理表现[18],当植株体内的抗氧化酶无法清除自身体内过多活性氧时,则引起膜脂过氧化,产物积累[20]。本研究结果显示在高温、干旱及复合胁迫早期,雄穗SOD、POD活性和$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量、MDA含量快速增加,且活性氧和过氧化产物含量的高峰期早于抗氧化酶活性的高峰期2—4 d,说明逆境下先是活性氧和过氧化产物的生成打破了平衡,进而引起植株抗氧化酶活性增加来增强其清除能力,但随着胁迫时间延长,胁迫后期酶活性快速下降,酶促清除速度低于活性氧和过氧化产物产生速度,生理机能整体下降并趋于衰老,形成了持续逆境下的酶系统与产物生成的新动态平衡,因此表现出持续胁迫后期先是酶活性下降迅速,过氧化物则是缓慢下降,之后二者趋于平缓。但不同胁迫下抗氧化酶和代谢产物变化幅度不同,二者形成动态平衡的时间点存在差异,高温胁迫下$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量和MDA含量增减变幅小于干旱及复合胁迫,表现出高温胁迫一方面造成过氧化产物迅速积累,另一方面其酶清除能力(SOD、POD酶活性)也随之增大,能够较快形成高温胁迫下的新动态平衡,而干旱及复合胁迫下过氧化物产物含量累积量大于高温胁迫,但其抗氧化酶清除能力(SOD、POD酶活性)未能有与之相匹配的速度增加,由于酶系统不能及时清除过量的活性氧和过氧化产物,因此造成$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量和MDA含量增幅较大,因此干旱和复合胁迫下$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量和MDA含量的峰值出现时间早于高温胁迫,尤其复合胁迫下$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量峰值比干旱胁迫平均早1.5 d左右,使得复合胁迫的雄穗更早进入衰老状态。本研究结果还发现,雄穗MDA含量胁迫后第6天达到最大值,$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$含量胁迫后第4—6天达到最大值,酶清除系统(POD、SOD活性)则在第8天达到最大值,表现为过氧化物(MDA)积累高峰略晚于活性氧($\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$)积累高峰,因此与之对应的POD酶活性的峰值晚于SOD酶活性的峰值。这主要是在植物的生理反应中,通过自氧化反应、电子传递链、内质网细胞色素反应等均可生成$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$,逆境胁迫下则更易产生较多$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$,而$\mathop{{O}}_{2}^{{\mathop{}_{\ •}^{-}}}$进一步转化成羟自由基,羟自由基氧化性较强且可以穿透细胞膜,启动脂质过氧化反应,为膜脂过氧化反应及过氧化产物的产生造就了充分条件,因此逆境胁迫下超氧阴离子自由基的产生早于过氧化产物MDA的产生。热敏感型品种HN138抗氧化酶活性和过氧化产物含量峰值较钝感型品种ZD958提早1—2 d。综上,高温胁迫早期生理指标的迅速升高是作物对高温的应急反应,并未加速或导致雄穗的死亡,而干旱和复合胁迫各指标升高一方面是应急性生理响应,同时也是加速雄穗程序性死亡(衰老)的表现,其抗氧化酶活性在干旱和复合胁迫后迅速达到峰值随后迅速下降,就是其衰老死亡的直接生理表现。3.4 不同胁迫处理对玉米产量、产量构成要素的影响及其相关性
玉米穗行数、行粒数和粒重是产量构成要素,穗长、穗粗、秃尖以及籽粒容库协调性是影响产量构成的重要因素[39]。本研究发现,胁迫处理下玉米产量及其构成因素显著低于对照,以复合胁迫影响大于干旱大于高温;穗行数不受胁迫影响,这与本试验逆境处理开始时小穗分化已完成有关。高温胁迫下,行粒数减少是穗粒数大幅减少和产量大幅度下降的重要原因之一,且与花粉数量、活力极显著相关;干旱胁迫下产量下降与行粒数减少和粒重降低均显著相关,复合胁迫则在干旱胁迫基础上增大了行粒数减少的幅度,尤其是有效籽粒数大大减少,同时伴随粒重降低,这主要和花粉活力进一步降低,花粉量锐减有关。2个品种比较,胁迫下ZD958产量高于HN138,原因是高温胁迫下ZD958产量下降是行粒数减少所致,HN138产量降低则是行粒数、粒重双重因素减少所致;干旱和复合胁迫下行粒数和粒重下降,且降幅增大是二者产量下降的主要原因,但复合胁迫下玉米的行粒数降幅更大,出现严重败育和不结实现象。综合2个品种雄穗农艺性状和产量构成因素发现,高温、干旱胁迫下雄穗短轴、多分枝的品种(ZD958),可以通过单穗花粉数量多来弥补逆境造成的花粉量减少、花粉活力低的劣势,提高行粒数和结实性,因此比主轴长、分枝少的品种(HN138)更能抵御高温、干旱伤害。
4 结论
花期高温、干旱及复合胁迫导致玉米雄穗主轴变短,分枝数变少和分枝小花密度降低,花药和花粉结构劣变,花粉数量减少,活力下降;酶促清除和过氧化产物产生平衡打破,穗粒数减少,产量降低。高温干旱复合胁迫对雄穗生理指标及产量的胁迫程度大于干旱大于高温,以雄穗短轴、多分枝、散粉量多的品种抵御高温、干旱胁迫能力更强。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.2135/cropsci2012.09.0545URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1011.2010.00377URL [本文引用: 1]
DOI:10.3724/SP.J.1011.2010.00377URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1016/S1002-0160(06)60059-3URL [本文引用: 1]
DOI:10.3724/SP.J.1006.2012.00857URL [本文引用: 2]
DOI:10.3724/SP.J.1006.2012.00857URL [本文引用: 2]
DOI:10.3724/SP.J.1006.2012.01884URL [本文引用: 1]
DOI:10.3724/SP.J.1006.2012.01884URL [本文引用: 1]
DOI:10.1105/tpc.108.065318URL [本文引用: 1]
[本文引用: 1]
DOI:10.1105/tpc.110.074369URL [本文引用: 1]
DOI:10.1016/j.cell.2009.11.006URL [本文引用: 1]
[本文引用: 1]
DOI:10.1104/pp.107.115121URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/nph.2014.203.issue-1URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2018.00442URL [本文引用: 1]
DOI:10.3724/SP.J.1006.2018.00442URL [本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}