Effects of Density and Row Spacing on Seedling Traits of Rapeseed and Seed Yield
KUAI Jie,1, LI Zhen1,2, WANG Bo1, LIU Fang3, YE Jun4, ZHOU GuangSheng,11College of Plant Science and Technology, Huazhong Agricultural University/Key Laboratory of Crop Ecophysiology and Farming System in the Middle Reaches of the Yangtze River, Ministry of Agriculture and Rural Affairs, Wuhan 430070 2College of Agriculture, Jinhua Polytechnic, Jinhua 321007, Zhejiang 3National Agricultural Technology Extension Service Center, Beijing 100125 4Hubei Provincial Cultivated Land Quality and Fertilizer Station, Wuhan 430070
Abstract 【Objective】The relationship between seedling growth and yield formation of Brassica napus L. under different densities and row spacings was studied, which laid a theoretical foundation for further increasing yield and reducing yield gap, and it also helped to understand the mechanism of yield regulation in rapeseed with high-density planting. 【Method】The genotype HZ 62 was selected as the test material in 2016-2017, and HZ 62 and variety 1301, with different plant architecture, were grown in 2017-2018. The planting density was set as main plots at 15×104plants/hm2 (D1), 30×104 plants/hm2 (D2) and 45×104 plants/hm2 (D3), with the row spacing as subplots of 15 cm (R15), 25 cm (R25) and 35 cm (R35). The changes of dry matter accumulation and distribution in different organs, carbon and nitrogen metabolism in stem and leaf, root activity and yield at maturity were studied. 【Result】 The results showed that: the root diameter, root dry weight, shoot dry weight and plant height decreased at maturity, and the effective branch number decreased with increased plant density, and the decrease rate became smaller with narrowing row spacing at the same density. Under the density of D1, D2 and D3, the above indices had the best performance at R25, R15 and R15. Compared with D1R25, the yield of HZ62 and 1301 decreased by 57.14% and 55.73% respectively, but the population yield increased by 21.55% and 30.92% in 2017-2018 under D3R15. The correlation analysis showed that there was a highly significant positive correlation between leaf dry matter allocation rate and individual plant yield, also, between stem and root dry matter allocation rate and population yield. Further analysis of the growth indexes showed that with the increase of plant density, leaf SPAD value, root biomass per plant, root bleeding sap and root activity decreased significantly, while the root biomass increased significantly. Under the same density, the leaf SPAD value, leaf and stem C/N ratio, leaf LAI and root biomass of the population increased by adjusting row spacing and decreasing the difference between plant spacing and row spacing, which laid a foundation for the yield at maturity. In 2017-2018, compared with D1R25, the C/N ratio of the stem decreased by 22.95%, the root biomass, bleeding snap and root activity of individual plant decreased by 35.60%, 16.07% and 15.51% respectively, while leaf C/N ratio and root biomass in population increased by 16.11% and 83.44% respectively for HZ62; the stem C/N ratio decreased by 19.71%, the root biomass per plant, bleeding snap and root activity decreased by 30.87%, 22.63% and 22.85% respectively, while leaf C/N ratio and root biomass in population increased by 14.84% and 108.21% respectively for 1301 under D3R15. 【Conclusion】Under the condition of this experiment, compared with the traditional arrangement of plant density and row spacing, the increasing plant density and narrowed row spacing could promote the nitrogen metabolism of individual leaf and increase the leaf SPAD value, the photosynthetic leaf area and the biomass as well as activity of root at the seedling stage to achieve higher yield. Keywords:rapeseed;density;row spacing;seedling traits;yield
PDF (1064KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 蒯婕, 李真, 汪波, 刘芳, 叶俊, 周广生. 密度和行距配置对油菜苗期性状及产量形成的影响[J]. 中国农业科学, 2021, 54(11): 2319-2332 doi:10.3864/j.issn.0578-1752.2021.11.006 KUAI Jie, LI Zhen, WANG Bo, LIU Fang, YE Jun, ZHOU GuangSheng. Effects of Density and Row Spacing on Seedling Traits of Rapeseed and Seed Yield[J]. Scientia Acricultura Sinica, 2021, 54(11): 2319-2332 doi:10.3864/j.issn.0578-1752.2021.11.006
Table 1 表1 表1试验材料主要株型特征参数 Table 1The main characteristics of plant architecture for tested materials
品种 Variety
株高 Plant height (cm)
有效分枝部位 Branch height (cm)
主花序长度 Main inflorescence length (cm)
分枝角度 Branch angle (°)
分枝数 Branch number
华杂62 HZ62
184.5a
86.5a
53.4a
50.4a
8.7a
1301
163.8b
59.6b
45.2b
31.9b
7.8a
表中所用数据为两品种(系)在15×104株/hm2习惯种植时株型参数,取3年均值 The data were recorded when they are conventionally planted at 15×104 plants/hm2, and the average values of 3 years are taken
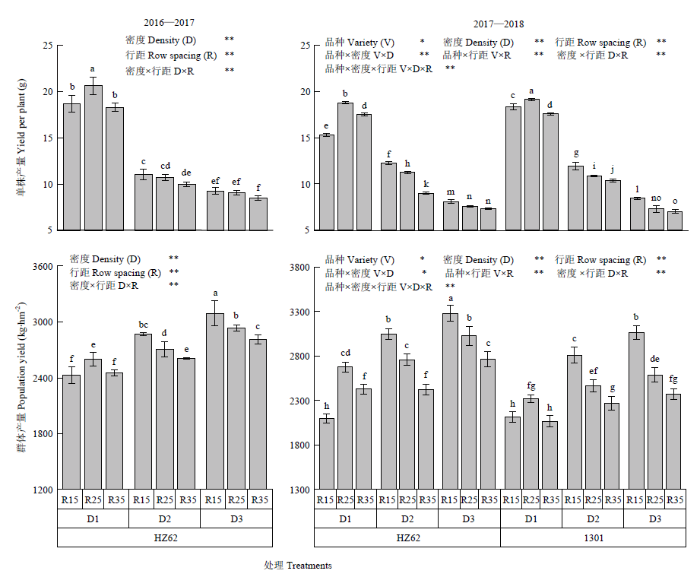
不同字母表示处理间差异达0.05水平;V:品种;D:种植密度;R:行距;FV×FD、FV×FR、FD×FR、FV×FD×FR分别表示因素间互作。*和**分别表示互作效应在0.05和0.01水平上显著。NS:差异不显著。下同 Fig. 1Seed yield at maturity of rapeseed under different densities and row spacing
Different letters indicated a difference of 0.05. V: Variety; D: Density; R: Row spacing; V×D, V×R, D×R, V×D×R represent interactions among treatments. * and ** means significant at the 0.05 and 0.01 probability level, respectively. NS: Difference is not significant according to ANOVA. The same as below
Table 2 表2 表2密度和行距配置对油菜成熟期农艺性状的影响 Table 2Effect of density and row spacing on agronomic characters of rapeseed at maturity stage
年份 Year
品种 Variety
密度 Density
行距 Row Spacing
根颈粗 Root crown diameter (mm)
根干重 Root biomass (g/plant)
株高 Plant height (cm)
有效分枝起点 Branch height (cm)
有效分枝数 Branch number
地上部干重 Shoot biomass (g/plant)
2016—2017
华杂62 HZ62
D1
R15
16.19b
14.37b
188.6ab
71.1f
7.5a
68.70c
R25
16.84a
16.11a
193.8a
77.0e
7.2a
79.00a
R35
15.51c
13.30c
187.4ab
79.3e
6.5b
74.69b
D2
R15
13.98d
8.73d
182.8bc
85.3d
6.1c
50.12d
R25
13.66d
7.62e
181.8bc
86.7d
6.0c
48.30e
R35
13.05e
5.26f
176.4c
95.0b
5.4d
46.80f
D3
R15
12.07f
4.92g
175.6c
86.7d
6.1c
40.20g
R25
10.69g
4.32h
174.4c
91.4c
5.4d
36.98h
R35
10.65g
4.52h
173.5c
101.1a
5.0e
36.90h
方差分析 Variance analyses
FD
**
**
**
**
**
**
FR
**
**
NS
**
**
**
FD×FR
**
**
NS
**
**
**
2017—2018
华杂62 HZ62
D1
R15
18.57c
13.05b
171.2b
68.7d
6.8c
66.22c
R25
21.74a
13.91a
179.6a
67.7d
8.3a
87.72a
R35
20.08b
13.26b
175.1ab
69.2d
7.4b
77.29b
D2
R15
15.59d
7.39c
171.7b
79.3c
5.9d
51.43d
R25
14.67e
6.65d
159.5c
78.5c
5.5e
43.58e
R35
14.07f
6.19d
150.9d
77.8c
4.1g
33.60fg
D3
R15
12.67g
4.90e
145.2de
84.1b
4.9f
34.96f
R25
12.09h
4.65e
143.4e
85.1b
4.0g
32.93fg
R35
12.48gh
4.86e
142.5e
90.9a
4.2g
30.60h
1301
D1
R15
16.83b
5.99a
161.6a
61.0e
8.0b
66.38b
R25
18.08a
6.04a
163.9a
59.3e
8.3a
75.20a
R35
16.83b
5.65b
157.8b
59.4e
7.2c
58.43c
D2
R15
14.82c
5.08c
157.6b
74.1d
6.8d
46.01d
R25
13.20d
4.21d
154.5bc
86.6b
5.5f
43.70e
R35
12.41e
3.27e
153.9bc
81.8c
6.0e
37.19f
D3
R15
11.56f
3.09e
151.8cd
88.4ab
4.6g
34.61g
R25
10.87g
2.73f
148.5de
90.6a
4.3i
32.30h
R35
9.96h
2.72f
146.8e
90.3a
4.4h
28.62i
方差分析 Variance analyses
品种Variety (V)
**
**
**
**
**
**
密度Density (D)
**
**
**
**
**
**
行距Row spacing (R)
**
**
NS
**
**
**
品种×密度V×D
**
**
**
**
**
**
品种×行距V×R
**
**
NS
**
**
**
密度×行距D×R
**
**
**
**
**
**
品种×密度×行距V×D×R
**
**
**
**
**
**
D1、D2和D3分别表示种植密度为15×104、30×104和45×104株/hm2;R15、R25和R35分别表示行距15、25和35 cm;同一列不同字母表示差异达0.05水平;*和**分别表示差异达0.05和0.01水平差异显著,NS:差异不显著;V:品种;D:种植密度;R:行距;FV×FD、FV×FR、FD×FR、FV×FD×FR分别表示因素间互作。下同 D1, D2 and D3 represent the planting densities of 15×104 plants/hm2, 30×104 plants/hm2 and 45×104 plants/hm2, respectively. R15, R25 and R35 represent row spacing of 15 cm, 25 cm and 35 cm respectively. Different letters in the same column indicated a difference of 0.05. * and ** mean the difference is at the level of 0.05 and 0.01 respectively. NS: Difference is not significant. V: Variety; D: Density; R: Row spacing; V×D, V×R, D×R, V×D×R represent interactions among treatments. The same as below
Table 4 表4 表4苗期根系生物量、伤流量和根系活力 Table 4Effect of density and row spacing on root biomass, root blooding sap and root activity of rapeseed at seedling stage
ZHAOY G, ZHAOS Y, CHENGY, LUOL X, FUG P, GUOR X, HUANGZ Y, MAH Q, LIUQ Y, ZHANGX K, CAIJ S. Rapeseed yield comparison between China and Canada in national trials of main producing areas Chinese Journal of Oil Crop Sciences, 2017,39(3):420-426. (in Chinese) [本文引用: 1]
WANGR, CHENGT, HUL Y. Effect of wide-narrow row arrangement and plant density on yield and radiation use efficiency of mechanized direct-seeded canola in Central China Field Crops Research, 2015,172:42-52. DOI:10.1016/j.fcr.2014.12.005URL [本文引用: 2]
KUAIJ, SUNY, ZUOQ, HUANGH, LIAOQ, WUC, LUJ, WUJ, ZHOUG. The yield of mechanically harvested rapeseed (Brassica napus L.) can be increased by optimum plant density and row spacing Scientific Reports, 2016,5(1):18835 DOI:10.1038/srep18835URL [本文引用: 1]
KUAIJ, SUNY, ZHOUM, ZHANGP, ZUOQ, WUJ, ZHOUG. The effect of nitrogen application and planting density on the radiation use efficiency and the stem lignin metabolism in rapeseed (Brassica napus L.) Field Crops Research, 2016,199:89-98. DOI:10.1016/j.fcr.2016.09.025URL [本文引用: 1]
LIX Y, ZHOUM, WANGT, ZHANGL, ZHOUG S, KUAIJ. Effects of planting density on the mechanical harvesting characteristics of semi-winter rapeseed Acta Agronomica Sinica, 2018,44(2):278-287. (in Chinese) DOI:10.3724/SP.J.1006.2018.00278URL [本文引用: 2]
STAMPP, KIELC. Root morphology of maize and its relationship to root lodging Journal of Agronomy & Crop Science, 2008,168(2):113-118. [本文引用: 1]
SNIDERJ L, RAPERR L, SCHWABE B. The effect of row spacing and seeding rate on biomass production and plant stand characteristics of non-irrigated photoperiod-sensitive sorghum (Sorghum bicolor (L.) Moench) Industrial Crops and Products, 2012,37(1):527-535. DOI:10.1016/j.indcrop.2011.07.032URL [本文引用: 1]
XUEJ, GOUL, ZHAOY, YAOM, YAOH, TIANJ, ZHANGW. Effects of light intensity within the canopy on maize lodging Field Crops Research, 2016,188:133-141. DOI:10.1016/j.fcr.2016.01.003URL [本文引用: 1]
HEITHOLTJ J, SASSENRATH-COLEG F. Inter-plant competition: growth responses to plant density and row spacing//STEWART J M, OOSTERHUIS D M, HEITHOLT J J, MAUNEY J R. eds. Physiology of Cotton. New York: Springer Dordrecht Heidelberg London, 2010. [本文引用: 1]
HUQ, JIANGW Q, QIUS, XINGZ Q, HUY J, GUOB W, LIUG D, GAOH, ZHANGH C, WEIH Y. Effect of wide-narrow row arrangement in mechanical pot-seedling transplanting and plant density on yield formation and grain quality of japonica rice Journal of Integrative Agriculture, 2020,19(5):1197-1214. DOI:10.1016/S2095-3119(19)62800-5URL [本文引用: 2]
FISCHERR A, MORENORAMOS O H, ORTIZMONASTERIO I, SAYREK D. Yield response to plant density, row spacing and raised beds in low latitude spring wheat with ample soil resources: An update Field Crops Research, 2019,232:95-105. DOI:10.1016/j.fcr.2018.12.011URL [本文引用: 1]
WIDDICOMBEW D, THELENK D. Row width and plant density effects on corn grain production in the northern Corn Belt Agronomy Journal, 2002,94:1020-1023. DOI:10.2134/agronj2002.1020URL [本文引用: 2]
GOBEZEY L, CERONIOG M, VANRENSBURG L D. Effect of row spacing and plant density on yield and yield component of maize (Zea mays L.) under irrigation Journal of Agricultural Science & Technology B, 2012,2:263-271. [本文引用: 1]
TURGUTI, DUMANA, BILGILIU, ACIKGOZE. Alternate row spacing and plant density effects on forage and dry matter yield of corn hybrids (Zea mays L.) Journal of Agronomy and Crop Science, 2005,191(2):146-151. DOI:10.1111/jac.2005.191.issue-2URL [本文引用: 2]
SEITERS, ALTEMOSEC E, DAVISM H. Forage soybean yield and quality responses to plant density and row distance Agronomy Journal, 2004,96(4):966-970. DOI:10.2134/agronj2004.0966URL [本文引用: 2]
BOQUETD J. Cotton in ultra-narrow row spacing: Plant density and nitrogen fertilizer rates Agronomy Journal, 2005,97(1):279-287. DOI:10.2134/agronj2005.0279URL [本文引用: 3]
GARSIDEA L, BELLM J, ROBOTHAMB G. Row spacing and planting density effects on the growth and yield of sugarcane: 2. Strategies for the adoption of controlled traffic Crop & Pasture Science, 2009,60(6):544-554. [本文引用: 1]
YANY H, YANGW J, ZHANGX Q, CHENX L, CHENZ Q. Improve soybean seeding growth by uniconazole under shading by corn in relay strip intercropping system Chinese Journal of Oil Crop Sciences, 2011,33(3):259-264. (in Chinese) [本文引用: 2]
ZHENGW, YEC, XIAOG B, CHENM, LIY Z, HUANGT B, XIAOX J, LIUX S, ZHUC. Effects of symbiotic period on seedling traits and yield components of interplanting rapeseed in rice Scientia Agricultura Sinica, 2015,48(21):4254-4263. (in Chinese) [本文引用: 2]
COSTAC, DWYERL M, HAMILTONR I, HAMELC, NANTAISL, SMITHD L. A sampling method for measurement of large root systems with scanner-based image analysis Agronomy Journal, 2000,92(4):621-627. DOI:10.2134/agronj2000.924621xURL [本文引用: 2]
BRARG, THIESW. Contribution of leaves, stem, siliques and seeds to dry matter accumulation in ripening seeds of rapeseed, Brassica napus L Zeitschrift Fü Pflanzenphysiologie, 1977,82(1):1-13. [本文引用: 2]
SEEBAUERJ R, SINGLETARYG W, KRUMPELMANP M, RUFFOM L, BELOWF E. Relationship of source and sink in determining kernel composition of maize Journal of Experimental Botany, 2010,61(2):511-519. DOI:10.1093/jxb/erp324URL [本文引用: 2]
GIRONDEA, ETIENNEP, TROUVERIEJ, BOUCHEREAUA, LECAHEREC F, LEPORTL, ORSELM, NIOGRETM F, NESIN, CAROLED, SOULAYF, MASCLAUX-DAUBRESSEC, AVICEJ C. The contrasting N management of two oilseed rape genotypes reveals the mechanisms of proteolysis associated with leaf N remobilization and the respective contributions of leaves and stems to N storage and remobilization during seed filling BMC Plant Biology, 2015,15(1):1-21. DOI:10.1186/s12870-014-0410-4URL [本文引用: 2]
READS M, NORTHCOTED H. Minimization of variation in the response to different protein of the Coomassic Blue G dye - binding: assay to protein Analytical Biochemistry, 1981,116(1):53-64. DOI:10.1016/0003-2697(81)90321-3URL [本文引用: 1]
XIEY F, WANGY, ZHANGZ M, SHANGX L, YANGW X, FANGS Z. Effects of uniconazole on growth and physiological characteristics of Cyclocarya paliurus plantlets Journal of Plant Resources and Environment, 2010,19(4):50-55. (in Chinese) [本文引用: 1]
MARTINSN, GON?ALVESS, ROMANOA. Aluminum inhibits root growth and induces hydrogen peroxide accumulation in Plantago algarbiensis and P. almogravensis seedlings Protoplasma, 2013. 250(6):1295-1302. DOI:10.1007/s00709-013-0511-1URL [本文引用: 2]
LIX, ZUOQ, CHANGH, BAIG, ZHOUG. Higher density planting benefits mechanical harvesting of rapeseed in the Yangtze River Basin of China Field Crops Research, 2018,218:97-105. DOI:10.1016/j.fcr.2018.01.013URL [本文引用: 1]
PARKS E, BENJAMINL R, WATKINSONA R. The theory and application of plant competition models: an agronomic perspective Annals of Botany, 2003,92(6):741-748. DOI:10.1093/aob/mcg204URL [本文引用: 1]
ZHANGZ J, CHUG, LIUL J, WANGZ Q, WANGX M, ZHANGH, YANGJ C, ZHANGJ H. Mid-season nitrogen application strategies for rice varieties differing in panicle size. Field Crops Research, 2013,150:9-18. DOI:10.1016/j.fcr.2013.06.002URL [本文引用: 1]
WEIH Y, ZHANGH C, HANGJ, DAIQ G, HUOZ Y, XUK, ZHANGS F, MAQ, ZHANGQ, ZHANGJ. Characteristics of N accumulation and translocation in rice genotypes with different N use efficiencies Acta Agronomica Sinica, 2008,34(1):119-125. (in Chinese) DOI:10.3724/SP.J.1006.2008.00119URL [本文引用: 1]
ZHENGC Y, YUZ W, WANGX Z, WUT H. Effects of irrigation amount and stage on nitrogen accumulation, distribution, translocation and soil-N content in high-yield wheat Plant Nutrition and Fertilizer Science, 2009,15(6):1324-1332. (in Chinese) [本文引用: 1]
ZUOQ S, LIUH, KUAIJ, FENGQ N, FENGY Y, ZHANGH X, LIUJ Y, YANGG, ZHOUG S, LENGS H. Effects of nitrogen and planting density on accumulation, translocation and utilization efficiency of carbon and nitrogen transplanting rapeseed with blanket seedling Scientia Agricultura Sinica, 2016,49(18):3522-3531. (in Chinese) [本文引用: 1]
ZUOQ S, YANGH Y, LENGS H, CAOS, ZENGJ X, WUJ S, ZHOUG S. Effects of nitrogen fertilizer on nitrogen accumulation, translocation and nitrogen use efficiency in rapeseed (Brassica napus L.) Acta Agronomica Sinica, 2014,40(3):511-518. (in Chinese) DOI:10.3724/SP.J.1006.2014.00511URL [本文引用: 1]
VAMERALIT, SACCOMANIM, BONAS, MOSCAG, GUARISEM, GANISA. A comparison of root characteristics in relation to nutrient and water stress in two maize hybrids Plant Soil, 2003,255(1):157-167. DOI:10.1023/A:1026123129575URL [本文引用: 1]
ZHOUY, YANGX W, ZHOUS M, WANGY J, YANGR, XUF D, MEIJ J, SHENG Y, LIQ J, HED X. Activities of key enzymes in root NADP-Dehydrogenase system and their relationships with root vigor and grain yield formation in wheat Scientia Agricultura Sinica, 2018,51(11):2060-2071. [本文引用: 1]
YANGY M, SUNY M, JIAL L, JIAS L, MENGC X. Effects of phosphorus fertilization depth on yield and root distribution of summer maize Scientia Agricultura Sinica, 2018,51(8):142-150. (in Chinese) [本文引用: 1]
GUOX H, WANGX C, MENGT, ZHANGH J, AOX, WANGH Y, XIEF D. Comparison of root morphological and activity of representative soybean cultivars(MG Ⅲ) developed in the USA and China Scientia Agricultura Sinica, 2015,48(19):3821-3833. (in Chinese) [本文引用: 1]
YANGG, ZHANGH J, SONGS H, WANGW B, AOX, XIEF D. Comparison on some root related traits of super-high-yielding soybean Soybean Science, 2013,32(2):176-181. (in Chinese) [本文引用: 1]
ZHANGH B, RENW J, YANGW J. Relationship between root characteristics and yield formation in relay-planting soybean under the nitrogen application Soil and Fertilizer Sciences China, 2007,2:46-49. (in Chinese) [本文引用: 1]
 ,1, 李真1,2, 汪波1, 刘芳3, 叶俊4, 周广生
,1, 李真1,2, 汪波1, 刘芳3, 叶俊4, 周广生

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT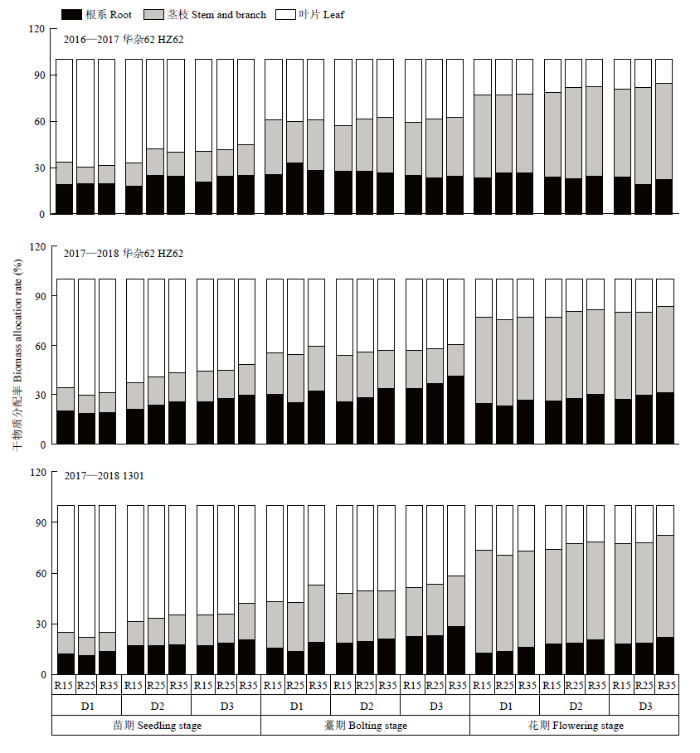 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT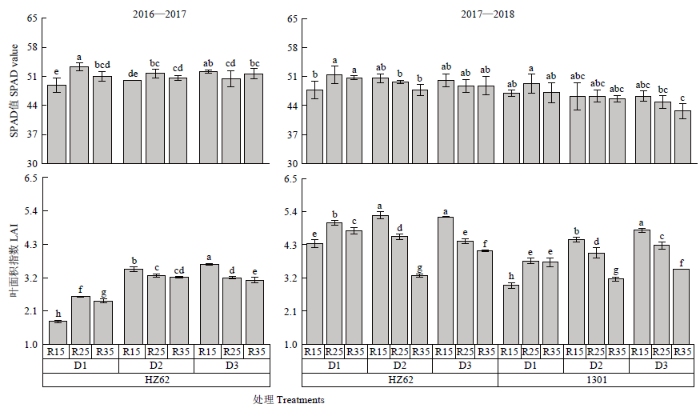 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT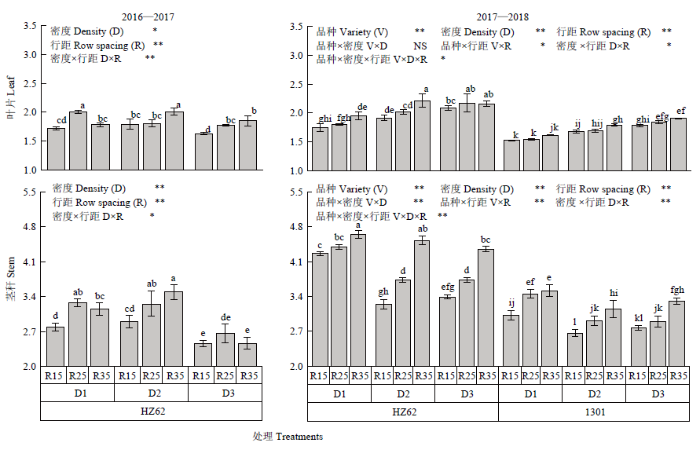 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}