Implantation and Persistence of Inoculated Active Dry Yeast in Industrial Wine Fermentations
SUN Yue,1, YANG HuiMin2, HE RongRong1, ZHANG JunXiang,11School of Food and Wine, Ningxia University, Yinchuan 750021 2College of Enology, Northwest A&F University, Yangling 712100, Shaanxi
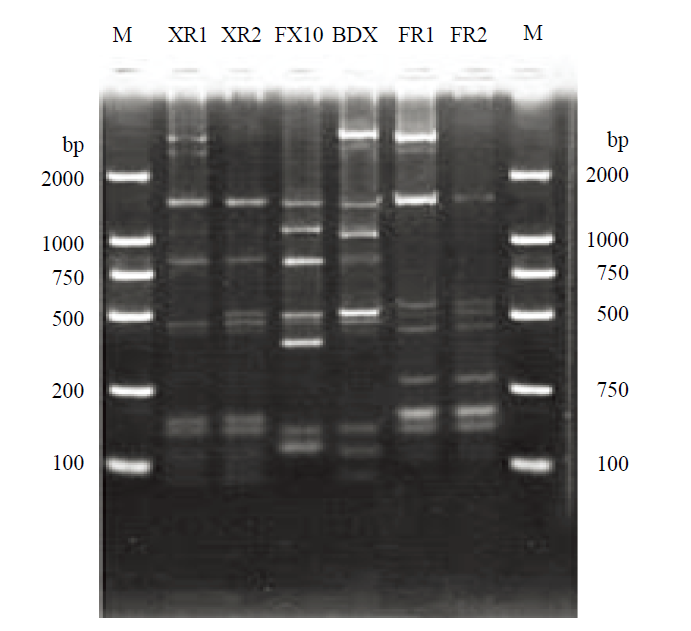
Abstract 【Objective】The purpose of this study was explore the implantation and persistence of commercial active dry yeast (ADY) during industrial wine fermentations, and their competitive relationship between Chinese indigenous Saccharomyces cerevisiae during fermentation process, so as to provide the theoretical basis for the breeding of indigenous S. cerevisiae strains and provide the reference for the use of ADY in wine production. 【Method】Industrial wine fermentations were carried out at wineries in Eastern Foot of Helan Mountain in Ningxia. Four vats of Cabernet Sauvignon gape must were inoculated with BDX, XR, FR and FX10, respectively. Samples were collected and analyzed at 1 d, 3 d and 5 d after the inoculation. Interdelta and SSR analysis were used to investigate the genotypes of different S. cerevisiae strains. Therefore, the number and proportion of S. cerevisiae strains in different fermentation stages were analyzed, and the colonization ability of commercial ADY was tracked. Genetic diversity parameters were calculated by PopGen32 software. The genetic correlation between commercial yeast and Ningxia indigenous yeast was revealed by NTsys2.10e software. 【Result】Interdelta fingerprint showed 6 kinds of fingerprints, namely 6 genotypes. And XR and FR showed more than one genotype; BDX and FX10 showed one genotype, respectively. SSR analysis showed that there was one genotype in each ADY for the 9 locus. 225 S. cerevisiae isolates were isolated from the 4 inoculated fermentations. Interdelta fingerprint showed 42 genotypes, of which 36 genotypes were indigenous strains. The degree of variability (16%, 42/225) was intermediate. SSR analysis showed 20 genotypes, of which 16 genotypes were indigenous strains. The analyzed 9 microsatellite prime pairs generated a total of 75 polymorphic bands, 8.3333 alleles for per locus. The heterozygosity observed was 0.2000-0.5000. The polymorphism information contents (PIC) of all strains at 9 loci were 0.6339-0.8620, suggesting that the 9 SSR loci were hypervariable. The indigenous genotypes were the most abundant in the fermentation inoculated with BDX (11 Interdelta types and 8 SSR types), followed by FR (11 Interdelta types and 6 SSR types). ADY did not dominate all three stages. Moreover, the genotypes of the dominant strains were also different for different stages in the same fermentation. Interdelta and SSR analysis showed FR was not dominant in the corresponding fermentation. Although BDX existed in the whole fermentation process, it was only dominant at d 3 after the inoculation. In the fermentation inoculated with XR and FX10, Interdelta analysis showed that they were not the dominant strains, while SSR analysis showed that they were the dominant strains in the corresponding fermentations, respectively. Indigenous strains of genotype β (SSR genotype BDX-7), genotype γ (SSR genotype BDX-6), genotype A (SSR genotype XR), genotype a (SSR genotype FX10), genotype b (SSR genotype FX10), genotype bb (SSR genotype FR-2) and genotype ee (SSR genotype FR-4) showed strong competitiveness in the corresponding fermentations. Cluster analysis showed that the genetic diversity among the S. cerevisiae strains isolated from the same fermentation was large. 【Conclusion】The genotypes of indigenous S. cerevisiae strains in the industrial wine fermentations were rich. The inoculated fermentations were completed by both indigenous strains and commercial ADY, and they competed with each other in the same fermentations and showed dynamic succession of different strains. Keywords:ADY;inoculated fermentation;strain typing;wine;SSR
PDF (615KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 孙悦, 杨慧敏, 何荣荣, 张军翔. 商业酵母在葡萄酒工业化生产中的定殖情况分析[J]. 中国农业科学, 2021, 54(9): 2006-2016 doi:10.3864/j.issn.0578-1752.2021.09.016 SUN Yue, YANG HuiMin, HE RongRong, ZHANG JunXiang. Implantation and Persistence of Inoculated Active Dry Yeast in Industrial Wine Fermentations[J]. Scientia Acricultura Sinica, 2021, 54(9): 2006-2016 doi:10.3864/j.issn.0578-1752.2021.09.016
A:接种BDX的发酵;B:接种XR的发酵;C:接种FR的发酵;D:接种FX10的发酵 Fig. 3Dynamics changes of different S. cerevisiae strains during inoculated fermentation of BDX, XR, FR and FX10
A: Inoculated with BDX; B: Inoculated with XR; C: Inoculated with FR; D: Inoculated with FX10
GIL-DíAZM, VALEROE, CABALLOSJ M, GARCIAM, ARROYOT. The impact of active dry yeasts in commercial wineries from the denomination of origen “Vinos de Madrid”, Spain 3 Biotech, 9(11):382. doi: 10.1007/s13205-019-1913-3. [本文引用: 5]
BARRAJóNN, ARéVALO-VILLENAM, RODRíGUEZ-ARAGóNL J, BRIONESA. Ecological study of wine yeast in inoculated vats from La Mancha region Food Control, 2009,20(9):778-783. [本文引用: 4]
CORDEROBUESOG, RODRíGUEZM E, GARRIDOC, CANTORALJ M. Rapid and not culture-dependent assay based on multiplex PCR-SSR analysis for monitoring inoculated yeast strains in industrial wine fermentations Archives of Microbiology, 2017,199(1):135-143. [本文引用: 1]
DECELIS M, RUIZJ, MARTíN-SANTAMARíAM, MARíA,ALONSN A, MARQUINAD, NAVASCUéSE, GOMEZ-FLECHOSOM A, BELDAI, SANTOSA. Diversity of Saccharomyces cerevisiae yeasts associated to spontaneous and inoculated fermenting grapes from Spanish vineyards Letters in Applied Microbiology, 2019,68(6). doi: 10.1111/lam.13155. [本文引用: 1]
VIGENTINII, FABRIZIOV, FACCINCANIM, PICOZZIC, COMASIOA, FOSCHINOR. Dynamics of Saccharomyces cerevisiae populations in controlled and spontaneous fermentations for Franciacorta D.O.C.G. base wine production Annals of Microbiology, 2014,64:639-651. [本文引用: 2]
LANGEJ N, FAASSEE, TANTIKACHORNKIATM, GUSTAFSSONF S, HALVORSENL C, KLUFTINGERA, LEDDERHOFD, DURALLD M. Implantation and persistence of yeast inoculum in pinot noir fermentations at three Canadian wineries International Journal of Food Microbiology, 2014,180:56-61. [本文引用: 2]
MATURANOY P, LERENAM C, MESTREM V, CASASSAL F, TOROM E, VAZQUEZF, MERCADOL, COMBINAM. Inoculation strategies to improve persistence and implantation of commercial S. cerevisiae strains in red wines produced with prefermentative cold soak LWT-Food Science and Technology, 2018,97:648-655. [本文引用: 4]
SUNY, YED Q, CHUY, ZHANGY F, LIUY L. Effects of different inoculation methods & temperatures on fermentation of active dry yeast Brewing Technology, 2019(3):24-28, 37. (in Chinese) [本文引用: 3]
HEJ, CAOP X, HUANGY Z, LIUY L. Dynamics of wine related yeasts during industrial icewine fermentations of Italian Riesling Brewed in China, 2014,33(5):30-33. (in Chinese) [本文引用: 1]
SONGY Y, PEIY F, WANGG P, LIUY L. Monitoring of yeast changes during inoculated fermentation process of Pinot Noir grape Chinese Journal of Food Science, 2010,10(2):125-130. (in Chinese) [本文引用: 2]
LIUS T, LIQ, LüW, QINP. Present situation and development trend of Chinese wine industry: A case study of producing area at Eastern Foot of Helan Mountain in Ningxia Modern Agricultural Science and Technology, 2019(9):241-243. (in Chinese) [本文引用: 1]
SUNY. Effects of different nitrogen levels on the characteristics and metabolites of Saccharomyces cerevisiae during mixed fermentation. [D]. Yangling: Northwest A & F University, 2016. (in Chinese) [本文引用: 1]
SUNY, QINY, PEIY F, WANGG P, JOSEPHC M, BISSONL, LINDAF, LIUY L. Evaluation of Chinese Saccharomyces cerevisiae wine strains from different geographical origins American Journal of Enology and Viticulture, 2017,68:73-80. [本文引用: 2]
SUNY, ZHANGF F, CHUS X, LIJ X, SHAOS, ZHANGJ X. Effects of Saccharomyces cerevisiae strains with different killer activity on the yeast diversity in inoculated fermentation of Cabernet Sauvignon Food Science, 2020,41(2):166-172. (in Chinese) [本文引用: 1]
LEGRASJ L, RUHO, MERDINOGLUD, KARSTF. Selection of hypervariable microsatellite loci for the characterization of Saccharomyces cerevisiae strains International Journal of Food Microbiology, 2005,102(1):73-83. [本文引用: 9]
JUBANYS, TOMASCOI, DELEO′N I P, MEDINAK, CARRAUF, ARRAMBIDEN, NAYAH, GAGGEROC. Toward a global database for the molecular typing of Saccharomyces cerevisiae strains FEMS Yeast Research, 2008,8:472-484. [本文引用: 2]
HALLB, DURALLD M, STANLEYG. Population dynamics of Saccharomyces cerevisiae during spontaneous fermentation at a British Columbia Winery American Journal of Enology and Viticulture, 2011,62(1):66-72. [本文引用: 5]
STEFANINII, ALBANESED, SORDOM, LEGRASJ L, DEFILIPPO C, CAVALIERID, DONATIC. SaccharomycesIDentifier, SID: Strain-level analysis of Saccharomyces cerevisiae populations by using microsatellite meta-patterns Scientific Reports, 2017,7:15343. [本文引用: 7]
JIANGW H, YANB, TAOY S. Isolation and screening of Sacchamyces cerorevisiae from grape harvested in Changli Food Industry Technology, 2014,35(12):202-206, 209. (in Chinese) [本文引用: 1]
RICHARDSK D, GODDARDMATTHEW R, GARDNERRICHARD C. A database of microsatellite genotypes for Saccharomyces cerevisiae Antonie van Leeuwenhoek, 2009,96(3):355-359. [本文引用: 1]
TORIJAM J, ROZEèSN, POBLETM, GUILLAMóNJ, MASA. Yeast population dynamics in spontaneous fermentations: Comparison between two different wine-producing areas over a period of three years Antonie van Leeuwenhoek, 2001,79:345-352. [本文引用: 1]
BOTSTEIND, WHITER L, SKOLNICKM, DAVISR W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms American Journal of Human Genetics, 1980,32(3):314-331 [本文引用: 1]
YANGK, MAOR Z, ZHAOY, HEC, WANGH L, CAOJ H, SUW, HEX H. Biodiversity of wine-related yeasts isolated from Shangri-La wine-producing region of Yunnan Bulletin of Microbiology, 2018,45(12):2708-2721. (in Chinese) [本文引用: 1]
JIAJ, WANGG Q, ZHUL X, HANP J, GUOD Q, CHENM. Structure characteristics of Saccharomyces cerevisiae population associated with grape and wine in Xinjiang Journal of Dalian Polytechnic University, 2018,37(2):94-99. (in Chinese) [本文引用: 1]
FENGM, WANGC X, LIUY L. Genetic diversity of Saccharomyces cerevisiae strains revealed by Microsatellite sequence polymorphism Scientia Agricultura Sinica, 2012,45(12):2537-2543. (in Chinese) [本文引用: 1]
ZHANGL Y, HUANGY Z, LIUY L. Genetic diversity analysis of Saccharomyces cerevisiae strains by Microsatellite marker technique Food Science, 2014,35(1):130-133. (in Chinese) [本文引用: 1]
YUANW. Comparative study of the vinification characteristic of selected wine yeast [D]. Yangling: Northwest A&F University, 2010: 4-7. (in Chinese) [本文引用: 1]
LIS S, LIS, LIW L, WUZ M, SUN. Application of indigenous yeast strain in the wine making Food Science and Technology, 2012,37(8):87-91. (in Chinese) [本文引用: 1]
ORTIZM J, BARRAJóNN, BAFFIM A, MARIAAV, BRIONESA. Spontaneous must fermentation: Identification and biotechnological properties of wine yeasts LWT-Food Science and Technology, 2013,50(2):371-377. [本文引用: 1]
?URANSKáH, VRáNOVáD, OMELKOVáJ. Isolation, identification and characterization of regional indigenous Saccharomyces cerevisiae strains Brazilian Journal of Microbiology, 2016,47(1):181-190. [本文引用: 1]
GAROFALOC, BERBEGALC, GRIECOF, MARIAT, GIUSEPPES, VITTORIOC. Selection of indigenous yeast strains for the production of sparkling wines from native Apulian grape varieties International Journal of Food Microbiology, 2018,285:7-17. [本文引用: 1]
 ,1, 杨慧敏2, 何荣荣1, 张军翔
,1, 杨慧敏2, 何荣荣1, 张军翔

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT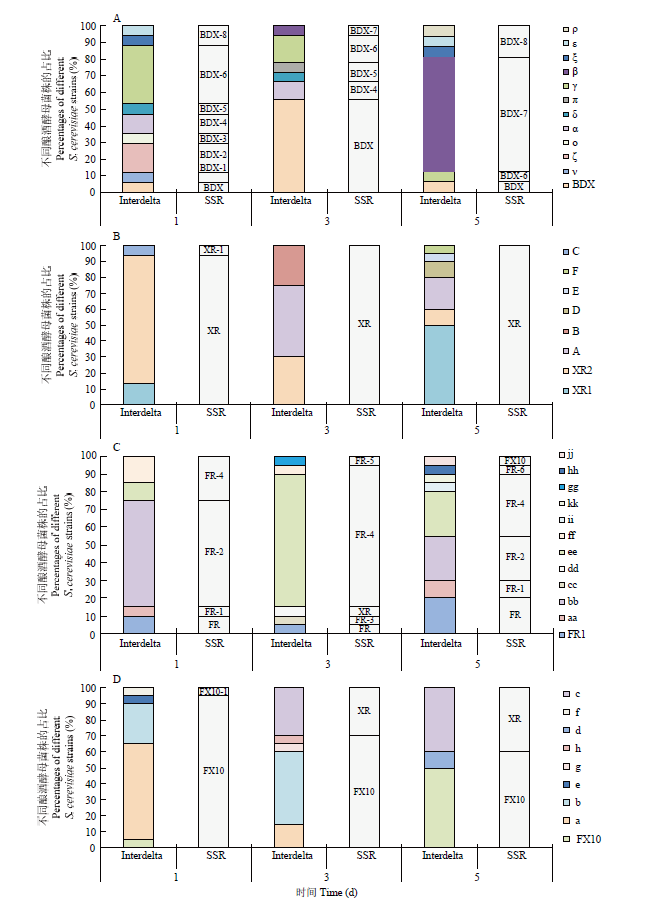 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT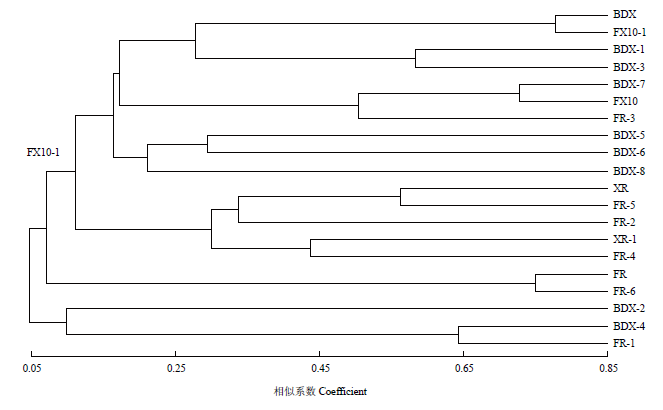 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}