 ,1, 仇贵生1, 佟兆国2, 张怀江1, 闫文涛1, 岳强1, 孙丽娜,1
,1, 仇贵生1, 佟兆国2, 张怀江1, 闫文涛1, 岳强1, 孙丽娜,1Ligands Binding Characteristics of Chemosensory Protein CsasCSP16 of Carposina sasakii
LIU XiaoHe,1, QIU GuiSheng1, TONG ZhaoGuo2, ZHANG HuaiJiang1, YAN WenTao1, YUE Qiang1, SUN LiNa,1通讯作者:
责任编辑: 岳梅
收稿日期:2020-05-11接受日期:2020-06-22网络出版日期:2021-03-01
| 基金资助: |
Received:2020-05-11Accepted:2020-06-22Online:2021-03-01
作者简介 About authors
刘孝贺,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (4349KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
刘孝贺, 仇贵生, 佟兆国, 张怀江, 闫文涛, 岳强, 孙丽娜. 桃小食心虫化学感受蛋白CSP16配体结合特性[J]. 中国农业科学, 2021, 54(5): 945-958 doi:10.3864/j.issn.0578-1752.2021.05.007
LIU XiaoHe, QIU GuiSheng, TONG ZhaoGuo, ZHANG HuaiJiang, YAN WenTao, YUE Qiang, SUN LiNa.
开放科学(资源服务)标识码(OSID):

0 引言
【研究意义】桃小食心虫(Carposina sasakii)属鳞翅目(Lepidoptera)蛀果蛾科(Carposidae),雌虫将卵产在苹果、桃、梨、山楂等果实的萼洼处或者果面,幼虫孵化后蛀入果实内部纵横串食,造成“豆沙馅”似的虫果,给果农造成严重的经济损失[1]。目前对桃小食心虫的防控主要依靠化学农药,但是大量化学农药的使用导致天敌被误杀并引起环境问题。随着对昆虫嗅觉机制研究的逐渐深入,引诱剂和趋避剂的开发为害虫防控提供了新的思路和策略。研究桃小食心虫化学感受蛋白(chemosensory protein,CSP)与寄主挥发物的识别和结合分子机制,对明确该虫产卵定位和取食选择以及建立基于昆虫行为学的生态防控具有重要理论意义。【前人研究进展】嗅觉在昆虫生理活动中发挥关键作用,例如定位产卵场所、寻找寄主植物和避免天敌危害等[2,3]。气味受体(odorant receptor,OR)或离子型受体(ionotropic receptor,IR)具有分辨和识别不同气味信号的功能。然而,疏水性的气味分子到达神经元树突激活气味受体或离子型受体之前,通常需要与气味结合蛋白(odorant binding protein,OBP)或化学感受蛋白相结合以复合物的形式穿过亲水性的淋巴液,到达嗅觉神经元附近刺激嗅觉受体,化学信号转化为电信号[4]。尽管OBP或CSP在嗅觉形成过程中的具体功能尚未完全探明,但是,在基因组和转录组中发现大量的OBP和CSP基因可能发挥关键作用[5,6,7,8]。CSP通常由100—120个氨基酸残基组成,结构紧凑,主要由6个α-螺旋形成一个疏水性的空腔。不同种类昆虫的CSP都含有4个保守的半胱氨酸残基,会形成两个二硫键用以保持CSP三维结构的稳定[9,10]。与OBP相比,CSP除了具有嗅觉功能外,还具有其他生理功能。CSP被鉴定出分布在不同的组织中。CSP首先在黑腹果蝇(Drosophila melanogaster)中被发现,命名为OS-D和A10,但当时关于CSP的功能还不清楚[11]。目前,有关CSP功能的信息在许多昆虫中得到验证。(1)CSP参与昆虫的再生和发育。在螳螂中,CSP10参与螳螂若虫断足的再生;蜜蜂的CSP5与受精卵和表皮的发育有关[12,13]。(2)使用亚致死浓度的菊酯类农药处理小菜蛾(Plutella xylostella)的研究中发现,有3个CSP(CSP3、CSP4和CSP8)基因表达量上调,推测CSP参与昆虫抗药性的形成[14,15]。(3)蝗虫由散居型变成聚居型时,体内的CSP基因表达量提高,CSP可能与蝗虫的聚集状态改变有关[16]。(4)一些CSP在昆虫喙和眼中作为营养吸收和视觉色素的载体[17]。(5)有研究表明,CSP在性腺或生殖器官中结合化学信息素信号,并协助它们释放到环境中[18,19,20]。最后,CSP结合气味分子和信息素分子,参与昆虫嗅觉的形成过程,CSP在蚊类的淋巴液中富集并且对化学信息素具有很高的亲和性;在蜜蜂触角中高表达的CSP3可以与信息素结合[21]。CSP如此广泛的表达谱和结合谱说明其在昆虫的生命活动中发挥重要的作用。因此,有关CSP的具体生理功能需要深入研究。【本研究切入点】笔者所在课题组前期从桃小食心虫的触角转录组中鉴定29个OBP基因,并发现2个GOBP和3个PBP基因,研究结果表明酯类、醛类和萜类等气味分子是潜在的吸引物质[22,23]。前期研究发现,CsasCSP16在雌、雄成虫的触角和翅中均有表达[24]。本研究采用原核表达和荧光竞争结合试验,研究CsasCSP16与35种小分子化合物的结合特性。使用分子对接和动力学模拟技术,预测CsasCSP16与小分子结合的关键位点。【拟解决的关键问题】研究桃小食心虫CsasCSP16与相关化合物的结合特性,并通过三维建模和分子动力学模拟方法研究CsasCSP16与配体结合的分子机制,为桃小食心虫的绿色防控技术提供新的思路和理论依据。1 材料与方法
试验于2018年7月至2020年3月在中国农业科学院果树研究所完成。1.1 供试昆虫
供试种群于2018年7月采自中国农业科学院果树研究所(40.61°N,120.73°E)试验园。采集被桃小食心虫幼虫蛀入带有“泪滴”危害状的金冠苹果,带回实验室,于(25±1)℃、相对湿度70%±5%、光周期L﹕D=15 h﹕9 h条件下饲养[25]。1.2 总RNA提取及cDNA第一链合成
分别取桃小食心虫雌、雄成虫全虫以及触角(100对)、头部(去触角,20头)、胸部(10个)、腹部(10个)、足(20头×6对)、翅(20对)等组织,按照TRIzol(TaKaRa)试剂盒说明书提取总RNA,用RNase-free水溶解,在检测其浓度和纯度后,再根据Prime ScriptTM RT Reagent Kit with gDNA Eraser(perfect real-time)试剂盒反转录合成cDNA第一链,-20℃保存备用或直接进行下一步试验。1.3 重组蛋白的原核表达和纯化
根据CsasCSP16的开放阅读框序列[24],于使用Ni-NTA亲和层析柱对表达在沉淀中的重组蛋白进行纯化后,放入透析袋中,经过(6、4、3、2、1、0 mol·L-1)梯度尿素缓冲液进行透析复性,每种梯度透析2 h[27]。透析缓冲液配方:5%甘油,1% L型精氨酸,0.1 mol·L-1 EDTA,尿素,使用1×PBS溶解。
1.4 荧光竞争结合试验
N-苯基-1-萘胺(1-NPN)用作CSP结合亲和性研究的探针,选择35种小分子化合物,包括33种苹果挥发物和2种桃小食心虫信息素组分。使用色谱级甲醇将1-NPN和气味物质配制成1 mmol·L-1的工作液,蛋白用50 mmol·L-1的Tris-HCl缓冲液(pH 7.4)稀释成2 μmol·L-1,设定激发波长为337 nm,扫描发射光波长范围350—600 nm。在浓度为2 μmol·L-1的CsasCSP16蛋白溶液中,逐一加入终浓度为0—20 μmol·L-1的1-NPN,直至荧光强度达到饱和,根据Scatchard方程计算CsasCSP16与1-NPN的结合常数。然后测定气味物质与CsasCSP16的结合能力。将气味物质加入CsasCSP16与1-NPN的混合液体系中,气味标样浓度依次从2 μmol·L-1梯度递增。每种气味物质重复测试3次。使用公式Ki= [IC50]/(1+[1-NPN]/K1-NPN)计算结合常数,其中IC50为荧光强度降至一半时加入的配基浓度,[1-NPN]是游离的1-NPN浓度,K1-NPN为重组蛋白CsasCSP16与1-NPN的结合常数[27]。1.5 同源建模和分子对接
使用Modeller(version-9.19)构建CsasCSP16的三维结构模型,选择CsasCSP16的氨基酸序列作为探针在PDB蛋白数据库中搜索,根据晶体结构的R-因子、序列连续性和结构相似性选择最佳模板。使用Align2D软件比对CsasCSP16与模板序列。Modeller中的自动建模程序生成100个构象,首先用共轭梯度法(conjugate gradient,CG)优化每个构象。其次使用DOPE(discrete optimized protein energy)值来衡量CsasCSP16构象的稳定性,最后使用Profile-3D方法和在线工具Swiss-model(运用openbabel(
1.6 分子动力学模拟
使用GROMACS(2019.3)进行能量最小化和分子动力学模拟[30]。蛋白的拓扑结构使用CHARMM36力场自动生成[31]。配体的拓扑结构使用CGenFF服务器(1.7 CsasCSP16蛋白复合物结合自由能计算
从分子动力学模拟过程中得到的配体与受体蛋白的相对结合自由能通常比分子对接得分具有更高的试验结果相关性[32]。运用MM-PBSA方法计算得到CsasCSP16与配体分子之间的相对结合自由能ΔGbind[33]。相对结合自由能计算中熵的变化可以忽略,ΔEint为配体与蛋白间的相互作用,ΔEele为静电作用力,ΔEvdW为范德华力,ΔGsol为溶剂化作用,ΔGPB为极性溶剂化作用,ΔGSA为非极性溶剂化作用。非极性化与极性化作用计算采用APBS软件,溶剂的介电常数设为80,复合物的介电常数设为1。从分子动力学模拟40—50 ns轨迹中每隔100 ps取一个构象,获得相对结合自由能值,配体分子与CsasCSP16蛋白的相对结合自由能表明配体与蛋白结合能力的强弱,也反映配体分子存在于活性口袋中保持稳定性的能力,这种能力往往是决定CsasCSP16与配体分子是否结合的关键因素。
1.8 CsasCSP16蛋白质复合物关键氨基酸能量分解
基于残基分解的方法,对配体分子与氨基酸残基之间的相互作用进行分析,定性描述配体分子与CsasCSP16的结合机理[34]。复合物中每个残基分解能可以进一步分解为主链、侧链和总的分解能。每个配体分子-残基对(ΔGCsasCSP16-residue)的相互作用表示为:使用g_mmpbsa计算出配体分子与每个CsasCSP16残基之间的范德华力(ΔEvdW)和静电作用力(ΔEele)。ΔGPB表示由PB模型计算得出的对溶剂化自由能的静电贡献。通过基于相应SASA的经验模型获得了对溶剂化自由能(ΔGCAVITY)的非极性贡献。内部能量(ΔEint)为零,因为它基于单个轨迹并且忽略了熵项(TΔS)。PB非极性溶剂化能量目前不可分解。
2 结果
2.1 CsasCSP16重组蛋白的表达与纯化
通过终浓度为1 mmol·L-1 IPTG 诱导结果显示,在14 kD左右出现一条蛋白条带。SDS-PAGE电泳显示CsasCSP16重组蛋白主要在包涵体中表达,因此对包涵体进行复性处理后过柱纯化获得了CsasCSP16重组蛋白。在SDS-PAGE条带上的大小与预测的分子量大小一致(图1)。图1
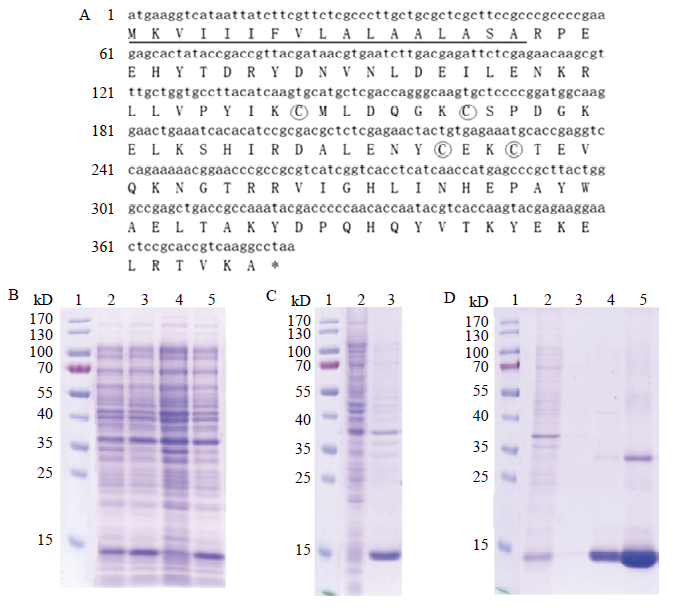 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1CsasCSP16重组蛋白的原核表达和蛋白纯化
Fig. 1The prokaryotic expression and protein purification of recombinant protein CsasCSP16
A:CsasCSP16 cDNA核苷酸序列及其推导的氨基酸序列,信号肽序列用下划线标注,保守的半胱氨酸残基用圆圈标注,终止密码子用星号表示 Nucleotide and deduced amino acid sequence of CsasCSP14 cDNA. Signal peptide is underlined, conserved cysteine residues are marked in circles, the stop codons are denoted by asterisks [25]。B:CsasCSP16在大肠杆菌中的表达Induciton of the CsasCSP16 in E. coli;1:蛋白分子量标签 Protein molecular weight marker;4:未诱导对照组 Control group without induction;2、3和5:IPTG诱导的表达产物 Expressed products induced by IPTG。C:超声破菌结果The result of sonication;1:蛋白分子量标签 Protein molecular weight marker;2:重组菌的上清蛋白The soluble fraction of recombinant E. coli;3:包涵体 Inclusion body。D:CsasCSP16重组蛋白的纯化 Purification of the CsasCSP16 recombinant protein;1:蛋白分子量标签 Protein molecular weight marker;2:上柱前蛋白 The proteins before purified by NTA-Ni affinity chromatography;3:流出液 Outflow;4:30 mmol·L -1 咪唑 Imidazole at 30 mmol·L -1;5:300 mmol·L-1咪唑 Imidazole at 300 mmol·L-1
2.2 CsasCSP16与配基的结合
为了进一步确定CsasCSP16在嗅觉方面的潜在作用,选择33种气味的化合物和2种信息素作为配体,运用荧光竞争结合试验验证CsasCSP16的结合特性(表1)。荧光探针1-NPN与CsasCSP16有良好的结合特性,其解离常数(Kd)为9.408 μmol·L-1,表明1-NPN可用于后续的竞争结合试验。根据荧光竞争结合曲线(图2)计算出CsasCSP16与配体分子结合的IC50和Ki值。结果表明,CsasCSP16与水杨酸甲酯、6-甲基-5-庚烯-2-酮、十五烷、庚酸丁酯和α-蒎烯有较强的结合活性,其Ki值分别为6.59、6.25、3.50、6.73和4.47 μmol·L-1。Table 1
表1
表1重组蛋白CsasCSP16 与不同寄主挥发物和性信息素分子结合的数据
Table 1
| 寄主挥发物和性信息素分子 Host volatile and sex pheromone molecule | CSP16 | 结合能 Binding energy (kJ·mol-1) | ||
|---|---|---|---|---|
| IC50 (μmol·L-1) | Ki (μmol·L-1) | |||
| 醇类 Alcohols | 2-己烯醇 2-hexenol | >50 | >50 | - |
| 苯乙醇 Phenethyl alcohol | >50 | >50 | - | |
| 醛类 Aldehydes | 反-2-己烯醛 (E)-2-hexenal | >50 | >50 | - |
| 己醛 Hexanal | >50 | >50 | - | |
| 苯甲醛 Benzaldehyde | >50 | >50 | - | |
| 庚醛 Heptanal | >50 | >50 | - | |
| 辛醛 Octanal | >50 | >50 | - | |
| 壬醛 Nonanal | >50 | >50 | - | |
| 癸醛 Decanal | >50 | >50 | - | |
| 酯类 Esters | 丁酸乙酯 Ethyl butyrate | >50 | >50 | - |
| 乙酸丁酯 Butyl acetate | >50 | >50 | - | |
| 乙酸异戊酯 Isoamyl acetate | >50 | >50 | - | |
| 乙酸-顺-3-己烯酯 (Z)-3-hexenyl acetate | >50 | >50 | - | |
| 丁酸丁酯 Butyl butyrate | >50 | >50 | - | |
| 己酸乙酯 Ethyl hexanoate | >50 | >50 | - | |
| 乙酸己酯 Hexyl acetate | >50 | >50 | - | |
| 水杨酸甲酯 Methyl salicylate | 18.69 | 6.59 | -25.52 | |
| 庚酸乙酯 Ethyl heptanoate | >50 | >50 | - | |
| 己酸丁酯 Butyl hexanoate | >50 | >50 | - | |
| 茉莉酸甲酯 Methyl jasmonate | >50 | >50 | - | |
| 乙酸二甲基丁酯 Dimethyl butyl acetate | >50 | >50 | - | |
| 辛酸正丁酯 Butyl octanoate | >50 | >50 | - | |
| 庚酸丁酯 Butyl heptanoate | 19.74 | 6.73 | -21.76 | |
| 辛酸丙酯 Propyl octanoate | >50 | >50 | - | |
| 萜类 Terpenoids | α-蒎烯 α-pinene | 11.50 | 4.47 | -27.61 |
| α-罗勒烯 α-ocimene | >50 | >50 | - | |
| 腈类 Benzonitriles | 苯甲腈 Benzonitrile | >50 | >50 | - |
| 烷烃类 Alkanes | 癸烷 Decane | >50 | >50 | - |
| 十四烷 Tetradecane | >50 | >50 | - | |
| 十五烷 Pentadecane | 7.90 | 3.50 | -24.27 | |
| 十六烷 Hexadecane | >50 | >50 | - | |
| 十八烷 Octadecane | >50 | >50 | - | |
| 酮类物质 Ketone | 6-甲基-5-庚烯-2-酮 6-methyl-5-hepten-2-one | 17.83 | 6.25 | -22.16 |
| Z-7-二十烯-11-酮 Z-7-eicosen-11-one | >50 | >50 | - | |
| Z-7-十九烯-11-酮 Z-7-nonadecen-11-one | >50 | >50 | - | |
新窗口打开|下载CSV
图2
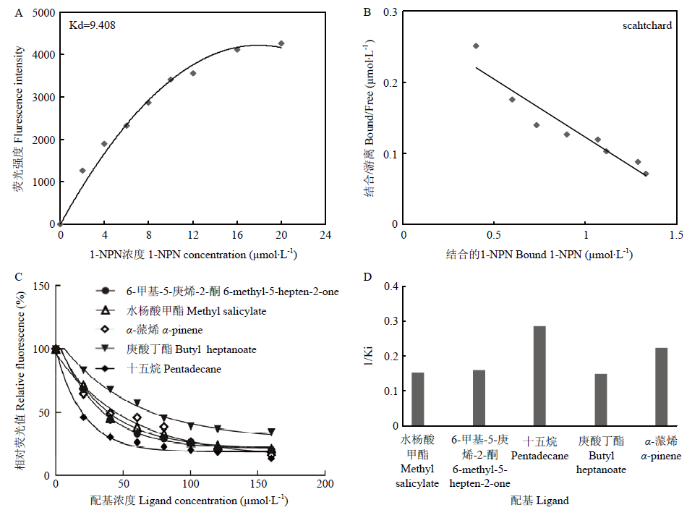 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2CsasCSP16 的配体结合试验
Fig. 2Ligand-binding assay for CsasCSP16
A、B:CsasCSP16与1-NPN的结合Binding curves of CsasCSP16 to 1-NPN;C:CsasCSP16与配体的荧光竞争结合Fluorescence competition of CsasCSP16 with ligands;D:CsasCSP16对不同气味物质的结合亲和力(以1/Ki表示)的比较Comparison of the binding affinities (indicated by 1/Ki) of CsasCSP16 with different components
2.3 CsasCSP16蛋白质三维模型构建
选择一致性(69%)最高的CSPMbraA6(登录号:1N8U)晶体结构为模板构建CsasCSP16蛋白质的三维模型(图3)。拉氏图评价结果表明有94.4%的氨基酸残基在最适区域内,5.6%的氨基酸残基在允许区域内(图4)。Profile-3D的方法得到Verify Score值为37.81,远远高于要求的最低Verify Score值。CsasCSP16与CSPMbraA6叠合非常好,RMSD值为0.159(图3-B)。综上,CsasCSP16的分子三维模型是合理的,可用于后续的研究工作。图3
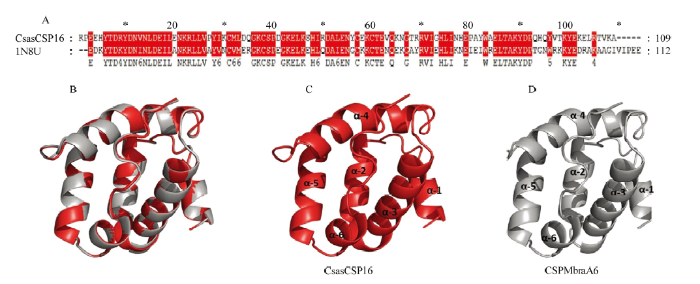 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3CsasCSP16的三维模型
Fig. 3Three-dimensional (3D) structure of CsasCSP16
A:CsasCSP16与1N8U序列比对Alignment of CsasCSP16 and 1N8U sequence;B:CsasCSP16与1N8U的三维结构比对Superimposed structure of CsasCSP16 and the template 1N8U;C:CsasCSP16三维结构模型Three-dimensional structure of CsasCSP16;D:1N8U的三维结构模型 Three-dimensional structure of 1N8U used as a template
图4
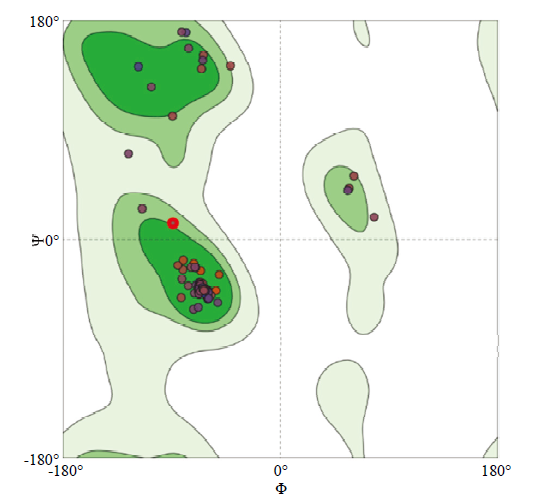 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4CsasCSP16三维结构拉氏图
深绿色区域内为氨基酸残基的最佳区域,绿色区域内为氨基酸残基较合适的区域,其他区域为氨基酸残基勉强许可区和不合理区
Fig. 4The Ramachandran plot of CsasCSP16
In the figure, the dark green regions are the best regions of the protein residues, the green regions are the appropriate regions of the protein residues, other regions are the irrational areas for protein residues
将CsasCSP16与水杨酸甲酯、6-甲基-5-庚烯-2-酮、十五烷、庚酸丁酯和α-蒎烯5种气味分子进行分子对接,得到的Vina Score值分别为-6.1、-5.3、-5.8、-5.2和-6.6。分子对接的结果与试验数据相吻合。由CB-DOCK预测的结合口袋得出,5种气味分子与CsasCSP16的结合部位均在疏水性结合腔内部的同一位置(图5)。
图5
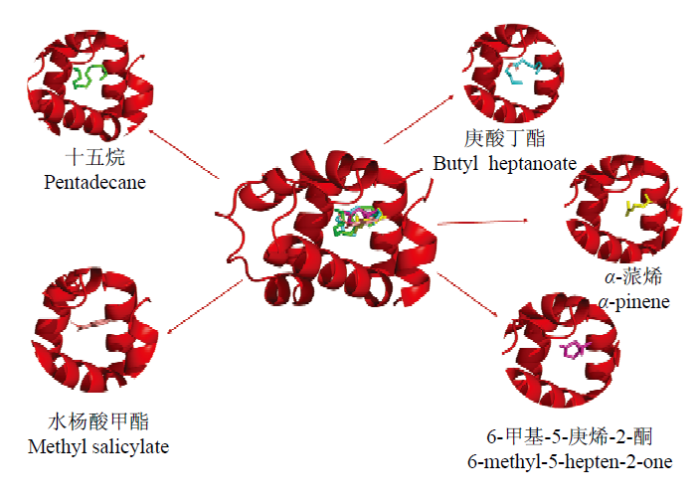 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5CsasCSP16在预测的结合口袋与不同气味分子结合模式
Fig. 5Binding modes of the different ligands in the putative binding pocket of CsasCSP16
2.4 CsasCSP16与气味分子的动力学模拟
为了使CsasCSP16与配体分子复合物的结构与晶体结构更接近,将CsasCSP16与配体复合物放置于TIP3P显性溶剂中进行了50 ns的动力学模拟。以CsasCSP16与配体分子在加热阶段的构象作为参考结构,使用RMSD值评估复合物的稳定性。结果如图6所示,CsasCSP16与水杨酸甲酯形成的复合物在10 ns时已经收敛达到平衡,RMSD值在0.2 nm左右波动;CsasCSP16与6-甲基-5-庚烯-2-酮形成的复合物在30 ns时达到平衡状态,RMSD值稳定在0.3 nm左右波动;CsasCSP16与十五烷形成的复合物在30 ns时达到收敛平衡,RMSD值在0.4 nm左右波动,CsasCSP16与十五烷的RMSD值波动较大,这可能与十五烷在结合腔内结构变化较大有关;CsasCSP16与庚酸丁酯在20 ns处达到收敛平衡,RMSD值稳定在0.25 nm左右。由于α-蒎烯在形成拓扑文件时,结构罚分值超过50,故未能进行动力学模拟。以上结果说明复合物已经达到平衡状态,动力学模拟的结果达到期望值。图6
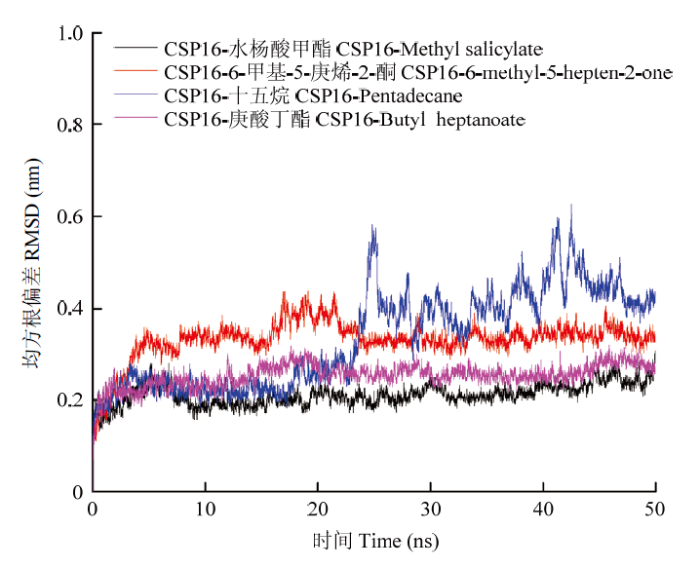 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6CsasCSP16与配体复合物主链Cα原子RMSD随时间变化曲线
Fig. 6RMSD value for the whole backbone of atoms of CsasCSP16-ligand complex
2.5 结合自由能
采用MM-PBSA的方法计算CsasCSP16与配体的结合自由能[33]。CsasCSP16-水杨酸甲酯、CsasCSP16-6-甲基-5-庚烯-2-酮、CsasCSP16-十五烷和CsasCSP16-庚酸丁酯的结合自由能分别为-50.264、-65.551、-136.035和-93.805 kJ·mol-1,显然CsasCSP16可以与4种配体分子形成稳定的复合物。该结果与荧光竞争结合试验一致。结合自由能可以反映出哪些能量分项影响配体的亲和力。从表2中发现范德华力能量向提供了主要的驱动力,其次为静电作用力和非极性溶剂化能。溶剂化自由能非极性部分能提供负能量,不利于CsasCSP16与配体的结合,这是由于非极性的溶剂与配体分子相互排斥造成的。残基与配体的空间结构和相互作用力的类型可以揭示这些氨基酸残基与配体的关键相互作用。从图7看出,CsasCSP16与配体分子主要是以范德华力连接在一起,并且水杨酸甲酯与苏氨酸68(Thr68)形成氢键。从CsasCSP16与配体气味分子三维结构图(图8)可以看出,气味分子大多被包围在由疏水性氨基酸组成的结合腔。Table 2
表2
表2CsasCSP16与配体分子的结合自由能和组分
Table 2
| 配体 Ligand name | 范德华力 Val der Waals (kJ·mol-1) | 静电作用力 Electrostatic force (kJ·mol-1) | 极性溶剂化能 Polar solvation energy (kJ·mol-1) | 非极性溶剂化能 Nonpolar solvation energy (kJ·mol-1) | 结合能 Binding energy (kJ·mol-1) |
|---|---|---|---|---|---|
| 水杨酸甲酯 Methyl salicylate | -82.466 (0.682) | -18.084 (0.850) | 61.336 (0.750) | -11.103 (0.055) | -50.264 (0.702) |
| 6-甲基-5-庚烯-2酮 6-methyl-5-hepten-2-one | -93.570 (0.927) | -16.158 (1.325) | 55.604 (0.973) | -11.428 (0.061) | -65.551 (1.080 ) |
| 十五烷 Pentadecane | -164.748 (0.845) | -0.969 (0.069) | 49.259 (0.848) | -19.537 (0.089) | -136.035 (1.047) |
| 庚酸丁酯 Butyl heptanoate | -109.800 (0.787) | -1.567 (0.211) | 33.365 (0.782) | -15.787 ( 0.079) | -93.805 (1.191) |
新窗口打开|下载CSV
图7
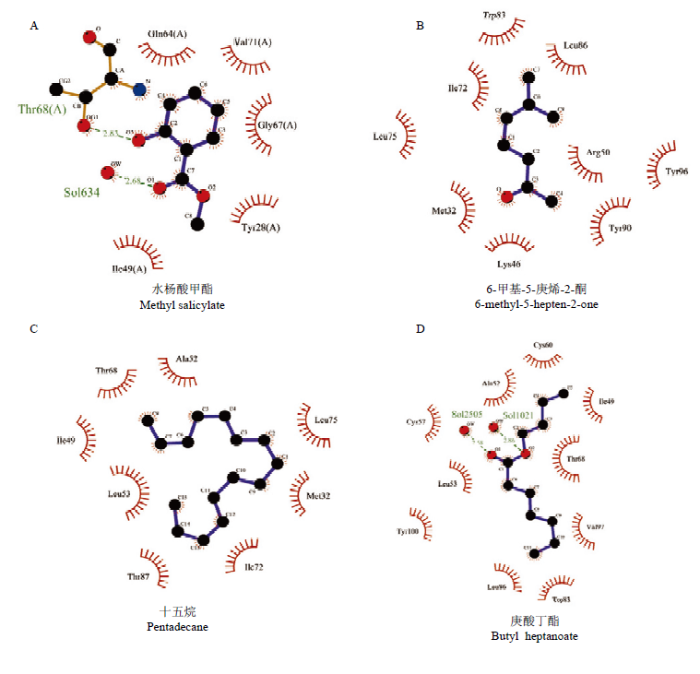 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7CsasCSP16与配体相互作用的关键氨基酸残基(二维结构)
Fig. 7The key residues of the different ligands that interact with CsasCSP16 (two dimensional)
图8
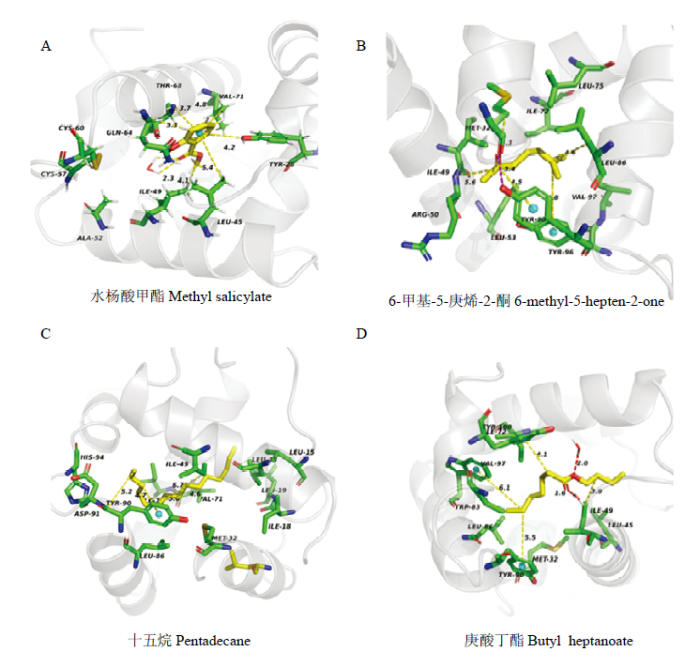 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8CsasCSP16 与配体相互作用的氨基酸残基(三维结构)
Fig. 8The key residues of the different ligands that interact with CsasCSP16 (three dimensional)
2.6 能量分解
使用MM-PBSA方法得到单个氨基酸残基水平(per-residue level)的结合自由能贡献值,其数值结果为单个氨基酸残基与配体分子相互作用的总和。单个残基能量贡献详细情况如图9和表3所示,在CsasCSP16与水杨酸甲酯形成的复合物中,有6个氨基酸残基的能量贡献>1 kJ·mol-1。其中异亮氨酸49(Ile49)贡献的能量高达6.2 kJ·mol-1。相反,酪氨酸28(Tyr28)、谷氨酰胺64(Gln64)和苏氨酸68(Thr68)不利于水杨酸甲酯的结合,提供了负面的能量贡献;在CsasCSP16与6-甲基-5-庚烯-2-酮形成的复合物中,有11个氨基酸残基贡献的能量>1 kJ·mol-1,相反,精氨酸50(Arg50)提供负面的能量贡献,这是由于Arg50属于亲水性氨基酸,不利于CsasCSP16与6-甲基-5-庚烯-2-酮形成复合物。CsasCSP16与十五烷形成的复合物中,有14个氨基酸的能量贡献>1 kJ·mol-1,精氨酸91(Asp91)和组氨酸94(His94)提供反向结合的能量。CsasCSP16与庚酸丁酯形成的复合物中有14个氨基酸残基贡献的能量>1 kJ·mol-1,利于CsasCSP16与配体相结合的氨基酸属于疏水性氨基酸,Ile49、缬氨酸71(Val71)、异亮氨酸72(Ile72)和酪氨酸90(Tyr90)贡献的能量>3 kJ·mol-1,并且Ile49参与了CsasCSP16与4种气味分子结合的过程,由此可以推断Ile49、Val71、Ile72和Tyr90可能是CsasCSP16与配体结合的关键氨基酸残基。图9
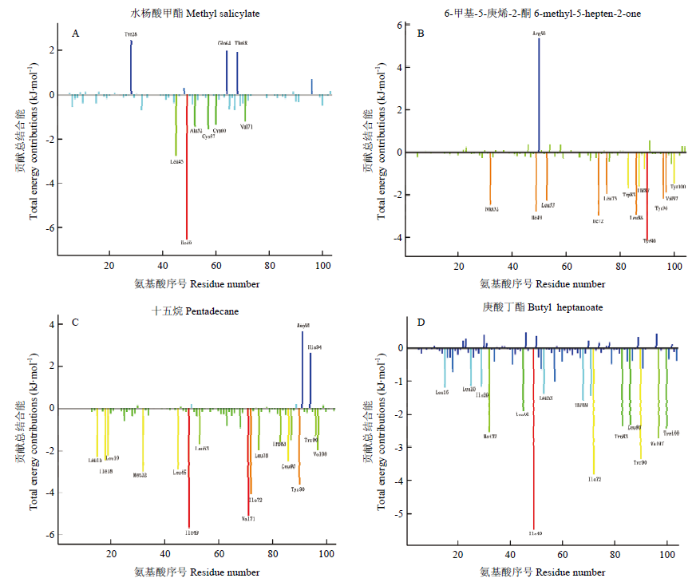 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9CsasCSP16 单个残基能量分解波谱图
X轴表示CsasCSP16氨基酸残基的序号数,Y轴表示配体与CsasCSP16氨基酸残基之间的相互作用能量大小。重要的残基用相应的文字标记
Fig. 9Single residue of CsasCSP16 energy decomposition spectrogram
The X-axis denotes the residue number of the CsasCSP16 and the Y-axis denotes the interaction energy between the ligand and the CsasCSP16 residues. The important residues for binding are marked by corresponding texts
Table 3
表3
表3主要残基对结合自由能的贡献
Table 3
| 气味分子 Volatile ligand | 相互作用能 Interaction energy (kJ·mol-1) | |||
|---|---|---|---|---|
| 范围 Range (>2.0 ) | 范围 Range (-1.0—-3.0 ) | 范围 Range (-3.0—-5.0) | 范围 Range (<-5.0 ) | |
| 水杨酸甲酯 Methyl salicylate | Tyr28, Gln64, Thr68 | Leu45, Ala52, Cys57, Cys60, Val71 | - | Ile49 |
| 6-甲基-5-庚烯-2酮 6-methyl-5-hepten-2-one | Arg50 | Met32, Ile49, Leu53, Ile72, Leu75, Trp83, Leu86, Thr87, Tyr96, Val 97, Tyr100 | Thr90 | - |
| 十五烷 Pentadecane | Asp91, His94 | Leu15, Ile18, Leu19, Met32, Leu45, Leu53, Leu75, Trp83, Leu86, Thr87, Tyr96, Val97 | Ile72, Tyr90 | Ile49, Val71 |
| 庚酸丁酯 Butyl heptanoate | - | Leu15, Leu25, Ile29, Met32, Leu45, Leu53, Cys57, Thr68, Val71, Trp83, Leu86, Val97, Tyr100 | Ile72, Tyr90 | Ile49 |
新窗口打开|下载CSV
3 讨论
在昆虫嗅觉感器中,化学感受蛋白结合疏水性的气味分子,穿过亲水性的淋巴液,传递给嗅觉神经元,将化学信号转化为电信号。本研究通过包涵体复性及Ni-NTA技术纯化得到了较高纯度的CsasCSP16蛋白,具有与天然蛋白一致的活性。ZHU等研究发现,含有His标签的融合蛋白与不带His标签的融合蛋白对气味分子的结合特性无影响[17]。结合特性研究结果表明,CsasCSP16与水杨酸甲酯、6-甲基-5庚烯-2-酮、十五烷、庚酸丁酯和α-蒎烯显示出较强的结合特性,推测CsasCSP16可能是桃小食心虫识别这些气味分子的靶标基因。在触角中高表达的其他昆虫的CSP与寄主挥发物分子也具有较高的结合活性。如苜蓿盲蝽(Adelphocoris lincolatus)AlinCSP4与法尼烯、β-法尼醇、β-紫罗兰酮有特异性的结合活性,AlinCSP1与3-己烯醇、2-己烯醛和正戊醛具有特异性结合的高亲和活性[35]。CsasCSP16不仅在触角中有表达,在雌、雄成虫翅中的表达量要高于其他组织,这与蜜蜂CSP3的表达谱比较一致,在翅中的功能有待于进一步研究[36]。ZHOU等研究发现,飞蝗(Locusta migratoria)翅上存在许多化学感觉器,推测在翅中高表达的化学感受蛋白与接收寄主挥发物的次生代谢物有关[37]。本研究通过荧光竞争结合试验筛选出5个与CsasCSP16结合的气味分子,为进一步开发高效的引诱剂提供了参考依据。荧光竞争结合试验可辨别CSP与不同气味分子的结合能力强弱,为了精确地揭示CSP与气味分子结合的具体作用机制,需要准确了解蛋白晶体结构。TIAN等通过对苹果蠹蛾(Cydia pomonella)性信息素结合蛋白(CpomPBP2)三维建模、分子对接预测了其与35种气味分子的亲和性,结果发现CpomPBP2与十二烷醇亲和性最高,荧光竞争结合表明CpomPBP2与十二烷醇抑制常数(Ki)值最小[38]。VENTHUR等用分子对接和分子动力学模拟预测欧洲葡萄蛾(Lobesia botrana)性信息素结合蛋白(LbotPBP1)与11种性信息素分子和6种寄主挥发物的结合机制,发现十二碳烯是LbotPBP1的最好配体[39]。FAN等在研究禾谷缢管蚜(Rhopalosiphum padi)RpadOBP3与EBF相互作用的机制时,在疏水结构域中发现了3个氨基酸残基与EBF结合紧密,分别为色氨酸71(Trp71)、色氨酸68(Trp68)、苯丙氨酸26(Phe26)[40]。这对于进一步研究桃小食心虫CSP与气味物质的作用机制具有指导性意义。
本研究进一步开展了CsasCSP16与荧光竞争结合筛选得到的5种气味分子的分子对接,发现水杨酸甲酯与CsasCSP16的苏氨酸68(Thr68)残基之间形成氢键。这与SANDLER等对家蚕(Bombyx mori)BmorPBP1与蚕蛾醇的研究结果一致,他们认为氢键在BmorPBP1专一性识别性信息素过程中发挥关键作用[41]。因此,CsasCSP16蛋白能够特异性识别水杨酸甲酯。水杨酸甲酯是许多植物的挥发物,由病虫害诱导产生,且可趋避害虫。如经斜纹夜蛾(Prodenia litura)危害的水稻产生大量的水杨酸甲酯,可显著趋避褐飞虱(Nilaparvata lugens)[42]。CHEN等研究发现,白背飞虱(Sogatella furcifera)CfurCSP5与水稻挥发物水杨酸甲酯也具有高亲和性[43]。CsasCSP16与高亲和性气味分子结合能研究发现,CsasCSP16-水杨酸甲酯结合自由能最高,为-50.264 kJ·mol-1,该结果与荧光竞争结合试验一致;CsasCSP16与气味分子的相互作用力主要是范德华力和疏水作用力,异亮氨酸49(Ile49)、缬氨酸71(Val71)、异亮氨酸72(Ile72)和酪氨酸90(Tyr90)贡献的结合能>3 kJ·mol-1,因此推测这4种氨基酸在CsasCSP16结合气味分子的过程中发挥关键作用。在CHEN等对白背飞虱CfurCSP5与水稻挥发物分子对接的研究中发现,CfurCSP5与具有高亲和性的气味分子结合过程中亮氨酸44(Leu44)、异亮氨酸64(Ile64)、丝氨酸98(Trp98)和苯丙氨酸101(Phe101)等疏水性氨基酸均参与其中[43]。同样地,在对小菜蛾CSPMbraA6与配体的研究中发现,亮氨酸(Leu)、异亮氨酸(Ile)和色氨酸(Trp)是与配体结合过程中的关键氨基酸[9]。因此,推测CsasCSP16与水杨酸甲酯结合最关键的氨基酸为异亮氨酸,水杨酸甲酯是否可以用作桃小食心虫寻找寄主的趋避物质有待于进一步探索。
4 结论
桃小食心虫CsasCSP16能与寄主植物的5种气味分子结合,可能参与桃小食心虫对寄主挥发物的识别。分子对接发现异亮氨酸49(Ile49)、缬氨酸71(Val71)、异亮氨酸72(Ile72)和酪氨酸90(Tyr90)可能是CsasCSP16与配体分子结合的关键氨基酸残基。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/brv.12339URLPMID:28480618 [本文引用: 1]

Odorant-binding proteins (OBPs) and chemosensory proteins (CSPs) are regarded as carriers of pheromones and odorants in insect chemoreception. These proteins are typically located in antennae, mouth organs and other chemosensory structures; however, members of both classes of proteins have been detected recently in other parts of the body and various functions have been proposed. The best studied of these non-sensory tasks is performed in pheromone glands, where OBPs and CSPs solubilise hydrophobic semiochemicals and assist their controlled release into the environment. In some cases the same proteins are expressed in antennae and pheromone glands, thus performing a dual role in receiving and broadcasting the same chemical message. Several reports have described OBPs and CSPs in reproductive organs. Some of these proteins are male specific and are transferred to females during mating. They likely carry semiochemicals with different proposed roles, from inhibiting other males from approaching mated females, to marking fertilized eggs, but further experimental evidence is still needed. Before being discovered in insects, the presence of binding proteins in pheromone glands and reproductive organs was widely reported in mammals, where vertebrate OBPs, structurally different from OBPs of insects and belonging to the lipocalin superfamily, are abundant in rodent urine, pig saliva and vaginal discharge of the hamster, as well as in the seminal fluid of rabbits. In at least four cases CSPs have been reported to promote development and regeneration: in embryo maturation in the honeybee, limb regeneration in the cockroach, ecdysis in larvae of fire ants and in promoting phase shift in locusts. Both OBPs and CSPs are also important in nutrition as solubilisers of lipids and other essential components of the diet. Particularly interesting is the affinity for carotenoids of CSPs abundantly secreted in the proboscis of moths and butterflies and the occurrence of the same (or very similar CSPs) in the eyes of the same insects. A role as a carrier of visual pigments for these proteins in insects parallels that of retinol-binding protein in vertebrates, a lipocalin structurally related to OBPs of vertebrates. Other functions of OBPs and CSPs include anti-inflammatory action in haematophagous insects, resistance to insecticides and eggshell formation. Such multiplicity of roles and the high success of both classes of proteins in being adapted to different situations is likely related to their stable scaffolding determining excellent stability to temperature, proteolysis and denaturing agents. The wide versatility of both OBPs and CSPs in nature has suggested several different uses for these proteins in biotechnological applications, from biosensors for odours to scavengers for pollutants and controlled releasers of chemicals in the environment.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/1744-7917.12116URLPMID:24677614 [本文引用: 1]
We analyzed 20 chemosensory protein (CSP) genes of the silkworm Bombyx mori. We found a high number of retrotransposons inserted in introns. We then analyzed expression of the 20 BmorCSP genes across tissues using quantitative real-time polymerase chain reaction (PCR). Relatively low expression levels of BmorCSPs were found in the gut and fat body tissues. We thus tested the effects of endectocyte insecticide abamectin (B1a and B1b avermectins) on BmorCSP gene expression. Quantitative real-time PCR experiments showed that a single brief exposure to insecticide abamectin increased dramatically CSP expression not only in the antennae but in most tissues, including gut and fat body. Furthermore, our study showed coordinate expression of CSPs and metabolic cytochrome P450 enzymes in a tissue-dependent manner in response to the insecticide. The function of CSPs remains unknown. Based on our results, we suggest a role in detecting xenobiotics that are then detoxified by cytochrome P450 anti-xenobiotic enzymes.
URLPMID:24796243 [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
URLPMID:21220518 [本文引用: 1]
URLPMID:21707107 [本文引用: 1]
[本文引用: 1]
URLPMID:30593726 [本文引用: 1]
DOI:10.1002/ps.5107URLPMID:29869368 [本文引用: 1]
BACKGROUND: The peach fruit moth, Carposina sasakii Matsumura (Lepidoptera: Carposinidae), is one of the most destructive pests of pome and stone fruits, while few studies of their molecular biology and physiology have been conducted. Research into CsasPBPs (Carposina sasakii pheromone binding proteins) and CsasGOBPs (Carposina sasakii general odorant binding proteins) may provide insights in to the mechanisms of olfaction in Carposina sasakii. RESULTS: In our study, results of real time quantitative polymerase chain reaction (qPCR) assays demonstrated that CsasPBP1-3 and CsasGOBP1-2 transcripts were abundantly expressed in the antennae of both sexes, suggesting they play a vital role in olfaction. In addition, to examine specific functional differences between pheromone binding proteins (PBPs) and general odorant binding proteins (GOBPs), fluorescence competitive binding assays were used to measured the binding affinities for the two sex pheromones and 18 apple plant volatiles. As a result, both PBPs and GOBPs showed stronger binding affinities to Z-7-eicosene-11-one than Z-7-nonadecene-11-one in two sex pheromones, whereas only PBP3 exhibited specific affinity towards both these two sex pheromone components, and PBP1 showed a high binding affinity to the sex pheromone components, and to other plant volatiles. In addition, GOBP1-2 displayed high binding affinity to general components of plant volatiles. CONCLUSION: Our study suggested CsasPBPs and CsasGOBPs play distinct physiological roles in the perception of sex pheromones and host plant volatiles. (c) 2018 Society of Chemical Industry.
URLPMID:30029592 [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1093/jee/tow170URLPMID:27498114 [本文引用: 2]
The purpose of this study was to evaluate the sublethal effects of the beta-cypermethrin on calling behavior and fecundity of a major fruit-boring pest of apple, Carposina sasakii Matsumura. The mating rate, fertility (total number of eggs laid per female), and adult longevity of adults were remarkably decreased as compared with that in control when the adults were exposed to 10% lethal concentrations (LC10) of beta-cypermethrin (LC10female symbol x LC10male symbol), and the age-specific survival rate (lx) was also negatively affected by sublethal beta-cypermethrin especially for the LC10female symbolx LC10male symbol mating combination. However, the age-specific fecundity (mx) was stimulated particularly in the combination of CKfemale symbolx LC10male symbol. Furthermore, the oviposition period was prolonged and the number of eggs was significantly increased for combinations of CKfemale symbolx LC10male symbol and LC10female symbolx CKmale symbol. In the mating experiments, males in control or LC10-beta-cypermethrin treatments preferred to mate with females in control. It might be because of lower calling rate of female survivors treated with sublethal beta-cypermethrin. Our data indicate that treatment of beta-cypermethrin had a sublethal effect on the development and production of C. sasakii, and their mating behavior changes in surviving adults that may contribute to assortative mating.
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 2]
URL [本文引用: 2]
URLPMID:19399780 [本文引用: 1]
DOI:10.1038/s41401-019-0228-6URLPMID:31263275 [本文引用: 1]
As the number of elucidated protein structures is rapidly increasing, the growing data call for methods to efficiently exploit the structural information for biological and pharmaceutical purposes. Given the three-dimensional (3D) structure of a protein and a ligand, predicting their binding sites and affinity are a key task for computer-aided drug discovery. To address this task, a variety of docking tools have been developed. Most of them focus on docking in the preset binding sites given by users. To automatically predict binding modes without information about binding sites, we developed a user-friendly blind docking web server, named CB-Dock, which predicts binding sites of a given protein and calculates the centers and sizes with a novel curvature-based cavity detection approach, and performs docking with a popular docking program, Autodock Vina. This method was carefully optimized and achieved ~70% success rate for the top-ranking poses whose root mean square deviation (RMSD) were within 2 A from the X-ray pose, which outperformed the state-of-the-art blind docking tools in our benchmark tests. CB-Dock offers an interactive 3D visualization of results, and is freely available at http://cao.labshare.cn/cb-dock/.
DOI:10.1093/bioinformatics/btt055URLPMID:23407358 [本文引用: 1]
MOTIVATION: Molecular simulation has historically been a low-throughput technique, but faster computers and increasing amounts of genomic and structural data are changing this by enabling large-scale automated simulation of, for instance, many conformers or mutants of biomolecules with or without a range of ligands. At the same time, advances in performance and scaling now make it possible to model complex biomolecular interaction and function in a manner directly testable by experiment. These applications share a need for fast and efficient software that can be deployed on massive scale in clusters, web servers, distributed computing or cloud resources. RESULTS: Here, we present a range of new simulation algorithms and features developed during the past 4 years, leading up to the GROMACS 4.5 software package. The software now automatically handles wide classes of biomolecules, such as proteins, nucleic acids and lipids, and comes with all commonly used force fields for these molecules built-in. GROMACS supports several implicit solvent models, as well as new free-energy algorithms, and the software now uses multithreading for efficient parallelization even on low-end systems, including windows-based workstations. Together with hand-tuned assembly kernels and state-of-the-art parallelization, this provides extremely high performance and cost efficiency for high-throughput as well as massively parallel simulations. AVAILABILITY: GROMACS is an open source and free software available from http://www.gromacs.org. SUPPLEMENTARY INFORMATION: Supplementary data are available at Bioinformatics online.
URLPMID:27819658 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1021/ci500020mURLPMID:24850022 [本文引用: 2]
Molecular mechanics Poisson-Boltzmann surface area (MM-PBSA), a method to estimate interaction free energies, has been increasingly used in the study of biomolecular interactions. Recently, this method has also been applied as a scoring function in computational drug design. Here a new tool g_mmpbsa, which implements the MM-PBSA approach using subroutines written in-house or sourced from the GROMACS and APBS packages is described. g_mmpbsa was developed as part of the Open Source Drug Discovery (OSDD) consortium. Its aim is to integrate high-throughput molecular dynamics (MD) simulations with binding energy calculations. The tool provides options to select alternative atomic radii and different nonpolar solvation models including models based on the solvent accessible surface area (SASA), solvent accessible volume (SAV), and a model which contains both repulsive (SASA-SAV) and attractive components (described using a Weeks-Chandler-Andersen like integral method). We showcase the effectiveness of the tool by comparing the calculated interaction energy of 37 structurally diverse HIV-1 protease inhibitor complexes with their experimental binding free energies. The effect of varying several combinations of input parameters such as atomic radii, dielectric constant, grid resolution, solute-solvent dielectric boundary definition, and nonpolar models was investigated. g_mmpbsa can also be used to estimate the energy contribution per residue to the binding energy. It has been used to identify those residues in HIV-1 protease that are most critical for binding a range of inhibitors.
URLPMID:18206907 [本文引用: 1]
URLPMID:22900060 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:26928635 [本文引用: 1]
URLPMID:31062883 [本文引用: 1]
DOI:10.1016/j.jinsphys.2017.07.014URLPMID:28778653 [本文引用: 1]
(E)-beta-farnesene (EBF) is the common active component of aphid alarm pheromone. Either or both of two orthologs of ordorant-binding proteins (OBPs), OBP3 and OBP7, recently reported in aphids, may be involved in EBF perception. The aim of this study was to investigate the respondence of the aphid Rhopalosiphum padi to its intraspecific alarm pheromone and which OBP is responsible for that response. We tested the olfactory response of the aphid R. padi to EBF and freshly crushed aphids. Then, we extracted the volatiles from crushed aphids using solid phase microextraction (SPME) for analysis with GCxGC-TOF/MS. We also cloned two OBPs cDNAs in R. padi (RpadOBP3 and RpadOBP7) and expressed them in competent Escherichia coli cells. Both recombinant proteins, RpadOBP3 and RpadOBP7, bound EBF well, with RpadOBP7 having specifically stronger affinity for EBF than for other volatiles. Based on the crystal structure of the OBPs with high identity, we performed homology modeling and analyzed the interactions between RpadOBPs and EBF. In conclusion, R. padi was repelled by both EBF and crushed aphids. EBF was identified as the only volatile that acted as the alarm pheromone. Our results indicated that OBP7 is a potential molecular target to control wheat aphids by disturbing their behaviors to the alarm pheromone.
URLPMID:10662696 [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1016/j.pestbp.2018.09.006URLPMID:30497699 [本文引用: 2]
The white-backed planthopper (WBPH) Sogatella furcifera is a notorious rice pest in Asia. Olfaction is crucial for the WBPH to seek and locate rice plants. However, its mechanism is still not fully understood. Chemosensory proteins (CSPs) are some of the important olfactory-related proteins. In this study, we first used a bacterial system to successfully express the recombinant, antennae-enriched protein SfurCSP5. Further, competitive fluorescence binding assays with 86 candidate ligands, including some known rice plant volatiles, showed that SfurCSP5 has high affinities for 2-tridecanone, 2-pentadecanone, and beta-ionone, which are known to be present in volatile mixtures that can attract rice planthoppers, and produced Ki values of 4.89, 4.09, and 1.39mumol/L, respectively. Additionally, homology modeling of the protein structure of SfurCSP5 showed that it possesses five alpha-helixes (alpha-1, alpha-2, alpha-3, alpha-4, and alpha-5), which is a non-typical feature of the insect CSPs. Finally, ligand docking results revealed that Leu-44, Ile-64, Phe-90, Trp-98, and Phe-101 are five hydrophobic residues that interact with all of the ligands, indicating their key involvement in the binding of SfurCSP5. Our study lays the foundation for an understanding of the olfaction mechanism of rice planthoppers.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}