 ,1, 王鹤潼1, 方东路2, 赵立艳2, 杨文建1, 裴斐1, 胡秋辉1
,1, 王鹤潼1, 方东路2, 赵立艳2, 杨文建1, 裴斐1, 胡秋辉1Nano-Packaging Preservative Mechanism of Flammulina filiformis After Harvest Based on Mitochondrial Energy Status Pathways
MA Ning,1, WANG HeTong1, FANG DongLu2, ZHAO LiYan2, YANG WenJian1, PEI Fei1, HU QiuHui1责任编辑: 赵伶俐
收稿日期:2020-03-21接受日期:2020-05-27网络出版日期:2020-08-16
| 基金资助: |
Received:2020-03-21Accepted:2020-05-27Online:2020-08-16
作者简介 About authors
马宁,Tel:13951729396;E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (3957KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
马宁, 王鹤潼, 方东路, 赵立艳, 杨文建, 裴斐, 胡秋辉. 基于线粒体能量代谢途径的金针菇采后纳米包装保鲜机制[J]. 中国农业科学, 2020, 53(16): 3356-3371 doi:10.3864/j.issn.0578-1752.2020.16.013
MA Ning, WANG HeTong, FANG DongLu, ZHAO LiYan, YANG WenJian, PEI Fei, HU QiuHui.
0 引言
【研究意义】金针菇(Flammulina filiformis)又称毛柄金钱菌,由于其美味和高营养特性,目前已成为世界第三大食用菌,不仅销售量逐年增长,更有广阔的开发前景[1,2]。然而采后金针菇含水量高、呼吸作用强、能量代谢旺盛[3],极易发生开伞、褐变和腐烂等品质劣变现象[4]。目前对于金针菇采后品质劣变的研究大多数集中于失重率、褐变度、开伞率等表观指标,内在机理的研究尚缺乏,而线粒体是植物能量代谢及能量物质(adenosine triphosphate,ATP)生成的主要场所。有研究表明,能量状态失衡是果蔬品质劣变的核心因素[5],由此推测金针菇采后品质劣变也可能是由能量状态失衡引起[3],因此,基于线粒体探究金针菇冷藏过程的能量状态很有必要,对金针菇保鲜技术体系研究具有重要意义。【前人研究进展】目前国内外对线粒体提取方法的研究有很多,主要有差速离心法[6,7]、不连续密度梯度离心法[8]及试剂盒法(如酵母试剂盒法[9]、普利莱试剂盒法[10])。密克等[8]利用蔗糖密度梯度离心法提取板栗疫病菌的线粒体,通过测定发现所制备的线粒体完整性好,且受到污染较少。ZHAO等[9]采用普利莱试剂盒提取的线粒体研究在大鼠中内质网和线粒体是否共同参与椎间盘突出的细胞凋亡和椎间盘退变。GUIROLA等[10]利用酵母线粒体提取试剂盒提取线粒体,进而研究酿酒酵母中线粒体内微量物质pif1对细胞内蛋白及微量元素的影响。纳米材料近年来在食品包装的应用研究中越来越受关注,常见的有金属及其氧化物纳米材料、二氧化硅及硅酸盐纳米材料、多糖基纳米材料、蛋白基纳米材料和复合纳米材料[11]。在实验室前期的研究中开发了一种添加了纳米银、纳米二氧化钛、纳米凹凸棒土、纳米二氧化硅的复合包装材料[12],该包装材料能有效抑制金针菇开伞、褐变、孢子增加等品质劣变,保鲜效果显著[12,13]。研究表明,采后果蔬的质量下降与能量供应密切相关[14,15]。课题组前期对乙烯含量、活性氧积累和线粒体微观结构等进行了研究,这些都与能量代谢有关[13,16]。另外对蛋白组学的检测发现纳米包装组金针菇中参与碳水化合物代谢的相关蛋白在冷藏期间发生了显著性差异表达[17]。研究发现ATP供应不足或生产效率下降会导致膜结构完整性破坏,并导致果蔬采后迅速衰老和质量下降[18,19,20]。阚娟等[21]报道线粒体复合体活性的下降也会导致以上的后果,并抑制能量物质合成,从而导致细胞能量供应不足。【本研究切入点】线粒体的研究(包括提取纯化)目前均集中在人、动物或植物上,应用于金针菇的几乎没有。【拟解决的关键问题】通过对常见线粒体提取方法的对比,确定适用于金针菇线粒体的最优提取方法。同时,从线粒体角度阐明不同包装对金针菇能量代谢的影响。此外,通过动态检测金针菇在冷藏期间ATP代谢系统物质含量变化,探明纳米包装在贮藏中维持金针菇能量代谢水平的作用机制。1 材料与方法
试验于2019年1—11月在南京财经大学食品科学与工程学院进行。1.1 试验材料
新鲜金针菇(F-C01l)从南京金万辰生物科技有限公司采购,1 h内低温运输至实验室。筛选的金针菇大小为菌盖直径3—4 cm、重量为23—30 g、未开伞、没有机械损伤。挑选后的金针菇进行液氮研磨,并用于后续线粒体提取方法筛选优化。金针菇菌种由同公司提供同品种菌种,培养至菌丝体时对菌丝进行过滤冲洗,将培养基冲洗干净后进行冻干处理,最后使用液氮研磨供后续线粒体提取方法筛选优化。1.2 仪器与试剂
CL21R型微量台式冷冻离心机,德国Thermo Electron LED公司;SX500型快速自动高压灭菌仪,日本鹿儿岛地震保险公司;HH2型数显恒温水浴锅,国华电器有限公司;Axlo Vert.A1型倒置式显微镜,德国卡尔蔡司股份公司;THZ-C型恒温振荡器,上海玺袁科学仪器有限公司;FreeZone 2.5升台式冻干机,北京照生行仪器设备有限公司;日立JEM-1011型透射电子显微镜,日本Hitachi公司;Clark氧电极,汉莎科学仪器有限公司;ELX800型酶标仪,美国Bio-Tek公司;1260系列高效液相色谱仪,美国Agilent公司。乙二胺四乙酸二钠试剂(EDTA)、甘露醇、Tris-HCl试剂、牛血清蛋白(BSA)、蔗糖、β-巯基乙醇、苯甲基黄酰氟(PMSF)、健那绿、麦芽糖、葡萄糖、酵母提取物、蛋白胨、MgSO4·7H2O、KH2PO4、琼脂粉、马铃薯葡萄糖琼脂培养基(PDA)、Gluta固定液、二甲砷酸钠缓冲液、锇酸、丙酮、Epon-812环氧树脂、磷酸氢二钾、磷酸二氢钾、甲醛(分析纯)、三磷酸腺苷、二磷酸腺苷、一磷酸腺苷、乙醇脱氢酶(ADH)活性检测试剂盒、线粒体呼吸链复合体Ⅳ活性检测试剂盒、超氧化物歧化酶(SOD)活性检测试剂盒、线粒体呼吸链复合体I活性检测试剂盒、线粒体呼吸链复合体III活性检测试剂盒均采购于北京索莱宝有限公司;酵母线粒体提取试剂盒采购于北京百奥莱博科技有限公司;线粒体-胞浆蛋白制备试剂盒采购于北京普利莱基因技术有限公司。
1.3 试验设计
1.3.1 线粒体的提取试验 改进普利莱法(Improved Pulilai Kit Method,IPKM):由于传统普利莱法没有对于细胞壁破碎的过程,更适用于动物细胞,因此根据吴媛等[22]的方法改进,先对金针菇细胞进行破壁处理,再进行线粒体的提取。具体步骤如下:将样品浸没至巯基化合物中,在32℃下静置15—30 min。10 000 r/min离心1 min,弃上清,沉淀重悬于山梨醇,每克样品加入蜗牛酶30—40 mg,37℃保温1 h。800×g离心5 min,将上清液丢弃,沉淀物用PBS洗3次,并于4℃、800×g离心5—10 min收集沉淀。加入1.5 mL冰预冷的Mito-Cyto Buffer重悬沉淀,4℃、800×g离心5 min,收集上清再次800×g离心5 min,收集上清后12 000×g再次离心10 min,管底沉淀即为线粒体。不连续密度梯度离心法(discontinuous density gradient centrifugation method,DDGCM):参照蔺凯丽等[23]的方法,将样品提取液以1﹕6的比例向样品中加入4℃预冷提取液(内含1 mmol?L-1 EDTA、0.3 mol?L-1甘露醇、50 mmol?L-1 Tris-HCl、0.2% BSA、2 mmol?L-1β-巯基乙醇),在组织匀浆机中匀浆3次,15 s/次。将匀浆液1 500×g离心5 min,取上清液3 000×g离心5 min,再取上清液12 000×g离心15 min,沉淀为粗线粒体。将沉淀分别用3种浓度的2 mL洗涤液洗涤(由于洗涤液的浓度高低影响线粒体的产量,因此采用3种浓度的洗涤液,并最终取最佳的一组与另两种方法组进行比较),首先配置初始浓度洗涤液:内含1 mmol?L-1 EDTA、0.3 mol?L-1甘露醇、10 mmol?L-1 Tris-HCl、0.5 mmol?L-1 PMSF,然后将上述浓度洗涤液分别稀释200倍和400倍,从而得到3个浓度的洗涤液,以下分别用DDGCM-1、DDGCM-200、DDGCM- 400来表示。洗涤过后12 000×g离心15 min,得沉淀为粗线粒体。将粗线粒体用悬浮液(内含1 mmol?L-1 EDTA、0.3 mol?L-1甘露醇、10 mmol?L-1Tris-HCl)悬浮。将粗线粒体悬浮液小心铺在15%﹕23%﹕32%﹕60%蔗糖不连续密度梯度上。4℃下134 000×g离心45 min。取32%和60%之间不透明浅白色线粒体环,用2倍体积悬浮液洗涤。4℃下12 000×g离心20 min,得到的沉淀即为高纯度线粒体。
酵母线粒体提取试剂盒方法(yeast mitochondrial extraction kit method,YMEKM):参照北京百奥莱博科技有限公司提供的酵母线粒体提取试剂盒说明书进行提取。
1.3.2 不同包装袋贮藏保鲜试验 冷藏试验中将金针菇在(4±1)℃和90%相对湿度下于黑暗中预冷24 h,并将样品(250±5 g)随机包装到纳米包装袋和普通PE包装袋中进行贮藏,每个处理组3个平行样品,对各包装组金针菇分别进行21袋的包装处理。然后将样品在(4±1)℃和90%相对湿度下存储21 d。冷藏期间每3 d利用优化得到的方法对各处理组样品进行线粒体提取,并进行生理生化分析。本研究中,所用的纳米包装袋是参考笔者实验室FANG等[24]的生产工艺得到。按质量比取纳米粉体15%(其中纳米银30%、纳米二氧化钛35%、纳米凹凸棒土25%、纳米二氧化硅10%)、低密度聚乙烯46%、低密度线性聚乙烯22%、分散剂10%、润滑剂5%和偶联剂2%制得纳米母粒,并按质量比取经上述制备的纳米母粒7.5%、防雾剂母粒8%和低密度线性聚乙烯84.5%制得纳米保鲜膜。普通PE包装袋是没有纳米复合母料纳米粉的聚乙烯袋,其尺寸和厚度(25 cm×25 cm,40 μm),与纳米包装袋相同。
1.4 检测方法
1.4.1 线粒体结构完整性的检测 乙醇脱氢酶(ADH)活性的测定:参照ADH活性检测试剂盒说明书进行测定。细胞色素C氧化酶活性的测定:参照线粒体呼吸链复合体Ⅳ活性检测试剂盒进行测定。
呼吸速率的测定:参照高永彬[25]的方法,使用Clark(Hansatech Ltd, Hardwick industrial Estate, King’s Lynn, Norfold, UK)公司的氧电极装置分别对各组提取的线粒体、金针菇子实体以及菌丝体进行呼吸氧消耗的测定,测定过程中保持室内25℃恒温状态。(1)线粒体呼吸速率的测定:测定前,用反应液进行校正,待温度恒定。取100 μL线粒体悬浮液定容到1 mL反应液中(反应液内含0.3 mol?L-1甘露醇、2 mmol?L-1MgSO4、5 mmol?L-1 KH2PO4、10 mmol?L-1TES、10 mmol?L-1 NaCl、0.1%(w/v)BSA,加入终浓度为10 mmol?L-1琥珀酸盐、5 mmol?L-1抗坏血酸、10 μmol?L-1 ATP、0.5 mmol?L-1 ADP、1 mmol?L-1 NADH、25 μmol?L-1细胞色素C等底物和0.05%(w/v)曲拉通100),并迅速转移至反应室中。开始测定,待耗氧速率稳定时开始记录。(2)金针菇子实体呼吸速率的测定:测定前,用超纯水进行校正,待温度恒定。取两个菌盖直径为0.3 cm,菌柄长1.5 cm的金针菇子实体加入反应室并使反应室内超纯水定容到1 mL,开始测定,待耗氧速率稳定时开始记录。(3)金针菇菌丝体呼吸速率的测定:测定前,用CYM培养基(内含1%麦芽糖、2%葡萄糖、0.2%酵母提取物、0.2%蛋白胨、0.05% MgSO4·7H2O、0.46% KH2PO4)进行校正,待温度恒定。取0.5 mL菌丝体定容到1mL培养基中,并迅速转移至反应室中。开始测定,待耗氧速率稳定时开始记录。
透射电镜观察:参照TARZE等[26]的方法,取纯化后的线粒体少许,在2.5% Gluta固定液于4℃固定2 h以上,用0.2 mol?L-1二甲砷酸钠缓冲液洗两次,每次15 min,再用1%的锇酸(以0.2 mol?L-1二甲砷酸钠配制)于4℃下固定2 h。之后进行脱水处理:4℃下分别在30%、50%和70%丙酮浸15 min,然后在室温下分别在80%和90%丙酮中浸一次,再在95%丙酮中浸两次,最后在100%丙酮浸两次,以上每次均浸15 min。脱水后的样品用Epon-812环氧树脂包埋并进行切片,挑选出厚度在50—70 nm的切片置于铜网上,用醋酸双氧铀和柠檬酸铅双重染色,最后在日立JEM-1011型透射电子显微镜下观察拍照。
1.4.2 线粒体功能活性的检测 超氧化物歧化酶(SOD)活性的测定:参照SOD活性检测试剂盒进行测定。
健那绿染色线粒体计数:参照高永超等[27]的方法,取线粒体悬浮液,滴一滴于载玻片,标记涂片,滴一滴0.02%健那绿染液,盖上盖片染色20 min。放于倒置显微镜下观察并拍照。线粒体呈蓝绿色颗粒状。拍照通过ImageJ软件进行计数。
1.4.3 能量水平和能荷值测定 参考WU等[28]的方法测定三磷酸腺苷(adenosine triphosphate,ATP)、二磷酸腺苷(adenosine diphosphate,ADP)、一磷酸腺苷(adenosine monophosphate,AMP)含量。标准溶液的制备:将ATP、ADP、AMP标准溶液分别稀释为6.25、12.5、25、50和100 mg?L-1的5个浓度梯度,得到的溶液用1 mol?L-1 KOH调至pH 6.5—6.8并定容,经0.45 μm微孔滤膜过滤。样品的制备:取0.5 g金针菇样品,加入6 mL冰冷的0.6 mol?L-1高氯酸冰浴研磨并静置10 min,3 000 r/min离心10 min,上清液与标液进行同样的调pH和过滤处理,滤液用于ATP、ADP和AMP含量测定,以上操作均在4℃条件下完成。制备好的标准溶液及样品在下述色谱条件下上机进样,并记录各标准品及样品峰面积。以不同浓度各标样峰面积y为纵坐标,以标样质量浓度x为横坐标,绘制标准曲线,回归方程及相关系数见表1。色谱条件:色谱柱(Eclipse plus C-18,5 μm×150 mm×4.6 mm);流动相:A:0.06 mol?L-1的K2HPO4+0.04 mol?L-1的KH2PO4(pH 7.0),B:100%甲醇;柱温:30℃;流速:1 mL?min-1;检测波长:254 nm;洗脱条件:ATP:50%A,50%B;ADP、AMP:95%A,5%B。洗脱时间:4 min。
Table 1
表1
表1
Table 1
| 能量组分 Component of energy charge | 回归方程 Regression equation | 相关系数 Correlation coefficient |
|---|---|---|
| ATP | y=29.933x-4.275 | R2=0.9998 |
| ADP | y=27.48x+28.358 | R2=1 |
| AMP | y=36.92x+12.892 | R2=1 |
新窗口打开|下载CSV
能量负荷值测定:能量负荷值=(ATP+1/2ADP)/(ATP+ADP+AMP)。
1.4.4 线粒体关键复合体酶活性的测定 线粒体复合体I、Ⅲ、Ⅳ活性的测定:利用1.3.1中对比确定的最佳线粒体提取方法对金针菇线粒体进行提取,并参照北京索莱宝科技有限公司线粒体复合体I、Ⅲ、Ⅳ活性检测试剂盒中说明书提供的方法进行活性测定。
1.5 数据处理与分析
使用Excel进行数据处理,并用Origin 8.5绘图,SPSS Statistics17.0进行显著性分析,显著性水平设置为P<0.05。2 结果
2.1 不同提取方法对金针菇线粒体结构完整性的影响
2.1.1 乙醇脱氢酶活性 乙醇脱氢酶(alcohol dehydrogenase,ADH)和细胞色素C氧化酶(cytochrome c oxidase,COX)是分别稳定存在于胞液及线粒体内的标志性蛋白,当线粒体膜发生破碎时,二者会分别向线粒体及胞液中流动,因此,通过测定它们在线粒体及胞液中的活性,可以来反映线粒体膜的破损情况[20]。在不连续密度梯度离心法(DDGCM)中,使用的洗涤液浓度高低会影响线粒体的产量,因此本研究使用3种浓度的洗涤液(DDGCM-1、DDGCM-200、DDGCM-400)进行筛选以确定最佳提取参数。由图1可见,在子实体组,DDGCM和酵母试剂盒法(YMEKM)所提取的线粒体中ADH活性均比改进普利莱法(IPKM)的活性高。其中,虽然DDGCM未稀释洗涤液组所提取的线粒体中ADH活性最低,但仍然高于IPKM。另外,IPKM所得胞液中的ADH活性最高,与各组都存在显著性差异。由图2可见,在菌丝体组,虽然各方法得到的胞液中ADH活性结果与子实体组相同,但是线粒体ADH活性中YMEKM所得与DDGCM-200相比无显著性差异,而IPKM所得活性仍然最低。因此,相比之下,IPKM提取得到的线粒体受胞质污染最轻。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1金针菇子实体所提线粒体(A)及胞液(B)中乙醇脱氢酶活性
不同的字母表示差异显著(P<0.05)。下同
Fig. 1Activity of alcohol dehydrogenase in mitochondria (A) and cytosol (B) of F. filiformis fruiting body
Different letters on bars indicated significantly different among treatments (P<0.05). The same as below
图2
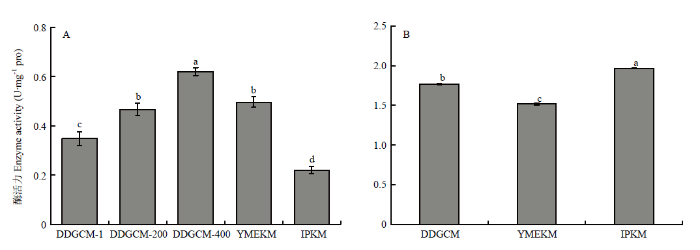 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2金针菇菌丝体所提线粒体(A)及胞液(B)中乙醇脱氢酶活性
Fig. 2Activity of alcohol dehydrogenase in mitochondria (A) and cytosol (B) of F. filiformis mycelium
2.1.2 细胞色素C氧化酶活性 由图3可见,在子实体组中,DDGCM提取的线粒体中COX活性随着洗涤液浓度的降低而降低,尽管未稀释洗涤液组在3个梯度中COX活性最高,并且显著高于YMEKM,但显著低于IPKM所提取线粒体中的COX活性。另外,胞液中的COX活性结果显示YMEKM最高,DDGCM其次,IPKM的最低,并且三者均有显著性差异。综上所述,IPKM所提取的线粒体受胞质污染最轻,与2.1.1得到的结果一致。此外,由图4可见,在菌丝体组中,所得结果与子实体趋势一致。
图3
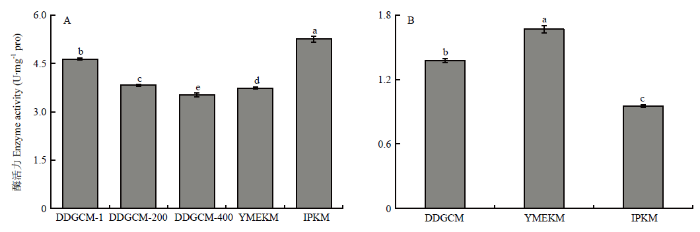 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3金针菇子实体所提线粒体(A)及胞液(B)中细胞色素C氧化酶活性
Fig. 3Activity of Cytochrome C oxidase in mitochondria (A) and cytosol (B) of F. filiformis fruiting body
图4
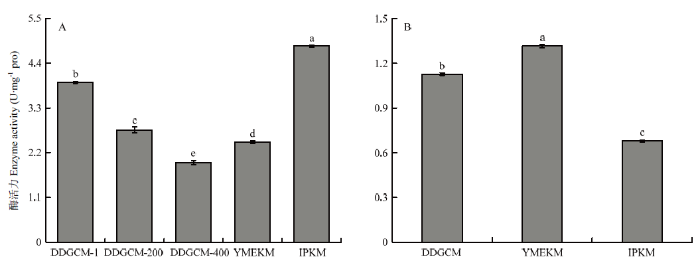 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4金针菇菌丝体所提线粒体(A)及胞液(B)中细胞色素C氧化酶活性
Fig. 4Activity of Cytochrome c oxidase in mitochondria (A) and cytosol (B) of F. filiformis mycelium
2.1.3 线粒体呼吸速率 细胞色素C氧化酶损失会直接导致线粒体呼吸能力下降,呼吸速率降低[29]。因此,为了进一步验证分离线粒体的呼吸能力,本研究使用氧电极对线粒体呼吸速率进一步评估。通过测定线粒体提取前后的呼吸速率,对比各方法在提取后的呼吸速率差异来评估它们对于线粒体膜的损伤程度。由图5可知,无论是子实体还是菌丝体中,IPKM所提的线粒体与未提取前差距最小,且显著高于其他两种方法所提线粒体的呼吸速率,证明该方法提取得到的线粒体能更好地维持其功能活性。虽然使用多个呼吸链底物配制线粒体反应液,但提取的线粒体呼吸速率与细胞内呼吸相比较低,这可能与反应液模拟细胞环境存在误差有关。随着菌丝生长,呼吸速率呈上升趋势。在不同生长时期,菌丝的呼吸速率增速不同[30],因此与子实体相比,菌丝体的呼吸速率相对较慢。
图5
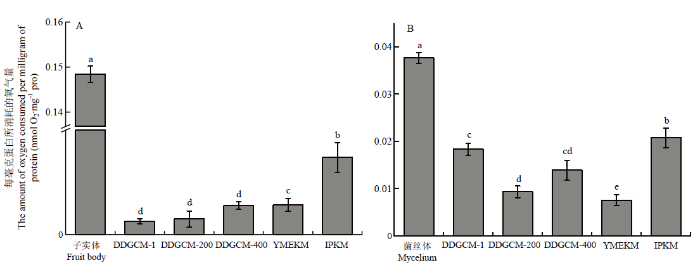 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5金针菇子实体(A)、菌丝体(B)线粒体的呼吸氧消耗
Fig. 5The number of active mitochondria extracted from the fruit body (A) and mycelium (B) of F. filiformis
2.1.4 透射电镜观察线粒体 在图6的显微镜照片中,所有样品组中都可见典型的椭圆形线粒体。其中,DDGCM随着洗涤液的逐级稀释,具有完整结构的线粒体也越来越少,并且肿胀现象也越来越严重。YMEKM制备的线粒体肿胀并表现出线粒体膜破碎严重,线粒体膜碎片在该组图片中清晰可见。而IPKM提取得到的线粒体虽然也存在少量的碎片,但得到的形态结构完整的线粒体数量最多,与其他组样品形成明显对比。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6透射电镜观察线粒体微观结构
M:线粒体;MF:线粒体碎片;放大倍数:20000倍
Fig. 6TEM observation of mitochondrial microstructure
Mitochondria; MF: Mitochondrial fragments; Magnification: 20000×
2.2 不同提取方法对金针菇线粒体功能活性的影响
2.2.1 超氧化物歧化酶活性 超氧化物歧化酶(superoxide dismutase,SOD)是细胞中重要的抗氧化酶,分为3大类,分别为Cu/Zn-SOD、Mn-SOD、Fe-SOD。其中存在于线粒体内膜基质中的是Mn-SOD,与线粒体的呼吸代谢有关[31],并维持线粒体膜的完整性和正常功能[32]。在不连续密度梯度离心法中,随着洗涤液浓度的降低,所提线粒体中SOD活性逐渐降低,因此,选取未稀释洗涤液组所提取的线粒体中SOD活性与其他两个方法进行对比。由图7可见,IPKM所提取的线粒体中SOD活性最高,酵母试剂盒法提取的线粒体中SOD活性最低,子实体组与菌丝体的趋势一致。图7
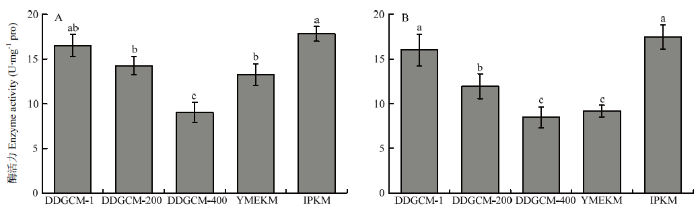 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7金针菇子实体(A)及菌丝体(B)所提线粒体中超氧化物歧化酶活性
Fig. 7Activity of Superoxide dismutase in mitochondria of F. filiformis fruiting body (A) and mycelium (B)
2.2.2 健那绿染色显微镜拍照及计数 由图8、9可见,在子实体中,DDGCM随着洗涤液稀释倍数的升高,所得活性线粒体数量逐渐减少。未稀释洗涤液组的活性线粒体数量在三者中最多,而IPKM所得活性线粒体数量整体最多,显著高于其他两组。在菌丝体中,仍然是IPKM所得活性线粒体数量最多,与其他组均具有显著差异;而YMEKM得到的活性线粒体数量最低。
图8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8健那绿染色活性线粒体
Fig. 8Janus green B staining active mitochondria
图9
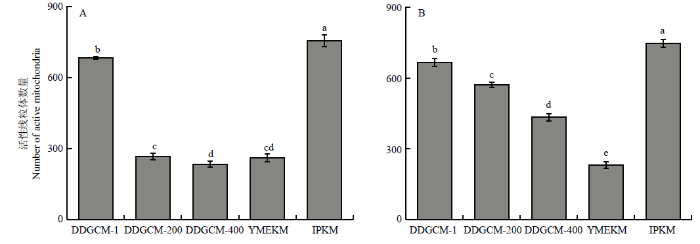 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9各提取方法的金针菇子实体(A)及菌丝体(B)中活性线粒体数量
Fig. 9The number of active mitochondria extracted from the fruit body (A) and mycelium (B) of F. filiformis by each extraction method
2.3 不同包装金针菇冷藏期间外观评估
由图10可见,与纳米包装相比,普通PE包装内表面的水分凝结现象更明显,分布更均匀。此外,在冷藏21 d后,两个包装处理组都呈现出一定程度的褐变及子实体开伞现象,而普通PE包装处理组的金针菇较纳米包装组更严重。图10
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10不同包装对金针菇冷藏期间外观质量的影响
Fig. 10Effects of different packaging materials on appearance quality of F. filiformis during refrigeration
2.4 不同包装金针菇冷藏期间能量水平和能荷值
由图11可见,在贮藏期间,纳米包装金针菇的ATP含量始终显著高于其他处理组,AMP含量始终显著低于其他处理组,表明纳米包装能有效控制ATP含量的降低,同时还抑制了AMP含量的增加。在整个贮藏过程中,随着ATP、ADP和AMP含量的变化,金针菇的能荷值也发生变化,而且在贮藏期间纳米包装组的能荷值始终显著高于普通PE包装组(P<0.05)。图11
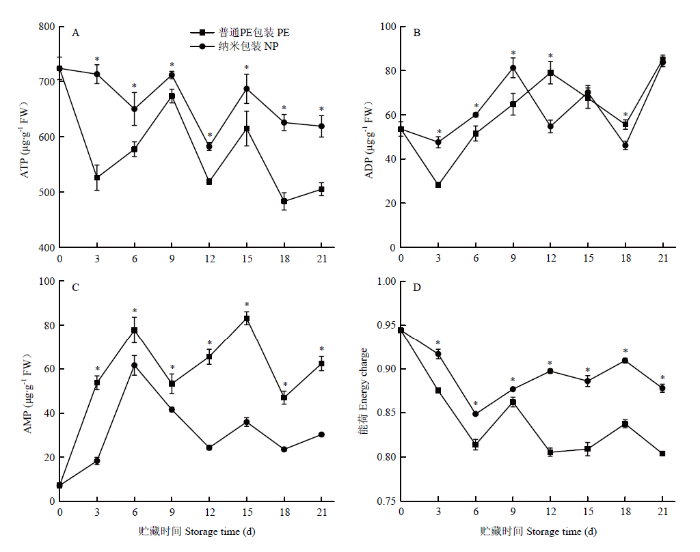 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图11不同包装对金针菇冷藏期间ATP含量(A)、ADP含量(B)、AMP含量(C)、能荷(D)的影响
*表示普通PE包装与纳米包装组差异显著(P<0.05)。下同
Fig. 11Effect of different packaging materials on changes of ATP content (A), ADP content (B), AMP content (C) and energy charges (D) of F. filiformis during refrigeration
*means significant difference between PE and NP groups (P<0.05). The same as below
2.5 不同包装金针菇冷藏期间线粒体关键复合体酶活性
由图12可见,在贮藏过程中,线粒体复合体I和复合体Ⅲ活性均呈现先升高再降低的趋势且两个包装组变化趋势一致。对于线粒体复合体Ⅳ活性,两个包装组都在0—6 d呈现先下降的趋势,普通PE包装组样品的复合体活性在贮藏第9天时达到峰值,随后逐渐下降,而纳米包装组的线粒体复合体Ⅳ活性则在贮藏第15天时才达到峰值然后下降。在整个贮藏期间,纳米包装组的3个线粒体复合体酶活性均显著高于普通PE包装组,说明纳米包装可以在贮藏期间使线粒体复合体酶维持高活性,从而维持金针菇的能量代谢水平。图12
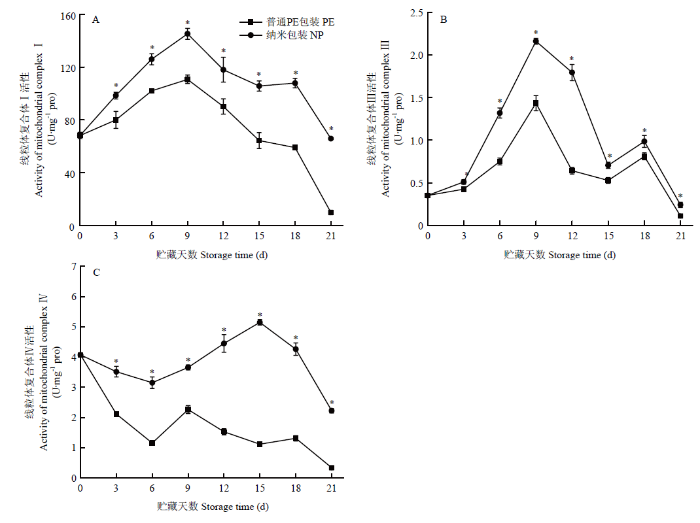 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图12不同包装对金针菇冷藏期间线粒体复合体I(A)、复合体Ⅲ(B)和复合体Ⅳ(C)活性的影响
Fig. 12Effects of different packaging materials on changes of mitochondrial complex I (A), complex Ⅲ (B) and complex Ⅳ (C) of F. filiformis during refrigeration
3 讨论
线粒体是能量代谢的主要场所,它为细胞活动提供了80%的能量,其中三羧酸循环发生于线粒体基质中,而电子传递链和氧化磷酸化的发生位点在线粒体内膜上[21]。提取高纯度的线粒体,是获得高质量线粒体蛋白的重要前提,有助于分子生物学以及蛋白质组学的进一步研究,为后续线粒体功能的研究及其蛋白质组的分析奠定了良好的基础[33]。然而在不同物种的细胞中,细胞结构有所差别,因此,针对不同物种细胞有不同的分离线粒体的方法[34,35,36]。本研究结果表明改进普利莱法所得的线粒体受胞质污染程度最低,更好地保持了线粒体的膜完整和活性。酵母线粒体提取试剂盒长时间的提取分离操作会对细胞壁、线粒体结构和活性造成损伤,而且过高或过低的缓冲液pH会引起线粒体肿胀和膜破碎[37],进而降低了线粒体活性。不连续蔗糖密度梯度离心法属于传统方法,尽管线粒体在分离过程中会从破碎的细胞中迅速分离出来,但在将线粒体分散到悬浮介质中之前,线粒体会与液泡的内容物有短暂的接触,其中一些水解酶将附着在线粒体外膜上。因此,线粒体外膜有被这些污染性的水解酶裂解的风险[38]。与此同时,多次的高速离心同样会使部分线粒体膜破损,并且高浓度蔗糖引起的渗透损伤也是造成线粒体内膜破损的重要原因[39]。相比之下,改进普利莱法能通过有效的破壁方法和较少的离心步骤得到结构较完整且活性较好的线粒体,而整个分离过程的关键点是对金针菇细胞壁的破碎及细胞膜的裂解。真菌的细胞壁主要由葡聚糖、几丁质和蛋白质组成[40],其中几丁质与植物中的纤维素类似,形成坚硬的微纤维,强化细胞壁的强度,并且与α-1,3-葡聚糖一起排列[41,42]。因此,破坏几丁质结构是破壁处理的关键。而蜗牛酶内含有多种酶,能够高效降解几丁质[43]。对于改进普利莱法,分离前用巯基化合物增强胞内蛋白稳定性的同时[44],在等渗条件下使用蜗牛酶进行酶解破壁,为得到大量完整的线粒体奠定了基础。有研究表明金针菇表面结构薄,不能有效阻止其表面水分的迅速散失[45]。而采后的金针菇子实体或包装袋内表面会因贮藏环境温差出现水汽凝结现象,该现象有利于微生物的繁殖、侵染和传播[46]。纳米包装材料可以有效控制金针菇采后蒸腾作用,减少包装内部水汽凝结,这与前期研究结果一致[13]。笔者实验室前期对冷藏期间不同包装金针菇的孢子数量检测发现,第15天无包装组和普通PE包装组孢子数量分别达到4.84×1011和4.09×1011/g cap,而纳米包装组只有3.44×1011/g cap,显著低于其他两组(P<0.05),表明纳米包装材料能有效调节包装袋内湿度和气体成分,减轻水汽凝结,控制微生物生长,减缓金针菇腐烂酸败。冷藏期间纳米包装材料可以抑制金针菇的菌柄伸长、开伞、褐变,该现象是采后金针菇后熟作用的重要表现[47]。在冷藏期间,金针菇后熟作用主要由包装内的温度、气体成分以及乙烯含量等因素所影响。有研究表明,金针菇在采后菌丝能产生高浓度的乙烯,促进其呼吸作用,从而加快其衰老速度[48]。前期对纳米包装在冷藏期间的气体成分进行了分析,结果未检测出乙烯,表明纳米包装材料中聚合的纳米颗粒可能会吸收金针菇释放的乙烯[22]。此外,纳米包装可以通过抑制金针菇的呼吸速率,减少活性氧积累,从而保护线粒体结构,维持金针菇的能量代谢水平[13]。
已有研究表明果蔬的生理失调和褐变与细胞能量的变化有关[49]。线粒体是细胞中进行生物氧化和产生ATP的主要场所。因此,果蔬组织及线粒体中主要能量物质的变化已成为细胞能量代谢研究的主要观察指标[50]。ATP是至关重要的能源,保障果蔬组织正常的生理代谢功能[51]。同时,能荷值的高低决定了线粒体磷酸化活性的高低。笔者实验室前期研究中发现纳米包装材料可以有效防止线粒体结构损伤[3],而线粒体结构受损时会导致磷酸化活性降低,进而诱导ATP和能荷低水平表达。而除了自然衰老劣变,线粒体的提取过程也会导致细胞裂解和线粒体结构损伤,这一点在线粒体提取方法筛选试验中已得到验证。这也能解释前人在贮藏试验中使用差速离心法提取线粒体时,测得的线粒体主要复合体活性指标并没有明显波动的原因[3]。电子传递链主要由线粒体复合体I(NADH脱氢酶)、线粒体复合体Ⅲ(细胞色素b/c1氧化还原酶)和线粒体复合体Ⅳ(细胞色素C氧化酶)组成,为细胞提供ATP发挥关键作用[14]。镶嵌于线粒体内膜上的线粒体复合体,它们的活性高低是表征线粒体功能变化的重要指标,同时也是三羧酸循环和氧化磷酸化等能量代谢过程中的关键酶,可以反映ATP合成情况和线粒体内膜损伤程度[52]。本研究结果表明,纳米包装组金针菇具有更高水平的ATP和能荷值,并且纳米包装延迟了能荷的下降;而ATP代谢系统各物质含量在金针菇冷藏期间均呈现波动变化,这是由于食用菌属于呼吸跃变型果蔬[53]。对于呼吸跃变型果蔬,研究人员常常把其呼吸跃变的高峰期作为水果由成熟走向衰老的转折期。有研究表明,能荷的降低也是衰老的体现[54]。因此,纳米包装材料在冷藏期间通过抑制ATP水平的下降和维持高水平的能荷值来延缓金针菇的衰老。与此同时,ATP供应不足可直接引起线粒体肿胀和嵴断裂,并进一步抑制线粒体的氧化磷酸化过程,这一现象在线粒体主要复合体活性的测定中得到了证实。在线粒体主要复合体中,线粒体复合体Ⅳ是线粒体呼吸和三羧酸循环(tricarboxylic acid cycle,TCA)中的关键酶[55],为氧化磷酸化提供能量,其活性高低直接影响到线粒体呼吸链中的电子传递。低活性的线粒体复合体Ⅳ会使ATP生成减少、能荷降低,从而导致细胞所需能量供应不足。此外,呼吸链障碍在导致氧化不完全的同时也会造成活性氧生成增多,以上均会导致细胞死亡[56]。在冷藏过程中,纳米包装显著延缓了该酶活性的下降趋势,与能荷值变化表现一致,进一步证明纳米包装材料延缓了金针菇氧化磷酸化速率的下降,保障了细胞ATP供应,维持基本能量代谢需要。对于不同包装金针菇线粒体差异蛋白表达谱以及能量代谢调控路径中关键性蛋白还不明确,后续还需利用线粒体蛋白组,从分子水平上进一步阐明纳米包装对金针菇能量和呼吸代谢的作用途径与机制。
4 结论
通过分析线粒体的结构完整性和功能活性,确定改进普利莱法最适合金针菇线粒体的提取。在此基础上,利用改进普利莱法对不同包装的金针菇线粒体进行提取,证明纳米包装通过维持高水平的ATP含量与线粒体复合体I、III活性,延缓能荷与线粒体复合体IV的下降,从而更好地保持了金针菇的能量状态并延缓了衰老。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
URLPMID:19960285 [本文引用: 2]
URLPMID:20858222 [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
URLPMID:29291867 [本文引用: 4]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:29111700 [本文引用: 1]
DOI:10.1016/j.foodchem.2017.07.118URLPMID:28946353 [本文引用: 1]

Effects of propyl gallate on metabolisms of respiration and energy of harvested 'Fuyan' longans and its relationship to pericarp browning were investigated. Compared to control longans, propyl gallate could reduce ascorbic acid oxidase (AAO) activity, lower cytochrome C oxidase (CCO) activity during early-storage and mid-storage, increase NADK activity, elevate contents of NADP and NADPH, decrease contents of NAD and NADH, in addition, lower the decreases of ATP content and energy charge (E.C.), increase activities of mitochondrial H(+)-ATPase, Ca(2+)-ATPase and Mg(2+)-ATPase during early-storage and mid-storage. Above results suggested that propyl gallate-retarded browning development in pericarp of harvested longans was resulted from decreases in activities of respiratory terminal oxidases like CCO and AAO, increase in proportion of pentose phosphate pathway (PPP) to Embden-Meyerhof pathway (EMP) and tricarboxylic acid (TCA) cycle, and maintenance of mitochondrial integrity via retaining higher levels of ATP content and energy charge, as well as higher activities of mitochondrial ATPase.
[本文引用: 1]
DOI:10.1016/j.foodchem.2017.02.045URLPMID:28317755 [本文引用: 2]
As compared with P. longanae-inoculated longans, DNP treatment for P. longanae-inoculated longans exhibited higher fruit disease index and pericarp browning index, lower ATP amount and energy charge level, lower activities of SOD, CAT and APX, lower amounts of AsA and GSH, lower levels of DPPH radical scavenging activity and reducing power, higher O2(-) generating rate and MDA amount. However, supply of ATP for P. longanae-inoculated longans showed the contrary effects. These results gave convincing evidence that DNP treatment for accelerating pericarp browning and disease development of harvested longans caused by P. longanae was due to decreases of energy production and ROS scavenging capacity, and increases of O2(-) accumulation and membrane lipid peroxidation. Whereas, supply of ATP for retarding pericarp browning and disease development of harvested longans caused by P. longanae was due to increases of energy production and ROS scavenging capacity, and reductions of O2(-) accumulation and membrane lipid peroxidation.
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1038/sj.onc.1210074URLPMID:17072346 [本文引用: 1]
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) is a pleiotropic enzyme that is overexpressed in apoptosis and in several human chronic pathologies. Here, we report that the protein accumulates in mitochondria during apoptosis, and induces the pro-apoptotic mitochondrial membrane permeabilization, a decisive event of the intrinsic pathway of apoptosis. GAPDH was localized by immunogold labeling and identified by matrix-assisted laser desorption/ionization-time of flight and nano liquid chromatography mass spectroscopy/mass spectroscopy in the mitochondrion of various tissues and origins. In isolated mitochondria, GAPDH can be imported and interact with the voltage-dependent anion channel (VDAC1), but not the adenine nucleotide translocase (ANT). The protein mediates a cyclosporin A-inhibitable permeability transition, characterized by a loss of the inner transmembrane potential, matrix swelling, permeabilization of the inner mitochondrial membrane and the release of two pro-apoptotic proteins, cytochrome c and apoptosis-inducing factor (AIF). This novel function of GAPDH might have implications for the understanding of mitochondrial biology, oncogenesis and apoptosis.
[本文引用: 1]
[本文引用: 1]
URLPMID:30409664 [本文引用: 1]
URLPMID:28870773 [本文引用: 1]
URL [本文引用: 1]
The changes of breath rares during the mycelial growth of three basidiomycetes(Ganoderma lucidum ,Hericium erinaceus,Flammulina velutipes)at the mycelial stage were studied.The results show that the mycelial breath rates of three basidiomycetes a11 increase with mycelial growth,but the value of each species is different.The mycelial breath rate is stable during the same growth period. Each curve of mycelial mean growth rate of the three basidiomycetes shows only one peak.The maximum of breath rate comes later than that of the mycelial growth rate.
URL [本文引用: 1]
The changes of breath rares during the mycelial growth of three basidiomycetes(Ganoderma lucidum ,Hericium erinaceus,Flammulina velutipes)at the mycelial stage were studied.The results show that the mycelial breath rates of three basidiomycetes a11 increase with mycelial growth,but the value of each species is different.The mycelial breath rate is stable during the same growth period. Each curve of mycelial mean growth rate of the three basidiomycetes shows only one peak.The maximum of breath rate comes later than that of the mycelial growth rate.
URL [本文引用: 1]
超氧化物歧化酶广泛存在于动物、植物、微生物体内,它能够专一性地清除生物氧化过程中产生的超氧化物自由基,是生物抗氧化系统的重要酶类之一.本文从超氧化物歧化酶的种类、作用机理、活性调控、超氧化物歧化酶基因工程以及分子进化五个方面综述超氧化物歧化酶研究的进展.
URL [本文引用: 1]
超氧化物歧化酶广泛存在于动物、植物、微生物体内,它能够专一性地清除生物氧化过程中产生的超氧化物自由基,是生物抗氧化系统的重要酶类之一.本文从超氧化物歧化酶的种类、作用机理、活性调控、超氧化物歧化酶基因工程以及分子进化五个方面综述超氧化物歧化酶研究的进展.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:6289753 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/jfds.2003.68.issue-5URL [本文引用: 1]
DOI:10.1099/00221287-92-2-251URL [本文引用: 1]
[本文引用: 1]
DOI:10.1016/S0008-6215(99)00154-8URL [本文引用: 1]
DOI:10.1016/j.foodchem.2017.06.024URLPMID:28873622 [本文引用: 1]
The blueberry fruit has a light-blue appearance because its blue-black skin is covered with a waxy bloom. This layer is easily damaged or removed during fruit harvesting and postharvest handling. We investigated the effects of wax removal on the postharvest quality of blueberry fruit and their possible mechanisms. The removal of natural wax on the fruit was found to accelerate the postharvest water loss and decay, reduce the sensory and nutritional qualities, and shorten the shelf-life. Wax removal decreased the activities of antioxidant enzymes and contents of antioxidants, and accelerated accumulation of ROS and lipid peroxidation, especially at the later period of storage. Moreover, the organellar membrane structure was disrupted in fruit with wax removed. These results indicate that cuticular wax plays an important role in maintaining the postharvest quality and delaying fruit senescence. The results should improve our understanding for better preservation of postharvest quality of blueberry fruit.
DOI:10.1111/j.1750-3841.2010.01520.xURLPMID:20492272 [本文引用: 1]
UNLABELLED: A novel nano-packing material with lower relative humidity, oxygen transmission rate and high longitudinal strength was synthesized by blending polyethylene with nano-powder (nano-Ag, kaolin, anatase TiO(2), rutile TiO(2)), and its effect on preservation quality of strawberry fruits (Fragaria ananassa Duch. cv Fengxiang) was investigated during storage at 4 degrees C. Results showed that nano-packaging was able to maintain the sensory, physicochemical, and physiological quality of strawberry fruits at a higher level compared with the normal packing (polyethylene bags). After a 12-d storage, decreases in the contents of total soluble solids, titratable acidity, and ascorbic acid of nano-packing were significantly inhibited. Meanwhile, decay rate, anthocyanin, and malondialdehyde contents were decreased to 16.7%, 26.3 mg/100g, 66.3 micromol/g for nano-packing and 26.8%, 31.9 mg/100g, 75.4 micromol/g for normal packing; polyphenoloxidase (PPO) and pyrogallol peroxidase (POD) activities were significantly lower in nano-packing than the control. These data indicated that the nano-packaging might provide an attractive alternative to improve preservation quality of the strawberry fruits during extended storage. PRACTICAL APPLICATION: Nano-packing exhibited identified quality benefits applicable to the preservation of fresh strawberry. Furthermore, nano-packing has the advantages of simple processing and feasibility to be industrialized in contrast with other storages. Thus, the utilization of nano-packing will likely assist commercial producers and retailers in extending the shelf life of products over a broader range in the future.
[D].
[本文引用: 1]
[D].
[本文引用: 1]
URLPMID:811760 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.foodchem.2016.01.142URLPMID:26920276 [本文引用: 1]
The effects of oxalic acid on the development of chilling injury (CI), energy metabolism and lycopene metabolism in tomato fruit (Solanum lycopersicum L.) were investigated. Mature green tomatoes were dipped in 10mmoll(-1) oxalic acid (OA) solution for 10min at 25 degrees C. Tomatoes were subsequently stored at 4+/-0.5 degrees C for 20days before being transferred to 25 degrees C for 12days. Oxalic acid treatment apparently alleviated CI development and membrane damage; maintained higher levels of ATP and ADP; increased activities of succinic dehydrogenase (SDH), Ca(2+)-adenosine triphosphatase (Ca(2+)-ATPase) and H(+)-adenosine triphosphatase (H(+)-ATPase); and elevated lycopene accumulation associated with the upregulation of PSY1 and ZDS expression in tomatoes during a period at room temperature following exposure to chilling stress. Thus, oxalic acid treatment benefited the control of CI and the maintenance of fruit quality in tomatoes stored for long periods (approximately 32days).
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.1186/1471-2229-13-55URL [本文引用: 1]
DOI:10.1002/jsfa.5973URLPMID:23208649 [本文引用: 1]
BACKGROUND: Energy status is associated with ripening, senescence and physiological disorder of various types of fruit after harvest, including peaches. Methyl jasmonate (MeJA) was effective in enhancing chilling tolerance in cold-stored peach fruit. The effect of MeJA on energy metabolism in peach fruit was investigated. RESULTS: MeJA treatment increased the contents of adenosine triphosphate and adenosine diphosphate, but decreased the content of adenosine monophosphate, which resulted in a higher level of energy charge in peach fruit. Enzyme activities including H+-adenosine triphosphatase, Ca2+-adenosine triphosphatase, succinic dehydrogenase and cytochrome c oxidase were also higher in MeJA-treated fruit than those in control fruit. Meanwhile, MeJA treatment markedly inhibited the increase of ion leakage and the accumulation of malondialdehyde, super-oxidant radical and hydrogen peroxide. CONCLUSION: These results indicated that MeJA could enhance chilling tolerance of peach fruit by inducing enzyme activities related to energy metabolism and maintaining high levels of ATP and energy charge.
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}