Molecular Ecological Network Analyses Revealing the Effects of Nitrogen Application on Soil Microbial Community in the Degraded Grasslands
ZHU RuiFen1, LIU JieLin1, WANG JianLi1, HAN WeiBo1, SHEN ZhongBao1, XIN XiaoPing,21Institute of Pratacultural Science, Heilongjiang Academy of Agricultural Sciences, Harbin 150086 2Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing 100081
Abstract 【Objective】 Nitrogen input affects the sustainability of the global grassland ecosystem. Paying attention to the soil microbial community and its molecular ecological network can provide a theoretical basis for grassland degradation restoration. 【Method】 Taking Songnen degraded Leymus chinensis grassland as the research object, the molecular ecological network of soil microbial community was constructed by applying high-throughput sequencing and random matrix network construction with and without nitrogen treatment. To explore the effects of nitrogen management on the soil microbial community structure and network in degraded Leymus chinensis grassland, the key microbial changes in the microbial network structure under the condition of nitrogen addition was studied, and the interaction between microorganisms during the process were investigated, and the conditions for external nitrogen addition key points and regularity of soil bacterial dynamic change were analyzed. 【Result】 At the level of phylum classification, there were 22 bacterial phylum in nitrogen-applied grassland and 23 without nitrogen. The 7 phylum were the dominant phylum of the nitrogen-applied and non-nitrogen-applied grasslands. Among them, Proteobacteria was the phyla containing the largest number of OTUs, accounting for about 30.46% of the total sequence. The next largest genus was about 30.15% of the total sequence. The genus Gemmatimonadetes was the third genus containing OTUs, accounting for 8.14% of the total sequence. Actinomycete accounted for about the total 6.15% of the sequence, while Chloroflexi, Bacteroidetes and Nitrospirae accounted for 17.16% of the total sequence. The relative abundances of Proteobacteria, Actinomycota and Bacteroides in soil microorganisms in nitrogen-applied grassland were significantly higher than those in non-nitrogen applied grassland soil; The relative abundances of Proteobacteria, Actinomycete and Bacteroidetes were significantly higher than the soils of nitrogen-applied grassland (P<0.01), and no significant difference was found between the nitrogen application and non-nitrogen treatment of other bacteria. The forward connection ratio, the average path length, the average clustering coefficient, and the modularity of the characterizing network were all significantly lower than the nitrogen-free treatment (P<0.001). In the molecular ecological network of soil, there were 16 modular hubs without nitrogen treatment (Zi>2.5, Pi≤0.62), and there were 6 modular hubs under nitrogen treatment, all of which belong to Acidobacteria, Gemmatimonadetes and Actinomycete. Nitrogen application led to changes in soil microbial species relationships, which in turn changed the overall soil ecological network. 【Conclusion】 Nitrogen application reduced the complexity and tightness of soil network structure of degraded grassland, and reduced the relative abundance of Acidobacteria and Chloroflexi in degraded grassland, while which improved the relative abundance of Proteobacteria, Actinomycete and Gemmatimonadetes. The number of microbial key species (OTU) in soil decreased from 16 (no nitrogen application) to 6 and there was no overlapping OTU in both soils, indicating that nitrogen application regulated key species of its community network and thus changes its molecular ecological network. Keywords:nitrogen application;degraded grassland;soil microbe;molecular ecology network
PDF (3307KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 朱瑞芬, 刘杰淋, 王建丽, 韩微波, 申忠宝, 辛晓平. 基于分子生态学网络分析松嫩退化草地土壤微生物群落对施氮的响应[J]. 中国农业科学, 2020, 53(13): 2637-2646 doi:10.3864/j.issn.0578-1752.2020.13.012 ZHU RuiFen, LIU JieLin, WANG JianLi, HAN WeiBo, SHEN ZhongBao, XIN XiaoPing. Molecular Ecological Network Analyses Revealing the Effects of Nitrogen Application on Soil Microbial Community in the Degraded Grasslands[J]. Scientia Acricultura Sinica, 2020, 53(13): 2637-2646 doi:10.3864/j.issn.0578-1752.2020.13.012
采集的土壤经过去除植物残体和石块等杂质均分后,一份用于土壤高通量测序,另一份用于土壤化学性质测定[21],包括全氮(soil total nitrogen content,STN)、全磷(soil total phosphorus content,STP)、速效氮(soil available nitrogen,SAN)、速效磷(soil available phosphorus,SAP)含量。STN、STP用H2SO4-K2SO4-Cu2SO4催化法消解,消煮后的样品经过滤、稀释、定容后用AutoAnalyzer3(AA3)连续流动分析仪测定。土壤速效氮含量是指硝态氮(soil nitrate nitrogen,SNO3)和铵态氮(soil ammonium nitrogen,SNH4)的总和,因此对SNO3和SNH4进行分别测定,土壤经过2 mol·L-1 KCl浸提、过滤、定容后,用AA3连续流动分析仪测定。SAP采用硫酸钼梯抗比色法测定。土壤样品的理化性质见表1。土壤高通量测序由百迈客公司完成,通过统计数据处理各阶段样品序列数目,评估数据质量。
Table 1 表1 表1施氮和未施氮处理草地土壤的化学性质 Table 1Chemical characteristic of no fertilized (NF) and fertilized (F) soils of grassland
处理 Treatment
pH
土壤全氮 STN (g·kg-1)
土壤全磷 STP (g·kg-1)
土壤铵态氮 SNH4 (mg·kg-1)
土壤硝态氮 SNO3 (mg·kg-1)
土壤速效磷 SAP (mg·kg-1)
F
7.95a
1.71a
0.53b
17.80a
21.62a
10.45a
NF
8.09a
1.31b
0.60b
12.61b
14.75b
10.49a
不同小写字母表示施氮土壤和未施氮处理在P<0.05水平上差异显著。下同 Different small letters represent significant difference between nitrogenous and non-nitrogenous treatments at P<0.05. The same as below
Table 2 表2 表2各样品测序数据评估结果 Table 2Results of sample sequencing data evaluation
样品名称Sample ID
双端序列 PE reads
原始序列 Raw tags
优化序列 Clean tags
有效序列 Effective tags
平均序列长度 Average length (bp)
GC (%)
Q20 (%)
Q30 (%)
Effective (%)
NF
47023
44654
39465
38754
421
57.14
97.15
94.39
82.41
F
66659
63320
56208
55106
420
56.61
97.26
94.61
82.67
GC为样品G和C类型的碱基占总碱基的百分比;Q20为质量值≥20的碱基占总碱基数的百分比;Q30为质量值≥30的碱基占总碱基数的百分比;Effective为有效序列占双端序列的百分比 GC is the percentage of bases of type G and C in the total bases; Q20 is the percentage of bases with a mass value greater than or equal to 20 in the total bases; Q30 is the mass value bases greater than or equal to 30 accounting for the percentage of total bases; Effective is the percentage of effective tags in double-ended sequences (PE reads)
Table 4 表4 表4施氮和未施氮退化草地土壤微生物群落的分子生态网络拓扑性质 Table 4Topological properties of the empirical molecular ecological networks (MENs) of microbial communities in the no fertilized (NF) and fertilized (F) soils
处理Treatment
阈值 Threshold
节点数 Total node
连接数 Total links
正向连接比 Positive link percentage (%)
平均连通度 Connectivity
平均路径长度 Average path distance
平均聚集系数 Clustering coefficient
模块性 Modularity
F
0.9
1872
19873
53.2*
23.6*
2.33*
0.435*
0.46*
NF
0.9
1913
20389
69.4*
28.7*
2.89*
0.462*
0.41*
*表示P<0.001,代表施氮土壤和未施氮土壤的显著性差异 *Represents a significant difference between nitrogenous and non-nitrogenous soils at P<0.001
圆圈代表物种,圆圈大小代表丰度;线条代表两物种间相关,线的粗细代表相关性的强弱。橙色代表正相关,绿色代表负相关。下同 Fig. 2Network of interactions of the main microbial genus of nitrogen-applied soil
The circle represents species, circle size represents abundance; The lines represent the correlation between the two species, the thickness of the lines represents the strength of the correlation. Orange represents positive correlation, and green represents negative correlation. The same as below
ZHANG NL, GUO JX, WANG XY, MA KP. Soil microbial feedbacks to climate warming and atmospheric N deposition Chinese Journal of Plant Ecology, 2007,31(2):252-261. (in Chinese) [本文引用: 2]
WENDU RL, LIG, YANG DL, ZHANG JN, YIJ. nifH gene diversity and community structure of soil nitrogen_fixing bacteria in Hulunbeier grassland, Inner Mongolia Chinese Journal of Ecology, 2007,30(4):790-797. (in Chinese) [本文引用: 1]
FAUSTK, RAESJ. Microbial interactions: From networks to models Nature Reviews Microbiology, 2012,10:538-550. [本文引用: 1]
ZHENGY, LI YY, DING CY, REN XM, CHEN ZJ. Effects of bioenergy cropping on rhizosphere bacteria networks structure in Cd contaminated soil Acta Scientiae Circumstantiae, 2016,36(7):2605-2612. (in Chinese) [本文引用: 1]
SCHLOTERM, NANNIPIERIP, SORENSEN SJ. Microbial indicators for soil quality Biology and Fertility of Soils, 2018,54:1-10. [本文引用: 1]
PACE NR. A molecular view of microbial diversity and the biosphere Science, 1997,276(5313):734-740. [本文引用: 2]
CAPORASO JG, LAUBER CL, WALTERS WA, BERG-LYONSD, HUNTLEYJ, FIERERN, OWENS SM, BETLEYJ, FRASERL, BAUERM, GORMLEYN, GILBERT JA, SMITHG, KNIGHTR. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms ISME Journal, 2012,6(8):1621-1624. [本文引用: 2]
MARTINY J BH, EISEN JA, PENNK, ALLISONSD, HORNER- DEVINE MC. Drivers of bacterial beta-diversity depend on spatial scale Proceedings of the National Academy of Sciences of the USA, 2011,108(19):7850-7854. [本文引用: 1]
LIU YY, WANGS, LI SZ, DENGY. Advances in molecular ecology on microbial functional genes of carbon cycle Microbiology China, 2017,44(7):1676-1689. (in Chinese) [本文引用: 1]
WANG ZH, WANGS, LIU YY, FENGK, DENGY. The applications of metagenomics in the detection of environmental microbes involving in nitrogen cycle Biotechnology Bulletin, 2018,34(1):1-14. (in Chinese) [本文引用: 1]
ALLISON SD, MARTINY J BH. Resistance, resilience, and redundancy in microbial communities Proceedings of the National Academy of Sciences of the USA, 2008,105(1):11512-11519. [本文引用: 1]
WALLENSTEIN MD, MC NULTYS, FERNANDEZ IJ. Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments Forest Ecology and Management, 2006,222(1/3):459-468. [本文引用: 2]
LUPWAYI NZ, CLAYTON GW, O'DONOVAN JT. Soil microbial response to nitrogen rate and placement and barley seeding rate under no till Agronomy Journal, 2011,103(4):1064-1071. [本文引用: 2]
GEISSELERD, LAZICKI PA, SCOW KM. Mineral nitrogen input decreases microbial biomass in soils under grasslands but not annual crops Applied Soil Ecology, 2016,106:1-10. [本文引用: 2]
PACE NR. A molecular view of microbial diversity and the biosphere Science, 1997,276(5313):734-740. [本文引用: 1]
RINKEC, SCHWIENTEKP, SCZYRBAA. Insights into the phylogeny and coding potential of microbial dark matter Nature, 2013,499(7459):431-437. [本文引用: 1]
PEURAS, BERTILSSONS, JONES RI. Resistant microbial co-occurrence patterns inferred by network topology Applied and Environmental Microbiology, 2015,81(6):2090. [本文引用: 1]
ZHANGB, HORVATHS. A general framework for weighted gene co-expression network analysis Statiscal Applications in Genetics and Molecular Biology, 2005,4(1):1-43. [本文引用: 1]
MUEGGE BD, KUCZYNSKIJ, KNIGHTSD. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans Science, 2011,332(6032):970-974. [本文引用: 1]
CHEN JS, ZHU RF, ZHANGQ, SHAN LW, WANG YX. Effects of clipping frequency and nitrogen fertilizer on the stoichiometric characteristics of N, P for soil and plant in Leymus chinensis meadow Chinese Journal of Grassland, 2019,41(1):25-30. (in Chinese) [本文引用: 4]
HAO YQ, XIEL, CHEN YM. Effects of nitrogen deposition on diversity and composition of soil bacterial community in a subtropical Cunninghamia lanceolata plantation Chinese Journal of Applied Ecology, 2018,29(1):53-58. (in Chinese) [本文引用: 2]
LINGN, CHEN DM, GUOH. Differential responses of soil bacterial communities to long-term N and P inputs in a semi-arid steppe Geoderma, 2017,292:25-33. [本文引用: 2]
YANGF, WU JW, ZHANG DD. Soil bacterial community composition and diversity in relation to edaphic properties and plant traits in grasslands of southern China Applied Soil Ecology, 2018,128:43-53. [本文引用: 1]
LIU HM. Effects of nitrogen deposition on soil carbon and nitrogen transformation and microbial characteristics in Stipa baicalensis steppe [D]. Beijing: Chinese Academy of Agricultural Sciences, 2019. (in Chinese) [本文引用: 1]
LUOF, YANGY, ZHONGJ. Constructing gene co-expression networks and predicting functions of unknown genes by random matrix theory BMC Bioinformatics, 2007,8(1):1-17. [本文引用: 1]
SUNX, GAOY, YANG YF. Recent advancement in environmental research with metagenomics tools Biodiversity Science, 2013,21(3):393-400. [本文引用: 1]
CORNWELL WK, SCHWILK L DW, ACKERLY DD. A trait-based test for habitat filtering: convex hull volume Ecology, 2006,87(6):1465-1471. [本文引用: 1]
DENGY, HE ZL, XIONG JB, YUH, XU MY, HOBBIE SE, REICH PB, SCHADT CW, KENTA, PENDALLE, WALLENSTEINM, ZHOU JZ. Elevated carbon dioxide accelerates the spatial turnover of soil microbial communities Global Change Biology, 2016,22(2):957-964. [本文引用: 1]
 ,2
,2
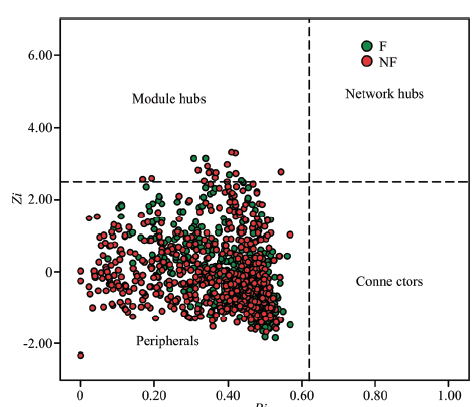 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT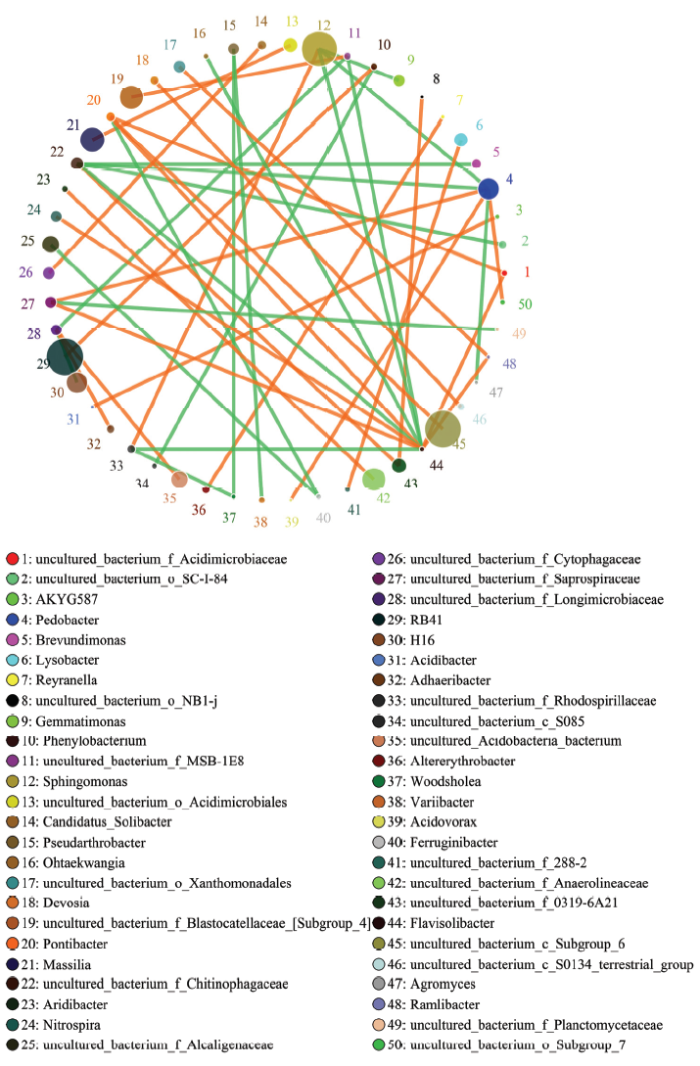 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT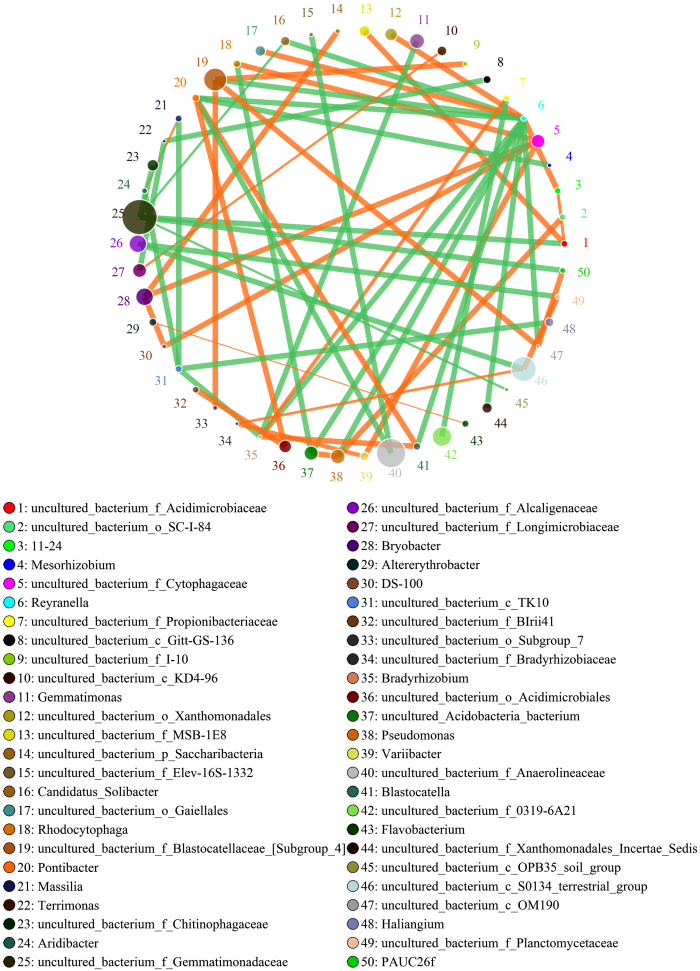 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}