Genome-Wide Identification and Characterization of Transposable Elements in Xichuan Black-Bone Chicken
LI DongHua, FU YaWei, ZHANG ChenXi, CAO YanFang, LI WenTing, LI ZhuanJian, KANG XiangTao,, SUN GuiRong,College of Animal Science and Veterinary Medicine, Henan Agricultural University, Zhengzhou 450046
Abstract 【Objective】The aim of the study was to analyze the whole genome re-sequencing data of Xichuan black-bone chicken to obtain the identification, classification, distribution of the transposon of Xichuan black-bone chicken, and to explore the pathways involved in transposon-related genes, which not only had important significance for studying the biological function of the Xichuan black-bone chicken transposon elements (TEs), but also provided important basic data for exploring genome amplification, genome function and evolution research. 【Method】In this study, whole genome resequencing of blood DNA of Xichuan black-bone chicken was performed, and the paired-end mapping methods alignment genome was used. The TEs was identified, constructed, corrected and classified by RepeaterModeler, TEclass, RepeatMasker and other processes. All TEs in the whole genome of Xichuan black-bone chicken were obtained to analyze the characteristics, distribution and relationship of their genes. All genes inserted in the TEs were subjected to GO and KEGG databases enrichment, and the functions were described in combination with GO and KEGG annotation results, respectively. 【Result】 After identification, classification and annotation, 370 252 TEs sequences were identified in Xichuan black-bone chicken, which were divided into 19 superfamilies, mainly CR1, TcMar-Mariner, ERVL, ERV1 and other superfamilies, further indicated that the TEs types of Xichuan black-bone chicken were major TEs. The number of TEs was related to chromosome length, and longer the chromosome was, the more the number of TEs was. Number of TEs was inversely proportional to the density of gene. TEs density was relatively low in gene dense regions. The insertion of TEs in genome was not random, LTR/ERVL, LTR/ERV1, DNA/PIF-Harbinger, DNA/Hat-Charlie, RC/Helitron tend to be inserted outside the gene. GO enrichment analysis indicated some of these genes related to TEs were enriched in the following biological process terms: cell process, single-organism process, metabolic process, biological regulation, response to stimulus. In addition, some of these genes related to TEs were enriched in the following molecular function terms: binding and catalytic activity. Furthermore, some of these genes related to TEs were enriched in the following cell component terms: cell parts, organelles, and membranes. KEGG enrichment analysis indicated that these genes related to TEs mainly focused on Glycerolipid metabolism, PPAR signaling pathway, PI3K-Akt signaling pathway, Insulin resistance, and Jak-STAT signaling pathway. This study mainly focused on the pathways related to the characteristics of Xichuan black-bone chicken, such as tyrosine metabolism related to pigmentation, lipid metabolism related to meat quality, fatty acid synthesis, and PI3K-Akt signaling pathway. 【Conclusion】 There was a positive correlation between TEs content and genome size of Xichuan black-bone chicken. Moreover, the TEs of Xichuan black-bone chicken had a certain preference in the distribution of genome. TEs related genes were enriched in the pigmentation related pathway, which might be related to the germplasm characteristics of Xichuan black-bone chicken. The specific regulatory mechanisms remained to be further studied. Keywords:chicken;whole genome resequencing;transposon elements;characterization analysis
PDF (1654KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 李东华, 付亚伟, 张晨曦, 曹艳芳, 李文婷, 李转见, 康相涛, 孙桂荣. 淅川乌骨鸡全基因组转座子的鉴定与分析[J]. 中国农业科学, 2020, 53(7): 1491-1500 doi:10.3864/j.issn.0578-1752.2020.07.017 LI DongHua, FU YaWei, ZHANG ChenXi, CAO YanFang, LI WenTing, LI ZhuanJian, KANG XiangTao, SUN GuiRong. Genome-Wide Identification and Characterization of Transposable Elements in Xichuan Black-Bone Chicken[J]. Scientia Acricultura Sinica, 2020, 53(7): 1491-1500 doi:10.3864/j.issn.0578-1752.2020.07.017
开放科学(资源服务)标识码(OSID):
0 引言
【研究意义】转座子(简称TEs),又称易位子,跳跃基因,指存在于基因组上可以自主复制和位移的一段DNA片段[1],构成了高等生物基因组的大部分并且其在不同生物基因组中的含量有非常大的变化。利用全基因组重测序筛选淅川乌骨鸡有关的转座子,从而发现与淅川乌骨鸡种质特性有关的转座子。【前人研究进展】转座子主要分为Class I 型转座子和Class Ⅱ型转座子两大类[2]。Class I 型转座子,又称逆转录转座子。以RNA为介导进行转座,属于“DNA-RNA- DNA”型,每转座一次相应拷贝数增加,基因组增大;且本身带有调控元件,所以会对宿主基因的表达造成影响。主要包括长末端重复元件(LTRs),内源性反转录病毒(ERV),长散布元件(LINEs)和短散布元件(SINEs),后两者又被统称为非长末端重复元件(Non-LTRs)。Class Ⅱ 型转座子,也称作DNA转座子。以DNA为介导进行转座,属“DNA-DNA”型,造成基因的突变或者重排,一般对基因组大小没有影响。此类转座子包括微型反向重复转座元件(MITEs)和滚筒式转座子(helitron)[3]。 转座子在20世纪50年代被McClintock在玉米中首次发现[4],在很长一段时间,一直被当做是基因组中的一类无用的“垃圾序列”[5]。然而,近年来,随着越来越多物种全基因组测序的应用,关于染色体结构[6]、基因组大小[7]、基因组重排[8]、新基因生成[9]和基因表达调控[10]等受转座子影响的研究报道越来越多,在分子生物学研究中被广泛应用于基因治疗、转基因动物制备和物种进化等研究[11,12,13]。目前,转座子插入对畜禽的影响也已被广泛报道,CLARK等[14]研究发现在狗silver基因SINE插入进而影响毛色表型。DALL'OLIO等[15]研究表明在赛马MSTN基因转座子的插入,影响马的赛跑距离。猪VRTN基因转座子的插入影响猪脊椎数发育[16]。【本研究切入点】淅川乌骨鸡是2011年列入的《中国畜禽遗传资源志-家禽志》中的一个新的遗传资源。具有“乌嘴、乌腿、乌皮、乌骨、乌肉、绿壳蛋”等特征;遗传性能稳定、觅食力强、适应性广、喜群居、耐寒性强、饲料消耗少、具有药用保健功能等优点。近年来,随着测序技术的发展,越来越多的生物基因组序列被测定,为从基因组水平进行转座子的鉴定、分类、注释等提供了便利,转座子几乎存在于所有生物体基因组中,并在真核生物基因组中占据重要组成部分,尤其是脊椎动物[17,18,19]。然而关于鸡转座子方面的研究还鲜有报道。【拟解决的关键问题】本研究利用淅川乌骨鸡全基因组重测序数据,从全基因组范围内进行转座子鉴定、分类和特征分析,为淅川乌骨鸡基因组功能和进化研究等提供重要基础数据。
OKADAN . Transfer RNA-like structure of the human Alu family: implications of its generation mechanism and possible functions Journal of Molecular Evolution, 1990,31(6):500-510. [本文引用: 1]
FINNEGAN DJ . Eukaryotic transposable elements and genome evolution Trends in Genetics, 1989,5(4):103-107. [本文引用: 1]
LISCHD . How important are transposons for plant evolution? Nature Reviews Genetics, 2013,14(1):49. [本文引用: 1]
MCCLINTOCKB . The origin and behavior of mutable loci in maize Proceedings of the National Academy of Sciences of the United States of America, 1950,36(6):344. [本文引用: 1]
DOOLITTLE WF, SAPIENZAC . Selfish genes, the phenotype paradigm and genome evolution Nature, 1980,284(5757):601-603. [本文引用: 1]
HOSKINS RA, CARLSON JW, KENNEDYC, ACEVEDOD, EVANSHOLMM, FRISEE, WAN KH, PARKS, MENDEZLAGOM, ROSSIF . Sequence finishing and mapping of Drosophila melanogaster heterochromatin Science,2007,316(5831):1625-1628. [本文引用: 1]
NENEV, WORTMAN JR, LAWSOND, HAASB, KODIRAC, TU ZJ, LOFTUSB, XIZ, MEGYK, GRABHERRM . Genome sequence of Aedes aegypti, a major arbovirus vector Science,2007,316(5832):1718-1723. [本文引用: 1]
GALLAGHER LA, SHENDUREJ, MANOILC . Genome-scale identification of resistance functions in Pseudomonas aeruginosa using Tn-seq mBio,2011,2(1):00315-00310. [本文引用: 1]
REARDENA, MAGNETA, KUDOS, FUKUDAM . Glycophorin B and glycophorin E genes arose from the glycophorin A ancestral gene via two duplications during primate evolution Journal of Biological Chemistry.1993,268(3):2260-2267. [本文引用: 1]
MANDAOKARA, KUMAR VD, AMWAYM, BROWSEJ . Microarray and differential display identify genes involved in jasmonate-dependent anther development Plant Molecular Biology,2003,52(4):775. [本文引用: 1]
MACIAA, BLANCO-JIMENEZE, GARC A-P REZ J L. Retrotransposons in pluripotent cells: Impact and new roles in cellular plasticity Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms,2015,1849(4):417-426. [本文引用: 1]
SLOTKIN RK, MARTIENSSENR . Transposable elements and the epigenetic regulation of the genome Nature Reviews Genetics, 2007,8(4):272-285. [本文引用: 1]
TIANZ, ZHAOM, SHEM, DUJ, CANNON SB, LIUX, XUX, QIX, LI MW, LAM HM . Genome-wide characterization of nonreference transposons reveals evolutionary propensities of transposons in soybean Plant Cell,2012,24(11):4422-4436. [本文引用: 1]
CLARK LA, WAHL JM, REES CA, MURPHY KE . Retrotransposon insertion in SILV is responsible for merle patterning of the domestic dog Proceedings of the National Academy of Sciences of the United States of America,2006; 103(5):1376-1381. [本文引用: 1]
STEFANIADO, EMILIOS, LUCAF, MARCOT . Analysis of the 227 bp short interspersed nuclear element (SINE) insertion of the promoter of the myostatin (MSTN) gene in different horse breeds Veterinaria Italiana, 2014; 50(3):193-197. [本文引用: 1]
MIKAWAS, SATOS, NIIM, MOROZUMIT, GOUY, IMAEDAN, YAMAGUCHIT, HAYASHIT, AWATAT . Identification of a second gene associated with variation in vertebral number in domestic pigs BMC Genetics,12,1(2011-01-14). 2011,12(1):5-5. [本文引用: 1]
IVICSZ, GARRELSW, MáTéS L, YAU T Y, BASHIR S, ZIDEK V, LANDA V, GEURTS A, PRAVENEC M, RüLICKE T, KUES W A, IZSVáK Z. Germline transgenesis in pigs by cytoplasmic microinjection of Sleeping Beauty transposons Nature Protocols, 2014, 9(4):810-827. [本文引用: 1]
BAILLIE JK, BARNETT MW, UPTON KR, GERHARDT DJ, RICHMOND TA, DE SF, BRENNAN PM, RIZZUP, SMITHS, FELLM . Somatic retrotransposition alters the genetic landscape of the human brain Nature, 2011, 479(7374):534-537. [本文引用: 1]
GOODIER JL . Retrotransposition in tumors and brains Mobile DNA,2014,5(1):11-17. [本文引用: 1]
WICKERT, SABOTF, HUAVANA, BENNETZEN JL, CAPYP, CHALHOUBB, FLAVELLA, LEROYP, MORGANTEM, PANAUDO . A unified classification system for eukaryotic transposable elements Nature Reviews Genetics, 2007(8):973-982. [本文引用: 1]
CONESAA, GTZS, GARC AG, MEZ JM, TEROLJ, TAL NM, ROBLESM . Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research Bioinformatics,2005,21(18):3674-3676. [本文引用: 1]
ASAFL, SCHRAGAS, GILA . Large-scale discovery of insertion hotspots and preferential integration sites of human transposed elements Nucleic Acids Research, 2009,38(5):1515-1530. [本文引用: 1]
LINHEIRO RS, BERGMAN CM . Whole genome resequencing reveals natural target site preferences of transposable elements in Drosophila melanogaster PLoS ONE, 2012,7(2):e30008. [本文引用: 1]
INITIATIVEAG . Analysis of the genome sequence of the flowering plant Arabidopsis thaliana Nature, 2000,408(6814):796-815. [本文引用: 1]
SCHNABLE PS, WARED, FULTON RS, STEIN JC, WEIF, PASTERNAKS, LIANGC, ZHANGJ, FULTONL, GRAVES TA . The B73 maize genome: complexity, diversity, and dynamics Science, 2009,326(5956):1112-1115. [本文引用: 1]
MAB, XINY, KUANGL, HOUF, HEN . Identification and characterization of reverse transcriptase fragments of long interspersed nuclear elements (LINEs) in the Morus notabilis genome American Journal of Molecular Biology, 2017, 7(3):138-152. [本文引用: 1]
MILLS RE, BENNETT EA, ISKOW RC, DEVINE SE . Which transposable elements are active in the human genome Trends in Genetics, 2007, 23(4):183-191. [本文引用: 1]
SMIT AF . Interspersed repeats and other mementos of transposable elements in mammalian genomes Current Opinion in Genetics & Development,1999,9(6):657-663. [本文引用: 1]
GAOB, WANGW, QIANY, CHENC, ZHONG JH, SHEND, CHENW, SONG CY . Temporal and spatial expression characteristics of transposons in zebrafish Chinese Journal of Bioinformatics, 2017, 15(4):201-206. (in Chinese) [本文引用: 1]
HAWKINS JS, GROVER CE, WENDEL JF . Repeated big bangs and the expanding universe: Directionality in plant genome size evolution Plant Science,2008,174(6):557-562. [本文引用: 1]
SANMIGUELP, BENNETZEN JL . Evidence that a recent increase in maize genome size was caused by the massive amplification of intergene retrotransposons. Annals of Botany, 1998, 82(suppl_1):37-44. [本文引用: 1]
PEREIRAV . Insertion bias and purifying selection of retrotransposons in the Arabidopsis thaliana genome Genome Biology, 2004,5(10):R79. [本文引用: 1]
WRIGHT SI, AGRAWALN, BUREAU TE . Effects of recombination rate and gene density on transposable element distributions in Arabidopsis thaliana Genome Research ,2003, 13(8):1897. [本文引用: 1]
LOWE CB, BEJERANOG, HAUSSLERD . Thousands of human mobile element fragments undergo strong purifying selection near developmental genes Proceedings of the National Academy of Sciences of the United States of America, 2007,104(19):8005-8010. [本文引用: 1]
IZSV KZ, IVICSZ, PLASTERK RH . Sleeping Beauty, a wide host-range transposon vector for genetic transformation in vertebrates Journal of Molecular Biology, 2000,302(1):93-102. [本文引用: 1]
HORIEK, KUROIWAA, IKAWAM, OKABEM, KONDOHG, MATSUDAY, TAKEDAJ . Efficient chromosomal transposition of a Tc1/mariner-like transposon Sleeping Beauty in mice Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(16):9191-9196. [本文引用: 1]
CARLSON CM, DUPUY AJ, FRITZS, ROBERGPEREZ KJ, FLETCHER CF, LARGAESPADA DA . Transposon mutagenesis of the mouse germline Genetics, 2003, 165(1):243. [本文引用: 1]
MISKEYC, IZSV KZ, PLASTERK RH, IVICSZ . The Frog Prince: A reconstructed transposon from Rana pipiens with high transpositional activity in vertebrate cells Nucleic Acids Research, 2003, 31(23):6873-6881. [本文引用: 1]
MISKEYC, PAPPB, MáTéS L, SINZELLE L, KELLER H, IZSV K Z, IVICS Z. The ancient mariner sails again: Transposition of the human Hsmar1 element by a reconstructed transposase and activities of the SETMAR protein on transposon ends Molecular & Cellular Biology, 2007,27(12):4589-4600. [本文引用: 1]
TAKEUCHIM, MATSUDAK, YAMAGUCHIS, ASAKAWAK, MIYASAKAN, LALP, YOSHIHARAY, KOGAA, KAWAKAMIK, SHIMIZUT . Establishment of Gal4 transgenic zebrafish lines for analysis of development of cerebellar neural circuitry Developmental Biology, 2015,397(1):1-17. [本文引用: 1]
ASAKAWAK, SUSTER ML, MIZUSAWAK, NAGAYOSHIS, KOTANIT, URASAKIA, KISHIMOTOY, HIBIM, KAWAKAMIK . Genetic dissection of neural circuits by Tol2 transposon-mediated Gal4 gene and enhancer trapping in zebrafish Proceedings of the National Academy of Sciences of the United States of America, 2008,105(4):1255. [本文引用: 1]
ZHONGJIEY, QIPENGL, SHANC, XIAOYUL . The function of LINE-1-encoded reverse transcriptase in tumorigenesis Hereditas, 2017,39(5):368-376. [本文引用: 1]
QIANL, JINHUIW, XIAOYUL, SHANC . The connection between LINE-1 retrotransposition and human tumorigenesis Hereditas, 2016: 93-102. [本文引用: 1]
BOULLIOUA, LE PJ, HUBERTG, DONALR, SMILEYM . The endogenous retroviral ev21 locus in commercial chicken lines and its relationship with the slow-feathering phenotype (K) Poultry Science, 1992,71(1):38. [本文引用: 1]
LU XQ, HAN JR, LIU XF, LIN TH, LI YL . The LTR of endogenous retrovirus ev21 retains promoter activity and exhibits tissue specific transcription in chicken Chinese Science Bulletin, 2009,54(24):4664-4670. [本文引用: 1]
GAVORA JS, KUHNLEINU, CRITTENDEN LB, SPENCER JL, SABOUR MP . Endogenous viral genes: Association with reduced egg production rate and egg size in White Leghorns Poult Sciences, 1991,70(3):618-623. [本文引用: 1]
WANGZ, QUL, YAOJ, YANGX, LIG, ZHANGY, LIJ, WANGX, BAIJ, XUG . An EAV-HP Insertion in 5′ flanking region of SLCO1B3 causes Blue Eggshell in the chicken Plos Genetics, 2013,9(1):e1003183. [本文引用: 1]
 ,, 孙桂荣
,, 孙桂荣

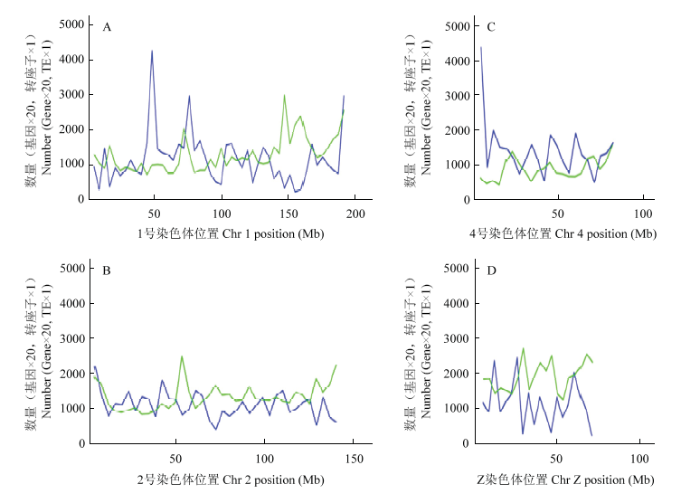 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT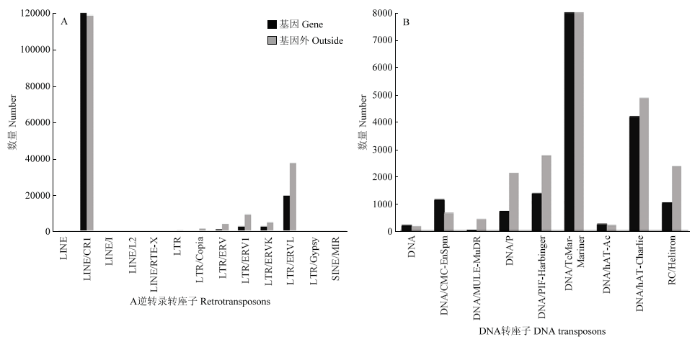 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT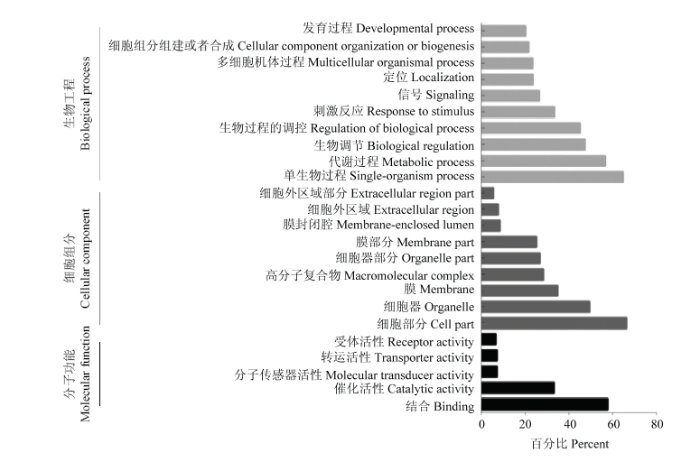 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT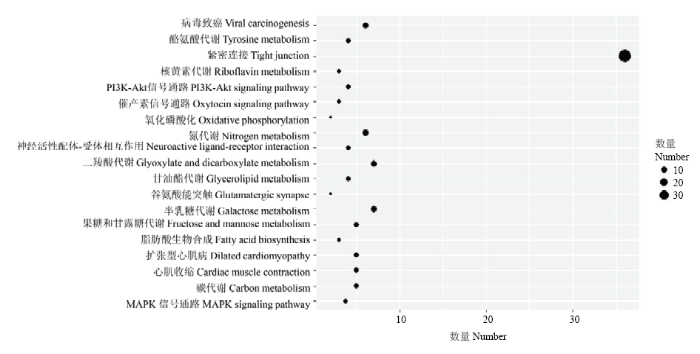 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}