 ,安徽省农业科学院水稻研究所/安徽省水稻遗传育种重点实验室,合肥 230001
,安徽省农业科学院水稻研究所/安徽省水稻遗传育种重点实验室,合肥 230001Transcriptome Research of Erect and Short Panicle Mutant esp in Rice
ZHOU KunNeng, XIA JiaFa, YUN Peng, WANG YuanLei, MA TingChen, ZHANG CaiJuan, LI ZeFu,Rice Research Institute, Anhui Academy of Agricultural Sciences/Key Laboratory for Rice Genetics and Breeding of Anhui Province, Hefei 230001通讯作者:
责任编辑: 李莉
收稿日期:2019-08-5接受日期:2019-10-21网络出版日期:2020-03-16
| 基金资助: |
Received:2019-08-5Accepted:2019-10-21Online:2020-03-16
作者简介 About authors
周坤能,E-mail:zhoukunneng1986@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (5500KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
周坤能, 夏加发, 云鹏, 王元垒, 马廷臣, 张彩娟, 李泽福. 水稻直立短穗突变体esp的转录组研究[J]. 中国农业科学, 2020, 53(6): 1081-1094 doi:10.3864/j.issn.0578-1752.2020.06.001
ZHOU KunNeng, XIA JiaFa, YUN Peng, WANG YuanLei, MA TingChen, ZHANG CaiJuan, LI ZeFu.
0 引言
【研究意义】水稻是最重要的粮食作物之一,供世界上近一半的人口食用。利用株型筛选提高水稻产量是育种的一个重要方向。株型直立不仅能够提高光合效率,而且能够改善温度、湿度、CO2浓度、通风等田间条件,有助于提高植株生长速率和产量。直立穗品种穗型直立、株高较矮,能够提高植株抗倒性和耐肥性。【前人研究进展】目前,水稻中大量控制株型结构的基因被克隆,这些基因通过控制分蘖角度、分蘖数、穗型等调控株型,进而影响水稻产量[1,2,3,4,5]。然而,与直立穗性状有关的基因报道较少。通过QTL定位发现一个控制直立穗性状的基因DEP1,该基因的等位变异能够提高分生组织的活力,减少花序节间长度,增加二次枝梗和二次枝梗粒数,进而增加每穗粒数,提高水稻产量[6,7,8];dep1与氮高效基因qNGR处于相同位点,具有耐肥特性,高氮条件下抗倒能力增强[9];研究表明dep1在增加产量的同时对品质没有显著影响[10];此外,dep1等位基因与gs3和OsMADS1lgy3聚合可以同时提高水稻产量和米质[11]。通过该基因的等位变异选育的直立穗型水稻品种在生产上大面积推广应用[12,13]。水稻直立穗基因DEP2/EP2编码一个植物特异的蛋白,该基因在幼嫩组织特别是幼穗中高度表达,DEP2/EP2突变抑制初级和次级枝梗生长,降低穗长,穗型直立,但不影响植株产量[14,15]。小圆粒基因SRS1与DEP2/EP2为等位基因,srs1系列突变体中株高降低,粒型变小,穗型表现为直立或半直立表型[16]。ep7突变体由中华11组培后代中获得,该突变体表现为穗部直立,株高、千粒重和穗数均显著降低,将其精细定位于水稻第7染色体,突变体存在大片段的DNA插入,该位点可能与DEP2/EP2等位[17]。DEP3突变导致穗型直立,穗长变短,每穗粒数增加,显著提高植株产量[18]。水稻EP3突变后导致穗数和穗粒数减少,穗长变短,结实率降低,穗型直立,同时光合能力下降,参与调控气孔保卫细胞的发育[19,20]。通过对直立穗基因的克隆,利用分子标记结合常规育种手段,可以快速、有效地选育出高产水稻品种[21,22]。【本研究切入点】尽管直立穗水稻品种的培育较早,但关于直立穗基因的研究较少,调控该性状的分子机制仍不够清楚。【拟解决的关键问题】本研究通过MNU(N-甲基-N-亚硝基脲)化学诱变水稻品种宁粳36,获得一个直立短穗突变体esp;通过图位克隆和基因组测序,克隆控制该突变表型的基因ESP;利用转录组研究分析该基因参与的生物学途径,进而解析穗部发育的分子机制。1 材料与方法
1.1 试验材料
水稻直立短穗突变体esp来源于粳稻品种宁粳36的MNU化学诱变突变体库,经过多代自交选择,性状稳定遗传。1.2 农艺性状调查
突变体和野生型于2017年5月在合肥同时播种移栽,成熟期调查株高、有效穗数、穗长、每穗粒数、剑叶长、剑叶宽、千粒重等主要农艺性状,10次重复。1.3 基因定位与克隆
构建突变体esp和PA64籼粳杂交F2定位群体,利用54个具有突变性状的F2典型单株和覆盖水稻12条染色体的分子标记对目的基因进行初定位(电子附表1)。通过基因组测序,分析定位区间内突变体和野生型之间的序列差异,进一步测序验证,发现变异位点,初步确定候选基因。Table 1
表1
表1突变体和野生型的农艺性状分析
Table 1
| 农艺性状Agronomic traits | 野生型WT | 突变体esp |
|---|---|---|
| 株高Plant height (cm) | 79.1±1.7 | 67.2±1.5** |
| 有效穗数Number of effective panicles | 14.8±1.7 | 14.9±1.2 |
| 穗长Panicle length (cm) | 21.06±0.85 | 17.03±0.70** |
| 每穗粒数Grain number per panicle | 190.5±15.2 | 136.7±8.8** |
| 剑叶长Flag leaf length (cm) | 26.60±1.53 | 36.07±2.64** |
| 剑叶宽Flag leaf width (cm) | 1.86±0.05 | 1.83±0.08 |
| 千粒重1000-grain weight (g) | 24.33±0.34 | 25.34±0.42* |
新窗口打开|下载CSV
1.4 生物信息学分析
通过RGAP数据库(1.5 转录组分析
利用天根RNA提取试剂盒(RNA Prep Pure Plant Kit)提取esp突变体和野生型幼穗中的总RNA,琼脂糖电泳和Nanodrop分别检测RNA的完整性和纯度,保证提取的RNA达到建库要求;转录组分析由北京诺禾致源公司完成。表达差异基因通过GO聚类分析,根据差异基因在分子功能、生物学过程和细胞组分上的分布,筛选不同的GO term进行分析;根据KEGG数据库,分析植物激素信号转导和内质网代谢途径中相关基因的表达。
1.6 RNA提取、反转录以及qRT-PCR分析
利用天根RNA提取试剂盒(RNA Prep Pure Plant Kit)提取突变体和野生型幼穗中的总RNA。剪取约100 mg孕穗期新鲜幼穗组织,液氮研磨成粉状,根据试剂盒操作步骤提取组织中的总RNA,琼脂糖电泳和Nanodrop分别检测RNA的完整性和纯度。进一步利用TaKaRa反转录试剂盒(SuperScript II Kit)将RNA反转成cDNA,-20℃保存备用。利用QuantStudio? 3(ABI)定量PCR仪,参考SYBR ? Premix Ex TaqTM Kit(TaKaRa)试剂盒说明书进行qRT-PCR分析,反应体系为cDNA模板2 μL(质量为50—100 ng)、10 μmol·L-1前后引物各0.8 μL、SYBR Premix Ex Taq II 10 μL、50×ROX Reference Dye II 0.4 μL,补ddH2O至20μL,3次样品重复和2次生物学重复。试验中所用到的定量引物见电子附表1,ubiquitin(LOC_Os03g13170,Ubq)为内参对照,利用2-ΔΔCT法分析相对基因表达结果。
2 结果
2.1 直立短穗突变体esp的表型分析
与野生型相比,突变体esp从抽穗期至成熟期株高降低,穗型由弯曲变为直立(图1-A和图1-B),突变体粒长变短约9.12%,粒宽增加约25.09%(图1-C、图1-D和图1-E)。农艺性状调查显示突变体株高、穗长和每穗粒数明显低于野生型,剑叶长和千粒重增加,然而有效穗数和剑叶宽与野生型无显著差异(表1)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1突变体esp和野生型的表型特点
A:成熟期植株表型;B:成熟期穗部表型;C、D和E:种子粒宽和粒长测定
Fig. 1Phenotypic characteristics of the esp mutant and wild type (WT)
A: Plant phenotypes of mature stage; B: Panicle phenotypes of mature stage; C, D and E: Determination of seed width and length
2.2 ESP的克隆
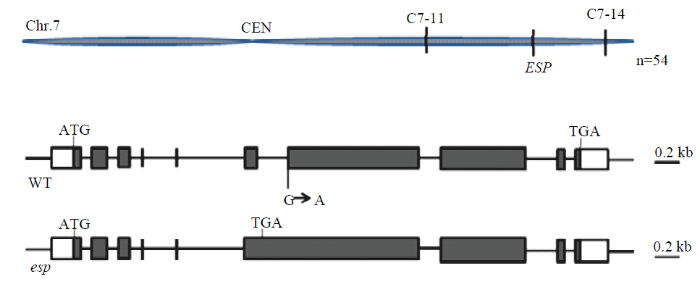
利用突变体esp与籼稻品种PA64杂交,F1植株表现为野生型性状,F2群体中出现弯穗和直立穗表型分离。利用54个具有突变表型的单株以及与突变表型连锁的分子标记将目的基因定位于水稻第7染色体长臂上Indel(insertion-deletion)标记C7-11和C7-14之间7.58 Mb区间内(图2-A)。通过对突变体和野生型进行基因组测序,结合定位结果以及测序验证,发现LOC_Os07g42410在其第6内含子与第7外显子连接位点由碱基G变异为A,导致第6内含子不能被剪切,蛋白翻译提前终止(图2-B);该基因与已报道的水稻基因DEP2/EP2为等位基因,dep2/ep2突变体与esp表型类似,表明LOC_Os07g42410即为控制esp突变表型的目的基因。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2ESP的图位克隆
A:ESP初定位;B:ESP结构和esp突变位点;ATG和TGA分别代表起始和终止密码子
Fig. 2Map-based cloning of the ESP gene
A: Initial mapping of the ESP gene; B: Structure of the ESP gene and mutant site of esp; ATG and TGA indicate start and stop codons, respectively
2.3 ESP及其同源蛋白的序列分析
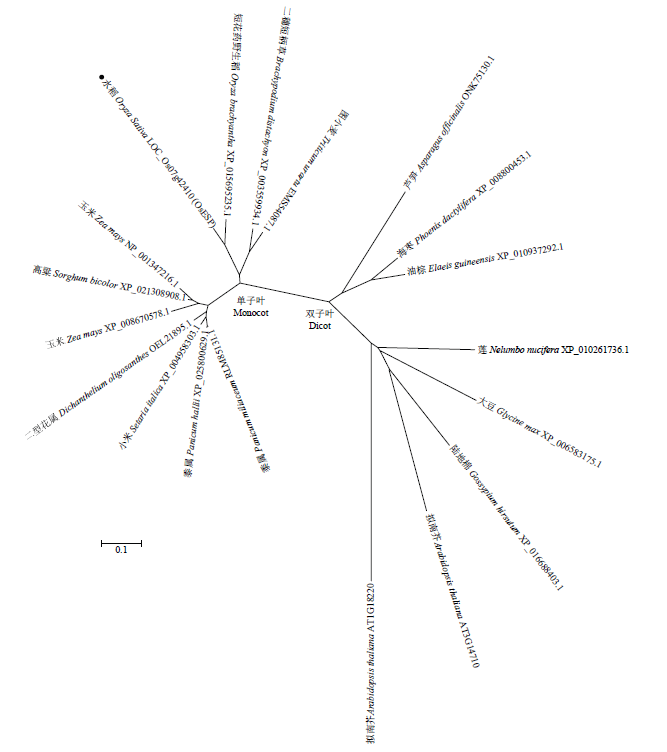
ESP由10个外显子和9个内含子组成,编码一个含1 365个氨基酸残基的蛋白。序列分析显示ESP不含任何结构域,功能未知。突变基因esp编码一个含191个氨基酸残基的蛋白(包括182个原有氨基酸和9个新氨基酸)。进化树分析显示ESP蛋白广泛存在于单子叶和双子叶植物中(图3)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3ESP及其相关蛋白的进化树分析
Fig. 3Phylogenetic analysis of ESP and its related proteins
2.4 ESP组织表达分析
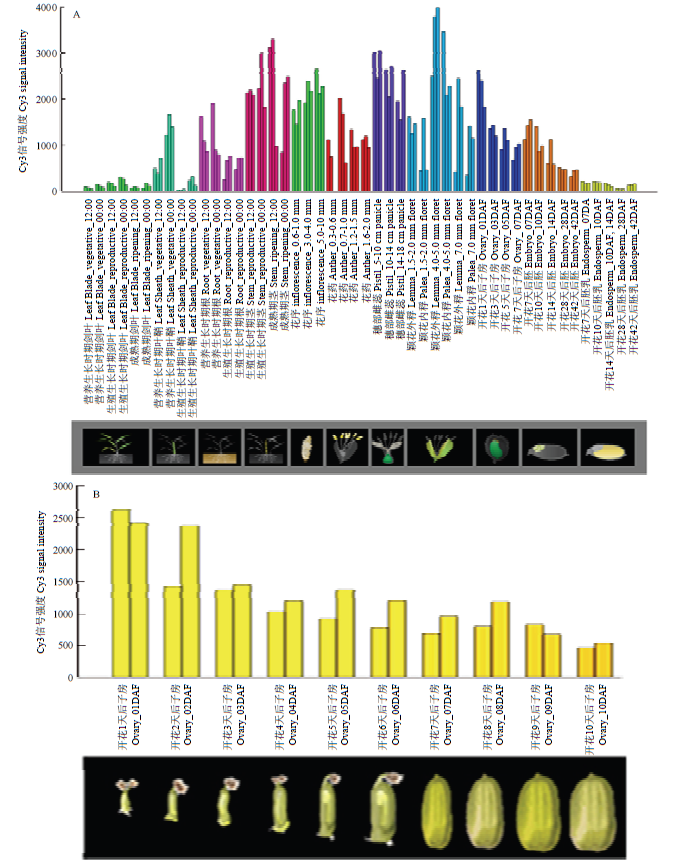
水稻表达模式数据库(RiceXPro)分析表明ESP是一个组成型表达基因,其在水稻发育的不同时期和不同组织器官中都有表达,尤其在茎秆、花序、雌蕊、内外稃和子房中高度表达(图4-A),ESP表达水平随着子房变大而逐渐降低(图4-B)。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4ESP的表达分析
A:不同时期不同组织中ESP的表达模式;B:ESP在子房发育过程中的表达分析。数据来源于水稻表达模式数据库RiceXPro
Fig. 4Expression analysis of ESP gene
A: Expression profile of ESP gene in different tissues at various stages; B: Expression analysis of ESP gene during the progress of ovary development; Data were collected from the rice expression profile database, RiceXPro
2.5 突变体esp和野生型幼穗的转录组分析
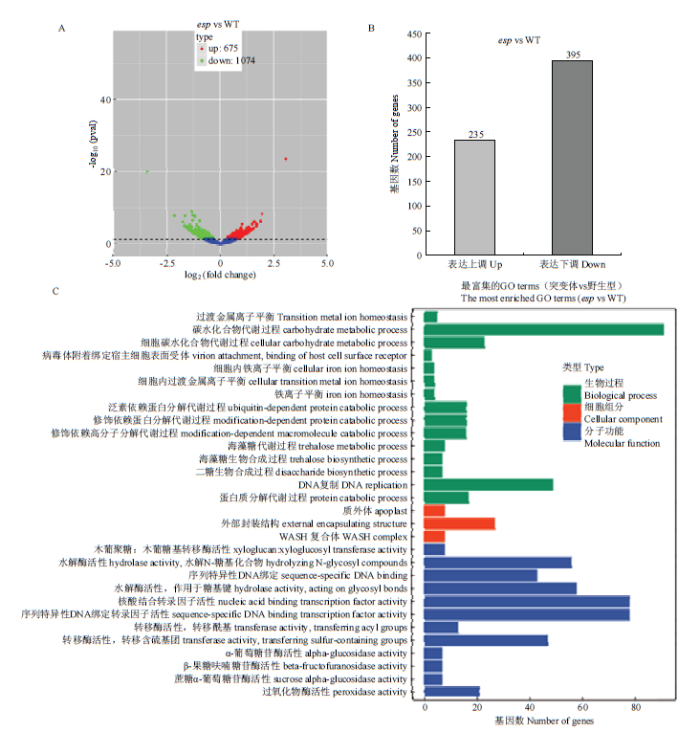
为了解析ESP的分子机理,利用转录组分析突变体esp和野生型幼穗中的基因表达,结果显示,与野生型相比,esp突变体幼穗中675个基因表达上调,1 074个基因表达下调(图5-A);表达差异>1.5倍的基因630个(P<0.01),其中235个表达上调, 395个表达下调(图5-B),这些基因涉及到15项生物学过程、3项细胞组分和12项分子功能(图5-C)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5突变体esp和野生型幼穗转录组分析
A:突变体esp和野生型幼穗转录组分析火山图;红点和绿点分别代表上调基因和下调基因,蓝点代表无显著差异表达基因;B:表达差异>1.5倍的上调和下调基因数目;C:表达差异基因的功能分类
Fig. 5Transcriptome analysis of young panicles in esp mutant and WT
A: Volcano Plot of transcriptome analysis of young panicles in esp mutant and wild type; Red and green dots separately represent up- and down-regulated genes, blue dots indicate genes with no significant difference; B: Numbers of up-regulated and down-regulated genes expressed differentially more than 1.5 time; C: Functional classification of differentially expressed genes
2.6 ESP突变对植物激素信号转导和内质网代谢途径的调控
根据转录组测序结果,与野生型相比,突变体esp中检测到包括次生代谢产物生物合成、淀粉和蔗糖代谢、内质网蛋白加工等11个代谢通路受到明显调控(P <0.1)(表2)。Table 2
表2
表2代谢通路及其差异性表达基因数量
Table 2
| 序号 No. | 通路名称 Pathway name | 差异基因数 DEGs | P值 P value |
|---|---|---|---|
| 1 | 次生代谢产物生物合成 Biosynthesis of secondary metabolites | 86 | 5.47×10-3 |
| 2 | 乙醛酸和二羧酸代谢 Glyoxylate and dicarboxylate metabolism | 12 | 6.19×10-3 |
| 3 | 淀粉和蔗糖代谢 Starch and sucrose metabolism | 22 | 8.09×10-3 |
| 4 | 内质网蛋白加工 Protein processing in endoplasmic reticulum | 28 | 9.59×10-3 |
| 5 | 苯丙素生物合成 Phenylpropanoid biosynthesis | 19 | 1.30×10-2 |
| 6 | 脂肪酸伸长 Fatty acid elongation | 6 | 2.36×10-2 |
| 7 | 植物激素信号转导 Plant hormone signal transduction | 24 | 2.69×10-2 |
| 8 | 氮代谢 Nitrogen metabolism | 6 | 3.63×10-2 |
| 9 | 氰基氨基酸代谢 Cyanoamino acid metabolism | 7 | 4.23×10-2 |
| 10 | 酪氨酸代谢 Tyrosine metabolism | 7 | 4.71×10-2 |
| 11 | 丙氨酸、天冬氨酸和谷氨酸代谢 Alanine, aspartate and glutamate metabolism | 7 | 6.99×10-2 |
新窗口打开|下载CSV
ESP突变后,植株多个性状发育受到抑制,推测ESP可能参与植物激素信号转导。结果表明,与野生型相比,突变体esp幼穗中与生长素信号转导有关的基因AUX1(OS05G0447200)、IAA8(OS02G0723400)、IAA15(OS05G0178600)、IAA23(OS06G0597000)和IAA24(OS07G0182400)上调表达,IAA6(OS01G0741900)下调表达;与细胞分裂素有关的基因ARR3(OS02G0631700)上调表达;与脱落酸有关的基因PYR/PYL(OS03G0297600)、PP2C(OS04G0167900)和SnRK2(OS03G0390200)下调表达;与乙烯有关的基因CTR1(OS02G0527600)、EIN3(OS07G0685700)和EBF1/2(OS06G0605900)下调表达,EIN2(OS07G0155600)上调表达;与茉莉酸有关的基因JAR1(OS01G0221000)和JAZ1(OS09G0439200)上调表达,JAZ2(OS04G0395800)下调表达(图6-A和图6-B)。
图6
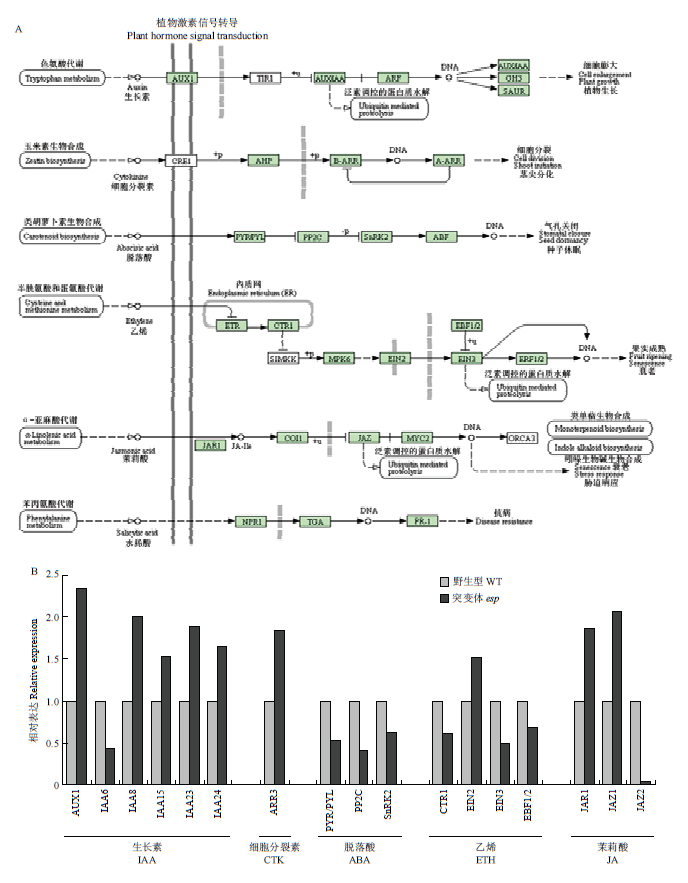 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6植物激素信号转导相关基因的表达分析
A:植物激素信号转导KEGG代谢途径;B:转录组分析植物激素信号转导途径相关基因的表达
Fig. 6Expression analysis of genes related to plant hormone signal transduction
A: Pathway of plant hormone signal transduction; B: Expression of genes related to plant hormone signal transduction in transcriptome analysis
亚细胞定位显示EP2是一个内质网蛋白[15],可能参与调控内质网代谢,转录组结果显示,内质网蛋白加工有关的代谢过程在突变体esp中受到明显的调控(图7-A)。包括与内质网中蛋白绑定和运输有关的基因Sec61(OS09G0347550)上调表达、Bip(OS02G0115900)和SAR1(OS06G0225000)下调表达;与内质网中多聚糖合成有关的基因OsTs-1(OS04G0675500)、OsTs-2(OS01G0911200)和OsTs-3(OS05G0519900)上调表达;参与内质网蛋白的折叠、分选和降解有关的基因DER1(OS05G0187800)、HSP70-1(OS01G0840100)、HSP70-2(OS03G0276500)、HSP70-3(OS11G0187500)、HSP70-4(OS11G0187600)、HSP70-5(OS05G0460000)、HSP90(OS08G0500700)、sHSF-1(OS06G0253100)、sHSF-2(OS06G0219500)、sHSF-3(OS02G0758000)、sHSF-4(OS07G0517100)、p97(OS08G0413000)、Ufd1(OS01G0144500)、UbcH5-2(OS02G0261100)、UbcH5-3(OS02G0721200)、Skp1(OS09G0275200)和Cul1-1(OS01G0369200)下调表达,HSP40(OS03G0648400)、RAD23(OS06G0264300)、UbcH5-1(OS01G0819466)和Cul1-2(OS05G0149600)上调表达(图7-B)。
图7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7内质网中蛋白质加工相关基因的表达分析
A:内质网蛋白质加工KEGG代谢途径;B:转录组分析内质网蛋白质加工途径相关基因的表达
Fig. 7Expression analysis of genes related to protein processing in endoplasmic reticulum
A: Pathway of protein processing in endoplasmic reticulum; B: Expression of genes related to protein processing of endoplasmic reticulum in transcriptome analysis
2.7 转录组验证
为了进一步验证转录组结果,对植物激素信号转导和内质网蛋白质加工途径相关基因进行定量分析。结果与转录组数据一致,同时发现与油菜素内酯信号转导相关基因BIN2(Os02g0236200)和BZR1/2(Os07g0580500)在突变体和野生型之间无显著差异(图8-A和图8-B),表明转录组结果准确。图8
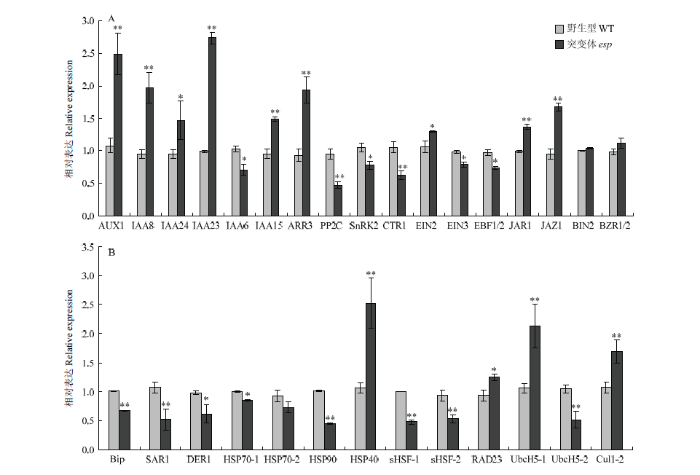 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8突变体和野生型中植物激素信号转导和内质网蛋白质加工途径相关基因的定量PCR分析
A:植物激素信号转导途径相关基因的表达分析;B:内质网蛋白质加工途径相关基因的表达分析。*和**分别代表t测验下差异显著和极显著水平
Fig. 8Quantitative PCR analysis of genes involved in plant hormone signal transduction and protein processing of endoplasmic reticulum between mutant and WT
A: Expression analysis of genes related to plant hormone signal transduction; B: Expression analysis of genes related to protein processing of endoplasmic reticulum; * and ** separately indicate significance at P=0.05 and P=0.01 by Student's t test
3 讨论
水稻株型一直是水稻遗传和育种中研究的重要性状之一,理想的株型能够通过提高植株光合效率、调节群体结构等增加产量;穗部结构是修饰水稻株型的重要因素之一。水稻直立穗株型可以增加植株的透光率,提高光能的利用效率,进而增加产量[6]。目前,大量的直立穗粳稻品种在生产上被广泛种植和推广[7]。然而,关于控制直立穗性状的基因研究较少,调控该性状的分子机制仍不够清楚。本研究克隆了一个控制直立短穗表型的基因ESP,该基因突变后株高降低,穗长变短,每穗粒数减少,粒长变短,粒宽和千粒重增加(图1-E和表1)。研究发现ESP与已克隆水稻基因DEP2/EP2为等位基因。ZHU等[15]鉴定2个分别由籼稻品种中籼3037和9311突变而来的等位突变体ep2-1和ep2-2;与野生型相比,ep2-1和ep2-2株高降低,穗长和粒长变短,粒宽增加,千粒重降低,然而不同的是ep2-1每穗粒数减少,ep2-2每穗粒数增加。此外,另2个等位突变体dep2-1和dep2-2分别由粳稻品种中花11和日本晴变异获得,dep2-1和dep2-2与野生型相比,株高降低,穗长和粒长变短,粒宽增加,千粒重降低,每穗粒数无显著差异[14]。由此可见,ESP等位变异的突变体相对于野生型,株高、穗长、粒长均降低和变短,粒宽增加;然而,不同突变体中每穗粒数和千粒重变化较大,esp和ep2-1中每穗粒数减少,ep2-2中每穗粒数增加,dep2-1和dep2-2中无明显差别;esp中千粒重增加,ep2-1、ep2-2、dep2-1和dep2-2中千粒重降低(表1和表3)。这些表型的差异可能与品种背景以及突变位点有关。
Table 3
表3
表3直立穗基因ESP等位突变体表型特点
Table 3
| 突变体 Mutant | 野生型 Wild type | 株高 Plant height | 穗长 Panicle length | 每穗粒数 Grain number per panicle | 粒长 Grain length | 粒宽 Grain width | 千粒重 1000-grain weight |
|---|---|---|---|---|---|---|---|
| esp | 宁粳36(粳) Ningjing 36 (Japonica) | 降低 Decrease | 变短 Shorten | 减少 Reduce | 降低 Decrease | 增加 Increase | 轻微增加 Slightly increase |
| ep2-1 | 中籼3037(籼) Zhongxian 3037 (Indica) | 降低 Decrease | 变短 Shorten | 减少 Reduce | 降低 Decrease | 增加 Increase | 降低 Decrease |
| ep2-2 | 9311(籼) 9311 (Indica) | 降低 Decrease | 变短 Shorten | 增加 Increase | 降低 Decrease | 增加 Increase | 轻微降低 Slightly decrease |
| dep2-1/dep2-2 | 中花11/日本晴(粳) Zhonghua 11/Nipponbare (Japonica) | 降低 Decrease | 变短 Shorten | 无差别 No difference | 降低 Decrease | 增加 Increase | 降低 Decrease |
新窗口打开|下载CSV
通过研究发现ESP突变导致不同组织、不同性状发生变异,可能影响多个代谢途径。利用转录组分析显示630个与次生代谢产物合成、内质网蛋白加工等多个生物学过程有关的基因在突变体中受到显著调控(图5-B和图5-C)。植物激素通过影响细胞增殖和生长,进而控制粒型的发育;dep2突变体不同组织细胞长度与野生型相比无显著差异,降低的穗长可能由于细胞增殖受到抑制[14],小圆粒突变体srs1-1中细胞长度和细胞数量都明显减少[16],表明DEP2可能与生长素、细胞分裂素、油菜素内酯等激素的信号转导有关;转录组研究和定量分析表明生长素和细胞分裂素信号转导有关的基因表达在esp突变体幼穗中受到调控,然而,油菜素内酯信号转导有关的基因在突变体和野生型之间无显著差异(图6-A、图6-B和图8-A);这表明ESP突变可能不参与调控突变体幼穗中油菜素内酯相关基因的表达。水稻基因TGW6编码IAA-葡萄糖水解酶,使IAA-葡萄糖水解成IAA和葡萄糖,通过调节生长素的水平调控粒型的发育[23];过量表达生长素响应因子OsARF19能够导致水稻粒型变小、株高降低[24,25];OsGAD1通过调节植株细胞分裂素的水平,调控粒长和穗粒数[26]。水稻DEP1等位基因DN1突变后导致植株矮化、每穗粒数增加,表现对油菜素内酯、赤霉素等敏感,但与油菜素內酯突变体间不存在遗传上位性,暗示DN1可能与植物激素无关[27]。说明ESP可能通过调节植物激素有关基因的表达,调控激素水平,进而调节植株穗型和粒型的发育。
直立穗株型能够改良株型结构和群体质量,提高植株生长速率和产量,改善抗倒性和耐肥性,育种利用潜力大[8, 12, 28-31]。解析ESP的不同等位变异与水稻产量之间的关系,不仅能够为水稻育种提供丰富的种质资源,为水稻高产育种提供潜在利用价值,同时有助于理解穗部发育的分子机制。
4 结论
鉴定一个直立短穗突变体esp,并克隆控制该突变性状的基因,其与已报道的水稻基因DEP2/EP2为等位基因。ESP广泛存在于单子叶和双子叶植物中,且在茎秆、花序、雌蕊、内外稃和子房中高度表达,随着子房变大表达水平逐渐降低。ESP可能通过调节植物激素信号转导、内质网蛋白加工过程中相关基因的表达进而影响穗部发育。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
URL [本文引用: 2]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}