 ,, 汤继华,河南农业大学农学院/省部共建小麦玉米作物学国家重点实验室,郑州 450002
,, 汤继华,河南农业大学农学院/省部共建小麦玉米作物学国家重点实验室,郑州 450002Transcriptome Analysis of Maize pTAC2 Effects on Chlorophyll Synthesis in Seedling Leaves
ZHANG Wen, MENG ShuJun, WANG QiYue, WAN Jiong, MA ShuanHong, LIN Yuan, DING Dong,, TANG JiHua,College of Agronomy, Henan Agricultural University/National Key Laboratory of Wheat and Maize Crop Science, Zhengzhou 450002通讯作者:
责任编辑: 李莉
收稿日期:2019-07-22接受日期:2019-09-30网络出版日期:2020-03-01
| 基金资助: |
Received:2019-07-22Accepted:2019-09-30Online:2020-03-01
作者简介 About authors
张稳,E-mail:
孟淑君,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (4009KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
张稳, 孟淑君, 王琪月, 万炯, 马拴红, 林源, 丁冬, 汤继华. 玉米pTAC2影响苗期叶片叶绿素合成的转录组分析[J]. 中国农业科学, 2020, 53(5): 874-889 doi:10.3864/j.issn.0578-1752.2020.05.002
ZHANG Wen, MENG ShuJun, WANG QiYue, WAN Jiong, MA ShuanHong, LIN Yuan, DING Dong, TANG JiHua.
0 引言
【研究意义】植物90%以上的干物质是通过光合作用产生,且作物生物学产量主要来源于叶片的光合产物[1]。叶绿素含量是决定作物光合效率和生物学产量的重要因素[2]。光合作用在叶绿体(chloroplast)中进行,除了进行光合作用之外,叶绿体中还发生与植物生命活动相关的代谢过程,如淀粉的合成和转运,叶绿素类、细胞色素类、蛋白质等的合成[3]。叶绿素的合成是受核基因和质体基因共同控制的复杂过程,其中,某些基因的突变会导致叶绿素合成减少,叶绿体发育受损,光合能力严重降低[4]。【前人研究进展】质体是植物细胞中的一种特定细胞器,负责光合作用,并参与植物生长和发育[5,6]。质体基因主要由2种RNA聚合酶转录,核编码RNA聚合酶(nuclear gene-encoded RNA polymerase,NEP)和质体编码RNA聚合酶(plastid-encoded RNA polymerase,PEP)[7,8]。质体基因至少有3种不同类型的启动子,2种不同来源的RNA聚合酶分别识别对应启动子调控基因的表达[9]。基于启动子的结构,叶绿体基因可分为3类:依赖PEP指导转录的基因,包括psaA、psbE、psbB、psaB、psbA、rbcL和petD等;依赖NEP指导转录的基因,包括rpoB、accD、rpoA、rpoB、rpoC1、rpoC2和Ycf2等;需要2种聚合酶共同指导转录的基因,包括ndhB、ndhF、clpP、atpB和atpE等[10]。NEP主要介导叶绿体发育早期持家基因的表达,包括PEP核心亚单位和参与质体基本功能的基因;而PEP负责80%以上叶绿体基因的表达,它指导产生完全活跃叶绿体所需的光合基因的转录[11,12]。PEP核心蛋白质有2种存在形式:可溶性RNA聚合酶复合物(soluble RNA polymerase,sRNAP)和转录激活染色体复合物(transcriptionally active chromosome,TAC)[13,14,15]。PFALZ等[15]从拟南芥(Arabidopsis thaliana)和芥菜(Sinapis alba)中鉴定出18个新的质体转录激活染色体蛋白质(plasid transcriptionally active chromosomes,pTACs),并对pTAC2、pTAC6和pTAC12等基因的缺失突变体进行研究,证实这三个基因是维持PEP的转录活性所必需的。在拟南芥中,pTAC12不仅影响叶绿体基因的表达,还调节细胞核基因的表达,尤其是在光敏色素信号传导中起作用[16]。拟南芥和芥菜中的铁超氧化物歧化酶FSD2和FSD3均是TAC复合物的组分[15]。双突变体fsd3-1fsd2-1表现为严重的白化表型,叶绿体严重受损,对氧化应激高度敏感,FSD2和FSD3通过保护叶绿体免受ROS的作用,清除早期叶绿体发育中的ROS进而维持叶绿体的正常发育[17]。拟南芥中AtpTAC4编码32 kD囊泡诱导的叶绿体蛋白质VIPP1,对类囊体的形成至关重要[18]。MARÉCHAL等[19]报道了拟南芥中TAC复合物2种组分pTAC1/AtWhy1和pTAC11/AtWhy3,编码这两个蛋白基因的突变会造成叶绿体基因组的异常重组,导致叶绿体发育异常,表现为白化或幼苗致死。对拟南芥AtpTAC14的敲除导致在叶绿体发育的初期类囊体形成受阻,造成苗期白化致死;同时AtpTAC14可与AtpTAC12形成复合体,作为光敏色素依赖性光信号调节叶绿体基因表达[20]。pTAC3在转录的起始,延伸和终止期间与PEP形成复合物,对质体PEP活性和叶绿素的合成是必需的[21]。拟南芥中AtpTAC7通过与4种pTACs组分FLN1、TAC10、TAC12和TAC14相互作用,在调节PEP依赖性叶绿体基因表达和叶绿体发育中起重要作用[22]。在水稻中对OsTRXz进行定点敲除,OsTRXz功能的缺失会造成水稻叶绿体的生物合成受阻,依赖PEP转录和翻译基因被严重抑制,OsTRXz与OsFLN1、OsHSA1相互作用,共同参与组成PEP复合物的形成,在水稻叶绿体基因的转录及叶绿体合成过程中必不可少[23]。在水稻叶色突变体中,叶绿素代谢异常通常会导致叶片黄化或滞绿的表型,如nyc1[24]和pao、rccr1[25]等突变体;而白化表型通常是由于叶绿体发育的异常造成的,如swl1[26]和al1[27]突变体。水稻中OspTAC2定位于叶绿体中,在osptac2纯合突变体中叶绿体发育受损,叶绿素合成受阻。且PEP(质体编码RNA聚合酶)依赖性基因的转录水平在突变体中较CK显着降低,而NEP(核编码RNA聚合酶)依赖性基因的转录水平显著增加。表明OspTAC2在叶绿体发育和叶绿素合成中起关键作用,并表明OspTAC2的分子功能在水稻和拟南芥中是保守的[28]。【本研究切入点】虽然pTACs复合物各成员在拟南芥中报道较多,但在大田作物(包括玉米)中的研究仍然较为欠缺。ZmpTAC2经鉴定是拟南芥和水稻质体转录活性染色体蛋白2(pTAC2)基因的直系同源基因,利用CRISPR/Cas9技术对ZmpTAC2进行定点突变,转基因纯合突变单株表型为叶片白化并且苗期致死,体内叶绿素含量较CK显著降低,叶绿素合成途径受阻且叶绿体发育缺陷。【拟解决的关键问题】本研究以白化表型的CRISPR/Cas9-ZmpTAC2转基因阳性纯合突变材料(zmptac2)为研究对象,对CK和zmptac2幼苗时期叶片取样进行RNA-seq研究,利用qRT-PCR技术验证转录组测序结果,并结合叶绿素含量测定及酵母双杂交等技术,筛选和鉴定参与叶绿素合成的相关基因。1 材料与方法
1.1 试验材料
以玉米自交系KN5585为背景材料,利用未米生物科技(江苏)有限公司开发的高通量CRISPR/Cas9靶点设计程序,对ZmpTAC2(Zm00001d022599)的第一个外显子上ATG下游1 319 bp的PPR结构域上设计了2个相距136 bp的编辑靶位点。构建目的载体CRISPR/Cas9-ZmpTAC2,运用农杆菌侵染的方法导入玉米自交系KN5585中。转基因后共获得15个独立T0转基因苗,其中,叶片表现为正常绿色7株,白化表型8株。1.2 转基因株系阳性鉴定
采用CTAB法提取基因组DNA[29],利用引物(JC-F:5′-CCGTGTCCTGCTCGATTT-3′和JC-R:5′-CTTCCTGCATACCGAGAAGGT-3′)检测转基因材料的编辑情况。PCR反应程序为95℃ 3 min;95℃ 30 s,58℃ 30 s,72℃ 30 s,35个循环;72℃ 8 min。将PCR产物送北京擎科生物技术有限公司测序,并分析测序结果。1.3 潜在脱靶位点分析
使用CRISPR RGEN Tools(http://www.rgenome. net/cas-offinder/)在线BLAST工具,将ZmpTAC2靶序列在玉米全基因组中进行比对,选取与靶序列同源性较高、错配≤4 bp,且具有PAM序列NGG的位点作为潜在脱靶序列,通过单克隆测序评估其脱靶效应。1.4 zmptac2突变体的叶绿体超微结构观察
参考文献[30]制作叶片的透射电镜样品。去除幼苗叶脉,切成2 mm×2 mm的小片,立即放入2.5%戊二醛(pH=7.4)中,抽真空30 min使叶片完全下沉后,室温放置2 h,4℃过夜;PBS缓冲液室温固定5 h;用不同浓度无水乙醇/丙酮溶液梯度脱水;酮/812包埋剂包埋64 h;使用超薄切片机制备60—80 nm超薄切片之后铀铅双染色,使用HITACHI HT7700型透射电子显微镜观察、拍照(武汉赛维尔生物科技有限公司)。1.5 叶绿素含量的测定
根据先前报道的分光光度法测定光合色素含量[31]。切下叶片(约0.2 g鲜重),并在1 mL 100%丙酮中均化,用80%丙酮处理叶片。使用UV-2000分光光度计(尤尼龙,UV-2000),在663 nm和645 nm波长下测定吸光值,按照下列公式计算叶绿素含量。叶绿素a含量(mg·g-1)= (12.72A663-2.59A645)×Vs×稀释倍数/Fw;
叶绿素b含量(mg·g-1)=(22.88A645-4.67A663)×Vs×稀释倍数/Fw;
叶绿素总含量(mg·g-1)=叶绿素a含量+叶绿素b含量。
式中,Fw(fresh weight)为样品鲜重;Vs为提取液总体积(mL)。
1.6 苗期叶片RNA提取、转录组建库及测序
使用TRIzol试剂(Invitrogen,USA)提取幼苗叶片总RNA,并用不含RNase的DNA酶I(TaKaRa,Japan)处理,以获得纯净RNA。使用NanoDrop One检测RNA浓度,并用1%琼脂糖凝胶电泳检测所提取RNA的质量,同时用Agilent 2100检测28S/18S以及RIN值。根据Illumina标准RNA-seq文库构建试剂盒构建链特异性文库,使用双端测序方法进行测序(北京贝瑞和康生物科技有限公司)。1.7 测序结果分析
测序得到原始数据后使用FastQC[32]进行质控,根据质控结果,使用Trimmomatic[33]进行过滤。以B73_V4基因组为参考(B73 AGPv4,ftp://ftp. ensemblgenomes.org/pub/plants/release-42/fasta/zea_mays/dna/),使用HiSat2[34]软件将过滤后的数据进行比对,reads的统计使用featureCounts[35]软件。使用R软件包DESeq2[34]进行基因的差异表达分析,其中p.adjust≤0.01及表达差异倍数大于2的基因认为是差异表达基因(DEGs)。利用R软件包中的Cluster Profiler[36]进行差异基因的GO富集及KEGG富集分析。1.8 qRT-PCR验证
选择15个差异表达的基因进行qRT-PCR验证。以CK和zmptac2的叶片总RNA为模板,反转录合成cDNA第一链,使用NCBI Primer Blast在线设计qRT-RCR反应基因特异引物(电子附表1)。cDNA反转录和qRT-PCR反应体系及反应程序按照PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa)和TB GreenTM Premix Ex TaqTM Ⅱ(TaKaRa)试剂盒说明书进行,每个试验均设3个生物学重复和3个技术重复。以Zm-β-actin作为内参基因,利用2-ΔΔCt法[37]分析基因相对表达水平。1.9 酵母单杂交转录激活活性分析
按照YE等[38]改进的方法进行转录激活活性分析。将含有pG221质粒的酵母菌株EGY48接进50 mL SD/-Ura培养基中,30℃,220 r/min培养16—20 h测A600=1.6—2.0,转接OD600=0.2—0.3,30℃,220 r/min 2.5—3.5 h,使终浓度OD600=0.4—0.6,取菌液制作酵母感受态,将pGBKT7-ZmpTAC2和pGBKT7- ZmpTAC3质粒转化感受态,涂SD/-Trp/-Ura培养基上,30℃培养2—3 d,将长出的酵母菌落在SD/-Trp/ -Ura+X-gal显色平板上划线,30℃培养2 d后观察显色情况。1.10 ZmpTAC2互作蛋白捕捉
使用酵母双杂交系统探究ZmpTAC2蛋白质的互作蛋白。以B73叶片cDNA为模板,PCR扩增获得ZmpTAC2的全长序列,并将此序列与PGBKT7载体连接构建pGBKT7-ZmpTAC2为诱饵载体,转化Y2HGold菌株。验证诱饵蛋白ZmpTAC2的毒性及诱饵载体pGBKT7-ZmpTAC2的自激活特性。将含有pGBKT7-ZmpTAC2的酵母Y2HGold与含有玉米cDNA文库的质粒用共转的方法进行杂交。在SD/-Trp/-Leu/-His/-Ade培养基上筛选,用pGBKT7-AD通用引物pGADT7-F:5′-TAATACGACTCACTATAG GG-3′和pGADT7-R:5′-AGATGGTGCACGATGCAC AG-3′进行菌液PCR扩增。扩增获得的PCR片段送交北京擎科生物技术有限公司测序,测序结果使用NCBI BLAST进行比对分析,挑选候选基因进行点对点回转验证。2 结果
2.1 zmptac2表型及转基因植株阳性编辑情况
对获得的15株T0幼苗进行转基因阳性鉴定,发现7株绿色幼苗中有3株为转基因阴性,4株为转基因阳性;8株白化幼苗全部表现为转基因阳性。对12株转基因阳性材料使用特异引物对含有双靶点的序列进行克隆和靶位点编辑检测,发现转基因阳性绿色幼苗中2株为未编辑,2株为杂合编辑突变,8株白化幼苗为纯合编辑突变,共包含多碱基位点编辑(5株)、158—165 bp的大片段的缺失(4株)及倒位片段重连接(1株)等多种不同编辑导致的突变方式(图1-B,电子附图1)。对转基因阴性材料命名为CK,白化表型且纯合编辑的8株突变材料命名为zmptac2(图1-A)。其中,白化表型的zmptac2-1缺失165 bp、zmptac2-2缺失163 bp、 zmptac-3缺失161 bp、 zmptac-4缺失158 bp,造成ZmpTAC2蛋白质翻译提前终止;zmptac2-5在靶位点1缺失1 bp,在靶位点2缺失3 bp;zmptac2-6在两靶位点间存在163 bp长片段的倒位(图1-B)。将zmptac2-1、zmptac2-2用于后续的转录组测序。RNA-seq结果显示,zmptac2突变体中ZmpTAC2表达量显著降低(CK:3 395.3 RPM,zmptac2:363.2 RPM)。利用qRT-PCR技术,测定CK与zmptac2中ZmpTAC2的相对表达量。结果显示,突变体中ZmpTAC2表达量显著降低,这与转录组测序结果一致。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1CK与zmptac2的表型鉴定及靶位点检测
A:CK与zmptac2苗期叶片表型;B:CK与zmptac2中靶位点检测测序结果
Fig. 1Phenotypic identification and target site detection of CK and zmptac2
A: Leaf phenotype of CK and zmptac2 seedlings; B: CK and zmptac2 target site detection sequencing results
2.2 zmptac2突变体脱靶效应评估
CRISPR/Cas9系统基因编辑的主要取决于sgRNA序列的特异性识别,但由于高等生物基因组较大且较为复杂,sgRNA可能会与非靶点DNA序列发生局部错配,造成CRISPR/Cas9系统基因编辑的脱靶。因此,需要对ZmpTAC2突变体脱靶效应进行评估,选取玉米基因组中与2个sgRNA序列错配碱基数在4 bp以内且具有PAM序列的位点作为潜在脱靶位点,每个靶位点选取3个,共6个(表1),使用NCBI Primer Blast在线设计检测引物(电子附表1)。所选取的8个突变单株的6个潜在脱靶,这6个潜在脱靶位点在T0均未产生脱靶效应(电子附图2),表明所设计的sgRNA序列具有较高的特异性。Table 1
表1
表1CRISPR/Cas9系统潜在脱靶位点突变检测
Table 1
| 编号 Serial number | 潜在脱靶位置 Putative off-target locus | 潜在脱靶序列 Sequence of the putative off-target site (5′-3′) | 错配碱基数 No. of mismatching bases | 测试株数 T0 No. of plants tested | 突变株数 T0 No. of plants with mutations |
|---|---|---|---|---|---|
| Off-target1 | Chr.7:109756398-109756420 | AgCTGGaGTAtAGCCTtATACGG | 4 | 8 | 0 |
| Off-target2 | Chr.1 :145160658-145160680 | AcCTGGGGTACAGCCaGATcAGG | 3 | 8 | 0 |
| Off-target3 | Chr.6:44283532-44283554 | cGaCCACGCAGCTATGTcTGAGG | 3 | 8 | 0 |
| Off-target4 | Chr.4:230687431-230687453 | cGaCCACGCAGCTATGTcTGAGG | 3 | 8 | 0 |
| Off-target5 | Chr.7:8936191-8936213 | TGGCCACGCgGCTAcGTgTGGGG | 3 | 8 | 0 |
| Off-target6 | Chr.7:8936289-8936311 | TGGCCACGCgGCTAcGTgTGGGG | 3 | 8 | 0 |
新窗口打开|下载CSV
2.3 zmptac2突变体的叶绿体超微结构及叶绿素含量测定
利用透射电镜观察了对照(CK)和突变体(zmptac2)幼叶的叶绿体超微结构结果(图2-A)。结果表明,转基因阴性(CK)具有完整的叶绿体结构,基粒类囊体折叠整齐而致密,而zmptac2突变体幼叶的叶绿体内基粒类囊体基粒数目明显少,且排列不规则,基质片层清晰可见,质体小球数目增多,表明叶绿体异常发育。由此推断突变体zmptac2的白化表型可能是通过叶绿体的异常发育而造成的。图2
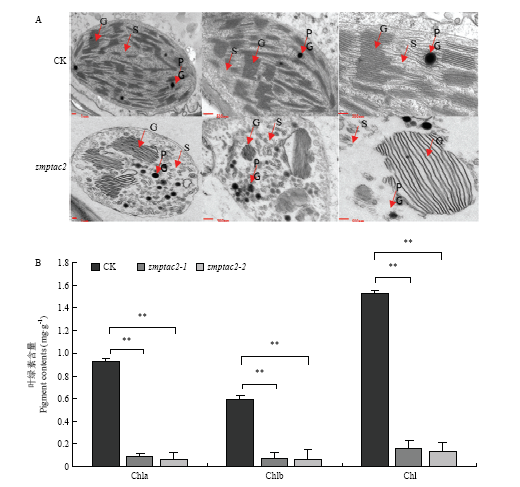 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2zmptac2纯合突变体叶绿体超微结构观察及叶绿素含量测定
A:zmptac2突变体的叶绿体超微结构。G:类囊体基粒;PG:质体小球;S:基质。B:叶绿素含量测定。Chla:叶绿素a;Chlb:叶绿素b;Chl:总叶绿素。*表示0.05水平差异;**表示0.01水平差异。下同
Fig. 2zmptac2 pure mutant chlorophyll content determination
A: Chloroplast ultrastructure of zmptac2 mutant. G: Thylakoid granule; PG: Plastid globule; S: Matrix. B: Chlorophyll content determination. Chla: Chlorophyll a; Chlb: Chlorophyll b; Chl: Total chlorophyll. * indicates difference at the 0.05 level; ** indicates difference at the 0.01 level. The same as below
针对白化表型,对CK和zmptac2突变体中叶绿素a、叶绿素b和总叶绿素含量进行了测定分析。结果显示,与CK相比zmptac2中叶绿素a、叶绿素b及总叶绿素含量显著降低(图2-B)。
2.4 CK与zmptac2幼苗叶片转录组测序质量分析
选取CK材料苗期叶片2份和zmptac2的纯合突变材料(图1,zmptac2-1和zmptac2-2)苗期叶片2份用于RNA提取及后续测序。所测数据经过严格的质控后,产生0.48亿对双末端序列,共计45.3 Gb的序列数据。所测样品Q30比例均超过90%,GC含量均低于50%,比对到参考基因组的比例均在95%以上,结合以上数据可以说明转录组测序质量较高,符合进一步的生物信息学分析要求(表2)。Table 2
表2
表2测序数据统计分析
Table 2
| 样品 Sample | 总碱基数 Total bases (Billion) | GC含量 GC percentage (%) | Q30 比例 Q30 percentage (%) | 比对序列比例 Total mapping reads percentage (%) |
|---|---|---|---|---|
| CK_1 | 15 | 47.52 | 92.90 | 98.71 |
| CK_2 | 16 | 47.50 | 91.94 | 98.75 |
| zmptac2_1 | 12 | 47.52 | 90.29 | 98.87 |
| zmptac2_2 | 16 | 47.92 | 92.17 | 98.56 |
新窗口打开|下载CSV
2.5 差异表基因的表达分析及GO功能富集分析
使用p.adjust≤0.01和差异倍数大于2倍以上为差异表达基因筛选的阈值。在zmptac2-vs-CK中共检测到1 367个差异表达基因,其中618个基因上调表达,749个基因下调表达(电子附表2和电子附表3)。对1 367个差异表达基因做GO(gene ontology)注释,按照基因的细胞组分(cellular component,CC)、分子功能(molecular function,MF)及参与的生物学过程(biological process,BP)进行分类统计。对差异表达基因进行GO功能富集分析,共得到次级分支45个,其中,生物学过程含有16个次级分支,分子功能含有12个次级分支,细胞组分含有17个次级分支(这里仅展示前22条,图3)。图3
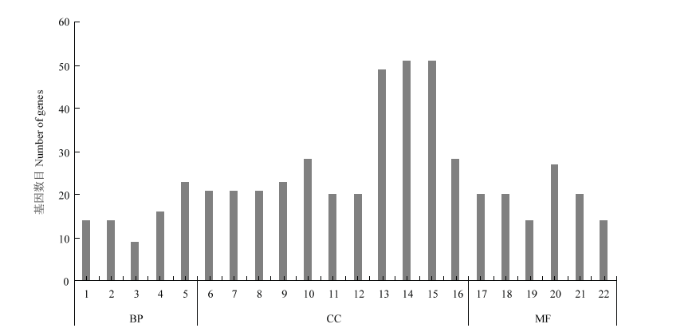 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3差异表达基因GO富集分析
1:细胞或亚细胞成分的运动Movement of cell or subcellular component;2:基于微管的运动Microtubule-based movement;3:有丝分裂细胞周期的调控Regulation of mitotic cell cycle;4:调节细胞成分的组织Regulation of cellular component organization;5:微管过程Microtubule-based process;6:核小体Nucleosome;7:蛋白质-DNA复合物Protein-DNA complex;8:DNA包装复合体DNA packaging complex;9:染色质Chromatin;10:染色体部分Chromosomal part;11:叶绿体基质Chloroplast stroma;12:质体基质Plastid stroma;13:细胞外区域Extracellular region;14:叶绿体Chloroplast;15:质体Plastid;16:染色体Chromosome;17:微管结合Microtubule binding;18:微管蛋白结合Tubulin binding;19:微管运动活动Microtubule motor activity;20:细胞骨架蛋白结合Cytoskeletal protein binding;21:蛋白质异二聚活性Protein heterodimerization activity;22:运动活动Motor activity。BP:生物学过程 Biological process;CC:细胞组分Cellular component;MF:分子功能Molecular function;差异表达基因GO注释,横坐标为GO功能分类,纵坐标为注释到GO条目上的基因数The differentially expressed gene GO annotation, the abscissa is the GO functional classification, and the ordinate is the number of genes annotated to the GO term
Fig. 3Analysis of differentially expressed genes in GO enrichment
根据GO注释结果,上调表达的基因主要富集在细胞周期过程的调节、细胞修饰的氨基酸代谢过程的调节等过程。下调表达的基因主要富集质体基质(20个)、叶绿体基质(20个)、叶绿体(51个)和质体(51个)等细胞组分,表明ZmpTAC2主要通过调控叶绿体相关途径来调节叶绿素的合成途径。
2.6 差异表达基因Pathway富集分析
对差异表达的1 367个基因进行生物学代谢途径(pathway)富集分析,差异基因主要富集在13个代谢途径上(图4)。其中上调基因主要富集在谷胱甘肽代谢途径(glutathione metabolism)、色氨酸生物合成(tryptophan biosynthesis)等途径(图4-A);下调基因主要富集在苯丙氨酸代谢(phenylalanine metabolism)、酪氨酸代谢(tyrosine metabolism)和异喹啉生物碱生物合成(isoquinoline alkaloid biosynthesis)等多种氨基酸代谢途径(图4-B)。图4
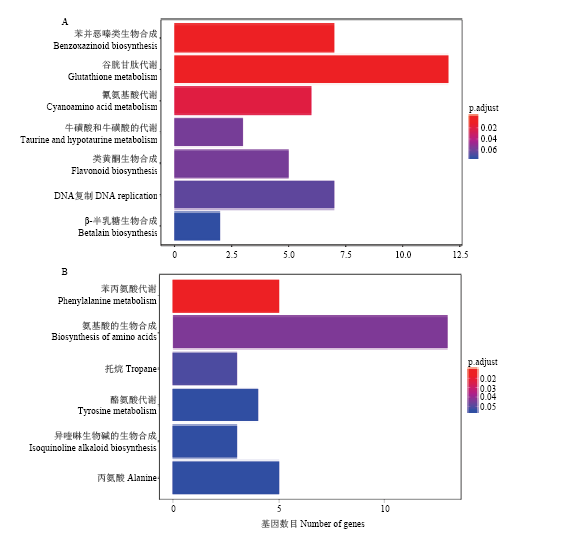 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4差异表达基因KEGG富集分析
A:上调表达基因KEGG(Pathway)富集分析;B:下调表达基因KEGG(Pathway)富集分析
Fig. 4KEGG enrichment analysis of differentially expressed genes
A: Up-regulation of the expression gene KEGG (Pathway) enrichment analysis; B: Down-regulation of expression gene KEGG (Pathway) enrichment analysis
2.7 qRT-PCR验证
根据差异表达的显著性及差异表达基因的功能,挑选15个差异表达基因(表3)进行实时荧光定量PCR以验证RNA测序数据的准确性。qRT-PCR结果显示(图5),这些差异基因在CK与zmptac2材料间的表达模式均与测序数据相一致,表明RNA-seq测序结果是可靠的。Table 3
表3
表3qRT-PCR验证的差异表达基因
Table 3
| 基因编号 Gene ID | zmptac2 | CK | 差异倍数 log2Fold change | P值 P-value | 基因名称 Synonyms | 描述 Description |
|---|---|---|---|---|---|---|
| Zm00001d039206 | 189.7162285 | 4.75059079 | 5.370830095 | 7.82E-06 | Bzip118 | bZIP转录因子超家族蛋白 Putative bZIP transcription factor superfamily protein |
| Zm00001d043825 | 80.63331881 | 0 | 8.789501589 | 0.00185833 | CtpA1 | 羧基末端加工肽酶1 Carboxyl-terminal-processing peptidase 1 chloroplastic |
| Zm00001d027476 | 176.7085969 | 0 | 9.921454443 | 4.88E-06 | DNA polymerases | 依赖DNA的DNA聚合酶 DNA-directed DNA polymerases |
| Zm00001d031503 | 1000.059518 | 22.40077408 | 5.473911936 | 0.00680168 | CPN60 | 伴侣蛋白亚基 Chaperonin 60 subunit alpha 2 chloroplastic |
| Zm00001d008221 | 0 | 351.1761292 | -21.80949 | 1.54E-09 | Cyc5 | 细胞周期蛋白5 Cyclin5 |
| Zm00001d022599 | 363.2261699 | 3395.326772 | -3.03756614 | 5.64E-14 | ZmpTAC2 | 质体转录活性染色体2 Plastid transcriptionally active chromosome 2 homolog |
| Zm00001d033365 | 0 | 362.4571584 | -23.97922 | 0.0000335 | Cyclin A3 1 | 细胞周期蛋白A3-1 Putative cyclin-A3-1 |
| Zm00001d027540 | 0 | 1170.714751 | -25.15007 | 0.0000113 | Gst12 | 谷胱甘肽S-转移酶12 Glutathione S-transferase12 |
| Zm00001d048353 | 0 | 168.1203061 | -22.56015 | 1.08E-08 | Gst13 | 谷胱甘肽转移酶13 Glutathione transferase13 |
| Zm00001d027760 | 0 | 1660.523115 | -24.97806 | 1.32E-05 | H2A | 组蛋白H2A Histone H2A |
| Zm00001d051591 | 10.74539904 | 2639.275824 | -7.938035 | 4.88E-05 | his2b5 | 组蛋白2B5 Histone 2B5 |
| Zm00001d042730 | 87.36474604 | 4052.072328 | -5.535586 | 5.51E-05 | H3 | 组蛋白H3 Histone H3 |
| Zm00001d012276 | 103.9578986 | 6414.512969 | -5.947576 | 0.000341 | H3.2 | 组蛋白H3.2 Histone H3.2 |
| Zm00001d039497 | 40.62122946 | 4971.062871 | -6.93312404 | 8.97E-28 | ClpC1 | 伴侣蛋白ClpC1(线粒体) Chaperone protein ClpC1 chloroplastic |
| Zm00001d042062 | 0 | 2272.297402 | -9.395263783 | 0.00021753 | ORG3 | 转录因子ORG3 Transcription factor ORG3 |
新窗口打开|下载CSV
图5
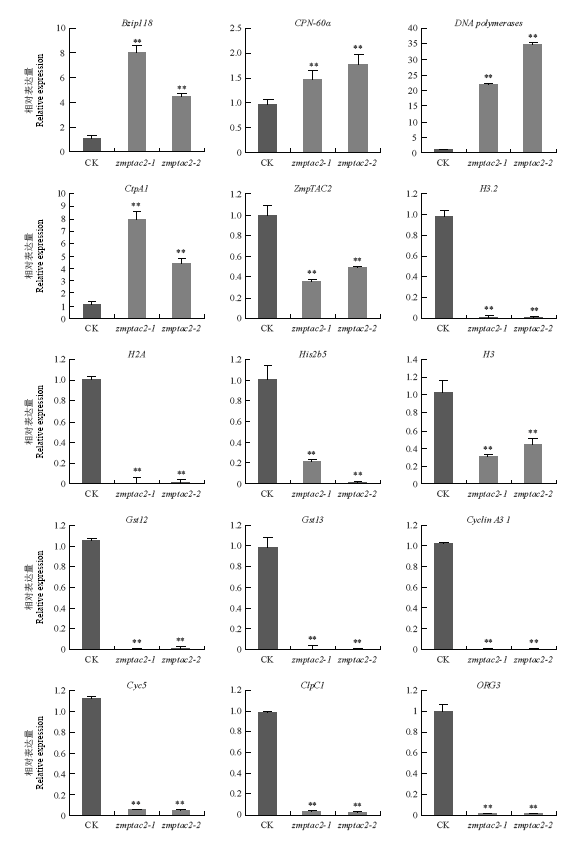 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5zmptac2与CK差异基因的qRT-PCR表达分析
Fig. 5qRT-PCR expression analysis of zmptac2 and CK differential genes
在拟南芥中,已经证实OspTAC2功能缺失时会造成PEP活性降低,进而引起PEP转录的质体基因表达降低,同时促进核编码的RNA聚合酶(NEP)转录基因的表达[24]。据此推测ZmpTAC2也参与调控玉米PEP的活性,进而影响玉米叶绿体基因的转录。为了验证这一推测,在ZmpTAC2转基因阳性株中,对PEP转录相关基因的表达量进行了检测。选择PEP依赖的基因rbcL、psa A、psbA、psaB和psbK,以及NEP依赖的基因rpoA、rpoB、rpoC1和rpoC2进行相对表达量的测定。结果显示,与CK相比,zmptac2中对PEP依赖的基因在RNA水平上表达量显著降低(图6-A),而NEP依赖的基因表达量则显著上升(图6-B)。说明ZmpTAC2对依赖PEP基因的转录具有直接影响,而对依赖NEP基因的转录似乎存在反馈调控。
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6zmptac2突变体内依赖PEP和NEP的相关基因相对表达量
A:依赖PEP的质体基因相对表达量;B:依赖NEP的质体基因相对表达量
Fig. 6zmptac2 mutations in the body rely on PEP, NEP related gene relative expression
A: The relative expression of plasmid genes that rely on PEP; B: The relative expression of plasmid genes that rely on NEP
2.8 酵母双杂及转录激活活性分析
为进一步研究ZmpTAC2在玉米细胞内的功能,通过筛选相互作用蛋白,找到了与ZmpTAC2共同调控叶绿体发育和叶绿素合成的蛋白。通过验证,在SD/-Trp培养基上诱饵蛋白ZmpTAC2生长良好,无毒性(图7-B);pGADT7+pGBKT7-ZmpTAC2在SD/-Trp/- Leu/-His和SD/-Trp/-Leu/-His/-Ade上无法生长,证明诱饵载体pGBKT7-ZmpTAC2无自激活(图7-C)。确定诱饵载体pGBKT7-ZmpTAC2可以用于直接与玉米cDNA文库杂交。通过筛选,共获得了120个阳性克隆。对阳性克隆进行菌落PCR的鉴定,随后在NCBI BLAST上进行序列比对。去除发生移码突变的基因及未注释的基因,共得到32个可能与pTAC2互作的蛋白(表4)。进一步分析这些基因,发现其中16个为核基因编码蛋白,10个为叶绿体基因编码蛋白。例如核编码的ZmpTAC3、α/β水解酶、histone H2A和CPN60等基因;叶绿体编码的CRS1、ATP-dependent DNA helicase等基因。结合转录组分析,将ZmpTAC3(Zm00001d032790)、CPN60(Zm00001d038857)和ATP-dependent DNA helicase(Zm00001d007417)定为候选基因进行点对点验证。结果表明,ZmpTAC2和ZmpTAC3在酵母系统中存在互作(图7-D)。图7
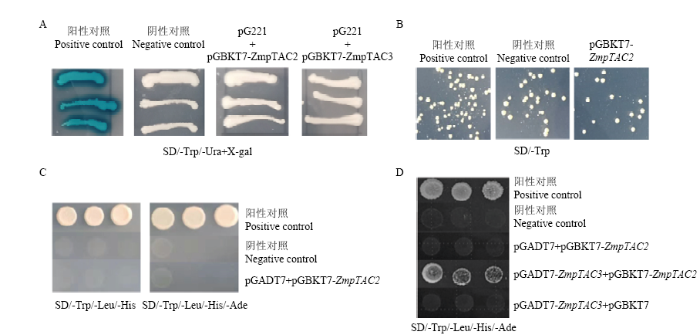 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7转录激活分析及互作蛋白筛选
A:酵母单杂交试验验证ZmpTAC2和ZmpTAC3的转录激活活性;B:诱饵蛋白ZmpTAC2毒性验证;C:诱饵载体pGBKT7-ZmpTAC2自激活验证;D:点对点验证
Fig. 7Transcriptional activation analysis and interaction protein screening
A: Yeast one-hybrid assay to verify the transcriptional activation activity of ZmpTAC2 and ZmpTAC3; B: Toxicity verification of bait protein ZmpTAC2; C: Self-activation verification of bait vector pGBKT7-ZmpTAC2; D: Point-to-point verification
Table 4
表4
表4酵母双杂筛库获得ZmpTAC2的互作蛋白质
Table 4
| 编号 No. | 基因编号 Gene ID_v4 | 亚细胞定位 Subcellular localization | 基因描述 Gene description |
|---|---|---|---|
| 1 | Zm00001d022242 | 细胞膜Cell membrane | 胞间连丝胼胝质结合蛋白5 PLASMODESMATA CALLOSE-BINDING PROTEIN 5 |
| 2 | Zm00001d010039 | 细胞壁Cell wall | 含纤维素酶的蛋白质Cellulase containing protein |
| 3 | Zm00001d023305 | 叶绿体Chloroplast | Protein LURP-one-related 2 |
| 4 | Zm00001d049153 | 叶绿体Chloroplast | 催化/水解酶Catalytic/ hydrolase |
| 5 | Zm00001d041550 | 叶绿体Chloroplast | 热休克蛋白3 Heat shock 70 kDa protein 3 |
| 6 | Zm00001d035270 | 叶绿体Chloroplast | SAM结构域蛋白Sterile alpha motif (SAM) domain-containing protein |
| 7 | Zm00001d028221 | 叶绿体Chloroplast | ATP合酶4 ATP synthase4 |
| 8 | Zm00001d046073 | 叶绿体Chloroplast | Protein trichome birefringence-like 26 |
| 9 | Zm00001d032373 | 叶绿体Chloroplast | 叶绿体RNA剪接体1 Chloroplast RNA splicing1 |
| 10 | Zm00001d007417 | 叶绿体Chloroplast | ATP依赖性DNA解旋酶ATP-dependent DNA helicase chloroplastic |
| 11 | Zm00001d024248 | 叶绿体Chloroplast | PPR蛋白Pentatricopeptide repeat*-59040 |
| 12 | Zm00001d016660 | 细胞质Cytoplasm | 肽基脯氨酰异构酶1 Peptidyl-prolyl isomerase1 |
| 13 | Zm00001d033623 | 细胞质Cytoplasm | 脂氧合酶 Lipoxygenase |
| 14 | Zm00001d018441 | 细胞质,细胞核 Cytoplasm. Nucleus | 核苷三磷酸水解酶超家族蛋白 P-loop containing nucleoside triphosphate hydrolases superfamily protein |
| 15 | Zm00001d040681 | 细胞质,细胞核 Cytoplasm. Nucleus | alpha/beta水解超家族蛋白 Alpha/beta-Hydrolases superfamily protein |
| 16 | Zm00001d038857 | 线粒体Mitochondrion | 伴侣蛋白CPN60 Chaperonin CPN60-like 2 mitochondrial |
| 17 | Zm00001d035201 | 线粒体Mitochondrion | 60S核糖体蛋白P060S acidic ribosomal protein P0 |
| 18 | Zm00001d053434 | 细胞核Nucleus | RING锌指结构域超家族蛋白 Putative RING zinc finger domain superfamily protein |
| 19 | Zm00001d032422 | 细胞核Nucleus | 休眠相关蛋白1 Dormancy-associated protein 1 |
| 20 | Zm00001d044246 | 细胞核Nucleus | 组蛋白H2A Histone H2A |
| 21 | Zm00001d044429 | 细胞核Nucleus | 硫氧还蛋白超家族蛋白 Thioredoxin superfamily protein |
| 22 | Zm00001d021954 | 细胞核Nucleus | MO25蛋白 Putative MO25-like protein |
| 23 | Zm00001d032646 | 细胞核Nucleus | RHC1A |
| 24 | Zm00001d016876 | 细胞核Nucleus | Trihelix转录因子ASIL2 Trihelix transcription factor ASIL2 |
| 25 | Zm00001d043914 | 细胞核Nucleus | Homeobox-DDT结构域蛋白RLT1 Homeobox-DDT domain protein RLT1 |
| 26 | Zm00001d014858 | 细胞核Nucleus | DNA结合蛋白MNB1B DNA-binding protein MNB1B |
| 27 | Zm00001d036494 | 细胞核Nucleus | GATA转录因子25 GATA transcription factor 25 |
| 28 | Zm00001d013159 | 细胞核Nucleus | α-微管蛋白3 Alpha-tubulin 3 |
| 29 | Zm00001d001847 | 细胞核Nucleus | NUMOD3基序家族蛋白 NUMOD3 motif family protein expressed; protein |
| 30 | Zm00001d032790 | 细胞核Nucleus | 质体转录活性3 Plastid transcriptionally active 3 |
| 31 | Zm00001d052298 | 细胞核Nucleus | 锌指(C3HC4型RING指)家族蛋白 Zinc finger (C3HC4-type RING finger) family protein |
| 32 | Zm00001d027652 | 液泡Vacuole | 水通道蛋白TIP1-1 Aquaporin TIP1-1 |
新窗口打开|下载CSV
将pG221(表达GAL4结合结构域)与ZmpTAC2和ZmpTAC3的融合蛋白的质粒转入酵母中,发现ZmpTAC2和ZmpTAC3各自并不能使GAL4顺式序列调控下的报告基因LacZ得到表达形成蓝色菌落(图7-A)。证明ZmpTAC2和ZmpTAC3蛋白在酵母中不具有转录激活活性。初步说明叶绿素的合成需要质体基因间及质体基因组和核基因组之间的相互协调作用。
3 讨论
已有大量研究证明大部分白化和幼苗致死突变体在叶绿体发育或叶绿素合成等方面存在缺陷,这表明叶绿素合成对植物生长和发育至关重要[39,40]。在本研究中,ZmpTAC2纯合突变转基因材料,显示出白化表型,白化苗不能正常存活,苗期死亡。当ZmpTAC2功能缺失时,zmptac2纯合突变转基因材料的总叶绿素、叶绿素a和叶绿素b的含量较CK显著降低(图2)。这表明白化表型是由ZmpTAC2突变引起的,ZmpTAC2是叶绿素合成和正常植物生长所必需的。这些结果与其他植物中报道的TAC相关突变体相似[24]。ZmpTAC2是水稻和拟南芥pTAC2的同源基因(80%和50%氨基酸同一性),且和水稻、拟南芥ptac2突变体有相似的表型,这表明pTAC2基因的分子功能在玉米、水稻和拟南芥之间是保守的。此外,ZmpTAC2与其在其他植物物种中的同源基因的氨基酸序列比对及系统发育分析表明ZmpTAC2在高等植物中是进化上保守的(电子附图3)。对ZmpTAC2定点编辑后,白色表型材料为编辑阳性纯合突变。经靶点检测分析结果表明,编辑主要为3种状态(图1-B),第一种为2个靶位点间存在约150 bp左右的长片段缺失,造成ZmpTAC2蛋白质翻译提前终止;第二种为第一个靶点单碱基缺失,第二个靶点缺失4个碱基;第三种为两靶点间片段倒位重连。这些编辑均造成了白化表型。在ZmpTAC2的相同基因区域发生这些不同类型的编辑,说明利用CRISPR/Cas9系统进行基因编辑可获得不同的编辑效果。包括两个靶位点间的片段缺失(图1-B,zmptac2-1、zmptac2-2、zmptac2-3 和zmptac2-4),是由Cas9蛋白对2个靶位点PAM序列同时编辑所致;2个靶位点分别编辑(图1-B,zmptac2-5),说明发生在2个位点的编辑事件是相对独立发生的;2个靶位点之间片段的倒位在1个编辑阳性事件中被检测到(图1-B,zmptac2-6),说明在2个靶位点间发生剪切后产生的DNA片段可用作断裂DNA双链修复的原料。综上,在使用CRISPR/Cas9系统进行基因编辑时,使用目标基因内相近(100—200 bp)外显子区域的2个靶位点可增加阳性编辑事件产生的类型,进而提高编辑效率。
根据GO富集分析及KEGG富集分析,下调基因GO注释富集在质体中;KEGG注释显示下调基因在苯丙氨酸代谢、酪氨酸代谢显著富集(图3和图4;电子附表2和电子附表3),与叶绿体、类囊体,PSⅠ和PSⅡ及DNA结合蛋白有关。苯丙氨酸在人体相关酶的作用下,可以转化成酪氨酸。若苯丙氨酸和酪氨酸代谢异常,则会产生白化病[41]。RNA-seq测序发现,CtpA3、RPS4A等在白化材料中的表达量显著高于CK。利用qRT-PCR技术验证测序的表达模式,证明这些基因在zmptac2突变体中较CK表达量显著上调,且差异倍数较大。NAKAMURA等[42]研究表明,光合相关基因的调控基因glk突变,叶绿素合成及叶绿体发育受到影响,导致植株光合效率降低。拟南芥中调控叶绿素合成HEMA1的基因突变,导致叶绿素合成过程中镁离子结合效率降低,光合效率下降[43]。核糖体蛋白(RPs)在核糖体生物合成和蛋白质合成中是必不可少的,并且在多种发育过程中起关键作用[44]。核糖体蛋白S4(ribosomal protein small subunit 4,RPS4)在原核生物和植物叶绿体的核糖体30S小亚基形成起始过程中起重要作用;在植物体中,该蛋白由叶绿体rps4编码[45]。CtpA是D1蛋白的羧基末端加工蛋白酶,在菠菜和大麦中CtpA由单拷贝核基因编码,CtpA蛋白仅在黄化材料中检测到,且定位于类囊体腔中[46]。本研究zmptac2突变体中光合作用相关基因CtpA、psb1和RPS4的表达量显著高于CK,而叶绿素的合成依然受阻。说明ZmpTAC2与Psb位于叶绿素合成的不同代谢途径,zmptac2突变直接影响叶绿素的合成,导致白化叶片表型;而叶绿素合成受阻作为一种反馈信号,诱导了CtpA、psb1和RPS4等基因的表达。
本研究中,Cyc基因家族中多个成员、Histone及ClpC1在zmptac2突变体中较CK显著下调。细胞分裂是所有生命体的基本特征之一,在植物生长发育过程中发挥着极其重要的作用[47]。细胞周期是细胞生命活动的重要过程,细胞周期和生长调控均取决于一类重要的细胞分裂调控因子-细胞周期蛋白(Cyclin)。如果细胞周期调控异常,细胞将进入病理状态,甚至死亡[48]。细胞周期蛋白cyclin B1与细胞周期蛋白依赖性蛋白激酶(cyclin-dependent protein kinases,CDKs)结合,通过磷酸化调节多种蛋白的活性与分布而参与纺锤体的形成和染色体的分离[49]。Cyclin A2型细胞周期蛋白,有助于微调植物发育过程中的局部增殖。CYCA2的转录下调代表了在植物发育过程中协调增殖的关键机制[50]。ClpPRS复合物是叶绿体生物发生,类囊体蛋白稳态和植物发育的核心[51]。CLP蛋白酶是ATP依赖性酪蛋白溶解蛋白酶,ClpC1可以促进蛋白酶复合物活性、叶绿体前蛋白的易位并介导蛋白质降解[52]。蛋白质组学分析表明clpc1中光系统蛋白质丰度降低,而Hsp70、Cpn60和一些RNA结合蛋白上调。拟南芥中ClpC1定位在类囊体表面,ClpC1突变后导致叶片褪绿和延迟生长[53]。本文中突变体表现为白化表型,叶绿体发育异常,叶绿素含量降低,ClpC1和CPN60显著降低。本研究在转录组测序数据中检测到zmptac2突变体中与细胞分裂相关的Cyclin、与染色质重塑相关的Histone及与叶绿体发育相关的ClpC1均较CK表达量降低。该结果暗示在突变体中,也许染色质趋于开放,细胞周期的异常,导致细胞分裂严重受阻,这可能是造成突变体叶绿体发育异常表型的部分原因。
4 结论
ZmpTAC2定点编辑的阳性纯合单株表现出严重的白化且幼苗致死;编辑阳性单株叶绿体发育异常,叶绿素含量显著降低;依赖PEP转录的基因表达量显著降低。ZmpTAC2是玉米叶绿素合成的重要调节基因,且是通过调控PEP相关基因表达而实现的。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}