 ,山东农业大学农学院/作物生物学国家重点实验室,山东泰安 271018
,山东农业大学农学院/作物生物学国家重点实验室,山东泰安 271018Effects of Phytase Q9 on Yield Formation of Summer Maize Shading in the Field
HUANG XinHui, GAO Jia, REN BaiZhao, ZHAO Bin, LIU Peng, ZHANG JiWang,College of Agriculture, Shandong Agricultural University/State Key Laboratory of Crop Biology, Taian 271018, Shandong通讯作者:
责任编辑: 杨鑫浩
收稿日期:2019-04-8接受日期:2019-07-19网络出版日期:2019-10-01
| 基金资助: |
Received:2019-04-8Accepted:2019-07-19Online:2019-10-01
作者简介 About authors
黄鑫慧,E-mail:1334112037@qq.com。

摘要
关键词:
Abstract
Keywords:
PDF (532KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
黄鑫慧, 高佳, 任佰朝, 赵斌, 刘鹏, 张吉旺. 植酶Q9对大田遮阴夏玉米产量形成的影响[J]. 中国农业科学, 2019, 52(19): 3309-3322 doi:10.3864/j.issn.0578-1752.2019.19.003
HUANG XinHui, GAO Jia, REN BaiZhao, ZHAO Bin, LIU Peng, ZHANG JiWang.
0 引言
【研究意义】光是玉米能量的主要来源,直接或间接地影响着玉米的产量形成[1,2]。玉米是高光效的C4作物,合理的光照强度和光照时间对玉米的高产优质至关重要。弱光胁迫影响夏玉米正常授粉,降低夏玉米的净光合速率、茎秆抗倒伏能力、内源激素水平、抗氧化酶系活性,进而影响产量形成[3,4,5,6,7]。近些年大气污染严重,地球表面太阳辐射量逐年下降[8,9],再加上黄淮海地区夏玉米的生长季高温多雨寡照频发以及农户生产上种植密度的增加,造成了夏玉米生长期间光照不足,这已成为制约黄淮海地区夏玉米产量进一步提高的重要限制因素之一。通过选用适宜的玉米品种和合理的农艺管理措施缓解弱光对夏玉米产量的影响,是当前黄淮海区夏玉米生产上急需解决的问题。【前人研究进展】遮阴影响夏玉米穗分化,生育期延长、雌雄间隔增加,雄穗小花败育率增加,导致雌穗小花受精率降低;另外遮阴后籽粒内源激素的不利变化引起籽粒败育,造成穗粒数减少。遮阴后夏玉米光合能力降低,干物质积累量减少,再加上遮阴减少了胚乳细胞数量,导致籽粒灌浆速率变慢,籽粒充实度降低,粒重降低。遮阴也降低夏玉米抗倒伏能力,田间倒伏率显著增加。这一系列的影响造成夏玉米减产,且减产程度表现为全生育期遮阴>穗期遮阴>花粒期遮阴[4-5,10]。化学调控是农业生产中常用的农艺管理措施,前人研究发现有许多植物生长调节剂对于谷子、大豆、小麦、玉米、油菜、甘蔗、水稻、棉花、草莓等的生长发育及品质具有正调控作用,可以促进花芽分化,提高叶面积指数,延长叶片功能期,提高光热利用率,促进氮代谢,延缓衰老,增强抗逆性和抗倒伏性能,缓解逆境危害,提高作物的品质和产量[11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26]。植酶Q9的主要成分是黄腐酸,黄腐酸在农业生产中有广泛应用,前人研究表明,黄腐酸可以提高苹果的氮素和光能利用率[27,28];延长马铃薯生育期、改善植株性状和光合性能,提高产量和商品薯率[29,30];增加苜蓿根瘤数、提高固氮酶活性,提高产量[31];缓解干旱胁迫下玉米的减产危害[32];促进番茄生长发育、改善叶片光合特性,提升食用品质,提高磷胁迫下植株中磷转运相关基因和质子泵基因的表达,降低无氧呼吸产物对植株的危害,进而缓解缺磷症状[33,34];提高硝酸盐胁迫下小白菜的抗氧化酶活性,降低膜脂过氧化水平,增强抗逆相关基因的表达,进而缓解硝酸盐胁迫[35]。张彩凤等[36]通过分子模拟软件对玉米喷施黄腐酸的作用机理研究表明,黄腐酸结合钙调蛋白后,形成的稳定配合物与SOD结合,使SOD的能量降低,从而促进植株体内SOD合成。【本研究切入点】植物生长调节剂对甘蔗、棉花、玉米、小麦、水稻等作物的调节研究已有很多报道[22,23,24,25],其中玉米的生长调节剂研究大多数是关于抗倒伏、淹水、干旱、冷胁迫的[18,20,26,37-38],鲜有关于缓解弱光胁迫的相关报道。【拟解决的关键问题】本论文旨在研究植酶Q9对遮阴胁迫夏玉米生长发育和产量的调控作用,以期为黄淮海区夏玉米的高产优质生产提供科学依据。1 材料与方法
1.1 试验地点
2013—2018年,本试验在山东农业大学试验农场(36.09°N,117.09°E)进行,当地为温带大陆性季风气候。土壤类型为棕壤土,土壤pH为7.12,播种前0—20 cm土壤养分状况为有机质9.34 g·kg-1,全氮0.76 g·kg-1,全磷0.88 g·kg-1,碱解氮80.61 mg·kg-1,速效磷37.19 mg·kg-1,速效钾84.23 mg·kg-1。1.2 试验设计
2013—2018年均选用夏玉米品种登海605(DH605)为试验材料,种植密度为67 500株/hm2,化控试剂为植酶Q9。设置3个遮阴处理,分别为开花至收获期遮阴(S1)、拔节至开花期遮阴(S2)和出苗至收获期遮阴(S3),以自然光照为对照(CK)。另外选用化控试剂植酶Q9对遮阴和正常光照处理进行外源调控,将植酶Q9原液按1﹕100的比例稀释,于拔节期(V6)、抽雄期(VT)分别对S1、S2、S3和CK的叶片正反面均匀喷施,即开花至收获期遮阴-植酶Q9(Z-S1)、拔节至开花期遮阴-植酶Q9(Z-S2)、出苗至收获期遮阴-植酶Q9(Z-S3)和自然光照-植酶Q9(Z-CK),并以同时期喷施清水为对照。遮阴处理透光率40%,利用脚手架和遮光率为60%的黑色遮阴网搭建可升降式遮阴棚,遮阴网与玉米冠层间距始终保持2 m,保证遮阴棚内小气候与大田自然光照条件基本一致。小区面积27 m2(9 m×3 m),3次重复,随机排列。肥料按12 000 kg·hm-2的产量水平施用(N 210 kg·hm-2、P2O5 75 kg·hm-2、KCl 150 kg·hm-2)。氮肥分别于拔节期(V6)施入40%,大喇叭口期(V12)施入60%,磷肥和钾肥于V6期一次性施入,按高产田水平进行田间管理。
1.3 测定项目及方法
1.3.1 田间小气候 光照强度、气温、CO2浓度、相对湿度、风速和地温(5 cm)等指标测定参照崔海岩等[5]的方法,开花后连续测定7 d,均在11:00开始测定(表1)。Table 1
表1
表1花粒期不同光照强度对田间小气候的影响
Table 1
| 处理 Treatment | 风速 Air speed (m·s-1) | 气温 Air temperature (℃) | 地温 Soil temperature (℃) | 相对湿度 Relative humidity (%) | 光照强度 Light intensity (μmol·m-2·s-1) | CO2浓度 CO2 concentration (μmol·mol-1) |
|---|---|---|---|---|---|---|
| CK | 0.91a | 29.8a | 24.4a | 50.6a | 1676a | 324a |
| S | 0.9a | 29.4a | 24.1a | 50.1a | 681b | 326a |
新窗口打开|下载CSV
1.3.2 生长发育进程 播种后观察出苗情况,记录出苗(VE)期。出苗后,每个处理选取有代表性的植株调查其生育进程。记录各处理的拔节期(V6)、大喇叭口期(V12)、抽雄期(VT)、吐丝期(R1)、乳熟期(R3)、成熟期(R6)的日期。
1.3.3 叶面积指数(LAI) 于V12、VT、R3、R6期,选择能代表小区整体长势的植株,每处理选10株,测量每片叶的长度和最大叶宽值。
单叶叶面积(cm2)=叶长(cm)×叶宽(cm)×0.75
LAI=(单株叶面积×每个小区的植株数)/小区面积
1.3.4 功能叶片SPAD值 使用SPAD-502便携式叶绿素仪(Soil-plant Analysis Development Section,Minolta Camera Co.,Osaka,Japan)测定叶绿素相对含量值(SPAD)。于V12、VT、R3和R6期,上午9:00—12:00在每小区选取能代表小区整体长势的10株植株进行测定(V12测定最新展开叶,VT及其以后各时期测穗位叶,测定时避开叶脉),取平均值。
1.3.5 干物质积累与分配 于V12、VT、R3和R6期取样,每个处理选取能代表小区整体长势的5株,V12、VT期植株分为茎、叶2部分,R3、R6期植株分为茎、叶、穗轴和籽粒4部分,置烘箱内110℃杀青30 min,然后80℃烘干至恒重称重。
1.3.6 植株性状 R3期各处理选取15株长势一致的代表性植株,测量植株的株高、穗位高,计算穗位系数;同时测量地上茎秆第3节茎粗,取平均值。玉米倒伏后,调查小区植株倒伏情况。
穗位系数=穗位高/株高×100%
倒伏率=倒伏株数/小区总株数×100%
1.3.7 收获、考种和测产 于每小区中间3行,随机选取30株玉米的果穗,自然风干用于室内考种、计算产量(按14%含水量折算)。收获穗数为田间调查有效穗数。
1.4 数据分析
采用Microsoft Excel 2010和SPSS 10.0软件进行数据统计和分析。利用软件SigmaPlot 10.0作图。2 结果
2.1 产量及其构成因素
2013—2018年,遮阴显著降低夏玉米产量, 减产程度为S3>S1>S2,分别较CK平均降低61%、47%、28%。喷施植酶Q9后,Z-S3的产量较S3平均增加21%,Z-S2较S2平均增加9%,Z-S1较S1平均增加14%,Z-CK较CK无明显差异(表2)。Table 2
表2
表2植酶Q9对大田遮阴夏玉米产量及其构成因素的影响
Table 2
| 年份 Year | 处理 Treatment | 产量 Yield (kg·hm-2) | 千粒重 1000-grain weight (g) | 穗粒数 Kernels per ear | 穗数 Harvest ear number (ears/hm2) | 空秆率 Empty stem rate (%) |
|---|---|---|---|---|---|---|
| 2013 | S3 | 4874c | 329c | 276c | 53706b | 13.51 |
| Z-S3 | 5698b | 351b | 300b | 54237a | 10.91 | |
| CK | 9333a | 358a | 509a | 51285b | 8.08 | |
| Z-CK | 10165a | 368a | 517a | 53421b | 4.83 | |
| 2014 | S3 | 1666c | 246c | 162c | 41854b | 20.47 |
| Z-S3 | 2136b | 265b | 189b | 42595b | 17.05 | |
| CK | 10441a | 320b | 520a | 62670a | 4.41 | |
| Z-CK | 10856a | 339a | 524a | 61003a | 4.85 | |
| 2015 | S3 | 3056g | 311c | 260g | 37860g | 11.65 |
| Z-S3 | 4137f | 314b | 308f | 42778f | 10.95 | |
| S2 | 7888c | 314b | 450c | 55864e | 8.08 | |
| Z-S2 | 9523b | 313b | 488b | 62346c | 4.61 | |
| S1 | 6087e | 313b | 327e | 59506d | 2.95 | |
| Z-S1 | 7207d | 314b | 361d | 63580b | 2.33 | |
| CK | 11850a | 316a | 568a | 65965a | 1.30 | |
| Z-CK | 11566a | 313b | 563a | 65635a | 0.85 | |
| 年份 Year | 处理 Treatment | 产量 Yield (kg·hm-2) | 千粒重 1000-grain weight (g) | 穗粒数 Kernels per ear | 穗数 Harvest ear number (ears/hm2) | 空秆率 Empty stem rate (%) |
| 2016 | S3 | 5854g | 328g | 320e | 55864g | 8.59 |
| Z-S3 | 6862f | 333f | 350d | 58951e | 5.91 | |
| S2 | 9186d | 346d | 455c | 58333f | 5.97 | |
| Z-S2 | 9790c | 351c | 464c | 60185d | 4.41 | |
| S1 | 6798f | 342e | 322e | 61728c | 7.54 | |
| Z-S1 | 7505e | 343e | 357d | 61420c | 5.24 | |
| CK | 12852b | 364a | 550b | 64198b | 2.35 | |
| Z-CK | 13494a | 358b | 574a | 65741a | 1.39 | |
| 2017 | S3 | 5219e | 336e | 330e | 47084c | 30.25 |
| Z-S3 | 6137d | 340d | 377d | 47921c | 29.01 | |
| S2 | 8765b | 342d | 432b | 59252b | 12.22 | |
| Z-S2 | 9048b | 346c | 433b | 60324b | 10.63 | |
| S1 | 6360d | 335e | 310d | 61365b | 9.09 | |
| Z-S1 | 7511c | 341d | 360c | 61210b | 9.32 | |
| CK | 12371a | 366a | 527a | 64208a | 4.88 | |
| Z-CK | 11954a | 355b | 516a | 65350a | 3.19 | |
| 2018 | S3 | 6882f | 334e | 312f | 66069h | 2.12 |
| Z-S3 | 7462e | 337d | 334d | 66328g | 1.74 | |
| S2 | 10074c | 340c | 445b | 66529f | 1.44 | |
| Z-S2 | 10525b | 347b | 452b | 67083b | 0.62 | |
| S1 | 7294e | 334e | 326e | 67045d | 0.67 | |
| Z-S1 | 7881d | 345b | 342c | 66840e | 0.98 | |
| CK | 12712a | 356a | 532a | 67079c | 0.62 | |
| Z-CK | 12775a | 358a | 529a | 67500a | 0.00 | |
| ANOVA | ||||||
| 年份Year (Y) | ** | ** | ** | ** | ** | |
| 遮阴Shading (S) | ** | ** | ** | ** | ** | |
| 化控Regulation (R) | ** | ** | ** | ** | ** | |
| Y×R | ** | ** | ** | ** | ** | |
| Y×S | ** | ** | ** | ** | ** | |
| R×S | * | NS | ** | ** | ** | |
| Y×R×S | ** | ** | ** | ** | ** |
新窗口打开|下载CSV
遮阴显著降低夏玉米的千粒重、穗粒数、公顷穗数。与CK相比,千粒重降低程度S3最大(平均为10%),S1次之(平均为5%),S2最低(平均为4%);喷施植酶Q9后,Z-S3较S3增加3.2%,Z-S2较S2增加1.6%,Z-S1较S1增加1.4%。穗粒数降低程度S3最大(平均为48%),S1次之(平均为41%),S2最低(平均为18%);喷施植酶Q9后,Z-S3较S3增加12%、Z-S2较S2增加3%、Z-S1较S1增加11%。公顷穗数降低程度S3>S1>S2(除2013年变化不显著),喷施植酶Q9后,除2014年、2017年、2016年(Z-S1)以外,其他年份均显著增加。遮阴提高夏玉米空秆率,除2016年呈S3>S1>S2趋势之外,其他年份呈S3>S2>S1的变化趋势。喷施植酶Q9后,除2017年的Z-S1外,其他遮阴处理空秆率降低(表2)。
2.2 穗部性状
遮阴后夏玉米的雌穗穗长、穗粗降低,秃顶变长,其中S3、S1的变化最显著,与CK相比,S3、S1雌穗穗长分别降低23.2%、23.5%,穗粗分别降低11%、7%,秃顶长分别增加68%、70%。遮阴后喷施植酶Q9可改善穗部性状,Z-S3秃顶长减小,穗长显著增加,穗粗变化不明显,Z-S2、Z-S1、Z-CK处理穗部性状变化不显著(表3)。Table 3
表3
表3植酶Q9对大田遮阴夏玉米穗部性状的影响
Table 3
| 年份 Year | 处理 Treatment | 穗长 Ear length (cm) | 秃顶长 Barren ear length (cm) | 穗粗 Ear diameter (cm) |
|---|---|---|---|---|
| 2015 | CK | 20.2a | 1.35e | 4.63a |
| Z-CK | 19.9a | 1.36e | 4.43b | |
| S3 | 14.8d | 2.47a | 3.97ef | |
| Z-S3 | 15.8c | 1.92c | 4.13e | |
| S2 | 17.7b | 1.53d | 4.4bc | |
| Z-S2 | 17.8b | 1.48d | 4.3cd | |
| S1 | 14.6d | 2.39b | 4.26d | |
| Z-S1 | 15.0d | 2.33b | 4.39bc | |
| 2016 | CK | 19.8a | 1.61d | 4.68ab |
| Z-CK | 19.9a | 1.69d | 4.72a | |
| S3 | 15.1e | 2.34bc | 4.13d | |
| Z-S3 | 16.1d | 1.87cd | 4.17d | |
| S2 | 18.2c | 2.40b | 4.52bc | |
| Z-S2 | 19.1b | 2.30bc | 4.47c | |
| S1 | 15.0e | 2.96a | 4.28d | |
| Z-S1 | 15.1e | 2.34bc | 4.24d | |
| 2018 | CK | 18.8a | 1.07c | 4.35a |
| Z-CK | 18.9a | 0.97c | 4.30a | |
| S3 | 15.3c | 1.87a | 4.04b | |
| Z-S3 | 15.9b | 1.02c | 4.05b | |
| S2 | 18.6a | 1.16c | 4.19ab | |
| Z-S2 | 18.7a | 1.00c | 4.19ab | |
| S1 | 15.3c | 1.65ab | 4.14ab | |
| Z-S1 | 15.3c | 1.58b | 4.17ab |
新窗口打开|下载CSV
2.3 干物质积累与分配
遮阴显著降低了各时期的干物质积累量,降低程度S3>S1>S2,与CK相比,在V12、VT、R3、R6期,S3分别降低54%、54%、46%、50%,S2分别降低36%、44%、27%、24%,S1在R3、R6期分别降低34%、40%。喷施植酶Q9后,遮阴处理各时期干物质积累量增加(图1)。以2015年为例,在V12、VT、R3、R6期,Z-S3较S3分别增加21%、32%、26%、35%,Z-S2较S2分别增加23%、24%、17%、4%,Z-S1较S1在R3、R6期分别增加21%、4%。植酶Q9也提高了遮阴处理干物质在籽粒中的分配比例(表4)。图1
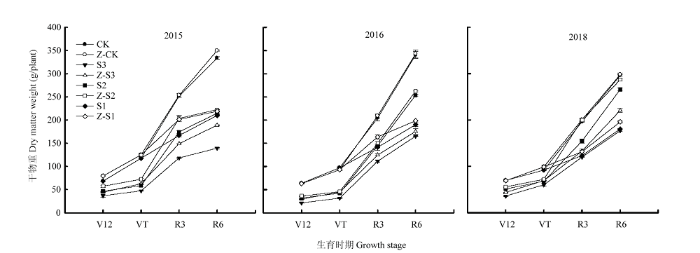 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1植酶Q9对大田遮阴夏玉米干物质积累量的影响
V12:大喇叭口期;VT:抽雄期;R3:乳熟期;R6:成熟期。下同
Fig. 1Effects of Phytase Q9 on dry matter accumulation of summer maize shaded in field
V12: Male tetrad stage; VT: Tassel stage; R3: Milk stage; R6: Maturity stage. The same as below
Table 4
表4
表4植酶Q9对大田遮阴夏玉米干物质分配的影响
Table 4
| 年份 Year | 处理 Treatment | 茎Stalk | 叶Leaf | 穗Kernel | 总干重 Total dry matter (g/plant) | |||
|---|---|---|---|---|---|---|---|---|
| 干重 Dry matter (g/plant) | 比例 Proportion (%) | 干重 Dry matter (g/plant) | 比例 Proportion (%) | 干重 Dry matter (g/plant) | 比例 Proportion (%) | |||
| 2015 | S3 | 51.8c | 34 | 27.9b | 19 | 56.8d | 38 | 150.6d |
| Z-S3 | 51.2c | 27 | 32.4a | 17 | 88.4c | 47 | 188.7c | |
| S2 | 51.6d | 23 | 25.6b | 12 | 120.5b | 54 | 222.3bc | |
| Z-S2 | 55.8c | 23 | 28.0b | 12 | 126.5b | 53 | 240.3b | |
| S1 | 73.9b | 38 | 37.7a | 19 | 72.0d | 37 | 196.5c | |
| Z-S1 | 82.2a | 39 | 35.9a | 17 | 97.8c | 42 | 230.5b | |
| CK | 87.5a | 28 | 36.3a | 12 | 166.1a | 53 | 313.8a | |
| Z-CK | 89.5a | 27 | 38.1a | 12 | 177.2a | 54 | 329.8a | |
| 2016 | S3 | 51.5d | 41 | 21.0c | 17 | 44.7e | 35 | 126.8f |
| Z-S3 | 48.6d | 30 | 22.7c | 14 | 80.8d | 50 | 162.5e | |
| S2 | 50.1d | 20 | 22.3c | 9 | 152.1c | 61 | 247.4c | |
| Z-S2 | 59.8c | 22 | 23.4c | 9 | 166.1b | 61 | 273.7b | |
| S1 | 67.0bc | 36 | 30.5b | 16 | 76.7d | 41 | 185.7d | |
| Z-S1 | 70.8b | 37 | 31.6b | 16 | 84.2d | 42 | 199.7d | |
| CK | 93.7a | 28 | 38.4a | 11 | 185.1a | 54 | 340.5a | |
| Z-CK | 92.3a | 27 | 37.2a | 11 | 188.9a | 56 | 340.2a | |
| 2018 | S3 | 64.1cd | 36 | 29.5c | 17 | 67.0h | 38 | 176.8g |
| Z-S3 | 66.2c | 30 | 31.7bc | 14 | 105.0e | 48 | 220.5d | |
| S2 | 73.0b | 28 | 29.6c | 11 | 140.0d | 53 | 265.7c | |
| Z-S2 | 75.9a | 26 | 31.9bc | 11 | 151.0c | 53 | 286.8b | |
| S1 | 61.2e | 34 | 31.5bc | 17 | 75.6g | 42 | 180.5f | |
| Z-S1 | 62.8de | 32 | 30.4c | 16 | 90.5f | 46 | 195.8e | |
| CK | 75.8a | 26 | 33.6ab | 11 | 164.4a | 56 | 296.1a | |
| Z-CK | 77.8a | 26 | 35.5a | 12 | 160.7b | 54 | 297.7a | |
新窗口打开|下载CSV
2.4 生长发育进程
遮阴延缓夏玉米的生长发育进程,雌雄间隔时间延长,S3、S2较CK抽雄期(VT)分别晚5 d和6 d,吐丝期(R1)分别晚6 d和7 d。喷施植酶Q9后,Z-S3、Z-S2的生育进程较各自对照S3、S2提前了1—2 d,雌雄间隔时间缩短了1 d(表5)。Table 5
表5
表5植酶Q9对大田遮阴夏玉米生长发育进程的影响(2018)
Table 5
| 处理 Treatment | 播种 Seeding | 出苗 VE | 拔节期 V6 | 大喇叭口 V12 | 抽雄期 VT | 吐丝期 R1 | 乳熟期 R3 | 成熟期 R6 |
|---|---|---|---|---|---|---|---|---|
| S3 | 6-6 | 6-11 | 7-6 | 7-29 | 8-3 | 8-7 | 9-3 | 10-7 |
| Z-S3 | 6-6 | 6-11 | 7-6 | 7-27 | 8-2 | 8-5 | 9-1 | 10-5 |
| S2 | 6-6 | 6-11 | 7-1 | 7-25 | 8-2 | 8-6 | 8-25 | 9-28 |
| Z-S2 | 6-6 | 6-11 | 7-1 | 7-23 | 8-1 | 8-4 | 8-24 | 9-27 |
| S1 | 6-6 | 6-11 | 7-1 | 7-21 | 7-28 | 7-31 | 8-26 | 9-30 |
| Z-S1 | 6-6 | 6-11 | 7-1 | 7-21 | 7-28 | 7-31 | 8-25 | 9-29 |
| CK | 6-6 | 6-11 | 7-1 | 7-21 | 7-28 | 7-31 | 8-23 | 9-26 |
| Z-CK | 6-6 | 6-11 | 7-1 | 7-21 | 7-28 | 7-31 | 8-23 | 9-26 |
新窗口打开|下载CSV
2.5 植株性状
遮阴后夏玉米株高、穗位高、茎粗降低、倒伏率增加。与CK相比,S3株高、穗位高、茎粗分别平均降低16%、20%、24%,S2分别平均降低13%、10%、16%,S1变化不显著。喷施植酶Q9后,Z-S3、Z-S2的株高、穗位高较各自对照S3、S2显著增加,以2015年为例,Z-S3株高、穗位高较S3分别增加13%、17%,Z-S2较S2分别增加10%、7%;而茎粗变化不明显。除2015年,其他年份喷施植酶Q9后倒伏率降低。植株性状Z-CK较CK没有显著差异(表6)。Table 6
表6
表6植酶Q9对大田遮阴夏玉米植株性状的影响
Table 6
| 年份 Year | 处理 Treatment | 株高 Plant height (cm) | 穗位高 Ear height (cm) | 穗位系数 Ear position coefficient | 第3节间茎粗 Stem diameter of the third node (cm) | 倒伏率 Lodging rate (%) |
|---|---|---|---|---|---|---|
| 2015 | CK | 268ab | 116a | 43 | 2.2a | 3.48 |
| Z-CK | 266b | 117a | 44 | 2.2a | 3.23 | |
| S3 | 220f | 90d | 41 | 1.6c | 5.34 | |
| Z-S3 | 249c | 105c | 42 | 1.7bc | 22.98 | |
| S2 | 224e | 105c | 47 | 1.9b | 2.30 | |
| Z-S2 | 246d | 112b | 45 | 1.9b | 4.20 | |
| S1 | 270a | 117a | 43 | 2.3a | 1.40 | |
| Z-S1 | 271a | 117a | 43 | 2.3a | 2.36 | |
| 2016 | CK | 268b | 118a | 44 | 1.9a | 2.19 |
| Z-CK | 257c | 117a | 46 | 1.9a | 2.07 | |
| S3 | 219f | 90d | 41 | 1.6bc | 8.32 | |
| Z-S3 | 244d | 106c | 43 | 1.7b | 5.31 | |
| S2 | 225e | 105c | 47 | 1.7b | 6.55 | |
| Z-S2 | 245d | 112b | 46 | 1.7b | 5.01 | |
| S1 | 269ab | 119a | 44 | 1.8ab | 2.38 | |
| Z-S1 | 272a | 119a | 44 | 1.9a | 2.15 | |
| CK | 247a | 94a | 38 | 2.1a | 0.62 | |
| Z-CK | 246a | 94a | 38 | 2.1a | 0.68 | |
| S3 | 215f | 82d | 38 | 1.5b | 3.89 | |
| Z-S3 | 221e | 87bc | 39 | 1.7b | 0.69 | |
| 2018 | S2 | 228d | 86c | 38 | 1.6b | 1.80 |
| Z-S2 | 231c | 88b | 38 | 1.7b | 0.93 | |
| S1 | 243b | 95a | 39 | 2.0a | 1.35 | |
| Z-S1 | 244b | 94a | 38 | 2.0a | 0.33 |
新窗口打开|下载CSV
2.6 叶面积指数(LAI)
各处理的LAI都呈单峰曲线变化,遮阴降低了夏玉米的LAI。在V12、VT、R3、R6期,S3较CK分别降低30%、22%、14%、44%,S2较CK分别降低18%、14%、10%、26%;在R3、R6期,S1较CK分别降低12%、28%。喷施植酶Q9后,各遮阴处理LAI有所增加,在V12、VT、R3、R6期,Z-S3较S3分别增加32%、16%、10%、13%,Z-S2较S2分别增加8%、6%、9%、11%;在R3、R6期,Z-S1处理2015年较S1分别增加12%、21%,2018年R6期增加12%,其他年份和时期变化不显著(图2)。图2
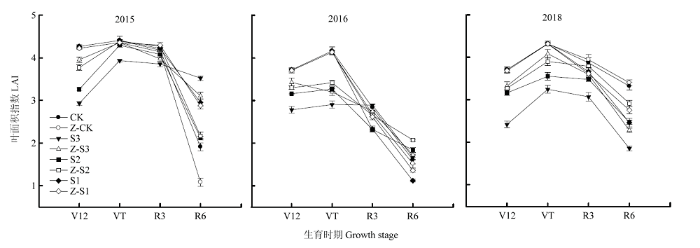 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2植酶Q9对大田遮阴夏玉米叶面积指数(LAI)的影响
Fig. 2Effects of Phytase Q9 on leaf area index (LAI) of summer maize shaded in field
2.7 功能叶片SPAD值
遮阴使夏玉米的功能叶片SPAD值降低(R6期除外),其中S3降幅最高(9%—19%),S2次之(5%—15%),S1降幅最小(0.3%—1%)。喷施植酶Q9后,Z-S3、Z-S2功能叶片SPAD值提高。在V12、VT、R3、R6期,Z-S3较S3分别提高了4%—7%、3%—5%、1.3%—5%、2%—9%;Z-S2在V12、VT、R3期较S2分别提高了1%—8%、2%—6%、3%—5%,差异显著;Z-S1在R3期增加效果不明显。Z-S2、Z-S1在R6期较各自对照的变化复杂,无明显规律(图3)。图3
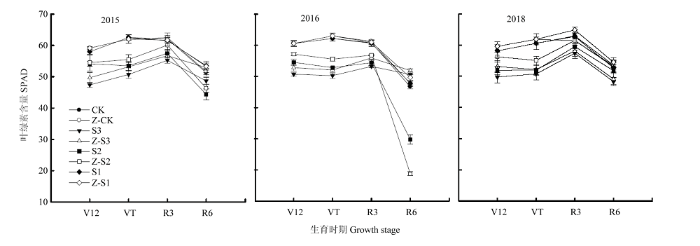 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3植酶Q9对大田遮阴夏玉米功能叶片SPAD值的影响
Fig. 3Effects of Phytase Q9 on SPAD values of functional leaves of summer maize shaded in field
3 讨论
3.1 植酶Q9对大田遮阴夏玉米产量及其构成因素的影响
作物的能量主要来源于光,弱光胁迫影响着作物的生长发育和形态建成,降低作物光合能力,导致产量急剧降低[39,40,41,42]。课题组前期研究结果表明,遮阴显著影响夏玉米光合能力、穗分化、茎秆强度、籽粒灌浆特性,降低产量,且不同生育阶段遮阴减产程度不同,其中开花至收获期遮阴减产幅度最大,拔节至开花期遮阴次之,苗期遮阴最小[5,10,43-44]。通过化学试剂调控夏玉米各器官生长发育和代谢过程,以增强其耐阴性,是提高玉米耐阴能力的可能途径。前人研究发现,玉米喷施矮丰王、增产胺、吨田宝、DA-6、85%乙烯利水剂等均增产,但喷施缩节胺和多效唑减产[38,45-46]。本研究结果表明,遮阴条件下的登海605喷施植酶Q9可显著增产,出苗至收获期遮阴、拔节至开花期遮阴和开花至收获期遮阴分别增产21%、9%和14%。但是,自然光照条件下喷施植酶Q9对夏玉米产量没有显著影响,这可能与植酶Q9的成分及喷施的环境条件有关,其调控机理还需进一步深入系统研究。玉米抗倒伏特性、籽粒中干物质积累、雌雄间隔是影响公顷穗数、千粒重、穗粒数的部分因素。崔海岩等[5]研究表明,遮阴导致玉米株高、穗位高、茎粗、茎秆穿刺强度降低,田间倒伏率升高,公顷穗数降低。李宁等[47]和张倩等[48]研究表明,喷施玉黄金和30%己·乙水剂都能降低玉米株高、穗高系数。株高、穗位高降低,植株重心降低,从而可以降低倒伏率[47,48,49,50]。本研究结果表明,遮阴条件下喷施植酶Q9提高了株高、穗位高,倒伏率却降低。因此,喷施植酶Q9降低倒伏率的原因可能是改善了茎秆皮层发育和显微结构,还需进一步研究。遮阴减少玉米籽粒中胚乳细胞数量,籽粒灌浆速率缓慢,籽粒充实度降低[10,44]。本研究结果表明,遮阴条件下喷施植酶Q9提高了登海605的干物质向籽粒中分配的比例,千粒重增加,即植酶Q9可能对弱光胁迫的玉米籽粒发育和灌浆特性有正调控效果。前人研究表明,遮阴影响穗分化进程,引起花粉量减少,雌雄间隔延长,花丝受精率降低,于是穗粒数减少[5]。本研究结果表明,遮阴条件下的登海605喷施植酶Q9,出苗至收获期遮阴和拔节至开花期遮阴处理的生育进程提前1—2 d,雌雄间隔缩短1 d,穗粒数增加,即大田遮阴喷施夏玉米植酶Q9可能有利于弱光胁迫下的夏玉米穗分化。
3.2 植酶Q9对大田遮阴夏玉米光合特性的影响
叶面积指数、SPAD值是影响夏玉米光合特性的部分因素,干物质积累量可以间接反映夏玉米光合特性。前人研究表明,遮阴显著降低夏玉米的叶面积指数[3,5],喷施增产胺、吨田宝可以提高玉米叶面积指数[46]。叶片SPAD值与叶绿素含量成正相关,叶绿素含量影响着叶片对光能的吸收,遮阴降低水稻叶片的SPAD值[51]。叶面积指数和SPAD值增加可以提升光合性能[3,51]。本研究结果表明,喷施植酶Q9后,出苗至收获期遮阴和拔节至开花期遮阴处理各时期叶面积指数和SPAD值提高,干物质积累量增加,开花至收获期遮阴不同年份叶面积指数和SPAD值表现不同,说明喷施植酶Q9延长了出苗至收获期遮阴和拔节至开花期遮阴处理的叶片功能期,利于提高光合特性,增加干物质积累量。植酶Q9对遮阴条件下夏玉米光合特性的影响还需要进一步研究。4 结论
喷施植酶Q9可以有效缓解夏玉米弱光胁迫导致的危害。喷施植酶Q9后,出苗至收获期遮阴和拔节至开花期遮阴生育进程提前1—2 d,雌雄间隔缩短1 d,叶面积指数、SPAD值、穗位高、株高显著增加;干物质积累及其向籽粒的分配比例增加,倒伏率和空秆率降低;全生育期遮阴的穗部性状改善,产量提高。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
DOI:10.7668/hbnxb.2011.04.016Magsci [本文引用: 1]

大田条件下,研究遮荫处理对玉米不同类型品种的茎秆形态特征及抗倒力学强度的影响。结果表明:稀植大穗品种JK519对弱光胁迫反应较为敏感,遮荫处理后玉米株高和穗位高均显著下降,叶面积指数降低。基部节间缩短、直径变小,单位节间长干质量减少。茎秆穿刺强度在30%和60%遮荫条件下分别下降36.4%和66.0%,田间倒伏率严重。而紧凑耐密品种CS1在遮荫30%胁迫下穗位高、LAI下降不明显;节间缩短,但直径和单位节间长干质量变化不大;穿刺强度仅下降5.9%,田间倒伏较轻;在遮荫60%胁迫下,穿刺强度明显下降,倒伏加重。同时各个品种均随遮荫程度的增加,最终玉米产量、收获穗数、单穗粒数和千粒重明显降低。因此,30%遮荫处理(弱光胁迫)可推荐作为玉米高产抗倒品种茎秆力学强度鉴定和筛选条件之一。
DOI:10.7668/hbnxb.2011.04.016Magsci [本文引用: 1]
大田条件下,研究遮荫处理对玉米不同类型品种的茎秆形态特征及抗倒力学强度的影响。结果表明:稀植大穗品种JK519对弱光胁迫反应较为敏感,遮荫处理后玉米株高和穗位高均显著下降,叶面积指数降低。基部节间缩短、直径变小,单位节间长干质量减少。茎秆穿刺强度在30%和60%遮荫条件下分别下降36.4%和66.0%,田间倒伏率严重。而紧凑耐密品种CS1在遮荫30%胁迫下穗位高、LAI下降不明显;节间缩短,但直径和单位节间长干质量变化不大;穿刺强度仅下降5.9%,田间倒伏较轻;在遮荫60%胁迫下,穿刺强度明显下降,倒伏加重。同时各个品种均随遮荫程度的增加,最终玉米产量、收获穗数、单穗粒数和千粒重明显降低。因此,30%遮荫处理(弱光胁迫)可推荐作为玉米高产抗倒品种茎秆力学强度鉴定和筛选条件之一。
Magsci [本文引用: 3]
<P>在大田条件下研究不同时期和不同程度遮荫对夏玉米光合特性及其产量的影响。结果表明,遮荫对玉米叶片的光合特性有显著影响,导致玉米籽粒产量显著降低,农大108(ND108)苗期遮荫50%和90%的籽粒产量分别比对照降低16.9%和24.5%,穗期遮荫分别降低34.1%和55.3%,花粒期遮荫分别降低67.5%和79.4%。遮荫后玉米的叶面积指数、叶绿素含量和净光合速率等都显著降低。不同生育时期对遮荫的响应不同,苗期遮荫50%和90%处理的叶面积指数分别较对照降低53.6%和64.3%,穗期分别降低22.1%和23.0%,花粒期分别降低66.2%和79.4%。遮荫50%和90%处理叶片的光合速率苗期分别较对照降低28.6%和49.0%,穗期分别降低36.6%和59.6%,花粒期分别降低43.9%和64.7%。遮荫使玉米叶片的磷酸烯醇式丙酮酸羧化酶(PEPCase)和核酮糖二磷酸羧化酶(RuBPCase)活性显著降低,不同遮荫时期对其影响不同,花粒期遮荫影响最显著。遮荫时期对玉米光合特性的影响大于遮荫程度对其影响。</P>
Magsci [本文引用: 3]
<P>在大田条件下研究不同时期和不同程度遮荫对夏玉米光合特性及其产量的影响。结果表明,遮荫对玉米叶片的光合特性有显著影响,导致玉米籽粒产量显著降低,农大108(ND108)苗期遮荫50%和90%的籽粒产量分别比对照降低16.9%和24.5%,穗期遮荫分别降低34.1%和55.3%,花粒期遮荫分别降低67.5%和79.4%。遮荫后玉米的叶面积指数、叶绿素含量和净光合速率等都显著降低。不同生育时期对遮荫的响应不同,苗期遮荫50%和90%处理的叶面积指数分别较对照降低53.6%和64.3%,穗期分别降低22.1%和23.0%,花粒期分别降低66.2%和79.4%。遮荫50%和90%处理叶片的光合速率苗期分别较对照降低28.6%和49.0%,穗期分别降低36.6%和59.6%,花粒期分别降低43.9%和64.7%。遮荫使玉米叶片的磷酸烯醇式丙酮酸羧化酶(PEPCase)和核酮糖二磷酸羧化酶(RuBPCase)活性显著降低,不同遮荫时期对其影响不同,花粒期遮荫影响最显著。遮荫时期对玉米光合特性的影响大于遮荫程度对其影响。</P>
[本文引用: 2]
[本文引用: 2]
[D].
[本文引用: 7]
[D].
[本文引用: 7]
Magsci [本文引用: 1]
以不同基因型玉米为材料,在玉米生长发育的3个主要阶段(苗期、穗期、粒期)进行分期遮光试验,研究不同时期弱光胁迫对不同基因型玉米生长发育和产量的影响。结果表明,遮光延缓了玉米叶片的出生速度,使叶片变薄;遮光可以延缓叶片的衰老,但遮光解除后则加速叶片的衰老;遮光造成植株高度增加,但恢复正常光照后,其株高却逐渐低于对照;遮光使干物质积累下降,抽雄吐丝日期推迟,尤其是吐丝日期推迟更多,并使产量降低,但不同基因型玉米不同遮光处理下降程度不同。试验的4个品种中,掖单2 2和豫玉2号受遮光影响较小,而掖单36 38和丹玉13受影响较大,即不同基因型玉米对弱光胁迫的敏感性不同。
Magsci [本文引用: 1]
以不同基因型玉米为材料,在玉米生长发育的3个主要阶段(苗期、穗期、粒期)进行分期遮光试验,研究不同时期弱光胁迫对不同基因型玉米生长发育和产量的影响。结果表明,遮光延缓了玉米叶片的出生速度,使叶片变薄;遮光可以延缓叶片的衰老,但遮光解除后则加速叶片的衰老;遮光造成植株高度增加,但恢复正常光照后,其株高却逐渐低于对照;遮光使干物质积累下降,抽雄吐丝日期推迟,尤其是吐丝日期推迟更多,并使产量降低,但不同基因型玉米不同遮光处理下降程度不同。试验的4个品种中,掖单2 2和豫玉2号受遮光影响较小,而掖单36 38和丹玉13受影响较大,即不同基因型玉米对弱光胁迫的敏感性不同。
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
该文利用时空数据模型引入时间维,在传统GIS平台上来分析了我国华北地区和华东部分地区40年来(1961年-2000年)温度、降水和日照时数的时空变化规律,以及三者的相互联系。结果表明,华北地区北部温度变化不大,而南部和华东部分地区温度升高明显,10年平均气温大约升高0.5℃/40年;华北地区降水呈东南高西北低的特点,并有明显的年代际变化,华北南部和北部的降水变化存在一定的反相关关系,华北北部降水减少,而东南部降水却呈上升势头;华北地区和华东部分地区的日照时数都有明显的降低。对三者的关系进行了定性分析,华北中北部温度和降水的关系为暖干型,而南部和华东地区为暖湿型;华北中北部日照时数的降低主要是因为大气污染加剧而引起,而华北东南部和华东部分地区是由于降水的增加,即云量的增加和晴天日数的减少而导致日照时数的降低。对温度、降水和日照时数时空变化规律的分析有助于对未来气候变化做出合理的预测,并制定相应的对策。
Magsci [本文引用: 1]
该文利用时空数据模型引入时间维,在传统GIS平台上来分析了我国华北地区和华东部分地区40年来(1961年-2000年)温度、降水和日照时数的时空变化规律,以及三者的相互联系。结果表明,华北地区北部温度变化不大,而南部和华东部分地区温度升高明显,10年平均气温大约升高0.5℃/40年;华北地区降水呈东南高西北低的特点,并有明显的年代际变化,华北南部和北部的降水变化存在一定的反相关关系,华北北部降水减少,而东南部降水却呈上升势头;华北地区和华东部分地区的日照时数都有明显的降低。对三者的关系进行了定性分析,华北中北部温度和降水的关系为暖干型,而南部和华东地区为暖湿型;华北中北部日照时数的降低主要是因为大气污染加剧而引起,而华北东南部和华东部分地区是由于降水的增加,即云量的增加和晴天日数的减少而导致日照时数的降低。对温度、降水和日照时数时空变化规律的分析有助于对未来气候变化做出合理的预测,并制定相应的对策。
DOI:10.11676/qxxb2005.090Magsci [本文引用: 1]
采用国家基准气候站和基本气象站的地面资料,系统地分析了中国大陆地区1951年以来近地表主要气候要素演化的时间和空间特征。结果表明,中国近50 a来年平均地表气温变暖幅度约为1.1 ℃,增温速率接近0.22 ℃/(10 a),比全球或半球同期平均增温速率明显偏高。地表气温增暖主要发生在最近的20余年,其季节和空间特征与前人分析结论基本一致。降水量变化趋势对所取时间段和区域范围敏感。1951年以来全国平均降水量变化趋势不明显,但1956年以来略有增加。降水变化的空间特征明显而相对稳定,东北北部、包括长江中下游的东南部地区和西部广大地区降水增加,而华北地区以及东北东南部和西北东部地区降水明显减少。分析还发现,近50 a来全国平均的日照时数、平均风速、水面蒸发等气候要素均呈显著下降趋势,但积雪地带的最大积雪深度却有所增加。中国日照时间和水面蒸发量变化的空间特征很相似,减少最明显的地区均发生在华北和华东,新疆次之。影响中国年代以上尺度气候变化的因子错综复杂,人类活动引起的大气中温室气体浓度增高可能在一定程度上影响了中国近50 a来的气候,但考虑到尚存的不确定性,目前仍不能给出明确结论。中国东部大部分地区日照时间和水面蒸发量减少可能均起源于人为排放的气溶胶影响,平均风速减弱也有利于水面蒸发量下降,而在西部地区云量和降水量的变化可能更重要。
DOI:10.11676/qxxb2005.090Magsci [本文引用: 1]
采用国家基准气候站和基本气象站的地面资料,系统地分析了中国大陆地区1951年以来近地表主要气候要素演化的时间和空间特征。结果表明,中国近50 a来年平均地表气温变暖幅度约为1.1 ℃,增温速率接近0.22 ℃/(10 a),比全球或半球同期平均增温速率明显偏高。地表气温增暖主要发生在最近的20余年,其季节和空间特征与前人分析结论基本一致。降水量变化趋势对所取时间段和区域范围敏感。1951年以来全国平均降水量变化趋势不明显,但1956年以来略有增加。降水变化的空间特征明显而相对稳定,东北北部、包括长江中下游的东南部地区和西部广大地区降水增加,而华北地区以及东北东南部和西北东部地区降水明显减少。分析还发现,近50 a来全国平均的日照时数、平均风速、水面蒸发等气候要素均呈显著下降趋势,但积雪地带的最大积雪深度却有所增加。中国日照时间和水面蒸发量变化的空间特征很相似,减少最明显的地区均发生在华北和华东,新疆次之。影响中国年代以上尺度气候变化的因子错综复杂,人类活动引起的大气中温室气体浓度增高可能在一定程度上影响了中国近50 a来的气候,但考虑到尚存的不确定性,目前仍不能给出明确结论。中国东部大部分地区日照时间和水面蒸发量减少可能均起源于人为排放的气溶胶影响,平均风速减弱也有利于水面蒸发量下降,而在西部地区云量和降水量的变化可能更重要。
[D].
[本文引用: 3]
[D].
[本文引用: 3]
DOI:10.7668/hbnxb.2010.S1.043Magsci [本文引用: 1]
在大田条件下,研究了氮肥和化学调控对小麦品质的影响。结果表明,氮肥和化控处理对籽粒蛋白组分和加工品质有一定的调节作用。研究表明,与180kg/hm<sup>2</sup>的施氮量相比,施氮270kg/hm<sup>2</sup>显著提高了清蛋白、干湿面筋含量以及面粉吸水率,但增氮肥不利于球蛋白和醇溶蛋白含量的提高;与对照相比,起身期喷施麦巨金对蛋白组分和加工品质有明显的正调节效应,其中对清蛋白、干湿面筋含量和吸水率的影响达显著水平;喷施乙膦水剂有利于粉质参数的改善;喷施多聚氨基酸钾盐对沉降值和吸水率有一定的正调节效应。
DOI:10.7668/hbnxb.2010.S1.043Magsci [本文引用: 1]
在大田条件下,研究了氮肥和化学调控对小麦品质的影响。结果表明,氮肥和化控处理对籽粒蛋白组分和加工品质有一定的调节作用。研究表明,与180kg/hm<sup>2</sup>的施氮量相比,施氮270kg/hm<sup>2</sup>显著提高了清蛋白、干湿面筋含量以及面粉吸水率,但增氮肥不利于球蛋白和醇溶蛋白含量的提高;与对照相比,起身期喷施麦巨金对蛋白组分和加工品质有明显的正调节效应,其中对清蛋白、干湿面筋含量和吸水率的影响达显著水平;喷施乙膦水剂有利于粉质参数的改善;喷施多聚氨基酸钾盐对沉降值和吸水率有一定的正调节效应。
[本文引用: 1]
[本文引用: 1]
DOI:10.11869/j.issn.100-8551.2015.03.0563Magsci [本文引用: 1]
为探讨聚丙烯酰胺(PAM)浸种对谷子抗旱萌发的影响,本研究通过聚丙烯酰胺(PAM)浸种晋谷21种子,用18%的PEG 6000溶液模拟土壤干旱,研究了不同浓度的PAM溶液(0.1%、0.5%、1.0%、5.0%、7.0%、10.0%)浸种对干旱胁迫下谷子种子萌发及幼苗生长中各项生理指标的影响。结果表明:干旱胁迫下的谷子发芽能力显著低于正常水分条件,幼苗中SOD、POD、CAT的活性、Pro、O<sub>2</sub><sup>-</sup>以及MDA的含量均增加,但可溶性蛋白的含量降低; 1.0%~7.0%的PAM溶液浸种可明显促进干旱胁迫下谷子种子的萌发和幼苗生长,有效提高幼苗活性氧清除系统中抗氧化酶的活性,增加渗透调节物质的含量,减少O<sub>2</sub><sup>-</sup>和MDA的积累。综合分析,筛选出用于谷子浸种的最佳PAM浓度为5.0%。结果表明,适当浓度的聚丙烯酰胺浸种可缓解干旱胁迫对谷子萌发和幼苗生长造成的伤害,增强谷子的抗旱性,这为PAM在生产实践中的应用提供了理论依据。
DOI:10.11869/j.issn.100-8551.2015.03.0563Magsci [本文引用: 1]
为探讨聚丙烯酰胺(PAM)浸种对谷子抗旱萌发的影响,本研究通过聚丙烯酰胺(PAM)浸种晋谷21种子,用18%的PEG 6000溶液模拟土壤干旱,研究了不同浓度的PAM溶液(0.1%、0.5%、1.0%、5.0%、7.0%、10.0%)浸种对干旱胁迫下谷子种子萌发及幼苗生长中各项生理指标的影响。结果表明:干旱胁迫下的谷子发芽能力显著低于正常水分条件,幼苗中SOD、POD、CAT的活性、Pro、O<sub>2</sub><sup>-</sup>以及MDA的含量均增加,但可溶性蛋白的含量降低; 1.0%~7.0%的PAM溶液浸种可明显促进干旱胁迫下谷子种子的萌发和幼苗生长,有效提高幼苗活性氧清除系统中抗氧化酶的活性,增加渗透调节物质的含量,减少O<sub>2</sub><sup>-</sup>和MDA的积累。综合分析,筛选出用于谷子浸种的最佳PAM浓度为5.0%。结果表明,适当浓度的聚丙烯酰胺浸种可缓解干旱胁迫对谷子萌发和幼苗生长造成的伤害,增强谷子的抗旱性,这为PAM在生产实践中的应用提供了理论依据。
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
DOI:10.3724/SP.J.1006.2012.00352Magsci [本文引用: 1]
采用盆栽试验, 以郑单958和丰单3号为材料, 研究了低温胁迫对玉米幼苗光合作用、叶绿素、叶绿素荧光参数和抗氧化酶活性的影响, 以及聚糠萘合剂(PKN)的调控效果。结果表明, 低温胁迫下, 玉米幼苗的光合作用和光系统II光化学最大效率受到抑制; 超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性降低; 过氧化氢、超氧阴离子的产生速率及丙二醛(MDA)含量显著升高。PKN处理提高了低温胁迫下玉米幼苗净光合速率(<em>P</em><sub>n</sub>)、气孔导度(<em>G</em><sub>s</sub>)、光系统II光化学的最大效率(<em>F</em><sub>v</sub>/<em>F</em><sub>m</sub>)、叶绿素含量(Chl a+Chl b)。低温处理7 d, ZDTR和FDTR的<em>P</em><sub>n</sub>、<em>G</em><sub>s</sub>、<em>F</em><sub>v</sub>/<em>F</em><sub>m</sub>、Chl a+Chl b分别比各自的对照提高了88.95%和61.11%、593.33%和1 741.67%、111.50%和145.16%、36.61%和54.03%; PKN处理延缓了SOD、POD、CAT活性的降低, 低温胁迫7 d, PKN处理使郑单958和丰单3的SOD、POD、CAT活性分别比对照高了292.59%和632.98%、295.07%和360.54%、254.55%和265.45%; 同时降低了过氧化氢、超氧阴离子的产生速率及MDA的含量。表明PKN处理有利于提高玉米幼苗的抗冷性。
DOI:10.3724/SP.J.1006.2012.00352Magsci [本文引用: 1]
采用盆栽试验, 以郑单958和丰单3号为材料, 研究了低温胁迫对玉米幼苗光合作用、叶绿素、叶绿素荧光参数和抗氧化酶活性的影响, 以及聚糠萘合剂(PKN)的调控效果。结果表明, 低温胁迫下, 玉米幼苗的光合作用和光系统II光化学最大效率受到抑制; 超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性降低; 过氧化氢、超氧阴离子的产生速率及丙二醛(MDA)含量显著升高。PKN处理提高了低温胁迫下玉米幼苗净光合速率(<em>P</em><sub>n</sub>)、气孔导度(<em>G</em><sub>s</sub>)、光系统II光化学的最大效率(<em>F</em><sub>v</sub>/<em>F</em><sub>m</sub>)、叶绿素含量(Chl a+Chl b)。低温处理7 d, ZDTR和FDTR的<em>P</em><sub>n</sub>、<em>G</em><sub>s</sub>、<em>F</em><sub>v</sub>/<em>F</em><sub>m</sub>、Chl a+Chl b分别比各自的对照提高了88.95%和61.11%、593.33%和1 741.67%、111.50%和145.16%、36.61%和54.03%; PKN处理延缓了SOD、POD、CAT活性的降低, 低温胁迫7 d, PKN处理使郑单958和丰单3的SOD、POD、CAT活性分别比对照高了292.59%和632.98%、295.07%和360.54%、254.55%和265.45%; 同时降低了过氧化氢、超氧阴离子的产生速率及MDA的含量。表明PKN处理有利于提高玉米幼苗的抗冷性。
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[D].
[本文引用: 2]
[D].
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2018.08.012Magsci [本文引用: 1]
【目的】磷是植物生长发育的关键元素之一,缺磷会严重影响作物的产量和质量。黄腐酸作为一种天然有机物,在促进植物生长的同时还能提高植物的抗逆能力。开展黄腐酸对番茄幼苗适应低磷胁迫生理调控作用研究,明确黄腐酸对番茄适应低磷胁迫的生理调控机制,对黄腐酸缓解番茄“遗传学缺磷”的概念和机理做出合理解释。【方法】采用Hogland营养液水培方式,以番茄‘金棚1号’为供试品种,待番茄幼苗长到三叶一心时移到水培盆中缓苗7 d,外源添加不同浓度(0、0.04、0.08、0.12、0.16和0.20 g·L<sup>-1</sup>)黄腐酸,研究其对低磷胁迫(10 μmol·L<sup>-1</sup>)下番茄幼苗生长、根系发育、光合作用性能、磷素吸收和分配、有机酸积累和分泌等生理过程的影响。【结果】低磷胁迫下,随着黄腐酸施用量的增加,番茄幼苗的各项生理指标基本呈现先增后降趋势。当黄腐酸的施用浓度为0.08 g·L<sup>-1</sup>时,显著提高根冠比;增加叶绿素含量,提高叶片光合作用性能;提高番茄不同组织中磷素积累、分配和转运;提高磷转运相关基因(PT1和PHO1)的表达,其中PT1能促进番茄幼苗根系向环境中吸收磷酸盐缓解低磷胁迫;PHO1能促进磷酸盐由根系向茎叶分配从而有效缓解地上部缺磷。黄腐酸能提高根系有机酸(草酸、苹果酸、柠檬酸、琥珀酸和酒石酸)积累,降低根系中无氧呼吸产物乳酸和乙酸含量,促进质子泵基因(HA1)的表达,促进根系泌酸,这种生理代谢变化有利于将环境中难溶性的磷转变为可溶性的磷,并促进植物根系对磷素吸收;增强生长发育相关转录因子(GRAS1)表达,降低叶片花青素积累。【结论】通过低磷胁迫下添加一定浓度的黄腐酸可明显改善番茄幼苗地上部生长及根系发育,可在一定程度上缓解植物的缺磷症状。
DOI:10.3864/j.issn.0578-1752.2018.08.012Magsci [本文引用: 1]
【目的】磷是植物生长发育的关键元素之一,缺磷会严重影响作物的产量和质量。黄腐酸作为一种天然有机物,在促进植物生长的同时还能提高植物的抗逆能力。开展黄腐酸对番茄幼苗适应低磷胁迫生理调控作用研究,明确黄腐酸对番茄适应低磷胁迫的生理调控机制,对黄腐酸缓解番茄“遗传学缺磷”的概念和机理做出合理解释。【方法】采用Hogland营养液水培方式,以番茄‘金棚1号’为供试品种,待番茄幼苗长到三叶一心时移到水培盆中缓苗7 d,外源添加不同浓度(0、0.04、0.08、0.12、0.16和0.20 g·L<sup>-1</sup>)黄腐酸,研究其对低磷胁迫(10 μmol·L<sup>-1</sup>)下番茄幼苗生长、根系发育、光合作用性能、磷素吸收和分配、有机酸积累和分泌等生理过程的影响。【结果】低磷胁迫下,随着黄腐酸施用量的增加,番茄幼苗的各项生理指标基本呈现先增后降趋势。当黄腐酸的施用浓度为0.08 g·L<sup>-1</sup>时,显著提高根冠比;增加叶绿素含量,提高叶片光合作用性能;提高番茄不同组织中磷素积累、分配和转运;提高磷转运相关基因(PT1和PHO1)的表达,其中PT1能促进番茄幼苗根系向环境中吸收磷酸盐缓解低磷胁迫;PHO1能促进磷酸盐由根系向茎叶分配从而有效缓解地上部缺磷。黄腐酸能提高根系有机酸(草酸、苹果酸、柠檬酸、琥珀酸和酒石酸)积累,降低根系中无氧呼吸产物乳酸和乙酸含量,促进质子泵基因(HA1)的表达,促进根系泌酸,这种生理代谢变化有利于将环境中难溶性的磷转变为可溶性的磷,并促进植物根系对磷素吸收;增强生长发育相关转录因子(GRAS1)表达,降低叶片花青素积累。【结论】通过低磷胁迫下添加一定浓度的黄腐酸可明显改善番茄幼苗地上部生长及根系发育,可在一定程度上缓解植物的缺磷症状。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.7668/hbnxb.2014.S1.048Magsci [本文引用: 2]
针对河北省高产夏玉米倒伏问题,以先玉335(XY335)和金海5号(JH5)为试验材料,通过设置6.75万,8.25万,9.75万株/hm<sup>2</sup>这3个密度梯度,研究了化学调控对玉米植株农艺性状、基部节间形态特征、力学性状、化学组分及产量的影响。结果表明,化学调控能显著降低植株高度、穗位、重心高度,缩短节间长度,增加节间粗度、降低长粗比值,显著增加茎秆可溶性糖含量、粗纤维含量和含钾量,同时基部节间硬度、抗折力和植株抗拉力值也显著提高,最终明显降低倒伏率。化控处理明显提高了高密条件下夏玉米产量。
DOI:10.7668/hbnxb.2014.S1.048Magsci [本文引用: 2]
针对河北省高产夏玉米倒伏问题,以先玉335(XY335)和金海5号(JH5)为试验材料,通过设置6.75万,8.25万,9.75万株/hm<sup>2</sup>这3个密度梯度,研究了化学调控对玉米植株农艺性状、基部节间形态特征、力学性状、化学组分及产量的影响。结果表明,化学调控能显著降低植株高度、穗位、重心高度,缩短节间长度,增加节间粗度、降低长粗比值,显著增加茎秆可溶性糖含量、粗纤维含量和含钾量,同时基部节间硬度、抗折力和植株抗拉力值也显著提高,最终明显降低倒伏率。化控处理明显提高了高密条件下夏玉米产量。
Magsci [本文引用: 1]
以耐荫性不同的玉米品种郑单958(ZD958,耐荫性较强)和豫玉22(YY22,耐荫性较弱)为材料,研究了苗期50%遮荫对玉米叶片形态结构和光合特性的影响。结果表明:形态结构上,苗期遮荫处理后,玉米叶片变薄,单位面积叶绿体数目减少,基粒数、基粒厚度和片层数增加,但是YY22的叶绿体大部分发育不良,肿胀呈球形,基粒片层和基质片层出现不同程度的松散;而ZD958大部分叶绿体结构良好,各部分发育基本正常。光合特性上,弱光胁迫使玉米叶片叶绿素含量升高,净光合速率(<em>Pn</em>)、PSⅡ最大光化学量子产量(<em>Fv/Fm</em>)和实际光化学效率(<em>Φ</em><sub>PS</sub><sub>Ⅱ</sub>)降低,胞间CO<sub>2</sub>浓度(<em>Ci</em>)和非光化学猝灭(<em>NPQ</em>)增加,但是YY22的<em>Pn、Fv/Fm</em>和<em>Φ</em><sub>PS</sub><sub>Ⅱ</sub>显著下降,<em>Ci</em>和<em>NPQ</em>显著升高;而ZD958的<em>Fv/Fm</em>和<em>Φ</em><sub>PS</sub><sub>Ⅱ</sub>下降幅度较小,且<em>NPQ</em>增加亦不显著。研究结果提示,弱光胁迫对玉米叶片形态结构和光合特性影响较大,且存在基因型差异,耐荫性较强的品种对弱光环境的适应性较强。
Magsci [本文引用: 1]
以耐荫性不同的玉米品种郑单958(ZD958,耐荫性较强)和豫玉22(YY22,耐荫性较弱)为材料,研究了苗期50%遮荫对玉米叶片形态结构和光合特性的影响。结果表明:形态结构上,苗期遮荫处理后,玉米叶片变薄,单位面积叶绿体数目减少,基粒数、基粒厚度和片层数增加,但是YY22的叶绿体大部分发育不良,肿胀呈球形,基粒片层和基质片层出现不同程度的松散;而ZD958大部分叶绿体结构良好,各部分发育基本正常。光合特性上,弱光胁迫使玉米叶片叶绿素含量升高,净光合速率(<em>Pn</em>)、PSⅡ最大光化学量子产量(<em>Fv/Fm</em>)和实际光化学效率(<em>Φ</em><sub>PS</sub><sub>Ⅱ</sub>)降低,胞间CO<sub>2</sub>浓度(<em>Ci</em>)和非光化学猝灭(<em>NPQ</em>)增加,但是YY22的<em>Pn、Fv/Fm</em>和<em>Φ</em><sub>PS</sub><sub>Ⅱ</sub>显著下降,<em>Ci</em>和<em>NPQ</em>显著升高;而ZD958的<em>Fv/Fm</em>和<em>Φ</em><sub>PS</sub><sub>Ⅱ</sub>下降幅度较小,且<em>NPQ</em>增加亦不显著。研究结果提示,弱光胁迫对玉米叶片形态结构和光合特性影响较大,且存在基因型差异,耐荫性较强的品种对弱光环境的适应性较强。
Magsci [本文引用: 1]
在田间池栽条件下,分别于小麦(品种:`济南17' 和 `鲁麦21')(Triticum aestivumcv. `Jinan17' and `Lumai21') 灌浆的前期(开花后1~10 d), 中期(11~20 d)和 后期(21~30 d)进行了遮去50%光合有效辐射的试验,研究了产量和品质的变化及其生理 原因。主要结论如下:1)弱光条件下,光合物质生产均受到严重抑制,产量下降,容重降 低;植株的氮素积累量减少、向子粒分配的比例低,但子粒蛋白质含量、湿面筋含量升高,其中,子粒灌浆前期遮光升高的幅度最大。2)遮光后小麦子粒麦谷蛋白和醇溶蛋白含量均升高,但麦谷蛋白升高的幅度大于醇溶蛋白,使麦谷蛋白与醇溶蛋白的比例升高,麦谷蛋白大聚合体(GMP)含量 也升高,粉质仪参数也显著提高;子粒灌浆前期或中期遮光对上述指标的影响则较小,子粒 品质的形成与灌浆后期的光照条件关系更为密切。3)灌浆期相对较弱的光照强度对改善品质有利,但以降低产量为代价,两个品种的小麦所表现出的趋势基本一致。
Magsci [本文引用: 1]
在田间池栽条件下,分别于小麦(品种:`济南17' 和 `鲁麦21')(Triticum aestivumcv. `Jinan17' and `Lumai21') 灌浆的前期(开花后1~10 d), 中期(11~20 d)和 后期(21~30 d)进行了遮去50%光合有效辐射的试验,研究了产量和品质的变化及其生理 原因。主要结论如下:1)弱光条件下,光合物质生产均受到严重抑制,产量下降,容重降 低;植株的氮素积累量减少、向子粒分配的比例低,但子粒蛋白质含量、湿面筋含量升高,其中,子粒灌浆前期遮光升高的幅度最大。2)遮光后小麦子粒麦谷蛋白和醇溶蛋白含量均升高,但麦谷蛋白升高的幅度大于醇溶蛋白,使麦谷蛋白与醇溶蛋白的比例升高,麦谷蛋白大聚合体(GMP)含量 也升高,粉质仪参数也显著提高;子粒灌浆前期或中期遮光对上述指标的影响则较小,子粒 品质的形成与灌浆后期的光照条件关系更为密切。3)灌浆期相对较弱的光照强度对改善品质有利,但以降低产量为代价,两个品种的小麦所表现出的趋势基本一致。
Magsci [本文引用: 1]
研究了灌浆期弱光对3个杂交稻组合的籽粒生长及籽粒品质的影响.结果表明,49%遮荫及69%遮荫下,籽粒空粒率升高,籽粒最终生长量、灌浆速率、灌浆相对起始势降低,以弱势粒及全穗籽粒受到的影响更大.遮荫处理后,糙米率、精米率、整精米率、透明度、胶稠度显著或极显著降低,组合间有差异;垩白米率、垩白度显著或极显著地升高,直链
Magsci [本文引用: 1]
研究了灌浆期弱光对3个杂交稻组合的籽粒生长及籽粒品质的影响.结果表明,49%遮荫及69%遮荫下,籽粒空粒率升高,籽粒最终生长量、灌浆速率、灌浆相对起始势降低,以弱势粒及全穗籽粒受到的影响更大.遮荫处理后,糙米率、精米率、整精米率、透明度、胶稠度显著或极显著降低,组合间有差异;垩白米率、垩白度显著或极显著地升高,直链
DOI:10.3864/j.issn.0578-1752.2016.11.004Magsci [本文引用: 1]
【目的】研究在不同生育时期遮阴处理下,不同大豆品种植株叶片叶面积、比叶重、叶绿素含量、光合参数、叶绿素荧光参数和产量构成因素的变化规律,为中国南方丘陵和山区大面积推广的玉米大豆间套作种植技术提供理论依据。【方法】采用透光率<span>50%的遮阳网对生育期和生育时期总天数不同的3个参试大豆品种桂夏2号、南豆12和C103进行遮阴,设置4个处理,分别为不遮阴(CK)、出苗至盛花期遮阴(VER2)、出苗至鼓粒期遮阴(VER5)和盛花期至完熟期遮阴(R2R8),测定叶面积、比叶重、叶绿素含量、光合参数、荧光参数和产量构成因素。【结果】VER2处理下大豆品种叶面积、叶绿素(a+b)含量和表观量子效率分别比对照高15.50%、12.95%和74.13%,比叶重、光补偿点和最大光合速率分别比对照低15.78%、26.16%和26.52%,R2R8处理下大豆品种叶面积、叶绿素(a+b)和表观量子效率平均分别比对照高0.3%、10.53%和28.07%,比叶重、光补偿点和最大光合速率平均分别比对照低10.15%、20.34%和12.13%;盛花期复光PS</span>Ⅱ最大量子产量平均比对照低<span>3.01%,非光化学荧光淬灭系数平均比对照高26.80%,鼓粒期复光PS</span>Ⅱ最大量子产量平均比对照低<span>8.47%,非光化学荧光淬灭系数平均比对照高40.79%;VER2、VER5和R2R8处理下,桂夏2号单株粒重分别比对照低40.84%、48.67%和59.16%,百粒重分别比对照低23.69%、39.31%和26.39%,南豆12单株粒重分别比对照低46.67%、54.16%和21.19%,百粒重分别比对照低3.91%、19.93%和26.14%,C103单株粒重分别比对照低69.8%、74.85%和73.89%,百粒重分别比对照低68.8%、69.55%和71.64%。【结论】出苗至盛花期遮阴对参试大豆品种叶片光合及叶绿素荧光特性的影响大于盛花期至完熟期遮阴,大豆植株遮阴</span>后在盛花期复光,叶片光合能力有一定程度的恢复,在鼓粒期复光,则表现为受到强光胁迫,因此,从减小遮阴对大豆叶片光合作用不良影响的角度考虑,在中国南方丘陵和山区玉米大豆间作优于玉米大豆套作,在玉米大豆套作模式下选择品种、播期及种植技术时,应确保大豆在盛花期前恢复光照,避免遮阴超过大豆鼓粒期。前期遮阴和后期遮阴对大豆产量的影响大小因大豆品种而异,但遮阴时间越长对大豆产量构成影响越大。
DOI:10.3864/j.issn.0578-1752.2016.11.004Magsci [本文引用: 1]
【目的】研究在不同生育时期遮阴处理下,不同大豆品种植株叶片叶面积、比叶重、叶绿素含量、光合参数、叶绿素荧光参数和产量构成因素的变化规律,为中国南方丘陵和山区大面积推广的玉米大豆间套作种植技术提供理论依据。【方法】采用透光率<span>50%的遮阳网对生育期和生育时期总天数不同的3个参试大豆品种桂夏2号、南豆12和C103进行遮阴,设置4个处理,分别为不遮阴(CK)、出苗至盛花期遮阴(VER2)、出苗至鼓粒期遮阴(VER5)和盛花期至完熟期遮阴(R2R8),测定叶面积、比叶重、叶绿素含量、光合参数、荧光参数和产量构成因素。【结果】VER2处理下大豆品种叶面积、叶绿素(a+b)含量和表观量子效率分别比对照高15.50%、12.95%和74.13%,比叶重、光补偿点和最大光合速率分别比对照低15.78%、26.16%和26.52%,R2R8处理下大豆品种叶面积、叶绿素(a+b)和表观量子效率平均分别比对照高0.3%、10.53%和28.07%,比叶重、光补偿点和最大光合速率平均分别比对照低10.15%、20.34%和12.13%;盛花期复光PS</span>Ⅱ最大量子产量平均比对照低<span>3.01%,非光化学荧光淬灭系数平均比对照高26.80%,鼓粒期复光PS</span>Ⅱ最大量子产量平均比对照低<span>8.47%,非光化学荧光淬灭系数平均比对照高40.79%;VER2、VER5和R2R8处理下,桂夏2号单株粒重分别比对照低40.84%、48.67%和59.16%,百粒重分别比对照低23.69%、39.31%和26.39%,南豆12单株粒重分别比对照低46.67%、54.16%和21.19%,百粒重分别比对照低3.91%、19.93%和26.14%,C103单株粒重分别比对照低69.8%、74.85%和73.89%,百粒重分别比对照低68.8%、69.55%和71.64%。【结论】出苗至盛花期遮阴对参试大豆品种叶片光合及叶绿素荧光特性的影响大于盛花期至完熟期遮阴,大豆植株遮阴</span>后在盛花期复光,叶片光合能力有一定程度的恢复,在鼓粒期复光,则表现为受到强光胁迫,因此,从减小遮阴对大豆叶片光合作用不良影响的角度考虑,在中国南方丘陵和山区玉米大豆间作优于玉米大豆套作,在玉米大豆套作模式下选择品种、播期及种植技术时,应确保大豆在盛花期前恢复光照,避免遮阴超过大豆鼓粒期。前期遮阴和后期遮阴对大豆产量的影响大小因大豆品种而异,但遮阴时间越长对大豆产量构成影响越大。
Magsci [本文引用: 1]
<P><FONT face=Verdana>在大田条件下研究了不同时期和不同程度遮荫对夏玉米产量及生长发育的影响.结果表</FONT><FONT face=Verdana>明,遮荫显著降低玉米产量.不同时期遮荫对其影响不同,花粒期(从开花到成熟期</FONT><FONT face=Verdana>)遮荫的影响最显著,农大108(ND108)和掖单13号(YD13)遮荫50%、90%处理分别减产67.</FONT><FONT face=Verdana>5%、79.4%和82.9%和86.7%,其次是穗期(从拔节到开花期)遮荫,ND108和YD13分别</FONT><FONT face=Verdana>减产34.1%、55.3%和47.2%、65.7%,而苗期(从出苗到拔节期)遮荫对其影响相对较小</FONT><FONT face=Verdana>,ND108和YD13分别减产16.9%、24.5%和18.9%、24.3%.遮荫对YD13产量的影响大于ND10</FONT><FONT face=Verdana>8.遮荫时期对玉米产量的影响显著地大于遮荫程度.遮荫后两个玉米品种的生育进程都延迟</FONT><FONT face=Verdana>,并随着遮荫程度的增加,对其影响加剧.穗期遮荫显著影响玉米的穗分化,花丝数和雄穗</FONT><FONT face=Verdana>分枝数显著降低,对YD13的影响大于ND108.苗期和穗期遮荫显著抑制玉米叶面积、株高和茎</FONT><FONT face=Verdana>节的生长.</FONT></P>
Magsci [本文引用: 1]
<P><FONT face=Verdana>在大田条件下研究了不同时期和不同程度遮荫对夏玉米产量及生长发育的影响.结果表</FONT><FONT face=Verdana>明,遮荫显著降低玉米产量.不同时期遮荫对其影响不同,花粒期(从开花到成熟期</FONT><FONT face=Verdana>)遮荫的影响最显著,农大108(ND108)和掖单13号(YD13)遮荫50%、90%处理分别减产67.</FONT><FONT face=Verdana>5%、79.4%和82.9%和86.7%,其次是穗期(从拔节到开花期)遮荫,ND108和YD13分别</FONT><FONT face=Verdana>减产34.1%、55.3%和47.2%、65.7%,而苗期(从出苗到拔节期)遮荫对其影响相对较小</FONT><FONT face=Verdana>,ND108和YD13分别减产16.9%、24.5%和18.9%、24.3%.遮荫对YD13产量的影响大于ND10</FONT><FONT face=Verdana>8.遮荫时期对玉米产量的影响显著地大于遮荫程度.遮荫后两个玉米品种的生育进程都延迟</FONT><FONT face=Verdana>,并随着遮荫程度的增加,对其影响加剧.穗期遮荫显著影响玉米的穗分化,花丝数和雄穗</FONT><FONT face=Verdana>分枝数显著降低,对YD13的影响大于ND108.苗期和穗期遮荫显著抑制玉米叶面积、株高和茎</FONT><FONT face=Verdana>节的生长.</FONT></P>
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
DOI:10.11674/zwyf.2014.0308Magsci [本文引用: 2]
<p>SPAD(soil-plant analysis and development)计是一种快速、 方便、 非破坏性的诊断植物叶片相对叶绿素或氮含量的仪器,与传统的氮营养诊断方法相比,此仪器节省时间、 劳力和资源。本试验通过未遮荫和遮荫的方法观察水稻冠层叶片SPAD值、 叶绿素含量、 叶绿素荧光、 光合参数、 叶片厚度和比叶重(叶片干重除以叶片的面积)等生理形态指标的变化,建立SPAD值与光系统II(PSII)最大量子产量(Fv/Fm)之间的回归关系。结果表明,遮荫条件下,甬优9号(YY9)和丙9363(B9363)冠层叶片变薄、 SPAD值、 叶绿素a/b、 比叶重、 电子传递速率(ETR)降低,但快速光曲线的初始斜率无明显变化; 同时, 遮荫导致了叶片的呼吸速率、 最大净光合(Pmax)、 量子效率、 光补偿点和饱和点降低,表明水稻叶片为适应弱光环境, 降低光合能力、 减少呼吸消耗, 以增加对有限光能的利用。不同光照条件下,水稻冠层叶片SPAD值与PSII的Fv/Fm的回归方程呈指数式关系(YY9 R<sup>2</sup>=0.896; B9363 R<sup>2</sup>=0.833), 表明SPAD计可以快速、 无损、 有效地评估水稻冠层叶片的光合作用进程,当SPAD值小于35时,其光合过程可能处于受损状态。</p>
DOI:10.11674/zwyf.2014.0308Magsci [本文引用: 2]
<p>SPAD(soil-plant analysis and development)计是一种快速、 方便、 非破坏性的诊断植物叶片相对叶绿素或氮含量的仪器,与传统的氮营养诊断方法相比,此仪器节省时间、 劳力和资源。本试验通过未遮荫和遮荫的方法观察水稻冠层叶片SPAD值、 叶绿素含量、 叶绿素荧光、 光合参数、 叶片厚度和比叶重(叶片干重除以叶片的面积)等生理形态指标的变化,建立SPAD值与光系统II(PSII)最大量子产量(Fv/Fm)之间的回归关系。结果表明,遮荫条件下,甬优9号(YY9)和丙9363(B9363)冠层叶片变薄、 SPAD值、 叶绿素a/b、 比叶重、 电子传递速率(ETR)降低,但快速光曲线的初始斜率无明显变化; 同时, 遮荫导致了叶片的呼吸速率、 最大净光合(Pmax)、 量子效率、 光补偿点和饱和点降低,表明水稻叶片为适应弱光环境, 降低光合能力、 减少呼吸消耗, 以增加对有限光能的利用。不同光照条件下,水稻冠层叶片SPAD值与PSII的Fv/Fm的回归方程呈指数式关系(YY9 R<sup>2</sup>=0.896; B9363 R<sup>2</sup>=0.833), 表明SPAD计可以快速、 无损、 有效地评估水稻冠层叶片的光合作用进程,当SPAD值小于35时,其光合过程可能处于受损状态。</p>

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}