 ,3, 杜远鹏,1
,3, 杜远鹏,1Effects of Different Light Intensity and Temperature on PSII Photochemical Activity in ‘Cabernet Sauvignon’ Grape Leaves Under Ozone Stress
GENG QingWei1,2, XING Hao1, ZHAI Heng1, JIANG EnShun,3, DU YuanPeng,1通讯作者:
收稿日期:2018-07-9接受日期:2019-01-3网络出版日期:2019-04-01
| 基金资助: |
Received:2018-07-9Accepted:2019-01-3Online:2019-04-01
作者简介 About authors
耿庆伟,E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (417KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
耿庆伟, 邢浩, 翟衡, 蒋恩顺, 杜远鹏. 臭氧胁迫下不同光强与温度处理对‘赤霞珠’葡萄叶片PSII光化学活性的影响[J]. 中国农业科学, 2019, 52(7): 1183-1191 doi:10.3864/j.issn.0578-1752.2019.07.006
GENG QingWei, XING Hao, ZHAI Heng, JIANG EnShun, DU YuanPeng.
0 引言
【研究意义】近几十年来O3污染已经成为对流层中首要污染物,对作物生长和农业生产造成很大影响,全球近1/4的国家或地区面临O3浓度超出60 nL·L-1的威胁[1,2]。由于人类活动和生物源排放,O3浓度仍以每年约0.5%—2.5%的速率增长[3],预计2015—2050年全球地表O3浓度将在现有基础上增加20%—25%,21世纪末将增加40%—60%。O3具有强氧化性,可对植物产生氧化伤害[4]。臭氧污染往往发生在特定的时间段内,不同季节以夏季最强,一天之中又以10:00—14:00最强,臭氧污染与强光和高温同时存在,3个生态逆境因素存在协同胁迫作用。因此,本研究开展3个生态逆境因素协同胁迫对葡萄光化学活性的影响,旨在为生产上采取相应措施缓解臭氧伤害提供理论依据。【前人研究进展】气孔是植物与外界进行气体交换的主要通道,是臭氧进入植物体的主要途径[5],O3通过气孔进入叶片后导致叶片光下最大光化学效率(Φ PSII)和PSII开放程度(qP)受到抑制[6],孙永江等[7]研究发现,随着O3浓度升高,叶片净光合速率(Pn)、气孔导度(Gs)、PSII的实际光化学效率(Φ PSII)、光下最大光化学效率(Fv′/Fm′)、捕获光能用于QA以后的电子传递的概率(ΨEo)、单位面积有活性反应中心的数量(RC/CSm)均呈下降趋势;在低O3浓度条件下(25—55 nL?L-1),Pn的下降主要是由于气孔限制引起的,O3浓度达到并超过80 nL·L-1时,叶绿素含量显著降低,非气孔限制占主导因素,葡萄叶片可以在短时间内(8 h)忍受一定浓度(不超过55 nL·L-1)的O3胁迫,O3浓度达到或超过80 nL·L-1时,PSII功能受到较严重的抑制[7],严重影响植物的蒸腾、光合、呼吸等重要生理过程[8,9]。光是植物进行光合作用所必需的环境因子,但在强光下,当光合机构吸收的光能超过光合作用的利用量时,即出现光抑制,若植物不能及时有效地利用或耗散,就会产生过量的活性氧[10],破坏光合色素和Dl蛋白等,引起PSⅡ的损伤[11,12]。高温胁迫会破坏叶片PSⅡ,导致植物叶片不能有效利用吸收的光能[13,14],最终会降低植物的光合作用、损坏细胞膜并打破活性氧平衡[15,16]。【本研究切入点】自然条件下,夏季强光和高温往往同时出现,伴随着臭氧的污染,植物遭受臭氧、强光、高温等3种逆境因子的交叉胁迫。但目前研究多从单一或两个胁迫因素开展研究,尚缺乏3个生态逆境因素协同胁迫影响机制研究。【拟解决的关键问题】以广泛栽培的酿酒葡萄品种‘赤霞珠’为试材,利用叶绿素荧光技术,在对植物叶片不造成伤害的前提下,探究光合机构的运转状况,探测和分析植物的光合功能,了解臭氧胁迫下强光、高温胁迫对葡萄叶片PSⅡ活性的影响,以期探寻减轻光抑制和提高逆境光合能力的有效途径,保证大田葡萄植株的生长和果实发育。1 材料与方法
1.1 试验时间、地点
本试验于2016年7月—9月在山东农业大学葡萄核心试验园内进行。1.2 试验材料和处理
试材为长势一致的‘赤霞珠’(Vitis vinifera L. Cabernet Sauvigon)一年生扦插苗,种植于直径25 cm、高35 cm的花盆中。基质:沙=1:1。待植株长至10—12片完全展开叶时进行试验处理。处理分为5组,分别在自然大气臭氧浓度条件下正常光照与适温(35 nL?L-1,800 μmol·m-2·s-1,26℃)、臭氧胁迫下正常光照与适温((120±20)nL?L-1,800 μmol·m-2·s-1,26℃)、臭氧胁迫下正常光照与高温((120± 20)nL?L-1,800 μmol·m-2·s-1,40℃)、臭氧胁迫下强光与适温((120±20)nL?L-1,1 600 μmol·m-2·s-1,26℃)和臭氧胁迫下强光与高温(120±20 nL?L-1,1 600 μmol·m-2·s-1,40℃)条件下处理3 h(5个处理分别表示为对照Control、T1、T2、T3、T4),臭氧浓度的选定依据前期试验确定[17]。处理结束后进行相关参数测定,每个处理重复5次,单株小区。
用于O3熏蒸的OTC[17]的框架由直径3 cm的镀锌钢管构成,四周用温室专用的聚乙烯塑料膜包被,顶部与大气相通。OTC内距离试验材料顶部50 cm有布气管,布气管上有一排斜向下与水平面呈45°的出气孔,O3发生器(WJ-HY5型)产生的O3在管道中与空气混合,在鼓风机(SF2-2型)的作用下被通入OTC内布气管。OTC内的O3浓度监测仪(DR70C-臭氧型)可以实时监测OTC内的O3浓度,并把数据传输到电脑上用于观察、保存。通过转子流量计调节O3发生器的进氧量,从而控制OTC内的O3浓度。在OTC气室内通过温度控制系统(Plant System,上海菲探仪器科技有限公司)调节叶片周围温度,用LED冷光源灯控制达到叶片表面光强。
1.3 测定项目和方法
采用英国Hansatech公司的FMS-2型便携脉冲调制式荧光仪测定荧光参数,测定程序如下:首先对叶片进行30 min暗适应,用饱和脉冲光(12 000 μmol·m-2·s-1)测定暗适应30 min后最大荧光(Fm)。对光适应下的叶片先用60 s作用光(1 000 μmol·m-2·s-1),然后用极弱的测量光(<0.05 μmol·m-2·s-1)测得叶片最小荧光(Fo'),再用饱和脉冲光(12 000 μmol·m-2·s-1)测定光适应下的最大荧光值Fm',打开作用光测定光下稳态荧光Ft。包括:天线转化效率:Fv'/Fm'=(Fm'-Fo')/Fm';线性电子传递速率:ETR=(ФPSII)×PFD×0.84× 0.5,其中PFD是被吸收的光通量密度(μmol·m-2·s-1),0.5代表光能在两个光系统间的分配系数,0.84指入射到叶片表面的光能平均有84%被叶片吸收;光化学淬灭:qP=(Fm'-Fs)/(Fm'-Fo'),其中Fs表示光适应下的稳态荧光。两个光系统之间的激发能分配系数按BRAUN等[18]的公式计算:光系统Ⅰ激发能分配系数α=f/(1+f),光系统Ⅱ激发能分配系数β=1/(1+f),f=(Fm'-Fs)/(Fm'-Fo');光系统Ⅰ和Ⅱ间激发能分配的不平衡性可用β/α-1表示。根据DEMMIG-ADAMS等[19]提出的公式计算光合机构吸收的光能用于光化学反应的份额P=Fv'/Fm'×qP,用于天线热耗散的份额D=1-Fv'/Fm',用于反应中心由非光化学反应耗散的份额(过剩光能,X耗散)Ex=Fv'/Fm'×(1-qP)。用连续激发式荧光仪(Handy PEA,Hansatech,英国)测量荧光诱导动力学曲线,参考李鹏民等[20]的计算方法,获得如下参数:PSII最大光化学效率Fv/Fm,PSII的潜在活性Fv/Fo,单位面积有活性反应中心数目(RC/CSm)=Fm·φPo·(Vj/Mo),Vj表示j点相对可变荧光,Mo表示QA被还原的最大速率。捕获的激子将电子传递到QA以后的其他电子受体的概率(Ψo)=ETo/TRo=(1-Vj),ETo/CSo表示用于电子传递的能量,TRo/CSo表示还原QA的激发能。
1.4 数据处理及统计分析
采用Microsoft Excel软件处理数据和制图,采用DPS软件的LSD法进行差异显著性检验。2 结果
2.1 对葡萄叶片Fv/Fm、Fv/F0、RC/CSm、Ψ0的影响
Fv/Fm是PSⅡ最大光化学量子产量,或称最大PSⅡ的光能转换效率,代表了光合机构把吸收的光能用于光化学反应的最大效率,反映光能吸收转化机构的完整性,非胁迫条件下该参数的变化极小,不受物种和生长条件的影响,常用Fv/Fm下降程度来反映光抑制程度的大小[21,22]。从图1可以看出,与对照相比,不同处理显著降低叶片的Fv/Fm值,与T1相比,T2、T3分别降低了10.3%和38.8% ,T4降幅最大,达到54.8%。Fv/Fo表示的是PSII的潜在活性,与T1相比,T2、T3和T4分别降低了和30.4%、69.6%和80.3%。RC/CSm反映了叶片单位面积内有活性反应中心的数量,Ψo表示捕获的激子将电子传递到QA以后的其他电子受体的概率。与对照相比,不同处理均降低了叶片的RC/CSm值和Ψo值,与T1相比,T2、T3和T4处理分别降低了26.8%、68.4%、70.2%和11.2%、21.6%、40.8%。表明臭氧胁迫下,1 600 μmol·m-2·s-1的强光和40℃高温均可加剧PSII最大光化学效率和潜在活性的降低,但1 600 μmol·m-2·s-1的强光影响程度更大;当强光和高温同时出现时,抑制程度最大。图1
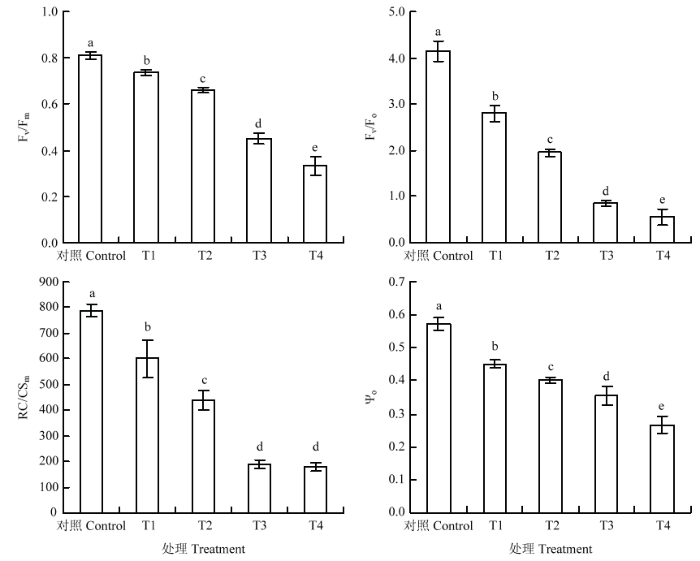 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1臭氧胁迫下不同光强与温度处理对‘赤霞珠’葡萄叶片Fv/Fm、Fv/Fo、RC/CSm、Ψo的影响
不同小写字母表示不同处理在0.05水平上的差异显著性。下同
Fig. 1Effects of light and temperature on Fv/Fm, Fv/Fo, RC/CSm and Ψo in ‘Cabernet Sauvignon’ grape leaf under ozone stress
Different small letters show significant difference between different treatments at 0.05 level. The same as below
2.2 对葡萄叶片Fv'/Fm'、ETR、qp的影响
Fv'/Fm'是PSⅡ有效光化学量子产量,为光条件下 PSⅡ天线转化效率,可以用来反映开放的PSII反应中心的激发能捕获效率[23]。从图2可以看出,与对照相比,T1处理降低了植物叶片实际光化学效率,与T1相比,T2、T3分别降低了7.9%、22.1%,T4下降最显著,降幅为42.3%。ETR可以表示从PSⅡ到PSⅠ的电子传递情况[24],与对照相比,T1处理显著降低了植物叶片线性电子传递速率,与T1相比,T4处理降幅最大,达到62.5%,T2和T3处理分别降低了17.6%和37.5%。qP表示荧光淬灭中的光化学淬灭,从图2可知,与对照相比,不同处理显著降低了葡萄叶片的光化学淬灭,与T1相比,T2、T3的qP值分别降低了10.7%和19.8%,T4变化最显著,降低了39.5%。这表明臭氧胁迫下1 600 μmol·m-2·s-1的强光和40℃高温均可降低PSII激发能捕获效率、线性电子传递速率和光化学淬灭,导致叶片吸收的光能更多的以热耗散形式消耗,被用来进行碳固定的能量显著减少,但1 600 μmol·m-2·s-1的强光影响程度更显著;当强光和高温同时出现时,Fv'/Fm'、ETR和qP降幅最大,影响更显著。图2
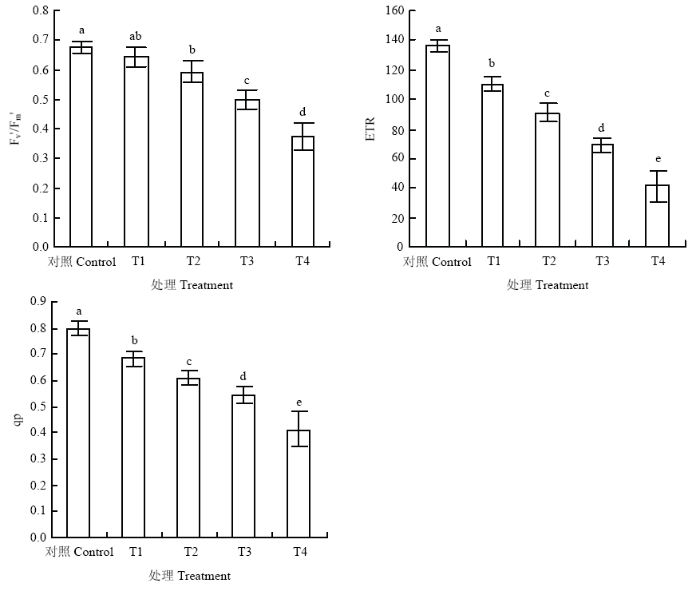 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2臭氧胁迫下不同光强与温度处理对‘赤霞珠’葡萄叶片Fv'/Fm'、ETR、qP的影响
Fig. 2Effect of light and temperature on Fv'/Fm', ETR and qP in ‘Cabernet Sauvignon’ grape leaf under ozone stress
2.3 对葡萄叶片吸收光能分配的影响
植物叶片吸收的光能有三条去路,一是在天线色素上进行热耗散(D),二是用于光化学反应(P),三是反应中心的过剩激发能(E)。图3反映了不同处理下吸收光能向光化学反应、热耗散及过剩光能分配的情况,由图可知,与对照相比,不同处理下吸收的光能用于光化学反应的比例均有所下降,与T1相比,T2、T3分别降低18.8%和38.8%,T4下降最明显,降幅为62.0%。与T1相比,T2、T3和T4处理吸收的光能用于热耗散的比例均增加,增幅分别为15.9%、36.2%和60.5%;过剩光能分别增加12.5%、19.1%和25.2%。这表明臭氧胁迫下1 600 μmol·m-2·s-1的强光和40℃高温均可增加吸收的光能用于热耗散和过剩光能的份额,降低用于光化学反应的比例,但1 600 μmol·m-2·s-1的强光造成过剩光能的份额更大。当强光和高温同时出现时,过剩光能的比例最大,这部分既没有被捕光天线色素耗散掉,也没有被光化学反应利用的过剩光能必然会导致葡萄叶片活性氧含量增加,进而对光合机构造成损伤。图3
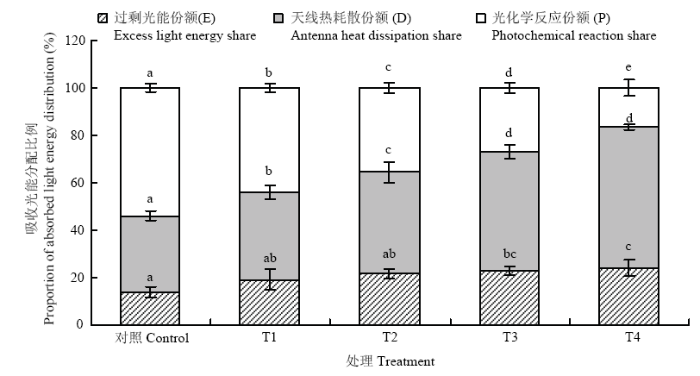 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3臭氧胁迫下不同光强与温度处理对‘赤霞珠’葡萄叶片吸收光能分配的影响
Fig. 3Effects of light and temperature on allocation of absorbed light energy in ‘Cabernet Sauvignon’ grape leaf under ozone stress
2.4 对葡萄叶片光系统I、光系统II间激发能分配的影响
由图4可知,与对照相比,不同处理显著降低叶片PSI激发能分配系数(α)。与T1相比,T2、T3分别下降了6.6%和12.8%,T4下降幅度最大,降幅为25.1%;与之相反,不同处理后叶片PSII激发能分配系数(β)均显著上升,与T1相比,T2、T3、T4分别升高了4.5%、8.7%、17.1%。双光系统间激发能分配不平衡偏离系数(β/α-1)变化趋势与PSII激发能分配系数一致,与T1相比,T2、T3分别升高37.7%和78.3%,T4升高幅度最大,达到187.8%。这表明臭氧胁迫下强光、高温严重影响激发能在两个系统之间的均匀分配,导致光系统之间激发能分配的不平衡,但强光的影响更为严重;当强光、高温同时出现时激发能在两个光系统间分配偏离最严重。图4
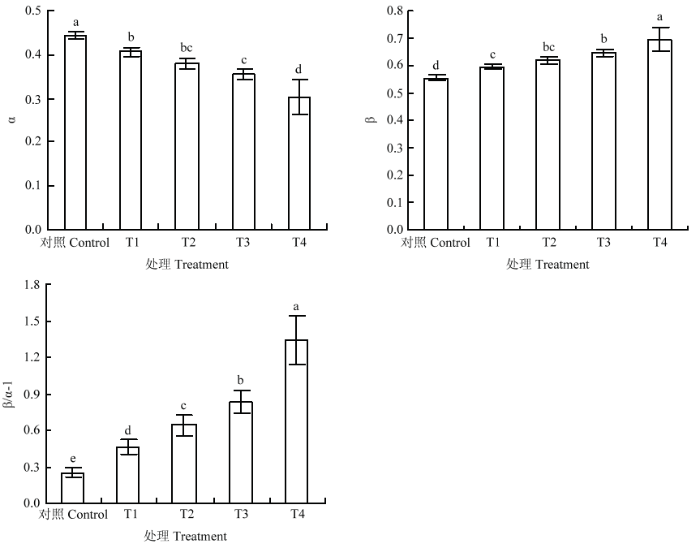 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4臭氧胁迫下不同光强与温度处理对‘赤霞珠’葡萄叶片光系统I、光系统II间激发能分配的影响
Fig. 4Effects of light and temperature on allocation of excitation energy in ‘Cabernet Sauvignon’ grape leaf under ozone stress
3 讨论
快速叶绿素荧光诱导动力学技术被认为是探测植物光合生理的无损探针,植物快速叶绿素荧光诱导曲线中包含着大量有关PSⅡ反应中心原初光化学反应的信息,其参数反映了植物原初反应和光合生理状况,常用来研究各种外界逆境因子对植物的影响[20]。大量研究表明,臭氧、强光和高温等非生物逆境因子跟农业生产和作物产量的有着紧密的联系。臭氧可以诱导植物体产生大量活性氧,造成过高的活性氧(ROS)积累,影响光抑制修复过程,从而导致叶片光抑制发生[25];强光可以使叶片发生光抑制,而且当光抑制达到一定程度时,光合机构中的D1蛋白发生降解,产生光破坏[26];在葡萄生长季节当气温高于40℃时会抑制新梢的生长;而当温度超过40℃时,由于细胞酶系统被钝化,葡萄体内的各种生理代谢活动受到严重阻碍,同时会造成PSII的结构和功能发生变化甚至损害,致使新梢停止生长、叶片发黄、果实日烧,最终导致葡萄减产并影响翌年葡萄生长发育。在自然条件下,特别是夏季晴天,高温往往伴随强光发生,同时高辐射会造成对流层形成大量O3[27],因此,在葡萄生长的旺季—夏季,葡萄叶片通常要遭受高温、强光和O3 3种胁迫因子。本研究中发现臭氧胁迫下葡萄叶片最大光化学效率(Fv/Fm)、单位面积有活性反应中心的数量(RC/CSm)以及捕获光能用于向QA以后的电子传递的能量比例(ΨEo)显著降低,这表明叶片PSII的反应中心和受体侧出现光抑制[28]。光系统I激发能分配系数(α)大幅降低,光系统II激发能分配系数(β)显著增大,两个光系统之间的激发能分配严重偏离平衡。光化学淬灭系数(qP)下降的同时非光化学淬灭系数(NPQ)升高,这表明叶绿素吸收的光能用于光化学反应的比例降低,叶片过剩的光能通过热耗散的形式散失以保护叶片光合结构,虽然叶片热耗散能力增加,但是由于PSII实际光化学效率(ФPSII)的下降,叶片通过光化学反应对激发能的利用率大幅下降,热耗散途径不足以清除大量的过剩光能,最终造成光系统II的伤害,这与前人研究结果一致。
强光和高温均对臭氧胁迫有加剧作用,1 000 μmol·m-2·s-1光强下的臭氧胁迫对菜豆叶片的影响显著大于单一臭氧或光照胁迫处理[29],说明高光强可以加剧臭氧对光合作用的抑制。笔者课题组前期试验通过响应面试验设计不同光照与臭氧组合也发现光照强度和O3浓度存在交互作用,强光不仅可以加剧O3对植物叶片PSII的伤害,并且随着光强的增加伤害程度在逐渐升高[30]。本研究结果发现40℃的高温、1 600 μmol·m-2·s-1的强光均加剧了臭氧胁迫对PSII造成的伤害,但1 600 μmol·m-2·s-1的强光影响程度显著大于40℃的高温;当强光和高温同时存在时,O3胁迫对PSII的伤害最明显,不仅加大了强光因素引起的光系统II伤害,也加大了高温因素引起的光系统II伤害。这是由于强光产生的过剩光能导致ROS的积累,本身加剧了O3对植物的ROS伤害,造成光抑制的出现,加之高温通过伤害细胞质膜和光合电子传递链[31],破坏PSII反应中心的D1蛋白,同时降低暗反应相关的酶活性[32],进一步导致植物叶片Pn降低[33]。虽然叶片通过热耗散的形式散失/疏散过剩光能以保护叶片光合结构,但是ROS和高温对光合机构、细胞质膜和光合电子传递链的氧化伤害,导致叶绿素吸收的光能用于光化学反应的比例大幅降低,大量的过剩光能积聚在光合结构,热耗散途径不足以及时有效清除的情况下过剩光能继续积累,ROS物质继续伤害抗氧化系统和光合机构,并最终造成了PSII的严重伤害。
生产中高温和强光天气往往同时发生,本研究结果提示臭氧胁迫下强光的伤害高于高温,由此提示可以通过降低光强缓解臭氧伤害。叶幕结构与植物光合作用联系密切,成果等[34]比较了新疆地区多主蔓扇形和“厂”字形等方式对‘赤霞珠’葡萄光合特性的影响,研究结果发现不同整形方式改变叶幕光照透射率以及果际周围光合有效辐射,影响叶片光合速率及果实品质。本研究团队前期在田间观察也发现棚架‘摩尔多瓦’葡萄叶片受臭氧伤害症状弱于篱架,由此提示可以通过架式调整或铺设遮阳网降低光强来缓解臭氧伤害。
4 结论
温、光逆境增加了PSII活性对臭氧胁迫的敏感性,40℃的高温、1 600 μmol·m-2·s-1的强光均可以显著降低叶片最大光化学效率(Fv/Fm)、线性电子传递速率(ETR)、单位面积有活性反应中心的数量(RC/CSm)以及光化学淬灭系数(qP),导致两个光系统之间的激发能分配严重偏离平衡,造成光系统II的伤害。1 600 μmol·m-2·s-1的强光影响程度显著大于40℃的高温,而高温、强光胁迫下过剩光能的比例最大,导致葡萄叶片活性氧含量增加,进而对光合机构造成损伤。(责任编辑 赵伶俐)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
URL [本文引用: 1]

采用开顶式气室(open top chambers,OTCs)装置,研究O浓度增加(E-O约150 nL·L)对湿地松幼苗的影响,包括伤害症状、植株生长、针叶光合色素含量、气体交换速率与叶绿素荧光、丙二醛(MDA)以及主要 抗氧化剂含量变化.经过E-O处理一个生长季后(AOT40值为38.24μL·L·h),湿地松幼苗当年生 新叶与老叶呈现出伤害症状,而株高与基径生长、生物量累积与分配未出现显著变化;与对照相比,O,暴露对当年生针叶光合作用具有一定 的抑制作用:净光合速率(P)、气孔导度(g)与蒸腾速率(T)分别显著降低42.5%、 48.2%与46.3%,而胞间CO:浓度(c)与水分利用效率(WUE)、叶绿素荧光指标无显著变化;叶绿素 a、叶绿素b与总叶绿素含量分别降低16.7%、21.1%与17.5%,叶片MDA含量、总还原能力、抗坏血酸与总酚含量均无显著变化.可见,光合色素 含量的降低是E-O,引起湿地松幼苗针叶光合速率下降的主要原因.
URL [本文引用: 1]
采用开顶式气室(open top chambers,OTCs)装置,研究O浓度增加(E-O约150 nL·L)对湿地松幼苗的影响,包括伤害症状、植株生长、针叶光合色素含量、气体交换速率与叶绿素荧光、丙二醛(MDA)以及主要 抗氧化剂含量变化.经过E-O处理一个生长季后(AOT40值为38.24μL·L·h),湿地松幼苗当年生 新叶与老叶呈现出伤害症状,而株高与基径生长、生物量累积与分配未出现显著变化;与对照相比,O,暴露对当年生针叶光合作用具有一定 的抑制作用:净光合速率(P)、气孔导度(g)与蒸腾速率(T)分别显著降低42.5%、 48.2%与46.3%,而胞间CO:浓度(c)与水分利用效率(WUE)、叶绿素荧光指标无显著变化;叶绿素 a、叶绿素b与总叶绿素含量分别降低16.7%、21.1%与17.5%,叶片MDA含量、总还原能力、抗坏血酸与总酚含量均无显著变化.可见,光合色素 含量的降低是E-O,引起湿地松幼苗针叶光合速率下降的主要原因.
URLMagsci [本文引用: 1]
采用开顶式气室(OTC),在大田试验条件下系统研究了地表臭氧浓度增加对冬小麦光合作用的胁迫效应。结果表明,在(100±8)nmol?mol-1的O3处理下,与对照相比,从抽穗期到成熟期冬小麦叶绿素a、叶绿素b和总叶绿素含量均呈先上升后降低的变化趋势,而类胡萝卜素的含量和Chla/b的比值则分别持续升高和下降,净光合速率和气孔导度在抽穗初期无明显下降,但是随着熏蒸时间的增加下降幅逐步变大。在整个试验期间冬小麦表观量子产额、单位面积叶片活性以及PSII最大光量子效率均无明显降低,而蒸腾速率、暗呼吸速率、光补偿点以及最大光合速率却受到较大程度的抑制。当采用(150±8)nmol?mol-1 O3处理时,各生育期中小麦叶片类胡萝卜素含量、暗呼吸速率和光补偿点均显著高于对照组,其余参数均较对照组明显降低。以上结果表明,地表臭氧浓度增加对冬小麦光合作用的影响阈值在100-150 nmol?mol-1之间,且随着熏蒸时间的增加,叶片光合作用受抑制的程度逐步增强。
URLMagsci [本文引用: 1]
采用开顶式气室(OTC),在大田试验条件下系统研究了地表臭氧浓度增加对冬小麦光合作用的胁迫效应。结果表明,在(100±8)nmol?mol-1的O3处理下,与对照相比,从抽穗期到成熟期冬小麦叶绿素a、叶绿素b和总叶绿素含量均呈先上升后降低的变化趋势,而类胡萝卜素的含量和Chla/b的比值则分别持续升高和下降,净光合速率和气孔导度在抽穗初期无明显下降,但是随着熏蒸时间的增加下降幅逐步变大。在整个试验期间冬小麦表观量子产额、单位面积叶片活性以及PSII最大光量子效率均无明显降低,而蒸腾速率、暗呼吸速率、光补偿点以及最大光合速率却受到较大程度的抑制。当采用(150±8)nmol?mol-1 O3处理时,各生育期中小麦叶片类胡萝卜素含量、暗呼吸速率和光补偿点均显著高于对照组,其余参数均较对照组明显降低。以上结果表明,地表臭氧浓度增加对冬小麦光合作用的影响阈值在100-150 nmol?mol-1之间,且随着熏蒸时间的增加,叶片光合作用受抑制的程度逐步增强。
DOI:10.1016/j.envpol.2015.01.016URLPMID:25618365 [本文引用: 1]
61Ozone concentrations are increasing in most of regions of China.61Ozone has caused high yield loss of food crops in China.61More species and local varieties should be investigated for ozone sensitivity.61Developing the air quality standards for crops is required in China.61More air quality stations in the rural are needed.
[本文引用: 1]
DOI:10.1111/j.1365-3040.2008.01923.xURLPMID:19054345 [本文引用: 1]
Experimental investigations of ozone (O 3 ) effects on plants have commonly used short, acute [O 3 ] exposure (>100 ppb, on the order of hours), while in field crops damage is more likely caused by chronic exposure (<100 ppb, on the order of weeks). How different are the O 3 effects induced by these two fumigation regimes? The leaf-level photosynthetic response of soybean to acute [O 3 ] (400 ppb, 6 h) and chronic [O 3 ] (90 ppb, 8 h d 1 , 28 d) was contrasted via simultaneous in vivo measurements of chlorophyll a fluorescence imaging (CFI) and gas exchange. Both exposure regimes lowered leaf photosynthetic CO 2 uptake about 40% and photosystem II (PSII) efficiency ( F q '/ F m ') by 20% compared with controls, but this decrease was far more spatially heterogeneous in the acute treatment. Decline in F q '/ F m ' in the acute treatment resulted equally from decreases in the maximum efficiency of PSII ( F v '/ F m ') and the proportion of open PSII centres ( F q '/ F v '), but in the chronic treatment decline in F q '/ F m ' resulted only from decrease in F q '/ F v '. Findings suggest that acute and chronic [O 3 ] exposures do not induce identical mechanisms of O 3 damage within the leaf, and using one fumigation method alone is not sufficient for understanding the full range of mechanisms of O 3 damage to photosynthetic production in the field.
[本文引用: 2]
探讨O3处理对‘赤霞珠’葡萄叶片光系统功能Ⅱ(PSⅡ)的伤害机制及O3胁迫发生阈值。以一年生‘赤霞珠’为试材,设置不同O3浓度(25、40、55、80和105 n L·L-1),测定叶片叶绿素含量、气体交换和叶绿素荧光参数,分析不同O3浓度处理对葡萄叶片PSII活性的影响。结果表明,随着O3浓度升高,叶片净光合速率(Pn)、气孔导度(Gs)、PSII的实际光化学效率(ΦPSII)、光下最大光化学效率(Fv’/Fm’)、捕获光能用于QA以后的电子传递的概率(ΨEo)、单位面积有活性反应中心的数量(RC/CSm)均呈下降趋势;PSII激发压(1-qP)、K点的相对可变荧光(Wk)、慢相荧光淬灭(qI)呈现上升趋势;在低O3浓度条件下(25~55n L·L-1),Pn的下降主要是由于气孔限制引起的,O3浓度达到并超过80 n L·L-1时,叶绿素含量显著降低,非气孔限制占主导因素,PSII功能开始受到抑制;浓度达到105 n L·L-1时叶片出现较明显的表观伤害症状,电子供体侧伤害程度大于受体侧。实验结果说明,葡萄叶片可以在短时间内(8 h)忍受一定浓度(不超过55 n L·L-1)的O3胁迫,O3浓度达到或超过80 n L·L-1时,PSII功能受到较严重的抑制。
[本文引用: 2]
探讨O3处理对‘赤霞珠’葡萄叶片光系统功能Ⅱ(PSⅡ)的伤害机制及O3胁迫发生阈值。以一年生‘赤霞珠’为试材,设置不同O3浓度(25、40、55、80和105 n L·L-1),测定叶片叶绿素含量、气体交换和叶绿素荧光参数,分析不同O3浓度处理对葡萄叶片PSII活性的影响。结果表明,随着O3浓度升高,叶片净光合速率(Pn)、气孔导度(Gs)、PSII的实际光化学效率(ΦPSII)、光下最大光化学效率(Fv’/Fm’)、捕获光能用于QA以后的电子传递的概率(ΨEo)、单位面积有活性反应中心的数量(RC/CSm)均呈下降趋势;PSII激发压(1-qP)、K点的相对可变荧光(Wk)、慢相荧光淬灭(qI)呈现上升趋势;在低O3浓度条件下(25~55n L·L-1),Pn的下降主要是由于气孔限制引起的,O3浓度达到并超过80 n L·L-1时,叶绿素含量显著降低,非气孔限制占主导因素,PSII功能开始受到抑制;浓度达到105 n L·L-1时叶片出现较明显的表观伤害症状,电子供体侧伤害程度大于受体侧。实验结果说明,葡萄叶片可以在短时间内(8 h)忍受一定浓度(不超过55 n L·L-1)的O3胁迫,O3浓度达到或超过80 n L·L-1时,PSII功能受到较严重的抑制。
URL [本文引用: 1]
为给O3等大气污染物胁迫下我国粮食生产和安全评估提供依据,利用开顶式气室(OTC)开展了3种O3熏蒸水平的大田试验(空气,CK;100nL·L^-1,T1;150nL·L^-1,T2),采用Diving—PAM叶绿素荧光仪和LCpro+光合仪测定了冬小麦(扬麦13)不同生育期的叶绿素荧光及气体交换参数.结果表明,T1的Fv/Fm均高于0.8,Pm、qP、(1-qP)/NPQ及Y(NO)与CK相似,NPQ及Y(NPQ)分别较CK上升13.5%~29.0%和13.3%~22.7%,实际光化学效率在自然光下(快速光曲线,RLC)和暗适应后(诱导曲线达稳态时,IC)分别下降4.6%~7.6%和11.3%~19.3%,Pn与Gs分别下降8.0%~9.8%和11.0%-23.1%,Ls在抽穗期和扬花期高于CK,在灌浆期和成熟期低于CK;T2的Fv/Fm均略低于0.8,Y(NO)、(1-qP)/NPQ及ci分别较CK增加37.9%~75.6%、157.1%~325.8%和3.4%~18.1%,实际光化学效率在RLC和IC条件下分别下降10.2%~13.6%和21.4%~29.1%,Ps、Ls、qP、Pm、NPQ及Y(NPQ)分别下降28.1%~39.9%、5.2%~21.3%、15.8%-30.4%、27.6%~45.6%、33.3%~52.9%和5.7%~17.9%.0,胁迫显著降低了冬小麦的光合能力,其胁迫效应和作用机制在生长季内呈动态变化;T1的Fv/Fm下降是对NPQ增加的响应,Ps与实际光化学效率降低是保护性调节的结果,其抗氧化系统及热耗散机制足以保护PSⅡ反应中心免遭光伤害;T2的CO2同化与QA重新氧化受限,热耗散机制受损,其光合能力下降是光合系统受到o,与过剩光能双重伤害的结果;O3损伤冬小麦PSⅡ的临界值在100—150nL·L^-1之间,接近100nL·L^-1,Fv/Fm难以指示O3对冬小麦的胁迫.尽管冬小麦对高浓度O3具有一定适应能力,地表O3浓度升高仍然是我国粮食生产中面临的一个重要的问题.
URL [本文引用: 1]
为给O3等大气污染物胁迫下我国粮食生产和安全评估提供依据,利用开顶式气室(OTC)开展了3种O3熏蒸水平的大田试验(空气,CK;100nL·L^-1,T1;150nL·L^-1,T2),采用Diving—PAM叶绿素荧光仪和LCpro+光合仪测定了冬小麦(扬麦13)不同生育期的叶绿素荧光及气体交换参数.结果表明,T1的Fv/Fm均高于0.8,Pm、qP、(1-qP)/NPQ及Y(NO)与CK相似,NPQ及Y(NPQ)分别较CK上升13.5%~29.0%和13.3%~22.7%,实际光化学效率在自然光下(快速光曲线,RLC)和暗适应后(诱导曲线达稳态时,IC)分别下降4.6%~7.6%和11.3%~19.3%,Pn与Gs分别下降8.0%~9.8%和11.0%-23.1%,Ls在抽穗期和扬花期高于CK,在灌浆期和成熟期低于CK;T2的Fv/Fm均略低于0.8,Y(NO)、(1-qP)/NPQ及ci分别较CK增加37.9%~75.6%、157.1%~325.8%和3.4%~18.1%,实际光化学效率在RLC和IC条件下分别下降10.2%~13.6%和21.4%~29.1%,Ps、Ls、qP、Pm、NPQ及Y(NPQ)分别下降28.1%~39.9%、5.2%~21.3%、15.8%-30.4%、27.6%~45.6%、33.3%~52.9%和5.7%~17.9%.0,胁迫显著降低了冬小麦的光合能力,其胁迫效应和作用机制在生长季内呈动态变化;T1的Fv/Fm下降是对NPQ增加的响应,Ps与实际光化学效率降低是保护性调节的结果,其抗氧化系统及热耗散机制足以保护PSⅡ反应中心免遭光伤害;T2的CO2同化与QA重新氧化受限,热耗散机制受损,其光合能力下降是光合系统受到o,与过剩光能双重伤害的结果;O3损伤冬小麦PSⅡ的临界值在100—150nL·L^-1之间,接近100nL·L^-1,Fv/Fm难以指示O3对冬小麦的胁迫.尽管冬小麦对高浓度O3具有一定适应能力,地表O3浓度升高仍然是我国粮食生产中面临的一个重要的问题.
DOI:10.3969/j.issn.1674-5906.2009.02.057URL [本文引用: 1]
随着全球气候变化对生态环境的影响日益增加,近地层臭氧(O3)污染的环境生态效应备受人们关注。现有研究表明,陆地生态系统的温室气体NOx和CH4释放、矿质能源消耗和机动车辆尾气排放量的增加将加剧近地层O3污染。O3污染通过降低植物叶片气孔导度、光合速率和净同化作用,改变同化物的分配,进而抑制植物生长和加速植物老化,导致作物和林木减产。O3污染导致植物-土壤系统碳积累和固定降低势必影响未来全球碳动力学和碳预算,而植物和根系生长受到抑制则不利于土壤养分、水分的吸收进而影响植物-土壤系统养分循环,但目前报导极少,尚无法准确判断对全球碳和养分循环的影响,亟待深入研究。由于环境因素间具有互作效应,目前模拟研究过多集中O3与CO2增加对陆地生态系统的复合效应方面,而与其它环境因子(如O3与NOx、SO2、水分、温度等)的复合效应研究偏少,不利于在全球气候变化背景下深入了解与预NO3污染对陆地生态系统的影响程度与趋势。基于研究现状,未来应加强:(1)地表O3监测网络建设和监测,结合田间试验和建模加强草地、森林和农田生态系统对O3污染的响应研究;(2)长期定位研究,侧重陆地生态系统对O3污染连合其它温室气体、温度增加等模拟未来气候情景下的环境响应研究;(3)O3污染下土壤-植物系统碳循环和固定研究;(4)O3污染条件下优势植物和农作物在不同时空条件下的土壤-植物系统养分利用研究;以期为判断和预测全球气候变化背景下陆地生态系统对近地层O3污染加剧的响应程度与趋势提供数据资料和科学依据。
DOI:10.3969/j.issn.1674-5906.2009.02.057URL [本文引用: 1]
随着全球气候变化对生态环境的影响日益增加,近地层臭氧(O3)污染的环境生态效应备受人们关注。现有研究表明,陆地生态系统的温室气体NOx和CH4释放、矿质能源消耗和机动车辆尾气排放量的增加将加剧近地层O3污染。O3污染通过降低植物叶片气孔导度、光合速率和净同化作用,改变同化物的分配,进而抑制植物生长和加速植物老化,导致作物和林木减产。O3污染导致植物-土壤系统碳积累和固定降低势必影响未来全球碳动力学和碳预算,而植物和根系生长受到抑制则不利于土壤养分、水分的吸收进而影响植物-土壤系统养分循环,但目前报导极少,尚无法准确判断对全球碳和养分循环的影响,亟待深入研究。由于环境因素间具有互作效应,目前模拟研究过多集中O3与CO2增加对陆地生态系统的复合效应方面,而与其它环境因子(如O3与NOx、SO2、水分、温度等)的复合效应研究偏少,不利于在全球气候变化背景下深入了解与预NO3污染对陆地生态系统的影响程度与趋势。基于研究现状,未来应加强:(1)地表O3监测网络建设和监测,结合田间试验和建模加强草地、森林和农田生态系统对O3污染的响应研究;(2)长期定位研究,侧重陆地生态系统对O3污染连合其它温室气体、温度增加等模拟未来气候情景下的环境响应研究;(3)O3污染下土壤-植物系统碳循环和固定研究;(4)O3污染条件下优势植物和农作物在不同时空条件下的土壤-植物系统养分利用研究;以期为判断和预测全球气候变化背景下陆地生态系统对近地层O3污染加剧的响应程度与趋势提供数据资料和科学依据。
[本文引用: 1]
DOI:10.1016/j.bbabio.2009.05.005URLPMID:19463778 [本文引用: 1]
Photosysthetic cleavage of water molecules to molecular oxygen is a crucial process for all aerobic life on the Earth. Light-driven oxidation of water occurs in photosystem II (PSII) 鈥 a pigment鈥損rotein complex embedded in the thylakoid membrane of plants, algae and cyanobacteria. Electron transport across the thylakoid membrane terminated by NADPH and ATP formation is inadvertently coupled with the formation of reactive oxygen species (ROS). Reactive oxygen species are mainly produced by photosystem I; however, under certain circumstances, PSII contributes to the overall formation of ROS in the thylakoid membrane. Under limitation of electron transport reaction between both photosystems, photoreduction of molecular oxygen by the reducing side of PSII generates a superoxide anion radical, its dismutation to hydrogen peroxide and the subsequent formation of a hydroxyl radical terminates the overall process of ROS formation on the PSII electron acceptor side. On the PSII electron donor side, partial or complete inhibition of enzymatic activity of the water-splitting manganese complex is coupled with incomplete oxidation of water to hydrogen peroxide. The review points out the mechanistic aspects in the production of ROS on both the electron acceptor and electron donor side of PSII.
DOI:10.1016/B978-0-12-405210-9.00007-2URL [本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2010.13.014URLMagsci [本文引用: 1]
<P><FONT face=Verdana>【目的】研究成熟期‘赤霞珠’葡萄的光合作用在高温胁迫下及恢复期间的变化,探讨葡萄光合作用抑制及恢复的机制。【方法】以‘赤霞珠’葡萄为试材,进行6 d 的高温(每日9:30—15:30 40℃,其余时间30℃)处理, 然后恢复 1 d。对照的温度每日6:30—18:30为30℃,其余时间20℃。恢复时的温度条件同对照的温度。测定高温处理前、处理过程中和恢复后葡萄叶片的气体交换参数和叶绿素荧光参数。【结果】每日40℃的高温处理后,葡萄叶片净光合速率(Pn)、气孔导度(Gs)、PSⅡ的实际光化学效率(ΦPSII)、光化学猝灭系数(qP)和单位面积有活性的反应中心的密度(RC/CSo)显著小于对照,细胞间隙二氧化碳浓度(Ci)、非光化学猝灭系数(NPQ)、光系统Ⅱ受体侧放氧复合体功能参数(Wk)和捕获的激子将电子传递到电子传递链中QA- 下游的其它电子受体的概率(ψEo)升高。经过1 d的恢复,高温处理的这些参数值与对照没有显著差异。【结论】40℃的高温胁迫导致‘赤霞珠’葡萄叶片的Pn显著下降,不过在常温下1 d就能够恢复。Pn下降不是气孔因子导致的结果,主要与PSⅡ供体侧和反应中心活性以及激发能的分配有关;Pn恢复上升,与上述因子都有关系。<BR></FONT></P>
DOI:10.3864/j.issn.0578-1752.2010.13.014URLMagsci [本文引用: 1]
<P><FONT face=Verdana>【目的】研究成熟期‘赤霞珠’葡萄的光合作用在高温胁迫下及恢复期间的变化,探讨葡萄光合作用抑制及恢复的机制。【方法】以‘赤霞珠’葡萄为试材,进行6 d 的高温(每日9:30—15:30 40℃,其余时间30℃)处理, 然后恢复 1 d。对照的温度每日6:30—18:30为30℃,其余时间20℃。恢复时的温度条件同对照的温度。测定高温处理前、处理过程中和恢复后葡萄叶片的气体交换参数和叶绿素荧光参数。【结果】每日40℃的高温处理后,葡萄叶片净光合速率(Pn)、气孔导度(Gs)、PSⅡ的实际光化学效率(ΦPSII)、光化学猝灭系数(qP)和单位面积有活性的反应中心的密度(RC/CSo)显著小于对照,细胞间隙二氧化碳浓度(Ci)、非光化学猝灭系数(NPQ)、光系统Ⅱ受体侧放氧复合体功能参数(Wk)和捕获的激子将电子传递到电子传递链中QA- 下游的其它电子受体的概率(ψEo)升高。经过1 d的恢复,高温处理的这些参数值与对照没有显著差异。【结论】40℃的高温胁迫导致‘赤霞珠’葡萄叶片的Pn显著下降,不过在常温下1 d就能够恢复。Pn下降不是气孔因子导致的结果,主要与PSⅡ供体侧和反应中心活性以及激发能的分配有关;Pn恢复上升,与上述因子都有关系。<BR></FONT></P>
DOI:10.3864/j.issn.0578-1752.2013.06.012URLMagsci [本文引用: 1]
【目的】探讨温度与光照交叉处理对‘赤霞珠’葡萄叶片光系统功能的伤害机制。【方法】以一年生‘赤霞珠’为试材,设置不同温度(28、34、37、40℃)与光强(800、1600 μmol?m-2?s-1)组合,测定叶片气体交换和叶绿素荧光参数,分析不同温度和光强处理对葡萄叶片PSⅡ活性的影响。【结果】无论高光强还是适宜光强下,随着处理温度升高,叶片净光合速率(Pn)、气孔导度(Gs)、PSⅡ的实际光化学效率(ΦPSⅡ)、电子传递速率(ETR)、光合性能指数(PIABS)、捕获光能用于QA以后的电子传递的概率(ΨEo)、单位面积有活性反应中心的数量(RC/CS)均呈下降趋势;PSⅡ激发压(1-qP)、非光化学淬灭系数(NPQ)、相对荧光曲线的初始斜率(Mo)呈现上升趋势;在适宜光强条件下,28—34℃内,叶片PSⅡ的RC/CS以及受体侧未发生显著变化,Pn的下降主要是由于气孔限制引起的,温度达到和超过37℃时,PSⅡ反应中心和受体侧开始出现伤害,叶黄素循环在植物光破坏防御过程中作用增加;在高光照条件下,高光强/28℃时的Pn已降至适光/34℃时的水平, PIABS已低于适光/40℃时的水平,适宜温度范围内(28—34℃),PSⅡ电子受体侧已受到伤害,温度超过37℃时,反应中心和电子受体侧出现更加严重的伤害,叶黄素循环受到抑制,最大光化学效率明显降低。【结论】适宜光强时,温度达到或超过37℃时,PSⅡ受到伤害,发生较严重的光抑制;强光胁迫在28℃时对PSⅡ功能造成伤害,随着温度的升高,伤害显著加剧,说明强光胁迫比高温胁迫更易造成PSⅡ功能的伤害。
DOI:10.3864/j.issn.0578-1752.2013.06.012URLMagsci [本文引用: 1]
【目的】探讨温度与光照交叉处理对‘赤霞珠’葡萄叶片光系统功能的伤害机制。【方法】以一年生‘赤霞珠’为试材,设置不同温度(28、34、37、40℃)与光强(800、1600 μmol?m-2?s-1)组合,测定叶片气体交换和叶绿素荧光参数,分析不同温度和光强处理对葡萄叶片PSⅡ活性的影响。【结果】无论高光强还是适宜光强下,随着处理温度升高,叶片净光合速率(Pn)、气孔导度(Gs)、PSⅡ的实际光化学效率(ΦPSⅡ)、电子传递速率(ETR)、光合性能指数(PIABS)、捕获光能用于QA以后的电子传递的概率(ΨEo)、单位面积有活性反应中心的数量(RC/CS)均呈下降趋势;PSⅡ激发压(1-qP)、非光化学淬灭系数(NPQ)、相对荧光曲线的初始斜率(Mo)呈现上升趋势;在适宜光强条件下,28—34℃内,叶片PSⅡ的RC/CS以及受体侧未发生显著变化,Pn的下降主要是由于气孔限制引起的,温度达到和超过37℃时,PSⅡ反应中心和受体侧开始出现伤害,叶黄素循环在植物光破坏防御过程中作用增加;在高光照条件下,高光强/28℃时的Pn已降至适光/34℃时的水平, PIABS已低于适光/40℃时的水平,适宜温度范围内(28—34℃),PSⅡ电子受体侧已受到伤害,温度超过37℃时,反应中心和电子受体侧出现更加严重的伤害,叶黄素循环受到抑制,最大光化学效率明显降低。【结论】适宜光强时,温度达到或超过37℃时,PSⅡ受到伤害,发生较严重的光抑制;强光胁迫在28℃时对PSⅡ功能造成伤害,随着温度的升高,伤害显著加剧,说明强光胁迫比高温胁迫更易造成PSⅡ功能的伤害。
DOI:10.1016/j.plantsci.2016.12.004URLPMID:28167040 [本文引用: 1]
Photosystem II (PSII) in plants is susceptible to high temperatures. The cyclic electron flow (CEF) around PSI is thought to protect both PSII and PSI from photodamage. However, the underlying physiological mechanisms of the photosynthetic electron transport process and the role of CEF in grape at high temperatures remain unclear. To investigate this issue, we examined the responses of PSII energy distribution, the P700 redox state and CEF to high temperatures in grape leaves. After exposing ‘Cabernet Sauvignon’ leaves to various temperatures (25, 30, 35, 40 and 4502°C) in the light (60002μmol photons m 612 02s 611 ) for 402h, the maximum quantum yield of PSII ( Fv / Fm ) significantly decreased at high temperatures (40 and 4502°C), while the maximum photo-oxidizable P700 ( Pm ) was not affected. As the temperature increased, higher initial rates of increase in post-illumination Chl fluorescence were detected, which were accompanied by an increase in high energy state quenching (qE). The chloroplast NAD(P)H dehydrogenase-dependent CEF (NDH-dependent CEF) activities were different among grape cultivators. ‘Gold Finger’ with greater susceptibility to photoinhibition, exhibited lower NDH-dependent CEF activities under acute heat stress than a more heat tolerant ‘Cabernet Sauvignon’. These results suggest that overclosure of PSII reaction centers at high temperature resulted in the photoinhibition of PSII, while the stimulation of CEF in grape played an important role in the photoprotection of PSII and PSI at high temperatures through contributing to the generation of a proton gradient.
DOI:10.1071/FP17269URL [本文引用: 1]
Changes in cyclic electron 04ow (CEF) around PSI activity after exposing grape (Vitis vinifera L.) seedling leaves to the combined stress of high temperature (HT) and high light (HL) were investigated. The PSII potential quantum ef03ciency (Fv/Fm) decreased significantly under exposure to HT, and this decrease was greater when HT was combined with HL, whereas the PSI activity maintained stable. HT enhanced CEF mediated by NAD(P)H dehydrogenase remarkably. Compared with the control leaves, the half-time of P700+ re-reduction decreased during the HT treatment; this decrease was even more pronounced under the combined stress, implying signi03cantly enhanced CEF as a result of the treatment. However, the heat-induced increase in nonphotochemical quenching (NPQ) was greater under HL, accompanied by a greater enhancement in high-energy state quenching. These results suggest that the combined stress of HT and HL resulted in severe PSII photoinhibition, whereas CEF showed plasticity in its response to environmental stress and played an important role in PSII and PSI photoprotection through accelerating generation of the thylakoid proton gradient and the induction of NPQ.
URL [本文引用: 2]
探讨O3处理对‘赤霞珠’葡萄叶片光系统功能Ⅱ(PSⅡ)的伤害机制及O3胁迫发生阈值。以一年生‘赤霞珠’为试材,设置不同O3浓度(25、40、55、80和105 n L·L-1),测定叶片叶绿素含量、气体交换和叶绿素荧光参数,分析不同O3浓度处理对葡萄叶片PSII活性的影响。结果表明,随着O3浓度升高,叶片净光合速率(Pn)、气孔导度(Gs)、PSII的实际光化学效率(ΦPSII)、光下最大光化学效率(Fv’/Fm’)、捕获光能用于QA以后的电子传递的概率(ΨEo)、单位面积有活性反应中心的数量(RC/CSm)均呈下降趋势;PSII激发压(1-qP)、K点的相对可变荧光(Wk)、慢相荧光淬灭(qI)呈现上升趋势;在低O3浓度条件下(25~55n L·L-1),Pn的下降主要是由于气孔限制引起的,O3浓度达到并超过80 n L·L-1时,叶绿素含量显著降低,非气孔限制占主导因素,PSII功能开始受到抑制;浓度达到105 n L·L-1时叶片出现较明显的表观伤害症状,电子供体侧伤害程度大于受体侧。实验结果说明,葡萄叶片可以在短时间内(8 h)忍受一定浓度(不超过55 n L·L-1)的O3胁迫,O3浓度达到或超过80 n L·L-1时,PSII功能受到较严重的抑制。
URL [本文引用: 2]
探讨O3处理对‘赤霞珠’葡萄叶片光系统功能Ⅱ(PSⅡ)的伤害机制及O3胁迫发生阈值。以一年生‘赤霞珠’为试材,设置不同O3浓度(25、40、55、80和105 n L·L-1),测定叶片叶绿素含量、气体交换和叶绿素荧光参数,分析不同O3浓度处理对葡萄叶片PSII活性的影响。结果表明,随着O3浓度升高,叶片净光合速率(Pn)、气孔导度(Gs)、PSII的实际光化学效率(ΦPSII)、光下最大光化学效率(Fv’/Fm’)、捕获光能用于QA以后的电子传递的概率(ΨEo)、单位面积有活性反应中心的数量(RC/CSm)均呈下降趋势;PSII激发压(1-qP)、K点的相对可变荧光(Wk)、慢相荧光淬灭(qI)呈现上升趋势;在低O3浓度条件下(25~55n L·L-1),Pn的下降主要是由于气孔限制引起的,O3浓度达到并超过80 n L·L-1时,叶绿素含量显著降低,非气孔限制占主导因素,PSII功能开始受到抑制;浓度达到105 n L·L-1时叶片出现较明显的表观伤害症状,电子供体侧伤害程度大于受体侧。实验结果说明,葡萄叶片可以在短时间内(8 h)忍受一定浓度(不超过55 n L·L-1)的O3胁迫,O3浓度达到或超过80 n L·L-1时,PSII功能受到较严重的抑制。
DOI:10.1016/0005-2728(90)90181-3URL [本文引用: 1]
The imbalance in photoactivity between the two photosystems in broken chloroplasts during steady-state electron transport was investigated using modulated chlorophyll a fluorimetry and oxygen evolution. No imbalance in favor of PS II (imbalance term equals zero) was found at low cation concentration (e.g., 10 mM NaCl) where the membranes are unstacked, while some imbalance in favor of PS II (imbalance term about 0.1–0.2) could be observed at ‘high’ cation medium (e.g., 100 mM for univalent, 5 mM for divalent and 100 μM for trivalent cations) where the membranes are stacked. At the high cation concentration the imbalance was particularly noticeable at a pH range 6–7.5 under conditions where the membranes were non-energized, e.g., in the presence of a range of uncouplers and ionophores (an imbalance term of between 0.44 and 1.1). In the absence of uncouplers or ionophores the imbalance term was initially high, decreasing to a low steady-state value during the light-induced energization of the membranes. The increase in the imbalance measured with the addition of gramicidin D was wavelength-dependent, implying changes in the allocation of excitation energy to the photosystems rather than any other mechanism. This effect was reversed at higher pH: At a pH higher than about 8, the imbalance in absence of uncouplers or ionophores was stronger than in their presence. The relation between the state of imbalance and the cross-membrane proton gradient (ΔpH) was not straightforward or simple, as follows: (i) Imbalance was induced by uncouplers and ionophores also at sufficiently low light intensities which produce only very small ΔpH. (ii) Valinomycin (+KCl) had the same effect as uncouplers like gramicidin D, nigericin, NH 4Cl and others, although it presumably abolishes membrane potential but not ΔpH. (iii) The effect of gramicidin D and NH 4Cl was close to saturation at concentrations which affect ΔpH still minutely. (iv) The same level of large imbalance could be achieved even without uncouplers or ionophores when a much higher cation concentration (approx. 10-fold) than that considered normal to achieve membrane stacking was used. It is therefore concluded that: (a) High cationic levels change the allocation of excitation energy, probably via the screening of negative charges on the thylakoid surface. (b) The effect of uncouplers and ionophores in the presence of high cationic levels most probably reflects, at least in part, the effect on external surface charges, their exposure by membrane energization and their screening by cations.
DOI:10.1016/s1360-1385(96)80019-7URL [本文引用: 1]
http://linkinghub.elsevier.com/retrieve/pii/S1360138596800197
DOI:10.3321/j.issn:1671-3877.2005.06.001URL [本文引用: 2]
JIP-测定(JIP-test)是以生物膜能量流动为基础建立的分析方法。利用该方法可以获得有关光系统II的大量信息。文章介绍了快速叶绿素荧光诱导动力学曲线的定义、数据分析方法及相关参数的意义,并举例说明如何利用该方法分析不同环境条件对光合机构主要是PSII的供体侧、受体侧及PSII反应中心的影响。
DOI:10.3321/j.issn:1671-3877.2005.06.001URL [本文引用: 2]
JIP-测定(JIP-test)是以生物膜能量流动为基础建立的分析方法。利用该方法可以获得有关光系统II的大量信息。文章介绍了快速叶绿素荧光诱导动力学曲线的定义、数据分析方法及相关参数的意义,并举例说明如何利用该方法分析不同环境条件对光合机构主要是PSII的供体侧、受体侧及PSII反应中心的影响。
[本文引用: 1]
URLMagsci [本文引用: 1]
以12年生‘黑叶’荔枝(Litchi chinensis Sonn.‘Heiye’)为研究材料,进行遮荫、环剥及遮荫 + 环剥处理,观测这些处理对新梢生长,以及叶片净光合速率、叶片光系统Ⅱ(PSⅡ)最大光量子效率(F<sub>v</sub>/F<sub>m</sub>)和淀粉含量的影响。结果表明:遮荫处理和环剥处理均能显著抑制枝梢生长,降低叶片的净光合速率,但两种处理抑制光合作用的机制各异。其中,环剥处理导致叶绿素含量降低,产生永久性光抑制,显示光系统Ⅱ(PSⅡ)受到破坏,因此,环剥处理导致光合作用的降低与光反应系统的破坏有关;遮荫下叶片最大光化学效率F<sub>v</sub>/F<sub>m</sub>维持正常,淀粉含量降低,但即使在相同的人工光源下,光合作用速率同样低于对照,可能主要与暗反应系统活性减弱有关;而遮荫可显著缓解环剥对光反应系统的破坏效应。
URLMagsci [本文引用: 1]
以12年生‘黑叶’荔枝(Litchi chinensis Sonn.‘Heiye’)为研究材料,进行遮荫、环剥及遮荫 + 环剥处理,观测这些处理对新梢生长,以及叶片净光合速率、叶片光系统Ⅱ(PSⅡ)最大光量子效率(F<sub>v</sub>/F<sub>m</sub>)和淀粉含量的影响。结果表明:遮荫处理和环剥处理均能显著抑制枝梢生长,降低叶片的净光合速率,但两种处理抑制光合作用的机制各异。其中,环剥处理导致叶绿素含量降低,产生永久性光抑制,显示光系统Ⅱ(PSⅡ)受到破坏,因此,环剥处理导致光合作用的降低与光反应系统的破坏有关;遮荫下叶片最大光化学效率F<sub>v</sub>/F<sub>m</sub>维持正常,淀粉含量降低,但即使在相同的人工光源下,光合作用速率同样低于对照,可能主要与暗反应系统活性减弱有关;而遮荫可显著缓解环剥对光反应系统的破坏效应。
URLMagsci [本文引用: 1]
为了探讨内源水杨酸(salicylic acid,SA)在黄瓜幼苗光合系统响应低温胁迫中的作用机制,</br>采用高效液相色谱法测定低温下黄瓜叶片中内源SA 含量的变化;通过SA 合成抑制剂Paclobutrazol(Pac,</br>100 μmol · L<sup>-1</sup>)喷施和外源SA(50 μmol · L<sup>-1</sup>)饲喂的方法调节内源SA 含量,并测定不同处理幼苗的叶</br>绿素荧光参数和光合碳同化关键酶基因的转录水平。结果显示:低温引起黄瓜幼苗内源SA 含量升高,Pac</br>预处理抑制SA 的积累。低温导致PSⅡ的最大光化学效率(<em>F </em>v/<em>F </em>m)、实际光化学效率(<em>Φ</em> PSII)、潜在光化</br>学活性(<em>F</em> v/<em>F </em>o)和光合电子传递效率(<em>ETR</em>)等降低,叶片光化学猝灭参数[(<em>Y</em>(<em>NO</em>)]升高;内源SA</br>含量降低使PSⅡ活性下降幅度增大,加重了叶片的光损伤程度。低温下PSⅡ吸收的光能分配于光反应的</br>部分减少,而以非光化学反应的过剩能量耗散Ex 为主要的光能分配途径,内源SA 含量降低会加剧光能</br>向Ex 的分配。低温时喷施Pac 的幼苗中Rubisco 小亚基基因(<em>RbcS</em>)和碳酸酐酶基因(<em>CA</em>)的表达水平</br>显著低于对照植株。对喷施Pac 的幼苗外源饲喂SA 后,内源SA 含量升高,低温下叶片光合活性得到有</br>效恢复,光损伤降低,光能分配趋于合理,<em>RbcS </em>和<em>CA</em> 的表达水平升高。上述结果表明,低温下内源SA</br>的积累有助于维持黄瓜叶片中较高的光系统活性和碳同化能力,从而保护光合系统,降低低温胁迫对植</br>物的损伤。
URLMagsci [本文引用: 1]
为了探讨内源水杨酸(salicylic acid,SA)在黄瓜幼苗光合系统响应低温胁迫中的作用机制,</br>采用高效液相色谱法测定低温下黄瓜叶片中内源SA 含量的变化;通过SA 合成抑制剂Paclobutrazol(Pac,</br>100 μmol · L<sup>-1</sup>)喷施和外源SA(50 μmol · L<sup>-1</sup>)饲喂的方法调节内源SA 含量,并测定不同处理幼苗的叶</br>绿素荧光参数和光合碳同化关键酶基因的转录水平。结果显示:低温引起黄瓜幼苗内源SA 含量升高,Pac</br>预处理抑制SA 的积累。低温导致PSⅡ的最大光化学效率(<em>F </em>v/<em>F </em>m)、实际光化学效率(<em>Φ</em> PSII)、潜在光化</br>学活性(<em>F</em> v/<em>F </em>o)和光合电子传递效率(<em>ETR</em>)等降低,叶片光化学猝灭参数[(<em>Y</em>(<em>NO</em>)]升高;内源SA</br>含量降低使PSⅡ活性下降幅度增大,加重了叶片的光损伤程度。低温下PSⅡ吸收的光能分配于光反应的</br>部分减少,而以非光化学反应的过剩能量耗散Ex 为主要的光能分配途径,内源SA 含量降低会加剧光能</br>向Ex 的分配。低温时喷施Pac 的幼苗中Rubisco 小亚基基因(<em>RbcS</em>)和碳酸酐酶基因(<em>CA</em>)的表达水平</br>显著低于对照植株。对喷施Pac 的幼苗外源饲喂SA 后,内源SA 含量升高,低温下叶片光合活性得到有</br>效恢复,光损伤降低,光能分配趋于合理,<em>RbcS </em>和<em>CA</em> 的表达水平升高。上述结果表明,低温下内源SA</br>的积累有助于维持黄瓜叶片中较高的光系统活性和碳同化能力,从而保护光合系统,降低低温胁迫对植</br>物的损伤。
DOI:10.1038/nature02598URLPMID:15175756 [本文引用: 1]
Abstract Photosynthesis provides at least two routes through which light energy can be used to generate a proton gradient across the thylakoid membrane of chloroplasts, which is subsequently used to synthesize ATP. In the first route, electrons released from water in photosystem II (PSII) are eventually transferred to NADP+ by way of photosystem I (PSI). This linear electron flow is driven by two photochemical reactions that function in series. The cytochrome b6f complex mediates electron transport between the two photosystems and generates the proton gradient (DeltapH). In the second route, driven solely by PSI, electrons can be recycled from either reduced ferredoxin or NADPH to plastoquinone, and subsequently to the cytochrome b6f complex. Such cyclic flow generates DeltapH and thus ATP without the accumulation of reduced species. Whereas linear flow from water to NADP+ is commonly used to explain the function of the light-dependent reactions of photosynthesis, the role of cyclic flow is less clear. In higher plants cyclic flow consists of two partially redundant pathways. Here we have constructed mutants in Arabidopsis thaliana in which both PSI cyclic pathways are impaired, and present evidence that cyclic flow is essential for efficient photosynthesis.
DOI:10.1007/s11120-007-9184-yURLPMID:17554634 [本文引用: 1]
Recent investigations of photoinhibition have revealed that photodamage to photosystem II (PSII) involves two temporally separated steps: the first is the inactivation of the oxygen-evolving complex by light that has been absorbed by the manganese cluster and the second is the impairment of the photochemical reaction center by light that has been absorbed by chlorophyll. Our studies of photoinhibition in Synechocystis sp. PCC 6803 at various temperatures demonstrated that the first step in photodamage is not completed at low temperatures, such as 10 C. Further investigations suggested that an intermediate state, which is stabilized at low temperatures, might exist at the first stage of photodamage. The repair of PSII involves many steps, including degradation and removal of the D1 protein, synthesis de novo of the precursor to the D1 protein, assembly of the PSII complex, and processing of the precursor to the D1 protein. Detailed analysis of photodamage and repair at various temperatures has demonstrated that, among these steps, only the synthesis of the precursor to D1 appears to proceed at low temperatures. Investigations of photoinhibition at low temperatures have also indicated that prolonged exposure of cyanobacterial cells or plant leaves to strong light diminishes their ability to repair PSII. Such non-repairable photoinhibition is caused by inhibition of the processing of the precursor to the D1 protein after prolonged illumination with strong light at low temperatures.
DOI:10.1111/j.1365-3040.2004.01172.xURL [本文引用: 1]
Photosynthesis is inhibited by high temperatures that plants are likely to experience under natural conditions. Both increased thylakoid membrane ionic conductance and ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) deactivation have been suggested as the primary cause. The moderately heat-tolerant crop Pima S-6 cotton ( Gossypium barbadense ) was used to examine heat stress-induced inhibition of photosynthesis. Previous field-work indicated that moderate heat stress ( T = 35鈥45 掳C) is associated with very rapid leaf temperature changes. Therefore, a system was devised for rapidly heating intact, attached leaves to mimic natural field heat-stress conditions and monitored Rubisco activation, carbon-cycle metabolites, thylakoid ionic conductance, and photosystem I activity. As a proxy for NADPH and stromal redox status the activation state of NADP-malate dehydrogenase (NADP-MDH) was measured. In dark-adapted cotton leaves, heating caused an increase in thylakoid permeability at temperatures as low as 36 C. The increased permeability did not cause a decline in adenosine 5'-triphosphate (ATP) levels during steady-state or transient heating. Rapid heating caused a transient decline in ribulose 1,5-bisphosphate without a decrease in Rubisco activation. Sustained heating caused a decline in Rubisco activation and also oxidized the stroma as judged by NADP-MDH activation and this is hypothesized to result from increased cyclic photophosphorylation, explaining the maintenance of ATP content in the face of increased thylakoid membrane ion leakiness.
DOI:10.1016/j.tplants.2005.11.002URLPMID:16359910 [本文引用: 1]
Farmers and breeders have long known that often it is the simultaneous occurrence of several abiotic stresses, rather than a particular stress condition, that is most lethal to crops. Surprisingly, the co-occurrence of different stresses is rarely addressed by molecular biologists that study plant acclimation. Recent studies have revealed that the response of plants to a combination of two different abiotic stresses is unique and cannot be directly extrapolated from the response of plants to each of the different stresses applied individually. Tolerance to a combination of different stress conditions, particularly those that mimic the field environment, should be the focus of future research programs aimed at developing transgenic crops and plants with enhanced tolerance to naturally occurring environmental conditions.
DOI:10.1016/S0167-8809(99)00111-5URL [本文引用: 1]
The aim of the study was to compare two ozone exposure patterns with different peak concentrations under equal dose conditions. Spring wheat ( Triticum aestivum cv. ‘Nandu’) was ‘pre-cultivated’ under glasshouse conditions and exposed to two different ozone exposure scenarios (A: 1700–0900 h: 20 ppb, 0900–1700 h: 65 ppb; B: 1600–1200 h: 20 ppb, 1200–1600 h: 110 ppb) in large fumigation chambers for two weeks during anthesis. Control plants received charcoal filtered ambient air (<5 ppb ozone 24 h/day) (CF). Chlorophyll content as well as chlorophyll fluorescence (Fv : Fm ratio) and carbohydrate composition (hexoses, sucrose, starch) were monitored in flag leaves and in the case of carbohydrate content also in the penultimate internode during and after ozone exposure. Thousand grain weight (TGW) was determined for estimating yield effects. Photosynthesis decreased markedly in both treatments expressed as Fv : Fm ratio. In treatment B chlorophyll content (a + b) of leaves was reduced by 40% after 7 and 14 days of exposure ( p ≤ 0.001). Carbohydrate content decreased significantly as compared with CF-plants to 20% ( p ≤ 0.001) in the case of sucrose and starch. Effects were especially pronounced in treatment B (110 ppb for 4 h) and could be observed even seven days after exposure under CF conditions. After 14 days of exposure during anthesis, plants were incapable of recovery as shown by a significant reduction in thousand grain weight of ozone treated plants compared with CF-plants (treatment B: 22% at p ≤ 0.001). Effects on photosynthesis and carbon content even under moderate daily ozone peaks as in treatment A induced a significant yield loss of 11% ( p ≤ 0.001) as well, when applied during anthesis. It can be concluded that under equal dose conditions ozone treatments with comparatively high peak concentration induce more pronounced effects than ozone patterns with moderate peaks.
[本文引用: 1]
DOI:10.1016/j.scienta.2017.09.051URL [本文引用: 1]
DOI:10.1111/j.1365-3040.2004.01171.xURL [本文引用: 1]
Restrictions to photosynthesis can limit plant growth at high temperature in a variety of ways. In addition to increasing photorespiration, moderately high temperatures (35–42 °C) can cause direct injury to the photosynthetic apparatus. Both carbon metabolism and thylakoid reactions have been suggested as the primary site of injury at these temperatures. In the present study this issue was addressed by first characterizing leaf temperature dynamics in Pima cotton ( Gossypium barbadense ) grown under irrigation in the US desert south-west. It was found that cotton leaves repeatedly reached temperatures above 40 °C and could fluctuate as much as 8 or 10 °C in a matter of seconds. Laboratory studies revealed a maximum photosynthetic rate at 30–33 °C that declined by 22% at 45 °C. The majority of the inhibition persisted upon return to 30 °C. The mechanism of this limitation was assessed by measuring the response of photosynthesis to CO 2 in the laboratory. The first time a cotton leaf (grown at 30 °C) was exposed to 45 °C, photosynthetic electron transport was stimulated (at high CO 2 ) because of an increased flux through the photorespiratory pathway. However, upon cooling back to 30 °C, photosynthetic electron transport was inhibited and fell substantially below the level measured before the heat treatment. In the field, the response of assimilation ( A ) to various internal levels of CO 2 ( C i ) revealed that photosynthesis was limited by ribulose-1,5-bisphosphate (RuBP) regeneration at normal levels of CO 2 (presumably because of limitations in thylakoid reactions needed to support RuBP regeneration). There was no evidence of a ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) limitation at air levels of CO 2 and at no point on any of 30 A – C i curves measured on leaves at temperatures from 28 to 39 °C was RuBP regeneration capacity measured to be in substantial excess of the capacity of Rubisco to use RuBP. It is therefore concluded that photosynthesis in field-grown Pima cotton leaves is functionally limited by photosynthetic electron transport and RuBP regeneration capacity, not Rubisco activity.
DOI:10.1023/A:1012446811036URL [本文引用: 1]
Effects of high temperature on the activity of photosynthetic enzymes and leaf proteins were studied in mulberry ( Morus alba L. cv. BC2-59). A series of experiments were conducted at regular intervals (120, 240 and 360 min) to characterize changes in activities of ribulose-1,5-bisphosphate carboxylase (RuBPC) and sucrose phosphate synthase (SPS), photosystem 2 (PS 2) activity, chlorophyll (Chl), carotenoid (Car), starch, sucrose (Suc), amino acid, free proline, protein and nucleic acid contents in leaves under high temperature (40 C) treatments. High temperature markedly reduced the activities of RuBPC and SPS in leaf extracts. Chl content and PS 2 activity in isolated chloroplasts were also affected by high temperature, particularly over 360 min treatment. Increased leaf temperature affected sugar metabolism through reductions in leaf starch content and sucrose-starch balance. While total soluble protein content decreased under heat, total amino acid content increased. Proline accumulation (1.5-fold) was noticed in high temperature-stressed leaves. A reduction in the contents of foliar nitrogen and nucleic acids (DNA and RNA) was also noticed. SDS-PAGE protein profile showed few additional proteins (68 and 85 kDa) in mulberry plants under heat stress compared to control plants. Our results clearly suggest that mulberry plants are very sensitive to high temperature with particular reference to the photosynthetic carbon metabolism.
DOI:10.1093/pcp/pcm118URLPMID:17804480 [本文引用: 1]
The responses of chlorophyll fluorescence, gas exchange rate and Rubisco activation state to temperature were examined in transgenic rice plants with 130 and 35% of the wild-type (WT) Rubisco content by transformation with rbcS cDNA in sense and antisense orientations, respectively. Although the optimal temperatures of PSII quantum efficiency and CO(2) assimilation were found to be between 25 and 32 degrees C, the maximal activation state of Rubisco was found to be between 16 and 20 degrees C in all genotypes. The Rubisco flux control coefficient was also the highest between 16 and 20 degrees C in the WT and antisense lines [>0.88 at an intercellular CO(2) pressure (Ci) of 28 Pa]. Gross photosynthesis at Ci = 28 Pa per Rubisco content in the WT between 12 and 20 degrees C was close to that of the antisense lines where high Rubisco control is present. Thus, Rubisco activity most strongly limited photosynthesis at cool temperatures. These results indicated that a selective enhancement of Rubisco content can enhance photosynthesis at cool temperatures, but in the sense line with enhanced Rubisco content Pi regeneration limitation occurred. Above 20 degrees C, the Rubisco flux control coefficient declined. This decline was associated with a decline in Rubisco activation. The activation state of Rubisco measured at each temperature decreased with increasing Rubisco content, and the slope of activation to Rubisco content was independent of temperature. We discuss the possibility that the decline in Rubisco activation at intermediate and high temperatures is part of a regulated response to a limitation in other photosynthetic processes.
URL [本文引用: 1]
【目的】探究不同整形方式对酿酒葡萄光合作用及果实品质的影响,进而对多主蔓扇形及厂字形在我国新疆产区表现的优劣作出评价。【方法】以多主蔓扇形和厂字形2种整形方式的‘赤霞珠’葡萄为试材,利用LI-6400便携式光合测定系统分别测定成熟叶片光合作用相关参数及光响应曲线,调查植株叶幕特征,测定采收期浆果的理化指标并利用HPLC-MS对花色苷总量进行分析。【结果】与多主蔓扇形相比,厂字形整形增强叶片光合速率的同时提高果实品质,这主要归因于厂字形整形方式为树体带来以下3方面的改变:1)增加植株叶面积总量;2)增强叶幕光照透射率;3)增加果际周围光合有效辐射(PAR)。与多主蔓扇形相比,厂字形整形下的叶片光合能力更强,代谢速率更快,并且具备更强的利用弱光和抵御强光的能力。虽然厂字形整形使得植株的产量降低,但其果穗质量更小,更加松散,且采收期果实含有的种子和果皮鲜质量更大,种子数量更多。另外,厂字形整形下的果实可溶性固形物含量以及花色苷浓度也高于多主蔓扇形。【结论】与传统的多主蔓扇形相比,厂字形整形方式更有利于果实酿酒品质的提升。
URL [本文引用: 1]
【目的】探究不同整形方式对酿酒葡萄光合作用及果实品质的影响,进而对多主蔓扇形及厂字形在我国新疆产区表现的优劣作出评价。【方法】以多主蔓扇形和厂字形2种整形方式的‘赤霞珠’葡萄为试材,利用LI-6400便携式光合测定系统分别测定成熟叶片光合作用相关参数及光响应曲线,调查植株叶幕特征,测定采收期浆果的理化指标并利用HPLC-MS对花色苷总量进行分析。【结果】与多主蔓扇形相比,厂字形整形增强叶片光合速率的同时提高果实品质,这主要归因于厂字形整形方式为树体带来以下3方面的改变:1)增加植株叶面积总量;2)增强叶幕光照透射率;3)增加果际周围光合有效辐射(PAR)。与多主蔓扇形相比,厂字形整形下的叶片光合能力更强,代谢速率更快,并且具备更强的利用弱光和抵御强光的能力。虽然厂字形整形使得植株的产量降低,但其果穗质量更小,更加松散,且采收期果实含有的种子和果皮鲜质量更大,种子数量更多。另外,厂字形整形下的果实可溶性固形物含量以及花色苷浓度也高于多主蔓扇形。【结论】与传统的多主蔓扇形相比,厂字形整形方式更有利于果实酿酒品质的提升。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}