 ,1,31
,1,31 2
3
Regulation Function of Trehalose-6-phosphate Synthase Genes on Chitin Synthesis in Sogatella furcifera
ZHANG DaoWei1, YU YaYa2, PAN BiYing3, KANG Kui1, ZENG BoPing1, CHEN Jing2, TANG Bin,1,31 2
3
ͨѶ����:
���α༭: ��÷
�ո�����:2019-05-5��������:2019-06-10�����������:2019-10-01
| ��������: |
Received:2019-05-5Accepted:2019-06-10Online:2019-10-01
����� About authors
�ŵ�ΰ,E-mail��zhangdw1000@163.com��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (1147KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
�ŵ�ΰ, �����, �˱�Ө, ����, ����ƽ, �¾�, �Ʊ�. �ױ���ʭ�����Ǻϳ�ø������ؼ����ʺϳɵĹ���[J]. �й�ũҵ��ѧ, 2019, 52(19): 3357-3366 doi:10.3864/j.issn.0578-1752.2019.19.007
ZHANG DaoWei, YU YaYa, PAN BiYing, KANG Kui, ZENG BoPing, CHEN Jing, TANG Bin.
0 ����
���о����塿ˮ����Oryza sativa�����ҹ�����Ҫ����ʳ����֮һ,����ˮ���ĸ߲����Ȳ����ҹ���ʳ��ȫ������ռ�м�����Ҫ�ĵ�λ��Ȼ��,ˮ����������϶�[1],���е���ʭ�ѳ�Ϊˮ����ͷ�ź���[2],����Ҫ�����ַ�ʭ��Nilaparvata lugens�����ױ���ʭ��Sogatella furcifera���ͻҷ�ʭ��Laodelphax striatellus��3�֡���ʭΪ��ʳ�Ժ���,����ˮ������Σ��,������������ڶ̡���ֳ�ٶȿ졢Ǩ����ɢ�����ͻ�����Ӧ��ǿ���ص�[3,4,5,6]��Ŀǰ,�ױ���ʭ�ѳ�Ϊ�ҹ��Լ������ֹ���ˮ���ϵ���Ҫ����,Ҳ��ˮ����ǰ����Ҫ��Ǩ���Ժ���֮һ,Σ����Χ��,�����Ѷȴ�[7]��̽���ױ���ʭ�����Ǻϳ�ø����SfTPS���Լ����ʺϳɵ�Ӱ��,��ɸѡ�ʺϵĺ�����ưб���ΰױ���ʭ������Ҫ���塣��ǰ���о���չ����������Ϊһ��˫��,����������ֲ������ϸ����,����������Ҳ�ֲ��Ϲ�,�ر���������Ѫ�ܰ���,�����Ǻ����dz���,��Ϊ����ġ�Ѫ�ǡ�,�������������������̬�ͷ�ֳ�ȹ����о����зdz���Ҫ�Ĺ���[8,9]�������ǵĺϳ�;����ֹһ��,����������Ҫ���õ���������,ͨ�������Ǻϳ�ø��trehalose-6-phosphate synthase,TPS��;����֬�����кϳ�[10,11],����������ʱͨ��������ø��trehalase,TRE������Ϊ������,�Ӷ�����������л�ͼ����ʺϳ�ͨ·��[12,13,14]��������֪,�����ʺ���Ӧ�ĵ��������������������Ҫ��ɲ���,���������Ƥ�����ܺ�ΧʳĤ�Ƚṹ����Ҫ�ɷ�Ҳ�Ǽ�����,���׳��������ض��Ľ�ʱ����Ҫ���ܼ����ʽṹ,�����Ƥ��Ϊ,���������Ҫ�ɼ����ʺϳ�ͨ·�ͼ�����ø���临���干ͬ���[15]�������ʺϳ�ͨ·��,�ܹ�����8����ͬ�Ļ������,������øΪ���漸���ʺϳ�ͨ·�ĵ�һ��ø,�����ʺϳ�ø��chitin synthase,CHS��Ϊ�����ʺϳɵ����һ��ø,Ҳ����Ϊ��Ҫ��[13,15]��CHEN���о�����,TRE��CHS�ı��ﱻ���ƺ�,����ļ����ʺϳɾ����ܵ����ص��谭,��������Ƥ����,���������������������������[12,16-17]�������о�����㡿�ںַ�ʭ�͵¹�СBlattella germanica���п�¡�ͷ�����2����ͬ��TPS�������[18,19],�����ںַ�ʭ�д����ŵ�3��TPS[14,20-21]������RNAi��������TPS�ı���,ͬ���ܹ������������Ƥ����,����������һ����������[19,22],�����TPS�ڼ����ʺϳ�������һ���ĵ�������,���������ĵ��������Լ���ͬTPS֮���Ƿ���ڲ�����о����١��ڰױ���ʭ��ͬ�����ִ���2����ͬ��SfTPS[23],�û����Ƿ��ܹ����ؼ����ʺϳ��Լ���ε��ؼ����ʺϳ���δ��ȷ,���ѡ��ױ���ʭΪ�о�����̽��SfTPS�Ĺ��ܡ��������Ĺؼ����⡿�о��ױ���ʭSfTPS���伸���ʺϳ�ͨ·�еĵ��ع���,�������溣���Ǻϳ�;���뼸���ʺϳ�;���Ĺ�����,̽��TPS�Ƿ�߱���Ϊ������εİб�����DZ��,Ϊ�����ױ���ʭ��ˮ�����������ũҩ�ṩ�������ݡ�1 �����뷽��
������2018��������ʦ��ѧԺ�ͺ���ʦ����ѧ��ɡ�1.1 ���Գ�Դ
�ױ���ʭ��������ʦ��ѧԺ������ũҵ�Ƽ�ѧԺ������Ⱥ,ˮ��Ʒ��ȫ�����øг�ˮ��TN-1���ױ���ʭ�����������4�俪ʼͳһ�ռ�,���䳤��5�������,���ں���ע�����顣1.2 ���鷽��
1.2.1 ��RNA�ij��ἰcDNA�ϳ� �ױ���ʭ�ij�����RNA�������Trizol��,�ϸ������Լ���˵������С���ȡ����1.2%����֬�Ǽ����RNA������,����NanoDropTM 2000�ֹ��ȼƲⶨ�䴿�Ⱥ�Ũ�ȡ�cDNA�ϳ�����PrimeScript?RT reagent Kit With gDNA Eraser�Լ���˵������С�1.2.2 dsRNA�ĺϳ� ������֪�İױ���ʭTPS����������,ѡ��2��SfTPS���ʵ�dsRNA������Ƭ�ν������������ϳɡ������������м���1,dsRNA�ĺϳɷ����ο�T7 RiboMAXTM Express RNAi System�Լ��е�˵�����С��ϳɵ�dsRNA����NanoDropTM 2000�ⶨŨ�ȡ�ͬʱ��GFPΪģ��,����ͬ���ķ����ϳ�GFP��dsRNA,��Ϊ������[17]�����,���ڰױ���ʭ��ע���dsRNAƬ�γ��ȷֱ�ΪdsSfTPS1��523 bp����dsSfTPS2��509 bp���Լ���Ϊ���յ�dsGFP��738 bp��,����SfTPS1��SfTPS2 dsRNA�������Ƚϼ�ͼ1��
Table 1
��1
��1dsRNA�ϳ���������
Table 1
| �������� Primer name | �������� Forward primer (5��-3��) | �������� Reverse primer (5��-3��) | ���ﳤ�� Length (bp) |
|---|---|---|---|
| dsSfTPS1 | CCCGTTGTGGTGAGAAATA | CAAGGTGGGAATGGAATG | 473 |
| dsSfTPS1-T7 | T7-CCCGTTGTGGTGAGAAATA | T7-CAAGGTGGGAATGGAATG | 523 |
| dsSfTPS2 | CGCATAGACCGCAACAAC | TCGCAACGGAGTAACCAG | 459 |
| dsSfTPS2-T7 | T7-CGCATAGACCGCAACAAC | T7-TCGCAACGGAGTAACCAG | 509 |
| dsGFP | AAGGGCGAGGAGCTGTTCACCG | CAGCAGGACCATGTGATCGCGC | 688 |
| dsGFP-T7 | T7-AAGGGCGAGGAGCTGTTCACCG | T7-CAGCAGGACCATGTGATCGCGC | 738 |
�´��ڴ�|����CSV
ͼ1
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1�ױ���ʭTPS1��TPS2���������бȶ�ͼ
Fig. 1Comparison of nucleotide sequence of TPS1 and TPS2 in S. furcifera
1.2.3 �ױ���ʭ����ע�䡢�۲�ͳ�Ƽ�����ѡȡ ������ע��IJ��Ϸ�ΪdsSfTPS1��dsSfTPS2�Լ���Ϊ���յ�dsGFP��ȡ5���1��İױ���ʭ������ע��,��ע���Dz�������Ϊע��ѹǿ200 hPa,ע��ʱ��0.5 s,����ѹ20 hPa,ע����Ϊ50 ng/ͷ��ע���ʭ���뺬ˮ���IJ�������,��������ס�ܿ�,�������ܷ����˹�������,������������Ϊ�¶ȣ�26��0.5����;������L�sD=16 h�s8 h,���ʪ�ȣ�50��5��%��24 h��ױ���ʭ�Ĵ����Ϊ��Чע������ע���ֳ�4������,ÿ����3���ظ�����1��ÿ��24 h�۲�ױ���ʭ�ķ������,��48 h��72 hͳ�ƻ�������������;��2��ע���48 h��72 h�ֱ�ȡ��,����ӫ�ⶨ����qRT-PCR������RNAi����Ч����SfCHS����;��3����ע���48 h ��72 h,���ڲⶨ�����ʺ���;���һ��������ƤΪ�ɳ��,�۲����������������ա�
1.2.4 RNAi��SfTPS����Ч����SfCHS�������ⶨ �ױ���ʭ��ע���48 h��72 hȡ�����������qRT-PCR���,�������2,���鷽��������¶��[24]��
Table 2
��2
��2ʵʱӫ�ⶨ��PCR�����������
Table 2
| �������� Primer name | �������� Forward primer (5��-3��) | �������� Reverse primer (5��-3��) | ���ﳤ�� Length (bp) |
|---|---|---|---|
| Q-SfTPS1 | CCGATTCGCTACATCTACG | GACAAACTCTTTCGCCACTAA | 123 |
| Q-SfTPS2 | GATGCTGAGGGCAAAGAC | TGTGGAAGCCGACAAAGT | 226 |
| Q-SfCHS1 | GATTGGTCATTGGCTTCAGA | GTAATGTCTTGCTTCGTCAG | 151 |
| Q-SfCHS1a | CTTCGGTGTTTGGTTTCTT | TGGGTAACATCATCATAGGA | 136 |
| Q-SfCHS1b | GAGAAGGCGAGAATAGCA | GCAGCAAGAACACGATTA | 103 |
| Q-18S rRNA | GCCCCGTAATCGGAATGAGT | GACAAGACGTCCCGCAAAAC | 205 |
�´��ڴ�|����CSV
1.2.5 �����ʺ����IJⶨ �ױ���ʭ��������ռ�����ļ�ⷽ���ο�����[24,25]�����ʵ��ġ���Ҫ�����������1�������ܣ���һ����Լ30��50 cm������ͷ��������������,��ȡһ���Թ����������,����һ�˵IJ�����Ƕ��������,��һ����ͨ��ˮ��,Ԥ����Һ����;��2����Ʒ�������ױ���ʭ��Ϊÿ��40ͷ,ƽ��3��,50������ں��������;��3������ɵİױ���ʭ���г���,��ΪW1;��4���Ѻ�ɺ���嵹����������,����5 mL��������������Һ,160�����ԡ,���������������Ĥ;��5������ֽ�Ƴ�©��״���˲���,С����ˮ��ϴ�ɾ�������50������к��,���ز���ΪW2;��6�����չ�ʽ���㼸���ʵ���Ժ����������ʵ���Ժ�����%��=��W2/W1����1.26��100,����1.26Ϊ���������������백�������ǵ���Է�������֮�ȡ�
1.2.6 ����ͳ������� ͨ��qRT-PCR�ⶨ�����CTֵ,ÿ������ѧ�ظ�����3�������ظ�,3�������ظ�����ֵ���������Ƚ�,��ֵ�����0.5���ڿ���,3����ֵ����2����3���ӽ�,��ֵ����,����������ÿ����Ʒ��3������ѧ�ظ�,�����õ�������Ϊƽ��ֵ��9���ظ���������2-����CT������,�ױ���ʭdsGFPע�����CTֵΪ�����顣��������ֵ
�ٽ��о��������2-����CT���㹫ʽ[26]��
2-����CT=2-[(CT������-CT����18S)- (CT������-CT������18S)]
Ӧ��Excel������������,��ʹ��SPSS��������ͳ�Ʒ���,One-Way ANOVA�����в��������Լ��飨P��0.05Ϊ��������,��*��ʾ;P��0.01Ϊ���켫����,��**��ʾ��,���ݱ�ʾΪƽ��ֵ��������������ݲ���SigmaPlot 10.0��ͼ��
2 ���
2.1 RNAi��SfTPS���P��Ƥ���
����dsRNAע�䵽�ױ���ʭ���ں�,����RNAi�ܹ���Ч����SfTPS�����dsSfTPS1��dsSfTPS2ע���48 h��72 h,SfTPS1��SfTPS2�ı���ֱ���������ע��dsGFP��ı���,����������������30%;ע��dsSfTPS1��dsSfTPS2��,�ǰб�SfTPS�ı���ͬ�����������ͣ�ͼ2-A��2-B����dsSfTPS1��dsSfTPS2���ܹ�����ױ���ʭ��ƤΪ�ɳ�����Σ�ͼ2-C��,���DZ�Ƥ��Ƥ�ϰ������ԡ�ͼ2
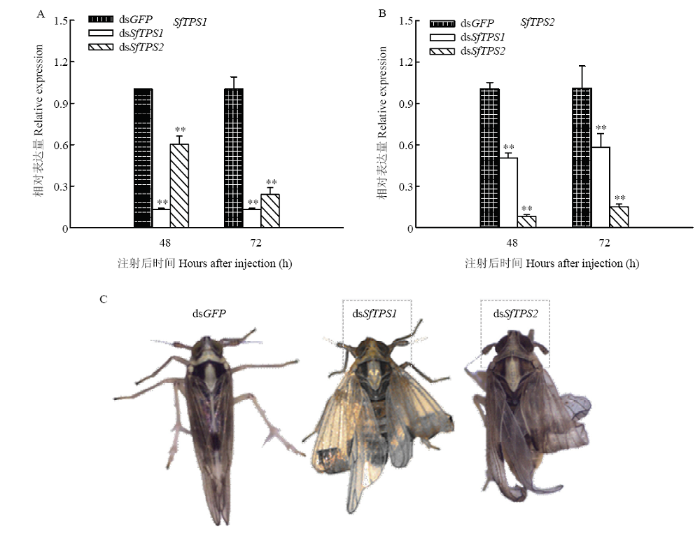 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2RNAi��ױ���ʭTPS����Ա��������ɳ���Ƥ���
Fig. 2The relative expression of two TPSs and moulting of adults of S. furcifera after RNAi
2.2 RNAi����SfTPS������ʺ����ı仯
����������,dsSfTPS1ע���48 h��72 h�����ʺ�������������,��dsSfTPS2ע���72 h�����ʵĺ���ͬ��������������ͼ3�����������,SfTPS�����½������ٽ��˰ױ���ʭ�����м����ʺ�����ߡ�ͼ3
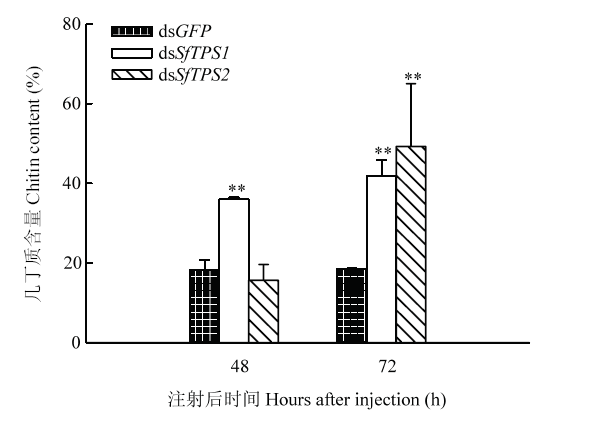 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3RNAi��ױ���ʭ���ڼ����ʵĺ����仯
Fig. 3Changes of chitin content in S. furcifera after RNAi
2.3 SfTPS RNAi��SfCHS���ɱ�ת¼�ӱ���仯
��ʭ�в�����CHS2�������,��CHS1����������ͬ�ļ�ӱ�����[27]������RNAi����SfTPS1��SfTPS2�����,��mRNAˮƽ�ϼ�⼸���ʺϳ�øSfCHS1����������ͬת¼��SfCHS1a��SfCHS1b�ı�������ʾSfCHS1��SfCHS1a��������һ��,�����ע��dsGFP�����,ע��dsSfTPS1��72 h����������������,����48 hʱ����������;ע��dsTPS2��48 h��72 hʱ��������������������ͼ4-A��4-B��������,ע��dsSfTPS1��dsSfTPS2���ܹ���Ч���SfCHS1b�ı���,SfCHS1b��SfTPS1��SfTPS2���ﱻ���ƺ��48 h��72 h�ı�������Ϊ���������ߣ�ͼ4-C����ͼ4
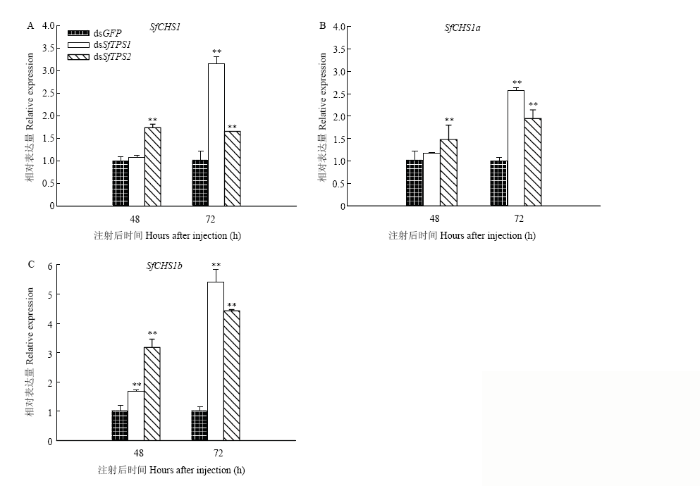 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4SfTPS RNAi��SfCHS���ɱ�ת¼�ӵ���Ա�����
Fig. 4The relative expression of SfCHS and variants transcripts after SfTPS RNAi
2.4 SfTPS RNAi��ױ���ʭ�Ļ����ʺ�������
Ϊ�˽⼸���ʺϳ���߶Ի����ʺ������ʵ�Ӱ��,�ⶨ��SfTPS1��SfTPS2���ﱻ���ƺ�48 h��72 h�Ļ����ʺ������ʡ��������,����dsSfTPS1��dsSfTPS2ע���48 h��72 h�ױ���ʭ��������Խϵ�,72 h��������δ����10%,����ע��dsGFP�����,�������������������죨ͼ5-A��;dsGFP�鴦����,�ױ���ʭ�����������,��SfTPS1��SfTPS2���ﱻ���ƺ�48 h��������������������,����72 h��������������ͼ5-B����ͼ5
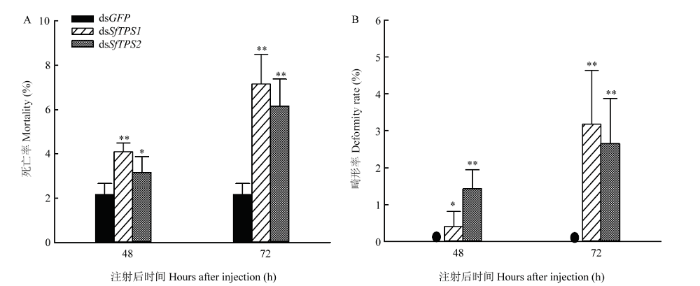 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ5SfTPS RNAi��ױ���ʭ�Ļ����ʺ�������

Fig. 5Deformity rate and mortality of S. furcifera after SfTPS RNAi
The deformity rate of dsGFP control group is 0
3 ����
��������֬�����кϳɺ�,�ᱻ�ͷŵ�Ѫ�ܰ���,ͨ���ܰ�ѭ�����͵�������֯�з��ӹ���[22,28]��AVONCE���о�����,�����Ǿ���5�ֲ�ͬ��DZ�ںϳ�;��,������Ҫͨ�������Ǻϳ�ø��TPS��/������-6-����֬ø��TPP��;��[29],Ҳ���п����ܹ�ֱ��ͨ����һ��TPS�����,TPS������TPS��TPP�������ؽṹ��,�Ҳ�������δ���ֵ���TPP [14]��Ŀǰ,�ڵ¹�С�ַ�ʭ�Ͱױ���ʭ�з���2����2�����ϵ�TPS [18,19],���ڻ�Į�׳�С�ر�ף�Microdera punctipennis���ĸ�ͨ�������з�����5��DZ��TPS��Unigenes[30],��Щ�����������ĺ����Ǻϳ�;��һֱ�ڽ�����,������ν����Լ�����Ľ�������ȴ�о����١����ں����ǵ���Ҫ����,TPS���о�Ҳ�ܵ�Խ��Խ��Ĺ�ע,�书���о�����Ҫ��������ͨ��RNAi����,����ע��dsRNA��siRNA���������ı���[31],̽������ص�DZ��ͨ·�ȡ�RNAi����ͨ�����г�Ĭ�ض�����Ĺ���,Ȼ�����µ��о����ֵ������õͻ��ó���,ͬ����������ڻ���ЧӦ,����������[32],���������ںַ�ʭTRE��ͬ������[33]��Ȼ��,�ںַ�ʭ���ڵ�������TPS1��TPS2,����������۸���TPS1����TPS2,TPS1��TPS2��ת¼��������[19];��������,��ע��dsSfTPS1��dsSfTPS2��,����һ��SfTPS�ı���ͬ���ܵ����ƣ�ͼ2-A��2-B��,ԭ�������SfTPS��Ա��س̶Ƚϸ��й�,��Ҫ��һ���о�ȷ��������Ĭ���淢���Ĺؼ������,ͨ�����ڲ�ͬ����֯��������ʾ����[34,35,36]�������ҹ�꣨Spodoptera exigua��TPS���ﱻ���ƺ�48 h��204 h,�������ʷֱ�Ϊ50.94%��66.76%,��������dsGFP������[22]��ͬ��,�ַ�ʭTPS1��TPS2���ﱻ���ƺ�,�ɳ������Ƥ�ϰ�,�����ȫ�γ�,�����ʴﵽ20%,�������ʽӽ�30%[19];�ַ�ʭTPS3���ﱻ���ƺ�,ͬ���������Ƶ���Ƥ��������[21];���ڰױ���ʭ��,δ������Ƥ���ѵĻ���,�����ֳ��ܹ���������,������һ���Ļ��Σ�ͼ2-C��,72 h��������Ҳ�Ȼ�����Ҫ�ߣ�ͼ5�����ۺϺַ�ʭ�Ͱױ���ʭTPS�Ĺ����о����,�������ַ�ʭ��TPS���ﱻ���ƺ�,�����ԵĻ��α�����,��ԭ������dz���ΧʳĤ�����ܵļ����ʺϳ��ܵ�Ӱ��[37],��Ϊ���ܵ�������������л�����ض���������[11]��
�����ʺϳɵ�ͨ·ʼ�ں�����ø,���ڼ����ʺϳ�ø��CHS��,�����ʺϳɺͽ���Ķ�̬ƽ����������Ƥ��������Ϊ��Ҫ������[13,15]�����ƺַ�ʭCHS1��CHS1a��CHS1b�������,��CHS1��CHS1a�ĸ��Żᵼ�ºַ�ʭ����Ρ�ϸ������Ƥ������������[27]������TRE���ﱻ����ʱ,����ͬ������ַ������Ρ����ؼ��ᡢ�����ʺϳɼ��١��������衢���м�����������������[10],�ر��Dz��ú�����ø���Ƽ������ַ�ʭ��,�伸���ʺ��������½�[38]����Щ�о�����,TRE��CHS�ȱ������ƺ�,�ή�ͼ����ʺ���,���������з��ְױ���ʭTPS1��TPS2���ﱻ���ƺ�,�����ʺ������������������ģ�ͼ3��,��ʭ�ɳ�ᷢ��Ϊ����,����С���ε�,Ҳ��ӱ��������ʺ����ϸߣ�ͼ2-C�������ڳ���ȵ���Tribolium castaneum����,TPS�������ƺ�,CHS1��72 h������������������,CHS2��48 h��72 h�������½�,���ֳ��岻��������Ӽ,���Ҽ����ʺ�������[39]�����Ƿ�˵��TRE��TPS���ܵ��ط�ʭ������ͨ·�Ľ����Ч����ͬ?TRE�ܹ�����CHS���������ø��chitiase,Cht���ı���[25,33],���ƺϳɵ�ƽ��,���¼����ʺ������͡���TPS�������ƺ�,�ܹ��ٽ�CHS�ı��ͼ4��,���Ӽ����ʺ�����ͼ3��,������ʱ��ڵ���,���Űױ���ʭTPS1��TPS2���ʺ�����CHS1a��CHS1b�ı仯����һ��,��CHS1��ʱ��ڵ�ı仯�����в��죨ͼ3��ͼ4��,��˵������TPS�Լ����ʴ�л��Ӱ����Ʋ���ȫ��ͬ��ͬ������������TPS���ﱻ���ƺ�Ҳ�ܹ�ʹ�����ʵĺϳ��ܵ�����[39],���¼����ʴ�лƽ������ҡ��Ժַ�ʭTPS1��TPS2�Ĺ����о�����,��TPS1��TPS2�������ƺ�48 h,CHS1a��CHS1b���オ�ͻ���������,��CHS1��CHS1a��72 hȴ������������������[33]��Ϊʲô�ڷ�ʭ�ͳ���ȵ��д��ڲ�һ�������,�������ʭ�Ķ̳�ͳ�������һ���Ĺ���,ͼ2-C�аױ���ʭΪ������,�伸���ʺ���������,���ַ�ʭ�̳����м����ʺϳ������ܵ��谭[19]����������,�����ưױ���ʭ�����ʺϳ�ͨ·��TPS������ø����ı���,�ᵼ�¼����ʺ������ʹӶ�������Ƥ��������;�������ʺ���������Ҳ�ܵ�������ᷢ�����μ�������
4 ����
�����Ǻϳ�ø����TPS����Ϊ�����ǺϳɵĹؼ�����,���ܹ��鵼�����ʺϳ�ø����CHS�������Ե��ؼ����ʺϳ�,�����ױ���ʭ�����Ǻϳ�;����TPS���ﱻ���ƺ�,��ͨ���ٽ�CHS�ı���,������ʺ���,Ӱ��������������ᷢ�����ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
Magsci [��������: 1]

������ø(Treh)������������л�ز����ٵ�һ��ø�� �����������ڼ����ʺϳ�ͨ·�ĵ�һ��ø�����������ø����ֱ�������������� ��Ƥ�� ��̬�Լ���ֳ��������Ҫ��������������ء�Ŀǰ���ж�������ĺ�����ø���ɹ���¡�� �Ӷ��������溣����ø��������ɶ����Ա��ɡ�������ø����������ĵ��״��������һ���ź���ǰ������ ���ֵ���ӵ��1~2����Ĥ�ṹ�� �����Ƿ���п�Ĥ�ṹ�� �ɽ����Ϊ�����Ժ�����ø(Treh1)��Ĥ����ͺ�����ø(Treh2)���࣬ Ĥ����ͺ�����ø����2�����еı�ǩ���У� ��“PGGRFREFYYWDSY”��“QWDYPNAWPP”��������ø����Ҫ�����ǽ�����Ͱ��ڵĺ����ǽ���������ǣ� Ϊ�����������ṩ�������������Ϊ�������棬 һ�Dz������漸���ʺϳ�;���� �Ӷ����ر�Ƥ�� �г��ȴ��ļ����ʺϳɣ� ����ͨ���뼤�ص�Эͬ���ã� �����������ں����Ǻ������ǵ��������ʵ�Ũ�ȱ仯�� �Ӷ���Ч��������ϸ������Ӧ���ɹ���Ӧ���澳������ ������俹�����������ں�����ø����Ҫ���ܣ� ���ѳ�Ϊ������Ƶ�DZ���°бꡣ��ͬ���ͺ�����ø�Ĺ����о���ø���Ƽ����з���Ӧ�ý���һ���ƶ�����������εķ�չ��
Magsci [��������: 1]
������ø(Treh)������������л�ز����ٵ�һ��ø�� �����������ڼ����ʺϳ�ͨ·�ĵ�һ��ø�����������ø����ֱ�������������� ��Ƥ�� ��̬�Լ���ֳ��������Ҫ��������������ء�Ŀǰ���ж�������ĺ�����ø���ɹ���¡�� �Ӷ��������溣����ø��������ɶ����Ա��ɡ�������ø����������ĵ��״��������һ���ź���ǰ������ ���ֵ���ӵ��1~2����Ĥ�ṹ�� �����Ƿ���п�Ĥ�ṹ�� �ɽ����Ϊ�����Ժ�����ø(Treh1)��Ĥ����ͺ�����ø(Treh2)���࣬ Ĥ����ͺ�����ø����2�����еı�ǩ���У� ��“PGGRFREFYYWDSY”��“QWDYPNAWPP”��������ø����Ҫ�����ǽ�����Ͱ��ڵĺ����ǽ���������ǣ� Ϊ�����������ṩ�������������Ϊ�������棬 һ�Dz������漸���ʺϳ�;���� �Ӷ����ر�Ƥ�� �г��ȴ��ļ����ʺϳɣ� ����ͨ���뼤�ص�Эͬ���ã� �����������ں����Ǻ������ǵ��������ʵ�Ũ�ȱ仯�� �Ӷ���Ч��������ϸ������Ӧ���ɹ���Ӧ���澳������ ������俹�����������ں�����ø����Ҫ���ܣ� ���ѳ�Ϊ������Ƶ�DZ���°бꡣ��ͬ���ͺ�����ø�Ĺ����о���ø���Ƽ����з���Ӧ�ý���һ���ƶ�����������εķ�չ��
[��������: 1]
[��������: 1]
[��������: 2]
DOI:10.3864/j.issn.0578-1752.2018.04.009Magsci [��������: 2]
������Ϊһ�ַǻ�ԭ����,�㷺������ϸ�������ࡢ�����ֲ����������С������DZ���Ϊ����“Ѫ��”,Դ�ڸ���Ϊ����Ѫ�ܰ��е���Ҫ��������,��������������������Ƥ�������������������Ҫ�����á�����ĺ��������ɺ����Ǻϳ�ø��trehalose-6-phosphate synthase,TPS�����ɺ�����-6-����,Ȼ��ͨ������������֬ø��trehalose-6-phosphate phosphatase,TPP�����պϳɺ�����,����������ʱͨ��������ø��trehalase,TRE������Ϊ������,��������������������Ϊ�����Ƥ���г�ΧʳĤ������ϵͳ����Ҫ��ɳɷ�,�����ڷ�����������Ҫ�ɵ��ɱ�Ƥ,�γ��µı�Ƥ,�ù���һֱ�Ǻ�����Ƶ���Ҫ�б�;����������øΪ���漸���ʺϳ�;���ĵ�һ��ø,�ڼ����ʺϳ�ͨ·��������Ҫ�Ĺ��ܡ���ô�����Ǵ�л������ε��ؼ����ʺϳ�;���������������Ƥ�������ʴ�л���أ����Ź����⺣���Ǵ�л��ػ������о������뿪չ,�о�����������溣���ǹ����ڼ����ʺϳ��о�����Ҫ������,������ø��Ϊ�����Ժ�Ĥ���������,������TRE��TPS�ڲ�ͬ���������о��ж��ͬԴ����,�������溣���Ǵ�л����;�������������,�����Ǵ�лֱ�ӵ��ؼ����ʺϳ�;��,�����Ǻ����Ǻϳ�ø���Ǻ�����ø����ĵͱ���,���ܿ��ƺ�����ʹ�乩����ƽ��,�Ӷ����¼����ʺϳ�;������,�ر��Ǽ����ʺϳ�ø������オ�Ͷ���ɼ����ʺ����½�,�õ������ÿɽ�һ������������Ƥ���ѡ��ᷢ�����ε�,���������������ٴ�,������ø���Ƽ��ܹ����ƿ����Ժ�Ĥ��������ຣ����ø���ԡ������ʺϳ�ͨ·��ػ�������ø�������������½�,���¼����ʺ�����������,�����߱��������������������Щ�����ֱ���,һ�����溣���Ǵ�л�Ĺ���ƽ�ⱻ����,��ֱ��Ӱ�쵽����ļ����ʺϳ������������Ƥ�뷢�����̡���������ǰ���ں����Ǵ�л���ؼ����ʺϳɷ���������о��ɹ�,Ϊ�������������ú�����ø���Ƽ��������Ǻϳ�ø���Ƽ�����ɫũҩ���κ����ṩ�������ݡ�
DOI:10.3864/j.issn.0578-1752.2018.04.009Magsci [��������: 2]
������Ϊһ�ַǻ�ԭ����,�㷺������ϸ�������ࡢ�����ֲ����������С������DZ���Ϊ����“Ѫ��”,Դ�ڸ���Ϊ����Ѫ�ܰ��е���Ҫ��������,��������������������Ƥ�������������������Ҫ�����á�����ĺ��������ɺ����Ǻϳ�ø��trehalose-6-phosphate synthase,TPS�����ɺ�����-6-����,Ȼ��ͨ������������֬ø��trehalose-6-phosphate phosphatase,TPP�����պϳɺ�����,����������ʱͨ��������ø��trehalase,TRE������Ϊ������,��������������������Ϊ�����Ƥ���г�ΧʳĤ������ϵͳ����Ҫ��ɳɷ�,�����ڷ�����������Ҫ�ɵ��ɱ�Ƥ,�γ��µı�Ƥ,�ù���һֱ�Ǻ�����Ƶ���Ҫ�б�;����������øΪ���漸���ʺϳ�;���ĵ�һ��ø,�ڼ����ʺϳ�ͨ·��������Ҫ�Ĺ��ܡ���ô�����Ǵ�л������ε��ؼ����ʺϳ�;���������������Ƥ�������ʴ�л���أ����Ź����⺣���Ǵ�л��ػ������о������뿪չ,�о�����������溣���ǹ����ڼ����ʺϳ��о�����Ҫ������,������ø��Ϊ�����Ժ�Ĥ���������,������TRE��TPS�ڲ�ͬ���������о��ж��ͬԴ����,�������溣���Ǵ�л����;�������������,�����Ǵ�лֱ�ӵ��ؼ����ʺϳ�;��,�����Ǻ����Ǻϳ�ø���Ǻ�����ø����ĵͱ���,���ܿ��ƺ�����ʹ�乩����ƽ��,�Ӷ����¼����ʺϳ�;������,�ر��Ǽ����ʺϳ�ø������オ�Ͷ���ɼ����ʺ����½�,�õ������ÿɽ�һ������������Ƥ���ѡ��ᷢ�����ε�,���������������ٴ�,������ø���Ƽ��ܹ����ƿ����Ժ�Ĥ��������ຣ����ø���ԡ������ʺϳ�ͨ·��ػ�������ø�������������½�,���¼����ʺ�����������,�����߱��������������������Щ�����ֱ���,һ�����溣���Ǵ�л�Ĺ���ƽ�ⱻ����,��ֱ��Ӱ�쵽����ļ����ʺϳ������������Ƥ�뷢�����̡���������ǰ���ں����Ǵ�л���ؼ����ʺϳɷ���������о��ɹ�,Ϊ�������������ú�����ø���Ƽ��������Ǻϳ�ø���Ƽ�����ɫũҩ���κ����ṩ�������ݡ�
[��������: 2]
[��������: 3]
[��������: 3]
[��������: 3]
[��������: 3]
[��������: 1]
[��������: 2]
Magsci [��������: 2]
��Ŀ�ġ������Ǻϳ�ø��trehalose-6-phosphate synthase, TPS���Dz�������Ѫ��-�����ǺϳɵĹؼ�ø�����о�ּ�ڿ�¡�¹�С� <em>Blattella germanica </em>TPS�����о�TPS�����ڵ¹�Сͬ��֯�еı���ģʽ���ڲ�ͬ�¶ȴ����µı����������������ͨ��RACE������¡�¹�С�TPS����ȫ�����У�����ӫ�ⶨ��PCR�ķ������TPS�����ڵ¹�С�5���׳治ͬ��֯�еı���ģʽ���ڸ���(40���46�洦��30 min)������(0���10�洦��1 h)�澳�µı������仯����������ӵ¹�С��п�¡���2��TPS���ֱ�����Ϊ <em>BgTPS</em>1 (GenBank��¼�ţ�KR050213) �� <em>BgTPS</em>2 (GenBank��¼�ţ�KR050214)�����У�<em>BgTPS</em>1����cDNA����ȫ��2 987 bp�������Ķ��� (ORF) 2 502 bp������833�������<em>BgTPS</em>2����cDNA����ȫ��3 212 bp�������Ķ���2 469 bp������822�������ᡣ<em>BgTPS</em>1��<em>BgTPS</em>2������Ҫ��5���׳�֬�����б����<em>BgTPS</em>2����ı�����Ϊ<em>BgTPS</em>1�����������3.9���������ֲ�ͬ�����¶��յ��£�<em>BgTPS</em>1��<em>BgTPS</em>2����mRNA���ϵ�������У�<em>BgTPS2 </em>�ı�����ʼ���������� <em>BgTPS</em>1����0��ʱ��<em>BgTPS</em>1��<em>BgTPS</em>2�ı�������ߡ������ۡ��¹�С�5���׳��д���2��TPS��������TPS�������֬�����и߱����<em>BgTPS</em>2����ı�������������<em>BgTPS</em>1�����º����յ��¾��ܴٽ���������ı������������ý��Ϊ��һ����ȷ���溣���ǵĺϳ�;��������������¶��澳�ķ�Ӧ�е������о��춨�˻�������
Magsci [��������: 2]
��Ŀ�ġ������Ǻϳ�ø��trehalose-6-phosphate synthase, TPS���Dz�������Ѫ��-�����ǺϳɵĹؼ�ø�����о�ּ�ڿ�¡�¹�С� <em>Blattella germanica </em>TPS�����о�TPS�����ڵ¹�Сͬ��֯�еı���ģʽ���ڲ�ͬ�¶ȴ����µı����������������ͨ��RACE������¡�¹�С�TPS����ȫ�����У�����ӫ�ⶨ��PCR�ķ������TPS�����ڵ¹�С�5���׳治ͬ��֯�еı���ģʽ���ڸ���(40���46�洦��30 min)������(0���10�洦��1 h)�澳�µı������仯����������ӵ¹�С��п�¡���2��TPS���ֱ�����Ϊ <em>BgTPS</em>1 (GenBank��¼�ţ�KR050213) �� <em>BgTPS</em>2 (GenBank��¼�ţ�KR050214)�����У�<em>BgTPS</em>1����cDNA����ȫ��2 987 bp�������Ķ��� (ORF) 2 502 bp������833�������<em>BgTPS</em>2����cDNA����ȫ��3 212 bp�������Ķ���2 469 bp������822�������ᡣ<em>BgTPS</em>1��<em>BgTPS</em>2������Ҫ��5���׳�֬�����б����<em>BgTPS</em>2����ı�����Ϊ<em>BgTPS</em>1�����������3.9���������ֲ�ͬ�����¶��յ��£�<em>BgTPS</em>1��<em>BgTPS</em>2����mRNA���ϵ�������У�<em>BgTPS2 </em>�ı�����ʼ���������� <em>BgTPS</em>1����0��ʱ��<em>BgTPS</em>1��<em>BgTPS</em>2�ı�������ߡ������ۡ��¹�С�5���׳��д���2��TPS��������TPS�������֬�����и߱����<em>BgTPS</em>2����ı�������������<em>BgTPS</em>1�����º����յ��¾��ܴٽ���������ı������������ý��Ϊ��һ����ȷ���溣���ǵĺϳ�;��������������¶��澳�ķ�Ӧ�е������о��춨�˻�������
[��������: 6]
[D].
[��������: 1]
[D].
[��������: 1]
DOI:10.3864/j.issn.0578-1752.2019.03.007Magsci [��������: 2]
<p id="C2"><b>����</b> ���溣���Ǻϳ�ø���������溣���Ǻϳɵ���Ҫ����,�����������ֻӵ��һ��������-6-����ϳ�ø��trehalose-6-phosphate synthase,TPS������,�����������һ��������-6-������ø��trehalose-6-phosphate phosphatase,TPP������ǰ���о����ַֺ�ʭ��<italic>Nilaparvata lugens</italic>����ӵ������<italic>TPS</italic>,���书���о�����<italic>TPS</italic>�����ܹ����غ����Ǵ�л,���ɽ鵼������ø���ؼ����ʺϳ��뽵��;��,�����������Ƥ���̡�<b>Ŀ��</b> ͨ���Ժַ�ʭת¼�������������һ���µ�<italic>TPS</italic>,���û����ںַ�ʭ��ͬ�����εı������,̽���û���Ĺ�����ǰ�ڷ��ֵ�����<italic>TPS</italic>������<b>����</b> �Ի�õ���<italic>TPS</italic>�������в��ÿ�¡�������ȫ��cDNA����,����֤��ȷ��,���䵰��һ���������������ṹ�������������TPS���бȶԷ���,������ʵʱӫ�ⶨ��PCR��qRT-PCR�������ⶨ3����ͬ<italic>TPS</italic>�ںַ�ʭ��ͬ�����εı���,������RNA���ţ�RNAi����������<italic>TPS3</italic>�ı��<b>���</b> ��ǰ���о��Ļ�����,��¡��һ���µ�<italic>TPS</italic>,������Ϊ<italic>TPS3</italic>��<italic>TPS3</italic>�����Ķ���Ϊ2 352 bp,����783��������,Ԥ�⵰������Ϊ88.9 kD,�ȵ��Ϊ5.47,������ˮ�Խṹ��������Ϣѧ��������,�ַ�ʭ3��TPS�����нϸߵ�ͬԴ��,������TPS��TPP�������ؽṹ��������������,����<italic>α</italic>-������<italic>β</italic>-�۵������������ռ�ı�����Ϊ�ӽ����ַ�ʭ��ͬ������3��<italic>TPS</italic>����Ա�������ͬ,<italic>TPS1</italic>����Ա�������4��0 h��ʼ������,���ɳ�δﵽ���,<italic>TPS2</italic>����Ա�������4��ĩ�ڿ�ʼ����������������5��ζ��нϸߵı���,<italic>TPS3</italic>����Ա�������5��ĩ�ںͳɳ���ڽϸߡ��������źַ�ʭ<italic>TPS3</italic> 48 h���Ż������Ա������½�,ds<italic>TPS3</italic>����Ч����<italic>TPS3</italic>�ı��<b>����</b> �ںַ�ʭ�з���һ���µ�<italic>TPS</italic>��<italic>TPS3</italic>��,����ַ�ʭ���Ѿ�������TPS1��TPS2���нϸߵ�ͬԴ�ԡ���ͬ�����α���������,3��<italic>TPS</italic>�ڷ�����������ʹ�Ĺ��ܲ�ͬ��RNAi�ܹ���Ч����<italic>TPS3</italic>�ı��ﲢ���ºַ�ʭ��Ƥ�ϰ��ͳᷢ�����Ρ�</p>
DOI:10.3864/j.issn.0578-1752.2019.03.007Magsci [��������: 2]
<p id="C2"><b>����</b> ���溣���Ǻϳ�ø���������溣���Ǻϳɵ���Ҫ����,�����������ֻӵ��һ��������-6-����ϳ�ø��trehalose-6-phosphate synthase,TPS������,�����������һ��������-6-������ø��trehalose-6-phosphate phosphatase,TPP������ǰ���о����ַֺ�ʭ��<italic>Nilaparvata lugens</italic>����ӵ������<italic>TPS</italic>,���书���о�����<italic>TPS</italic>�����ܹ����غ����Ǵ�л,���ɽ鵼������ø���ؼ����ʺϳ��뽵��;��,�����������Ƥ���̡�<b>Ŀ��</b> ͨ���Ժַ�ʭת¼�������������һ���µ�<italic>TPS</italic>,���û����ںַ�ʭ��ͬ�����εı������,̽���û���Ĺ�����ǰ�ڷ��ֵ�����<italic>TPS</italic>������<b>����</b> �Ի�õ���<italic>TPS</italic>�������в��ÿ�¡�������ȫ��cDNA����,����֤��ȷ��,���䵰��һ���������������ṹ�������������TPS���бȶԷ���,������ʵʱӫ�ⶨ��PCR��qRT-PCR�������ⶨ3����ͬ<italic>TPS</italic>�ںַ�ʭ��ͬ�����εı���,������RNA���ţ�RNAi����������<italic>TPS3</italic>�ı��<b>���</b> ��ǰ���о��Ļ�����,��¡��һ���µ�<italic>TPS</italic>,������Ϊ<italic>TPS3</italic>��<italic>TPS3</italic>�����Ķ���Ϊ2 352 bp,����783��������,Ԥ�⵰������Ϊ88.9 kD,�ȵ��Ϊ5.47,������ˮ�Խṹ��������Ϣѧ��������,�ַ�ʭ3��TPS�����нϸߵ�ͬԴ��,������TPS��TPP�������ؽṹ��������������,����<italic>α</italic>-������<italic>β</italic>-�۵������������ռ�ı�����Ϊ�ӽ����ַ�ʭ��ͬ������3��<italic>TPS</italic>����Ա�������ͬ,<italic>TPS1</italic>����Ա�������4��0 h��ʼ������,���ɳ�δﵽ���,<italic>TPS2</italic>����Ա�������4��ĩ�ڿ�ʼ����������������5��ζ��нϸߵı���,<italic>TPS3</italic>����Ա�������5��ĩ�ںͳɳ���ڽϸߡ��������źַ�ʭ<italic>TPS3</italic> 48 h���Ż������Ա������½�,ds<italic>TPS3</italic>����Ч����<italic>TPS3</italic>�ı��<b>����</b> �ںַ�ʭ�з���һ���µ�<italic>TPS</italic>��<italic>TPS3</italic>��,����ַ�ʭ���Ѿ�������TPS1��TPS2���нϸߵ�ͬԴ�ԡ���ͬ�����α���������,3��<italic>TPS</italic>�ڷ�����������ʹ�Ĺ��ܲ�ͬ��RNAi�ܹ���Ч����<italic>TPS3</italic>�ı��ﲢ���ºַ�ʭ��Ƥ�ϰ��ͳᷢ�����Ρ�</p>
[��������: 3]
[��������: 1]
[��������: 1]
DOI:10.3864/j.issn.0578-1752.2017.06.006Magsci [��������: 2]
��Ŀ�ġ����溣����ø�ܹ����ؼ����ʴ�л��������Ƥ���̡����о�ͨ��TRE���ﱻ���ƺ�,���ַ�ʭ(<em>Nilaparvata lugens</em>)��Ƥ״���������ʺ����������ʺϳ�ø��chitin synthase,CHS���ͼ�����ø��chitinase,Cht������������,̽����ͬ�ĺ�����ø��trehalase,TRE���ںַ�ʭ��Ƥ�жԼ����ʴ�л�ĵ������á�������������RNAi����,��ʵ����������Ⱥ�ַ�ʭΪ����,ͨ����������ע��˫��RNA��dsRNA���ֱ����Ƶ���������ø�����ͬʱ���ƶ��������ø����,ע��48 h��ͨ��Trizol����ȡ�ַ�ʭ��RNA,��ת¼�Լ��кϳɵ�һ��DNA�����ʵʱӫ�ⶨ��PCR��qRT-PCR���������û���ı������,ȷ��RNAiЧ�����������ط��ⶨ48 h�ַ�ʭ���弸���ʺ����仯������Ƥ���ѳ���������գ�������qRT-PCR���ַ�ʭCHS��Cht��mRNAˮƽ�ϵ���Ա������仯,����TRE�ڵ��ؼ����ʴ�л�е����á����������ע��dsGFP��Ƚ�,�����ע����ַ�ʭ���弸���ʺ��������½�,����dsTRE1���ע������Validamycinע����ʼ������½�,ͬʱ�ַ�ʭ������Ƥ���ѵ�����qRT-PCR�������ʾ����TRE��dsRNAע���û���ı��ﱻ����,���Dz���TRE�ı����л���������������TRE1-2��TRE2�ڸ�ע���鴦���±�����½�,dsTRE1s��TRE2�ı���Ҳ������Ч��,������dsTRE1���ע����ͺ�����ø���Ƽ�Validamycin����Ч�����ԣ�dsTREע��������CHS����Ч��������,Validamycin�ܹ���������CHS1��CHS1a�ڱ�Ƥ�еı���,��2��dsTRE1ע���CHS1����������,dsTRE1-2ע����Ƥ�е�CHS1a�ı���������Cht1��Cht8��dsTRE��ע���鼰Validamycin�����б����½��������½�,dsTRE1-1ע���Cht2��Cht5��������������dsTRE1-2ע���Cht1��Cht6��Cht8�����½�,Cht2��Cht4��������������dsTRE2��������Cht1��Cht8��Cht10�����½���Cht9��������������dsTRE1sע���,Cht1��Cht5���������½�,��Cht9��������������Validamycinע������10��������ø������ﶼ�������������½��������ۡ�TRE�ܹ�ͨ�����غַ�ʭ�����ʴ�л;�������Ƽ����ʵĺϳ�,�����Ϊ��չ��ɸѡ��Ч�ĺ�����ø���Ƽ����ƺַ�ʭ�Ⱥ����ṩ�������ݡ�
DOI:10.3864/j.issn.0578-1752.2017.06.006Magsci [��������: 2]
��Ŀ�ġ����溣����ø�ܹ����ؼ����ʴ�л��������Ƥ���̡����о�ͨ��TRE���ﱻ���ƺ�,���ַ�ʭ(<em>Nilaparvata lugens</em>)��Ƥ״���������ʺ����������ʺϳ�ø��chitin synthase,CHS���ͼ�����ø��chitinase,Cht������������,̽����ͬ�ĺ�����ø��trehalase,TRE���ںַ�ʭ��Ƥ�жԼ����ʴ�л�ĵ������á�������������RNAi����,��ʵ����������Ⱥ�ַ�ʭΪ����,ͨ����������ע��˫��RNA��dsRNA���ֱ����Ƶ���������ø�����ͬʱ���ƶ��������ø����,ע��48 h��ͨ��Trizol����ȡ�ַ�ʭ��RNA,��ת¼�Լ��кϳɵ�һ��DNA�����ʵʱӫ�ⶨ��PCR��qRT-PCR���������û���ı������,ȷ��RNAiЧ�����������ط��ⶨ48 h�ַ�ʭ���弸���ʺ����仯������Ƥ���ѳ���������գ�������qRT-PCR���ַ�ʭCHS��Cht��mRNAˮƽ�ϵ���Ա������仯,����TRE�ڵ��ؼ����ʴ�л�е����á����������ע��dsGFP��Ƚ�,�����ע����ַ�ʭ���弸���ʺ��������½�,����dsTRE1���ע������Validamycinע����ʼ������½�,ͬʱ�ַ�ʭ������Ƥ���ѵ�����qRT-PCR�������ʾ����TRE��dsRNAע���û���ı��ﱻ����,���Dz���TRE�ı����л���������������TRE1-2��TRE2�ڸ�ע���鴦���±�����½�,dsTRE1s��TRE2�ı���Ҳ������Ч��,������dsTRE1���ע����ͺ�����ø���Ƽ�Validamycin����Ч�����ԣ�dsTREע��������CHS����Ч��������,Validamycin�ܹ���������CHS1��CHS1a�ڱ�Ƥ�еı���,��2��dsTRE1ע���CHS1����������,dsTRE1-2ע����Ƥ�е�CHS1a�ı���������Cht1��Cht8��dsTRE��ע���鼰Validamycin�����б����½��������½�,dsTRE1-1ע���Cht2��Cht5��������������dsTRE1-2ע���Cht1��Cht6��Cht8�����½�,Cht2��Cht4��������������dsTRE2��������Cht1��Cht8��Cht10�����½���Cht9��������������dsTRE1sע���,Cht1��Cht5���������½�,��Cht9��������������Validamycinע������10��������ø������ﶼ�������������½��������ۡ�TRE�ܹ�ͨ�����غַ�ʭ�����ʴ�л;�������Ƽ����ʵĺϳ�,�����Ϊ��չ��ɸѡ��Ч�ĺ�����ø���Ƽ����ƺַ�ʭ�Ⱥ����ṩ�������ݡ�
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 3]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}