Biological Characteristics of Transcriptional Regulator GalR in Streptococcus suis Serotype 4
SUN Ke1,2, ZHU HaoDan1, HE KongWang1, WANG DanDan1, ZHOU JunMing1, YU ZhengYu1, Lü LiXin1, NI YanXiu,11 Institute of Veterinary Medicine, Jiangsu Academy of Agricultural Sciences, Nanjing 210014 2 College of Animal Science and Technology, Jilin Agricultural University, Changchun 130118
Abstract 【Objective】 The study was carried out to investigate the effect of the deletion of the transcriptional regulator GalR gene of Streptococcus suis type 4 virulent strain SH1510 on the biological characteristics of bacteria in order to further study GalR regulation of galactose metabolism pathway and its pathogenesis, so as to provide a theoretical basis.【Method】 The SH1510ΔGalR, an SS4 virulent strain SH1510 transcriptional regulator GalR gene deletion strain, was constructed by homologous recombination double-crossover. Then, a preliminary screening of the deletion strain was performed by the internal amplification primers I1/I2 of GalR gene, and further identified the deletion strain SH1510ΔGalR by PCR and Western blotting. We performed Gram staining on the parental strain SH1510 and the deletion strain SH1510ΔGalR to compare morphological differences. We configured the basal medium, added glucose, sucrose and D-galactose to culture the bacteria, and plotted the bacterial growth curve to compare the bacteria’s utilization of different sugars. The bacterial concentration of the parent strain and the deletion strain were adjusted to 2.5×10 9 CFU/mL, 5×10 8 CFU/mL, 1×10 8 CFU/mL and 2×10 7 CFU/mL, respectively, and BALB/c mice were intraperitoneally injected. Then, the lethality of the mice was observed, and the strain LD50 was calculated by using the Reed-Muench method. The parental strain and the deletion strain with a concentration of 2×10 7CFU/mL were mixed in an equal volume of 1﹕1 and intraperitoneally injected into BALB/c mice. After 24 h, brain, spleen and blood were counted on chloramphenicol-resistant and non-resistant THB plates to compare the colonization ability of bacteria in mice. The internal environment with the whole blood of healthy pigs was simulated, and then the parent strain and the deletion strain were separately and added to whole blood of healthy pigs containing SS4 antiserum, and incubated at 37℃ for 2 h for plate counting to compare the viability of bacteria in whole blood.【Result】 PCR was performed by using the internal detection primer I1/I2, and the result showed that the deletion strain SH1510ΔGalR was negative. Further, primers C1/C2, O1/C2, O2/C1, and O1/O2 were used to identify the deletion strain SH1510ΔGalR, and 1 056, 2 121, 2 094, and 3 147 bp fragments were amplified, respectively, and the results were in agreement with expectations. The results of Western blotting showed that the parental strain SH1510 could specifically bind to the crude rabbit anti-GalR polyclonal serum, and a single band appeared at 37 kD, but the deletion strain SH1510ΔGalR showed no band. The results of PCR and Western blotting indicated that the deletion strain SH1510ΔGalR was successfully constructed. The parent strain and the deletion strain were stained by Gram. The optical microscopic observation showed that the parent strain and the deletion strain were arranged in a chain, the length was similar, and the morphology was not significantly different. Under the growth conditions of glucose and sucrose as the sole glycogen, the growth of the parent strain and the deletion strain was similar; when the glycogen was D-galactose, the growth rate and OD600 value of the deletion strain were significantly lower than that of the parent strain. In addition, it could be seen from the highest OD600 value that the utilization rate of sugar by Streptococcus suis SH1510 was glucose>sucrose>D-galactose. In the mouse pathogenicity test, the LD50 of the parent strain and the deletion strain were 1×10 8 CFU and 1.62×10 8 CFU, respectively, and the pathogenicity of the deletion strain to the mouse was decreased by 1.62 times. In vivo competitive infection test results showed that the number of bacteria isolated from deleted strains in the brain, spleen and blood of the mice was much lower than that of the parent strains, and the difference was extremely significant (P<0.01). The whole blood survival test showed that the survival rate of the parent strain was 35.2%, the survival rate of the deletion strain was 27.3%, and the survival rate of the deletion strain SH1510ΔGalR in the whole blood was significantly lower than that of the parent strain SH1510 (P<0.05).【Conclusion】 In summary, the GalR gene could promote the utilization of galactose by Streptococcus suis serotype 4, and had direct or indirect regulation of the virulence of SH1510. Keywords:Streptococcus suis serotype 4;GalR;biological characteristics;virulence
PDF (793KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 孙珂, 祝昊丹, 何孔旺, 王丹丹, 周俊明, 俞正玉, 吕立新, 倪艳秀. 猪链球菌4型转录调控因子GalR的生物学特性[J]. 中国农业科学, 2019, 52(19): 3485-3494 doi:10.3864/j.issn.0578-1752.2019.19.017 SUN Ke, ZHU HaoDan, HE KongWang, WANG DanDan, ZHOU JunMing, YU ZhengYu, Lü LiXin, NI YanXiu. Biological Characteristics of Transcriptional Regulator GalR in Streptococcus suis Serotype 4[J]. Scientia Acricultura Sinica, 2019, 52(19): 3485-3494 doi:10.3864/j.issn.0578-1752.2019.19.017
0 引言
【研究意义】 细菌对环境的改变必须做出迅速的反应,营养的供给随时都可以发生变化,反复无常,要能得以幸存必需具有可以变换不同代谢底物的能力,细菌代谢与调控型因子可以让细菌更好的应对环境变化,帮助细菌生存和感染,所以对细菌代谢调控基因的研究有利于细菌致病机制的阐明。【前人研究进展】 猪链球菌(Streptococcus suis)是一种重要的猪病原菌,可导致猪关节炎、败血症、心内膜炎、脑膜炎、肺炎等病症,也可导致畜牧与兽医相关从业人员的发病及死亡[1,2,3]。根据细菌荚膜抗原,猪链球菌可分为33个血清型(1-31型,33型和1/2型),其中,猪链球菌2型被公认为是致病性最强血清型[4,5,6]。另外,在亚洲国家也经常出现3,4,5,7,8,1/2血清型[7,8,9,10];加拿大经常分离出3,4,8,22,1/2血清型[11]。猪链球菌4型(Streptococcus suis type 4,SS4)也是一种致病血清型,可导致人和动物发病甚至死亡。1968—1984年,荷兰分离出30株引起人脑膜炎的猪链球菌菌株,其中1株为SS4[12]。2009—2012年,泰国在患有猪链球菌病的人群中分离出了668株猪链球菌,血清型比例SS4为0.15%[13]。2001年,HAN于韩国不同生猪屠宰场406份扁桃体中分离出55株猪链球菌,SS4占5.5%[14]。PRUFER[15]将1996—2016年间从德国分离的711株猪链球菌进行血清学分型,结果1996—2004年间的189株分离株中出现19个猪链球菌血清型,其中SS4占10%;2015—2016年间的522株分离株中出现23个猪链球菌血清型,SS4占10.3%;将研究结果与猪链球菌的临床背景联系起来,SS4经常分离自具有呼吸道疾病和中枢神经系统疾病的患病猪,证明SS4可引起肺病和脑膜炎;将结果与毒力因子结合分析,SS4分离菌株高频率携带毒力因子mrp和sly。CHATURVEDI[16]等用4周龄仔猪为动物模型评价SS4的毒力,结果导致仔猪死亡。近几年来,在中国健康猪场和发病猪场均经常分离到SS4,且分离率逐渐上升[17],王娟[18]对2011—2014年间在广东省不同地区发病猪场分离的58株猪链球菌进行血清学分型,SS4血清型比例为3.45%。周明瑶[19]在苏南地区某健康猪场80份鼻拭子中检测到猪链球菌4型竟高达46.25%,这证明猪链球菌4型在我国某些地区已成为优势血清型,严重威胁猪群健康。当前,国内外对猪链球菌的研究主要集中在猪链球菌2型上[20],对SS4的研究较少,范围仅限于血清型分型PCR鉴定,对SS4毒力因子的研究还没有报道。江苏省农业科学院畜牧兽医研究所实验室前期通过比较蛋白组学分析猪链球菌4型强弱毒株的菌体蛋白表达谱,筛选到了16个差异表达蛋白,其中包括转录调控因子GalR。GalR是参与半乳糖转运和代谢的调控蛋白[21]。在大肠杆菌中,GalR可调控D-半乳糖代谢途径相关基因galETKM、galP、galS、mglBAC的转录[22]。肺炎链球菌中,在半乳糖作为糖源的生长环境下,galk是维持正常生长所必须的基因,转录因子GalR可作为galk的转录激活因子,促进半乳糖的代谢[23]。【本研究切入点】GalR蛋白是SS4强弱毒株中显著差异表达蛋白之一, 推测其可能与SS4的致病性相关。因此,通过同源重组的方法构建了SS4强毒株SH1510的GalR基因缺失株SH1510ΔGalR,对其生物学特性进行比较,包括细菌的形态、生长特性、糖的利用率、在猪全血中的存活能力及对BALB/c小鼠的致病性。【拟解决的关键问题】转录调控因子GalR对猪链球菌4型强毒株SH1510半乳糖代谢及毒力的影响。
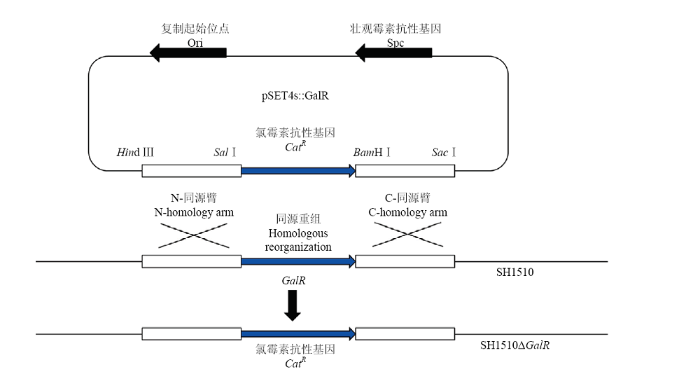
1.5.1 基因缺失质粒pSET4s::GalR的构建 基因缺失质粒pSET4s::GalR的构建如图1所示,使用引物L1/L2、R1/R2及SH1510 DNA模板扩增出GalR编码基因的上下游同源臂基因,使用引物C1/C2及pR326质粒为模板扩增氯霉素抗性基因。各扩增产物使用MiniBEST DNA Fragment Purification进行纯化回收,通过限制性内切酶对纯化后的DNA片段进行酶切,然后将酶切后的各产物进行纯化。最后利用DNA Ligation Kit,将3个片段以上游同源臂、氯霉素抗性基因、下游同源臂的顺序依次连接到pSET4s载体上,其间每一步通过转化进大肠杆菌DH5α进行大量克隆。每一步连接过后的质粒送上海英潍捷基生物有限公司进行测序,以确保质粒构建的准确性。
M:DNA分子质量标准;1、3、5、7、9:菌株SH1510;2、4、6、8、10:菌株SH1510ΔGalR。1、2:引物I1/I2;3、4:引物C1/C2;5、6:引物O1/C2;7、8:引物O2/C1;9、10:引物O1/O2 Fig. 2PCR identification of the deletion strain SH1510ΔGalR
M: DL5000 DNA Marker; 1, 3, 5, 7, 9: Strain SH1510; 2, 4, 6, 8, 10: Strain SH1510ΔGalR. 1, 2: Primer I1/I2; 3, 4: Primer C1/C2; 5, 6: Primer O1/C2; 7, 8: Primer O2/C1; 9, 10: Primer sO1/O2
2.2 Western blotting 鉴定
Western blotting 鉴定结果可见图3,亲本株SH1510可与粗制GalR多抗兔血清发生特异性结合,在37 kD处出现单一条带。缺失株SH1510ΔGalR不能发生反应,没有条带出现,结果表明缺失株SH1510ΔGalR构建成功。
Table 2 表2 表2BALB/c小鼠致死率和半数致死量 Table 2Mortality and LD50 in BALB/c mice
菌株 Strain
攻毒剂量 Challenge dose (CFU)
半数致死量 LD50/CFU
2.5×109
5×108
1×108
2×107
SH1510
10/10*
10/10
5/10
0/10
1×108
SH1510ΔGaIR
10/10
9/10
3/10
0/10
1.62×108
NaCl
0/10
0/10
0/10
0/10
The molecules represent the number of deaths in each group of mice and the denominator represents the total number of mice in each group *分子表示每组小鼠死亡数,分母表示每组小鼠总数
DU FS, LVQ, DUAND, CHENL, HUANG JH, WANG LP . Antimicrobial susceptibility and emergence of vancomycin resistance genes in Streptococcus suis. Animal Husbandry and Veterinary Medicine, 2018,50(12):32-36. (in Chinese) [本文引用: 1]
FAULDS-PAINA, SHAW HA, TERRA VS, KELLNERS, BROCKMEIER SL, WREN BW . The Streptococcos suis sortases SrtB and SrtF are essential for disease in pigs Microbiology, 2019,165(2):163-173. [本文引用: 1]
KONGD, CHENZ, WANGJ, LVQ, JIANGH, ZHENGY, XUM, ZHOUX, HAOH, JIANGY . Interaction of factor H-binding protein of Streptococcus suis with globotriaosylceramide promotes the development of meningitis Virulence, 2017,8(7):1290-1302. [本文引用: 1]
LIUJ, CHEN SS, ZHANGY, ZHONG XJ, PAN ZH, YAO HC . Influence of SstF on bacitracin resistance and virulence inStreptococcus suis serotype 2 Animal Husbandry and Veterinary Medicine, 2018(06):74-79. (in Chinese) [本文引用: 1]
XIA XJ, WANGL, CHENG LK, SHEN ZQ, LI SG, WANG JL . Expression and immunological evaluation of elongation factor Tu of Streptococcus suis serotype 2 Polish Journal of Veterinary Sciences, 2017,20(2):277-284. [本文引用: 1]
SHENX, LIUH, LIG, DENGX, WANGJ . Silibinin attenuates Streptococcus suis serotype 2 virulence by targeting suilysin Journal of Applied Microbiology, 2019,126(2):435-442. [本文引用: 1]
KATAOKAY, SUGIMOTOC, NAKAZAWAM, MOROZUMIT, KASHIWAZAKIM . The epidemiological studies of Streptococcus suis infections in Japan from 1987 to 1991 Journal of Veterinary Medical Science, 1993,55(4):623-626. [本文引用: 1]
WUC . Study on epidemiology characterization of Streptococcus suis and hp0197 genetic diversity analysis [D]. Wuhan: Huazhong Agricultural University, 2013. ( in Chinese) [本文引用: 1]
HAN DU, CHOIC, HAM HJ, JUNG JH, CHO WS, KIMJ, HIGGINSR, CHAEC . Prevalence, capsular type and antimicrobial susceptibility of Streptococcus suis isolated from slaughter pigs in Korea Canadian Journal of Veterinary Research-revue Canadienne de Recherche Veterinaire, 2001,65(3):151-155. [本文引用: 1]
WANY . The involvement of SntA protein in pathogenesis of Streptococcus suis serotype 2 [D]. Wuhan: Huazhong Agricultural University, 2017. ( in Chinese) [本文引用: 1]
GOTTSCHALKM, LACOUTURES, BONIFAITL, ROYD, FITTIPALDIN, GRENIERD . Characterization of italic> Streptococcus suis isolates recovered between 2008 and 2011 from diseased pigs in Quebec, Canada Veterinary Microbiology, 2013,162(2/4):819-825. [本文引用: 1]
ARENDS JP, ZANEN HC . Meningitis caused by Streptococcus suis in humans Reviews of Infectious Diseases, 1988,10(1):131-137. [本文引用: 1]
KERDSINA, AKEDAY, TAKEUCHID, DEJSIRILERTS, GOTTSCHALKM, OISHIK . Genotypic diversity of Streptococcus suis strains isolated from humans in Thailand European Journal of Clinical Microbiology & Infectious Diseases, 2018,37(5):917-925. [本文引用: 1]
HAN DU, CHOIC, HAM HJ, JUNG JH, CHO WS, KIMJ, HIGGINSR, CHAEC . Prevalence, capsular type and antimicrobial susceptibility of Canadian Journal of Veterinary Research, 2001,65(3):151-155. [本文引用: 1]
PRUFER TL, ROHDEJ, VERSPOHLJ, ROHDEM, DE GREEFFA, WILLENBORGJ, VALENTIN-WEIGANDP . Molecular typing of Streptococcus suis strains isolated from diseased and healthy pigs between 1996-2016 PLoS ONE, 2019,14(1):e210801. [本文引用: 1]
CHATURVEDI VK, SINGH DP, SRIVASTAVA AK . Pathogenicity of Streptococcus suis serotype 4 in piglets Veterinary Record, 1999,145(15):435-436. [本文引用: 1]
LIUQ, WANGJ, ZHOU YR, JIA AQ, WANG GP . The Streptococcus suis epidemiological analysis of healthy and infected swine in Guangdong Province China Animal Husbandry and Veterinary Medicine, 2017,44(6):1825-1831. (in Chinese) [本文引用: 1]
WANGJ . Epidemiologic investigation of Streptococcus suis in some areas of guangdong province from 2014 to 2015 [D]. Nanjing: Nanjing Agricultural University, 2015. ( in Chinese) [本文引用: 1]
ZHOU MY . The epidemiological investigation of Streptococcus suis isolated from health pigs in southern Jiangsu province in 2015 [D]. Nanjing: Nanjing Agricultural University, 2016. ( in Chinese) [本文引用: 1]
LIL, HUANG LZ, XIEB, ZHANG HL, GU WJ . Isolation, identification and gene sequence analysis of Streptococcus suis serotype 9 strain from Guangdong China Animal Husbandry and Veterinary Medicine, 2018(04):1016-1026. (in Chinese) [本文引用: 1]
GEANACOPOULOSM, ADHYAS . Functional characterization of roles of GalR and GalS as regulators of the gal regulon Journal of Bacteriology, 1997,179(1):228-234. [本文引用: 1]
QIANZ, TROSTELA, LEWIS DE, LEE SJ, HEX, STRINGER AM, WADE JT, SCHNEIDER TD, DURFEET, ADHYAS . Genome-wide transcriptional regulation and chromosome structural arrangement by GalR in E. coli Frontiers in Molecular Biosciences, 2016,3(4):156-164. [本文引用: 2]
AFZALM, SHAFEEQS, MANZOORI, KUIPERS OP . GalR acts as a transcriptional activator of galKT in the presence of galactose in Streptococcus pneumoniae Journal of Molecular Microbiology and Biotechnology, 2015,25(6):363-371. [本文引用: 3]
TAKAMATSUD, OSAKIM, SEKIZAKIT . Thermosensitive suicide vectors for gene replacement in Streptococcus suis Plasmid, 2001,46(2):140-148. [本文引用: 2]
TANG YL . Functional analysis of Sortases, CcpA and SodA of Streptococcus suis type 2 in relation to pathogenicity [D]. Hangzhou: Zhejiang University, 2012. ( in Chinese) [本文引用: 1]
WANGY . Biological functional of transcription regulator Rex in Streptococcus suis type 2 [D]. Nanjing: Nanjing Agricultural University, 2017. ( in Chinese) [本文引用: 1]
ZHENG CK . Contribution of the Spx regulators and NADH oxidase to the pathogenicity of Streptococcus suis serotype 2 [D]. Wuhan: Huazhong Agricultural University, 2017. ( in Chinese) [本文引用: 1]
NGUYEN CC, SAIER MJ . Phylogenetic, structural and functional analyses of the LacI-GalR family of bacterial transcription factors Federation of European Biochemical Societies Letter, 1995,377(2):98-102. [本文引用: 1]
XU ZM . The screening of STSLS related Streptococcus suis transcription factors, and its regulatory mechanism [D]. Wuhan: Huazhong Agricultural University, 2018. ( in Chinese) [本文引用: 1]
REN SJ . Research on identification and function of Type Ⅱ toxin-antitoxin system in Streptococcus suis serotype 2 [D]. Wuhan: Huazhong Agricultural University, 2017. ( in Chinese) [本文引用: 1]
 ,11
,11 
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT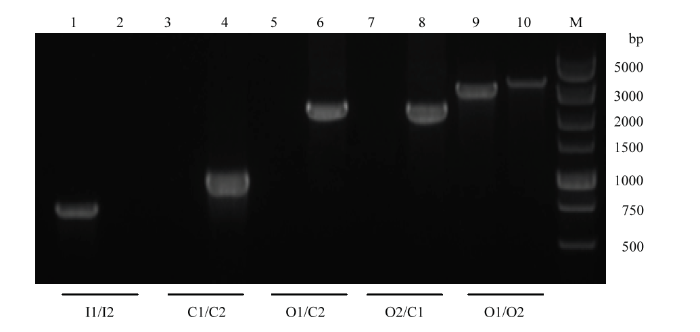 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT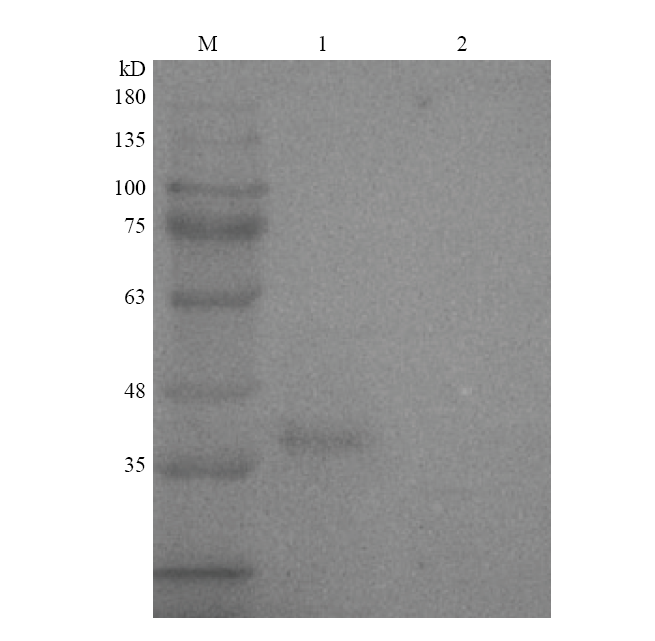 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT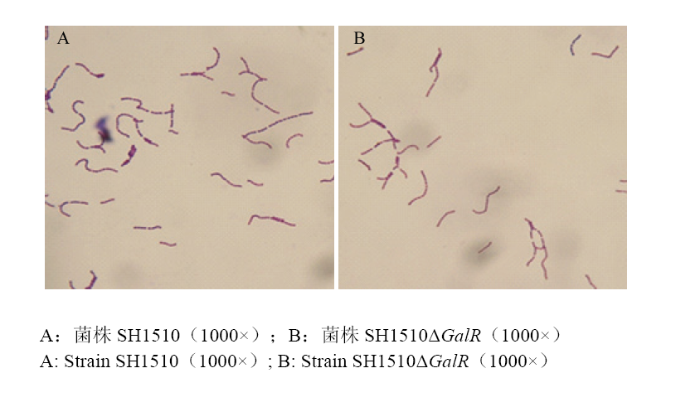 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT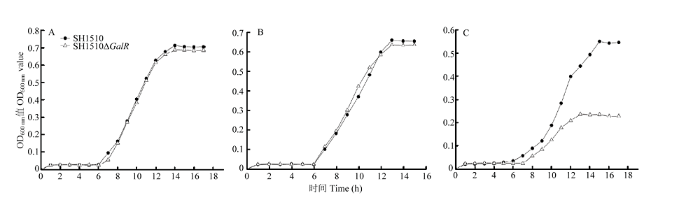 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT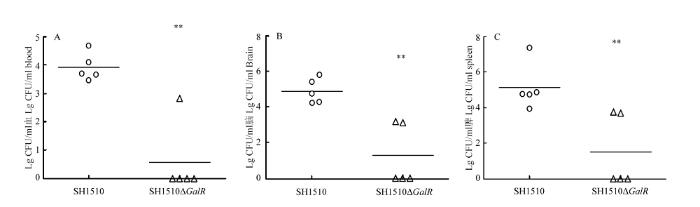 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT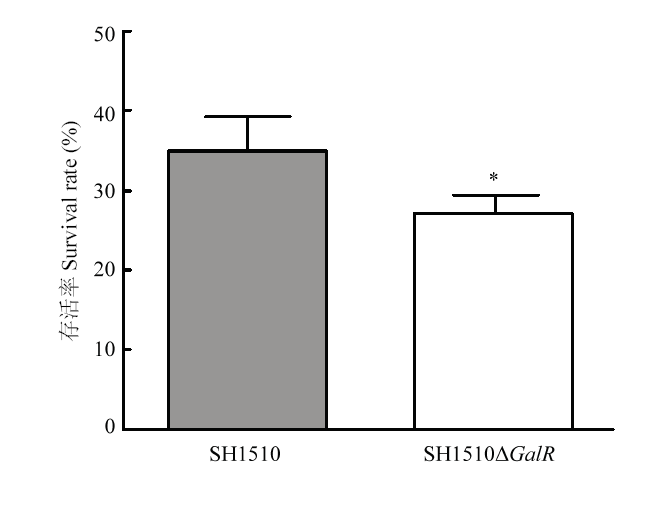 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}