 ,1
,1Coefficient of Parentage Analysis Among Naked Barley Varieties in Qinghai-Tibet Plateau
LI Jian1,2, FENG XianHong2, CAI YiLin,1通讯作者:
收稿日期:2019-04-3接受日期:2019-06-8网络出版日期:2019-08-16
| 基金资助: |
Received:2019-04-3Accepted:2019-06-8Online:2019-08-16
作者简介 About authors
李健,E-mail:lijian5028@163.com。

摘要
关键词:
Abstract
Keywords:
PDF (967KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
李健, 丰先红, 蔡一林. 青藏高原青稞育成品种的亲缘系数分析[J]. 中国农业科学, 2019, 52(16): 2758-2767 doi:10.3864/j.issn.0578-1752.2019.16.002
LI Jian, FENG XianHong, CAI YiLin.
0 引言
【研究意义】青稞既是青藏高原主要粮食作物,又是酿酒原料和牲畜饲料。在国际环境复杂多变的条件下,藏区“粮食安全”的实质就是以“青稞安全”为核心的食物安全[1]。青稞新品种选育工作,在藏区已经开展了近70年,以系统选择、杂交育种为技术支撑,培育了大批优良品种。由于长期对产量、品质、抗性等性状的连续定向选择,很可能造成育成品种的遗传基础日益狭窄,遗传多样性下降,不利于当前青稞新品种的选育[2,3]。【前人研究进展】近年来,利用分子标记(SSR、RAPD、SRAP)、同工酶、醇溶蛋白等技术,对青稞种质资源的遗传多样性进行了评估[4,5,6,7,8,9,10]。吴昆仑[4]研究认为,青藏高原青稞种质资源具有丰富的遗传多样性。曾兴权等[5]研究认为,青稞育成品种的平均遗传多样性高于西藏青稞地方品种的平均遗传多样性。孟凡磊等[9]、杨平等[10]研究认为西藏、四川的青稞育成品种遗传多样性较低,遗传基础相对狭窄。【本研究切入点】由于参试材料和研究方法的差异,研究结果不尽相同;虽然认为西藏、四川的青稞育成品种遗传基础相对狭窄,但对其成因还缺乏了解。其次,利用分子标记进行遗传多样性评价,如果存在标记数量少、密度低,与表型性状关联度低等缺陷,其结果的参考价值、实用价值有限,而大幅提升标记密度,同时强化与表型性状的关联度,又存在技术要求高、费用高的问题。亲缘系数分析是对遗传多样性进行评价的简便方法。近年来,对于自花授粉、谱系信息较为完整的作物,利用亲缘系数结合分子标记技术分析小麦、大麦、大豆、水稻、亚麻等作物遗传多样性,发现亲缘系数法与分子标记技术的分析结果具有较高的一致性[2,11-15]。另外,亲缘系数分析可以直观地展示育成品种遗传多样性的演变进程,揭示其成因,为品种选育工作提供参考。【拟解决的关键问题】青稞是高度自花授粉作物,且育成品种有较为完整的谱系信息。本研究选择1950s后藏区选育的105个推广面积较大、影响力较高的青稞育成品种以及3个引进品种,收集、整理其谱系信息,利用亲缘系数法分析青藏高原青稞育成品种的遗传多样性,了解其演变进程,探索其成因和改良对策,从而推动当前青稞新品种选育工作。1 材料与方法
1.1 材料
根据文献报道,选择1950年后,推广面积较大、影响力较高且谱系信息相对完整的藏区青稞育成品种105个,引进品种3个,作为参试品种(电子附表1)。1.2 方法
根据参试品种的谱系信息,参考王春江等[16]和刘章雄等[17]的方法,计算108个品种成对组合的亲缘系数(coefficient of parentage,COP)。计算原则如下:①一个品种分别从其双亲得到一半的基因;②所有祖先种亲本及其后代品种都是纯合的;③最早的祖先品种(系)间的亲缘系数为0;④混合授粉材料各花粉供体享有相同的等同于雌配子的亲缘系数概率,例如,A与5个父本混合授粉材料杂交得到B,则B与A的亲缘系数为1/2;B与各父本的亲缘系数均为1/2×5;⑤一个品种与其系选所得品种间的亲缘系数为0.75;⑥自然突变和诱导突变材料与其祖先的亲缘系数为0.75;⑦含有部分相同亲本的旁系品种间的亲缘系数计算公式为RSD=∑ (1/2)n1+n2;R代表品种S和品种D之间的亲缘系数,n1和n2分别代表S品种和D品种的共同亲本与品种S和品种D间的世代数;⑧一个品种与其自身的亲缘系数为1.00;⑨亲本与后裔品种间的亲缘系数可以表示其遗传物质的传递比率即遗传贡献率。根据参试品种的谱系信息,计算各品种的累积直系亲缘系数(cumulative direct coefficient of parentage,CD-COP)。首先分别计算该品种与其所有衍生种的COP系数,然后对该品种与其所有衍生种的COP系数求和。
利用DPS7.05软件,采用兰氏距离、类平均法,对COP矩阵(108×108)进行聚类分析。
2 结果
2.1 参试品种亲缘系数分析
108个参试品种形成5886对组合,其中,有1060对组合的COP值不为0,占比18.01%;各组合亲缘系数的变异范围为0.000—0.750,5 886对组合累积亲缘系数为314.146,平均值为0.053;品种间亲缘系数高于均值的组合数为771对,占比13.10%。2.1.1 各时期参试品种亲缘系数分析 以十年为期,将藏区青稞选育工作划分为7个时期。由于1950年前,藏区青稞育种工作进展极为缓慢,1950s的4个参试品种均为历史遗留的农家种,参试品种各组合间的COP值为0。1960s,参试品种数达到12个,形成78对品种间组合,组合累积亲缘系数为1.500,组合平均亲缘系数为0.019。1970s,参试品种数小幅增加至15个,形成组合120对,组合累积亲缘系数上升至13.750,组合平均亲缘系数骤然提升至各时期的最高水平,达到0.115。1980s,参试品种数达到26个,为各时期的最高值,形成351对组合,组合累积亲缘系数为37.516,组合平均亲缘系数小幅下降至0.107。1990s,参试品种数下降至17个,形成组合153对,组合累积亲缘系数陡然下降至6.078,组合平均亲缘系数也大幅下降至0.040。21世纪初,参试品种数为18个,形成组合171对,组合累积亲缘系数回升至13.469,组合平均亲缘系数大幅回升至0.079。最近十年,参试品种数为16个,形成组合136对,组合累积亲缘系数为4.414,组合平均亲缘系数为0.032,这两项指标均处于最近40年的历史最低值(图1)。
图1
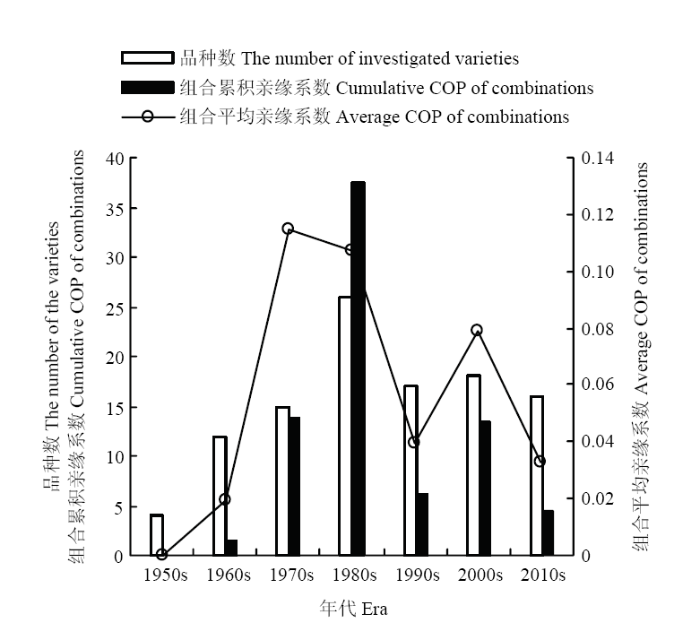 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1各时期参试品种亲缘系数分析
Fig. 1The COP analysis of the investigated varieties at each era
2.1.2 各地区参试品种亲缘系数分析 西藏自治区参试品种数达到48个,组合累积亲缘系数为110.969,是藏区参试品种最多,组合累积亲缘系数最高的地区。青海省参试品种数、组合累积亲缘系数位列藏区第二,分别为27个和46.160。计算各地区参试品种的组合平均亲缘系数,结果表明,甘肃省最高,达到0.220,西藏自治区、青海省、四川省的参试品种组合平均亲缘系数较为接近,分别为0.094、0.122和0.138(图2)。
图2
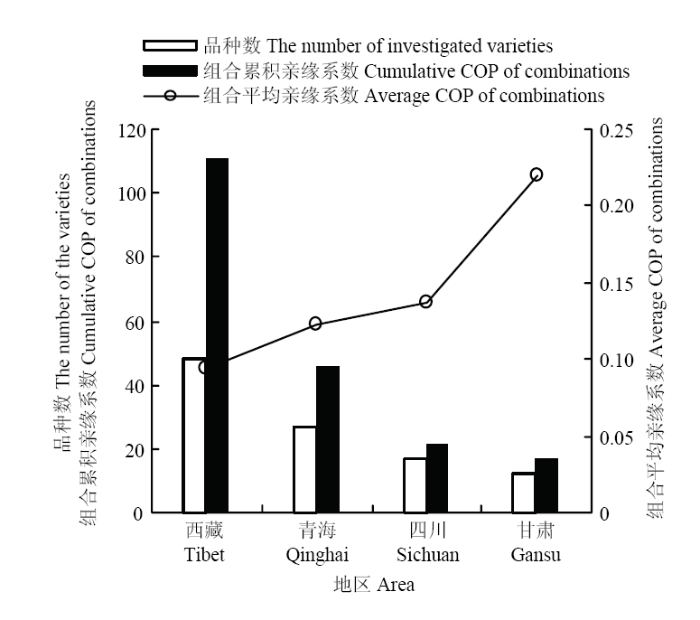 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2各地区参试品种亲缘系数分析
Fig. 2The COP analysis of the investigated varieties from each region
2.2 参试品种亲缘系数聚类分析
根据参试品种的亲缘系数,采用兰氏距离、类平均法进行聚类分析。108个参试品种聚为7个类群,不同类群间存在显著差异(电子附图1)。2.2.1 不同类群所包含品种呈现明显的地域差异 通过分析各类群所包含品种的地理来源,发现Ⅰ类群仅包含四川省、甘肃省的育成品种,占比分别为80.00%和20.00%;Ⅱ、Ⅲ类群所包含品种的地理来源呈多样化分布,西藏自治区、青海省的育成品种数占比较高,四川省、甘肃省、云南省的育成品种数占比较低;Ⅳ、Ⅴ类群以西藏自治区育成品种为主体,其中,Ⅴ类群所包含品种全部来源于西藏自治区;Ⅵ类群所包含品种以甘肃省育成品种为主体;Ⅶ类群50.00%的品种来自西藏自治区,40.00%的品种来自青海省,10.00%的品种来自四川省(图3)。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3各类群参试品种地理来源分布
Fig. 3The temporal distribution of investigated varieties that belong to seven groups
2.2.2 群内组合累积亲缘系数、平均亲缘系数差异显著 Ⅰ类群包含10个品种,群内组合累积亲缘系数为17.500,群内组合平均亲缘系数为0.318。Ⅱ类群包含12个品种,组合累积亲缘系数和组合平均亲缘系数均为0。Ⅲ类群包含13个品种,群内组合累积亲缘系数和平均亲缘系数都非常低,遗传多样性相对较高。Ⅳ、Ⅴ类群分别包含18个和11个品种,Ⅳ、Ⅴ类群的组合累积亲缘系数接近,但Ⅴ类群的组合平均亲缘系数为0.358,大幅高于Ⅳ类群。Ⅵ类群包含14个品种,组合累积亲缘系数大幅升高,组合平均亲缘系数为0.391,是各类群的最高值;Ⅶ类群包含30个品种,群内组合累积亲缘系数高达116.930,组合平均亲缘系数为0.252(图4)。
图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4各类群参试品种亲缘系数分析
Fig. 4The COP analysis of the investigated varieties from each group
2.2.3 不同类群品种构成分析 Ⅱ、Ⅲ类群的群内组合平均亲缘系数较低,所包含品种以农家种或农家系选种、引进品种为主,其他少数品种为影响力较大、推广面积大但谱系信息部分缺失的品种。
Ⅰ、Ⅳ、Ⅴ、Ⅵ、Ⅶ类群的群内组合平均亲缘系数较高;群内所包含品种中,存在亲缘关系的品种数占比也较高。Ⅰ类群90.00%的品种与813和康青3号存在亲缘关系。Ⅳ类群66.67%的品种与南繁2号、果洛存在亲缘关系。Ⅴ类群所有品种与白玉紫芒、藏青336存在亲缘关系。Ⅵ类群所有品种与肚里黄存在亲缘关系。Ⅶ类群所有品种与昆仑1号、藏青稞存在亲缘关系。因此,Ⅰ、Ⅳ、Ⅴ、Ⅵ、Ⅶ类群均存在骨干亲本,各类群以各自的骨干亲本及其衍生种为主体,将83个参试品种划分为5类(图5)。
图5
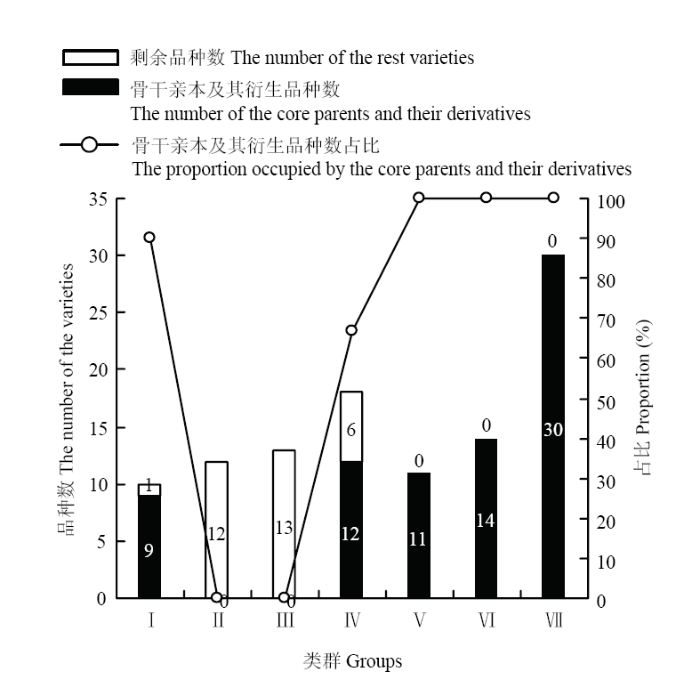 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5各类群骨干亲本及其衍生品种数占比
Fig. 5The proportion occupied by the core parents and their derivatives within each group
2.3 藏区青稞骨干亲本分析
根据各参试品种的累积直系亲缘系数,筛选出骨干亲本。结果表明,各参试品种的累积直系亲缘系数变异范围为0.000—7.563;其中,40个品种的累积直系亲缘系数不为0;昆仑1号、肚里黄、藏青336、藏青稞、白玉紫芒、果洛、康青3号、813、南繁2号的累积直系亲缘系数位居前列,是藏区主要的骨干亲本(电子附表2)。上述骨干亲本及其衍生种的数量合计达76个,占参试育成品种总量的72.38%。2.3.1 骨干亲本及其衍生种的时间分布 1960s,是藏区青稞品种选育的起步阶段,10年间,5个参试品种(藏青稞、藏青336、白玉紫芒、813和肚里黄)成为第一批骨干亲本,占该时期参试品种数的41.67%。1970s,育成昆仑1号和果洛2个骨干亲本,第一批骨干亲本迅速在藏区青稞品种选育活动中发挥显著作用。1970s,骨干亲本及其衍生种占该时期参试育成品种总量的80.00%。1980s是参试育成品种集中出现的时期,该时期育成青稞品种26个,其中21个是由9个骨干亲本及其衍生种组成,占比80.77%。随着时期的推移,9个骨干亲本及其衍生种占同时期参试育成品种总量的比例呈逐期下降趋势,但是降幅非常小。其后的3个时期,9个骨干亲本及其衍生种数量占同时期参试品种总量的比例分别为76.47%、72.22%和68.75%(图6)。1970s—1990s,以9个骨干亲本为亲本选育而成的品种数占参试育成品种总量的比例较高,分别达到66.67%、61.54%和70.59%;21世纪初,44.44%的参试育成品种仍以9个骨干亲本为亲本。最近十年,虽然以9个骨干亲本为亲本选育而成的参试品种数占比下降至12.50%,但是仍然有56.25%的参试育成品种以9个骨干亲本为祖先种(图6和图7)。
图6
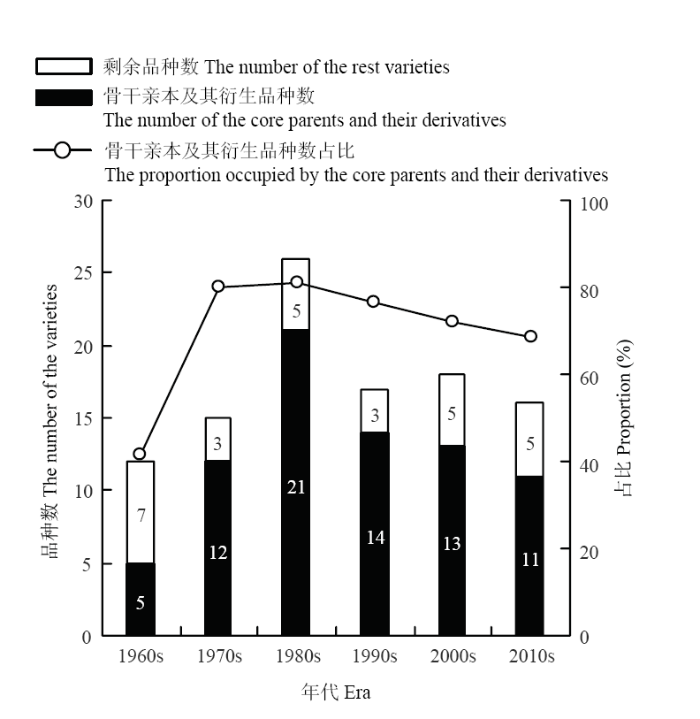 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6骨干亲本及其衍生品种数占各时期参试品种总量的比例
Fig. 6The proportion occupied by the core parents and their derivatives at each era
图7
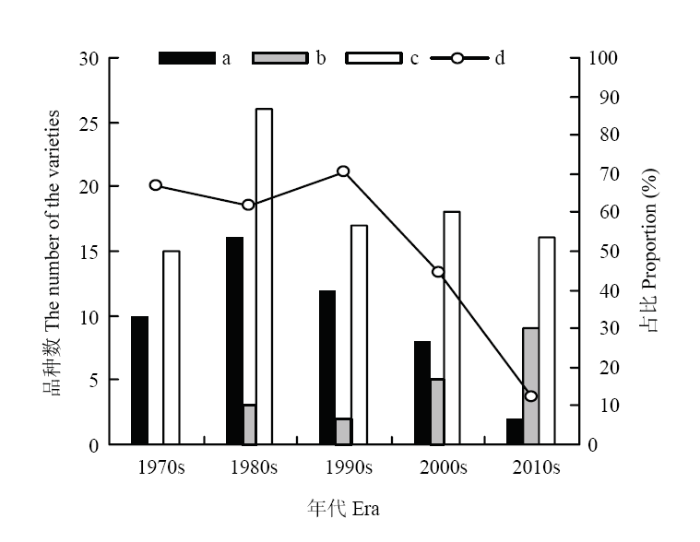 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7各时期以9个骨干亲本为亲本或祖先种选育而成的参试育成品种数
a:以9个骨干亲本为亲本选育而成的参试育成品种数;b:以9个骨干亲本为祖先种选育而成的参试育成品种数;c:各时期参试育成品种数;d:各时期以9个骨干亲本为亲本选育而成的品种数占参试育成品种总量的比例
Fig. 7The amount of varieties that were bred by using the nine core parents as parents or ancestors at each era
a: The amount of varieties that were bred by using the nine core parents as parents; b: The amount of varieties that were bred by using the nine core parents as ancestors; c: The amount of investigated varieties at each era; d: The percentage of the amount of varieties that were bred by using the nine core parents as parents to the amount of investigated bred varieties at each era
2.3.2 骨干亲本及其衍生种的地域分布 通过分析骨干亲本及其衍生种的地理来源,发现76个品种存在明显的地域划分,不同地域高频使用的骨干亲本也各不相同。西藏自治区育成的骨干亲本为藏青336、白玉紫芒、果洛和南繁2号,主要引进骨干亲本为昆仑1号和藏青稞;6个骨干亲本及其衍生品种数占西藏自治区参试育成品种数的79.17%。青海省育成的骨干亲本为昆仑1号和藏青稞,引进的骨干亲本为肚里黄,3个骨干亲本及其衍生品种数占青海省参试育成品种数的62.96%。四川省育成的骨干亲本为813和康青3号,引进的骨干亲本为昆仑1号和肚里黄,4个骨干亲本及其衍生品种数占四川省参试育成品种数的70.59%。甘肃省育成的骨干亲本为肚里黄,引进的骨干亲本为康青3号,2个骨干亲本及其衍生品种数占甘肃省参试育成品种数的75.00%(图8)。
图8
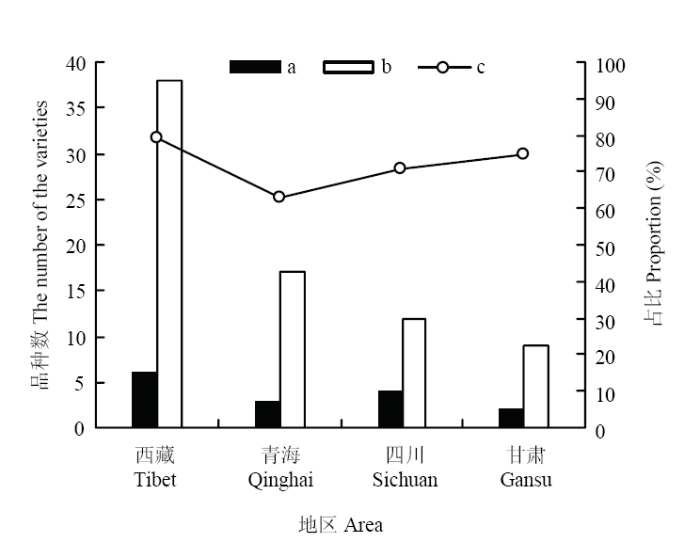 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8骨干亲本及其衍生品种占各地区参试育成品种总量的比例
a:常用骨干亲本数;b:骨干亲本及其衍生品种数;c:骨干亲本及其衍生品种数占各地区参试育成品种总量的比例
Fig. 8The proportion occupied by the core parents and their derivatives at each area
a: The amount of frequently-used core parents; b: The amount of core parents and their derivatives; c: The percentage of the amount of core parents and their derivatives to the amount of total varieties at each area
2.4 参试育成品种选育方法及选育周期分析
通过对参试育成品种的选育方法进行分析,结果表明,79.05%的参试育成品种是通过杂交育种技术选育而成,19.05%的参试育成品种是通过系统选育而成,仅有1.90%的参试育成品种是结合杂交育种技术和辐射诱变技术选育而成。利用系统选育技术选育而成的品种主要分布在1950s—1970s,占比达到80.00%。利用杂交育种技术选育而成的品种分布在1970s以后;结合辐射诱变技术和杂交育种技术选育而成的品种分布在1980s和1990s(图9)。图9
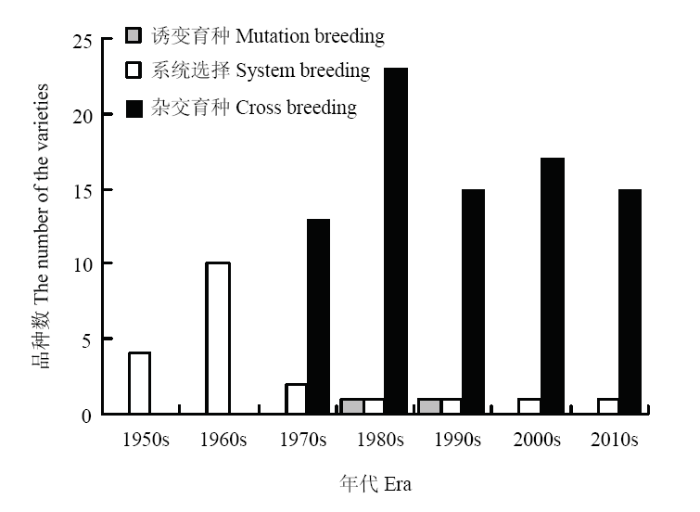 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9参试育成品种选育方法分析
Fig. 9The techniques used in the breeding process of the investigated varieties
52个参试品种包含选育周期信息。选育周期在5—10年的品种占比为25.00%,共13个品种,其中,76.92%的品种审定时间集中在1980s及1980s以前。选育周期在10—15年的品种占比为51.92%,共27个品种,其中,96.30%的品种审定时间集中在1980s及1980s以后。选育周期在15—20年的品种占比为23.07%,共12个品种,其审定时间全部集中在最近20年。
3 讨论
3.1 藏区青稞育成品种的遗传多样性分析
2018年,曾兴权等[18]对177份大麦种质资源进行重测序分析,其中包含69个藏区青稞地方品种和35个藏区青稞育成品种;结果表明,大麦野生种的遗传多样性最高,东、西方大麦种质资源的遗传多样性较高,而青稞地方品种和育成品种的遗传多样性最低,遗传基础相对狭窄。2016年,郭焕强等[19]对中国155个大麦主栽品种进行亲缘系数分析,结果表明,155个大麦品种的组合平均亲缘系数为0.013。2008年MIKEL等[20]对1970—2006年间育成的北美大麦品种进行亲缘系数分析,结果显示,363个大麦品种的组合平均亲缘系数为0.023,其中69个啤大麦品种的组合平均亲缘系数为0.111,274个饲用大麦品种的组合平均亲缘系数为0.020。与本研究结果相比,108个藏区青稞品种的组合平均亲缘系数为0.053,显著高于中国大麦主栽品种以及北美大麦品种的组合平均亲缘系数。因此,青藏高原青稞育成品种的遗传多样性低,遗传基础相对狭窄、脆弱。
3.2 藏区青稞育成品种遗传基础狭窄的成因及对策
3.2.1 骨干亲本在藏区青稞育种中的重要作用 在麦类作物育种工作中,骨干亲本都起到了不可替代的重要作用。阿夫、阿勃、欧柔、南大2419、矮孟牛、小偃6号等16个小麦骨干亲本,对中国小麦品种的选育做出了突出贡献,阿勃衍生了217个品种,阿夫衍生了165个品种[21]。1950年后,藏区选育的9个骨干亲本,在各个时期、各个地区的青稞选育工作中,都起到了不可替代的重要作用。1970s,以矮化育种为理论指导,以萧山立夏黄、沧州裸大麦、喜马拉1号为矮源与藏青稞、白玉紫芒等当地骨干亲本组配,培育了以昆仑1号为代表的突破性青稞品种,大幅提高了青稞单产[22]。同时,又以昆仑1号为亲本与当地骨干亲本组配,培育衍生品种4个,骨干亲本及其衍生种数量占1970s参试品种数的80%。1980s,仅以昆仑1号为骨干亲本选育而成的衍生种数量就达到8个,占同时期参试品种总量的30.77%;9个骨干亲本及其67个衍生种占同时期参试品种总量的80.77%。21世纪初,72.22%的参试育成品种仍然由9个骨干亲本及其衍生种构成,44.44%的参试育成品种仍然以9个骨干亲本为亲本。
藏区各地参试育成品种的谱系信息表明,西藏自治区79.17%的参试育成品种由6个骨干亲本(藏青336、藏青稞、白玉紫芒、昆仑1号、果洛和南繁2号)及其衍生种构成;青海省62.96%的参试育成品种由3个骨干亲本(昆仑1号、肚里黄和藏青稞)及其衍生种构成;四川省70.59%的参试育成品种由4个骨干亲本(813、肚里黄、昆仑1号和康青3号)及其衍生种构成;甘肃省75%的参试育成品种由2个骨干亲本(肚里黄和康青3号)及其衍生种构成。
由于9个本地骨干亲本在藏区青稞育种工作中的重要作用,使得藏区在青稞播种面积有所下降的情况下,青稞总产量到达解放初期的3倍以上,青稞平均单产到达解放初期的4倍左右。
但是,由于骨干亲本的高频、长期使用,客观上造成了藏区青稞育成品种的遗传多样性大幅下降,遗传基础趋于狭窄。1970s、1980s和21世纪初的3个时期,参试品种的组合平均亲缘系数到达0.115、0.107和0.079,是藏区青稞育成品种遗传多样性最低的3个时期。但是,1990s和最近十年,参试育成品种的组合平均亲缘系数分别为0.040和0.032,育成品种的遗传基础得以拓宽(图1)。通过对2个时期参试品种的构成进行分析,结果表明,育种目标的调整是2个时期参试品种遗传多样性提高的主要原因。1974年,藏区由单一的春青稞育种思路发展为冬青稞、春青稞并重的育种思路。1990s,优良冬青稞品种大量涌现,该时期38.88%的参试品种为冬青稞品种,使得参试品种的组合平均亲缘系数显著下降,藏区青稞育成品种的遗传基础得以拓宽。基于青稞育种的经验总结,矮秆、早熟很难实现高产;其次,21世纪初,藏区的农业基础设施、耕作水平大幅提升,防灾抗灾能力明显增强。因此,通过提高生物产量,延长灌浆时间以提高青稞单产的育种理论得以普及。最近十年,以藏青2000为代表的“中高秆”、“中晚熟”品种开始出现。由于育种理论的进一步发展,导致最近十年参试育成品种的组合平均亲缘系数大幅下降,藏区青稞育成品种的遗传基础得以再次拓宽。因此,加强青稞基础研究,适时调整育种目标,有利于青稞育成品种遗传基础的拓宽和突破性青稞品种的选育。
3.2.2 由于生态条件和最终用途的显著差异,外引大麦种质的利用率低 DAI等[23]利用DarT标记技术,对不同来源的大麦种质进行遗传多样性分析,结果表明,西藏大麦栽培种与西藏野生大麦的遗传相似性高于地中海地区的野生大麦和栽培大麦。ZENG等[24]通过对青稞地方品种“拉萨勾芒”的基因组进行测序并与Morex的基因组数据进行比对分析,结果显示,“拉萨勾芒”所包含的104个基因家系存在显著扩张,这些基因家系与大麦的抗逆性、抗寒性相关;信号通路分析显示这些基因存在于环境应答或环境适应信号通路之中。由于青藏高原特殊的自然环境,推动了本地青稞种质的适应性进化,而外引大麦品种则表现出较差的环境适应性,很难直接推广使用。1960s以后,没有一个外引大麦品种,在藏区进行大面积推广使用[25]。
由于籽粒最终用途的显著差异,导致外引大麦种质作为供体亲本而加以利用的频率很低。在北美、欧洲、澳洲以及国内其他地区,大麦主要作为酿酒原料和饲料[26],由于麸皮对大麦籽粒、麦芽具有一定的保护作用且裸粒性状为隐性性状,因而皮大麦种质的数量远高于裸大麦种质[27,28,29]。根据中国作物种质信息网的查询结果,中国国家种质库中保存的6 452份大麦种质中,皮大麦种质的数量占比高达91.00%。在青藏高原,青稞籽粒的最终用途是食品原料,因而,裸粒性状是刚性育种目标。裸大麦与皮大麦组配,将有大量不符合育种目标的有麸后代产生,降低了选育效率;增加了皮大麦种质在藏区青稞育种工作中的利用难度。即便引入裸大麦种质,在藏区特殊的生态条件下,多棱型品种的产量表现普遍优于二棱型品种。因此,育种人员更倾向于使用多棱型大麦种质与本地青稞种质组配;否则,育种效率会显著下降。105个藏区育成品种所使用的亲本种质为118个,分析各亲本种质的地理来源信息,结果表明,82.20%的亲本属于藏区本地青稞种质,5.09%的种质原产地信息不详,从国内外其他地区引进的种质仅占12.71%。引进的大麦种质中,多棱裸大麦种质、二棱皮大麦种质、二棱裸大麦种质的数量占比分别为6.67%、13.33%和33.34%。
因此,由于适应性和最终用途的显著差异,外引大麦种质在藏区青稞育种工作中的使用频率很低,是导致藏区青稞育成品种遗传多样性较低的重要原因。
CHEN等[30]通过筛选120个代表性小麦品种并对其进行全基因组重测序分析,结果表明,现代中国小麦育成品种与欧洲小麦地方种质的遗传相似性甚至高于与中国小麦地方种质的遗传相似性;由此可见,高效利用外引种质,对于现代麦类作物育种的重要作用。随着育种技术的不断发展,利用分子标记辅助育种技术,“speed breeding”快速加代技术等先进育种手段[31],提高外引优异突变与藏区本地种质的融合速度,充分发挥外引种质在遗传改良过程中的作用,是拓宽藏区青稞育成品种遗传基础,推动藏区青稞新品种选育的重要途径。
4 结论
藏区青稞的9个骨干亲本及其67个衍生品种构成了育成品种的主体,外引大麦种质利用率低,从而导致青藏高原青稞育成品种的遗传多样性较低,遗传基础相对狭窄、脆弱。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOI:10.7606/j.issn.1009-1041.2011.06.006Magsci [本文引用: 2]

为了明确青藏高原青稞种质资源的遗传多样性,并为青稞育种提供依据,利用分布于青稞14条染色体长、短臂上的14对SSR引物对来自青藏高原区域内西藏、青海、四川和云南的55份青稞材料的遗传多样性进行了分析。结果表明,每个位点检测出的等位基因数为2~10个,共检测出总位点数48个,平均3.7个,各多态位点检测出基因型为1~21种,Nei’s基因多样性指数为0~0.4998,平均为0.2921,Shannon信息指数为0~0.6930,平均为0.4461,表明青藏高原青稞种质资源具有丰富的遗传多样性。不同生态区青稞材料在遗传上存在较大的差异,云南参试材料的Nei’s基因多样性指数最高,Shannon信息指数最低,是四个区域中遗传多样性最丰富的地区。聚类分析将材料分为五组,部分材料的来源与所研究的SSR引物位点相关性状存在独立性,但总体上基于SSR多态性的聚类与材料来源地区存在一定的相关性。
DOI:10.7606/j.issn.1009-1041.2011.06.006Magsci [本文引用: 2]
为了明确青藏高原青稞种质资源的遗传多样性,并为青稞育种提供依据,利用分布于青稞14条染色体长、短臂上的14对SSR引物对来自青藏高原区域内西藏、青海、四川和云南的55份青稞材料的遗传多样性进行了分析。结果表明,每个位点检测出的等位基因数为2~10个,共检测出总位点数48个,平均3.7个,各多态位点检测出基因型为1~21种,Nei’s基因多样性指数为0~0.4998,平均为0.2921,Shannon信息指数为0~0.6930,平均为0.4461,表明青藏高原青稞种质资源具有丰富的遗传多样性。不同生态区青稞材料在遗传上存在较大的差异,云南参试材料的Nei’s基因多样性指数最高,Shannon信息指数最低,是四个区域中遗传多样性最丰富的地区。聚类分析将材料分为五组,部分材料的来源与所研究的SSR引物位点相关性状存在独立性,但总体上基于SSR多态性的聚类与材料来源地区存在一定的相关性。
DOI:10.7606/j.issn.1009-1041.2013.02.009Magsci [本文引用: 2]
为了评价西藏青稞种质资源的遗传多样性,用260对SSR引物对来自青藏高原主要农区的75份青稞育成品种、17份野生大麦、39份青稞地方品种和44份国外大麦材料的遗传多样性进行了分析。结果表明,从260对SSR引物中筛选出23对多态性高的引物,占筛选引物的8.1%。23对SSR引物在西藏野生大麦中共扩增出稳定、清晰的条带92条,多态性条带为81条,其比例为88.04%;在西藏青稞地方品种中共扩增出稳定、清晰的条带89条,多态性条带为78条,其比例为87.6%;在青稞育成品种中共扩增出稳定、清晰的条带109条,多态性条带98条,其比例为89.9%;在引进材料中共扩增出稳定、清晰的条带64条,多态性条带53条,其比例为82.8%。遗传多样性分析结果为青稞育成品种(0.9882)>西藏野生大麦(0.8033)>西藏青稞地方品种(0.5820)>国外大麦材料(0.4218)。聚类与主坐标分析表明,实验材料可以清楚的分为4类,西藏野生大麦分在两个类群,西藏青稞地方品种分在两个类群,引进大麦材料分在一个类群,青稞育成品种分在四个类群。同一来源地区的青稞育成品种遗传基础较为狭窄,不同地区间的青稞育成品种遗传差异较大。以上结果说明,在西藏青稞新品种选育和遗传改良中,在发掘和利用西藏野生大麦资源、西藏青稞地方品种资源的同时,更应该加强不同地区青稞育成品种资源的交换和配合使用。
DOI:10.7606/j.issn.1009-1041.2013.02.009Magsci [本文引用: 2]
为了评价西藏青稞种质资源的遗传多样性,用260对SSR引物对来自青藏高原主要农区的75份青稞育成品种、17份野生大麦、39份青稞地方品种和44份国外大麦材料的遗传多样性进行了分析。结果表明,从260对SSR引物中筛选出23对多态性高的引物,占筛选引物的8.1%。23对SSR引物在西藏野生大麦中共扩增出稳定、清晰的条带92条,多态性条带为81条,其比例为88.04%;在西藏青稞地方品种中共扩增出稳定、清晰的条带89条,多态性条带为78条,其比例为87.6%;在青稞育成品种中共扩增出稳定、清晰的条带109条,多态性条带98条,其比例为89.9%;在引进材料中共扩增出稳定、清晰的条带64条,多态性条带53条,其比例为82.8%。遗传多样性分析结果为青稞育成品种(0.9882)>西藏野生大麦(0.8033)>西藏青稞地方品种(0.5820)>国外大麦材料(0.4218)。聚类与主坐标分析表明,实验材料可以清楚的分为4类,西藏野生大麦分在两个类群,西藏青稞地方品种分在两个类群,引进大麦材料分在一个类群,青稞育成品种分在四个类群。同一来源地区的青稞育成品种遗传基础较为狭窄,不同地区间的青稞育成品种遗传差异较大。以上结果说明,在西藏青稞新品种选育和遗传改良中,在发掘和利用西藏野生大麦资源、西藏青稞地方品种资源的同时,更应该加强不同地区青稞育成品种资源的交换和配合使用。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2016.00180Magsci [本文引用: 1]
<p>利用92个SSR标记对108份青稞亲本材料进行多态性扫描,分析其遗传多样性,旨在寻找与农艺性状相关联的分子标记,为青稞杂交组合的配制及分子标记辅助育种提供依据。挑选48个多态性标记进行群体遗传结构分析,在此基础上采用Tassel 2.1 GLM (general linear model)和MLM (mixed linear model)方法进行标记与农艺性状的关联分析。共检测出156个等位变异,每个位点2~6个等位变异。供试群体的Shannon指数为<a name="OLE_LINK10">0.6727~1.1368</a>,材料间遗传相似系数为0.2250~1.0000,平均0.7585。通过群体遗传结构分析将供试材料划分成4个亚群。以GLM分析,发现12个与株高、穗长、穗粒数和分蘖数相关联的标记,<span>对表型变异的解释率分别为</span>11.5%~17.6%、19.4%~45.4%、15.4%~22.1%和29.2%;以MLM分析,发现8个与株高、分蘖数和小穗数相关的标记,各标记对表型变异的解释率分别为31.7%~49.8%、28.1%~37.2%、22.7%~32.7%。关联标记分布在基因组全部6个连锁群上。</p>
DOI:10.3724/SP.J.1006.2016.00180Magsci [本文引用: 1]
<p>利用92个SSR标记对108份青稞亲本材料进行多态性扫描,分析其遗传多样性,旨在寻找与农艺性状相关联的分子标记,为青稞杂交组合的配制及分子标记辅助育种提供依据。挑选48个多态性标记进行群体遗传结构分析,在此基础上采用Tassel 2.1 GLM (general linear model)和MLM (mixed linear model)方法进行标记与农艺性状的关联分析。共检测出156个等位变异,每个位点2~6个等位变异。供试群体的Shannon指数为<a name="OLE_LINK10">0.6727~1.1368</a>,材料间遗传相似系数为0.2250~1.0000,平均0.7585。通过群体遗传结构分析将供试材料划分成4个亚群。以GLM分析,发现12个与株高、穗长、穗粒数和分蘖数相关联的标记,<span>对表型变异的解释率分别为</span>11.5%~17.6%、19.4%~45.4%、15.4%~22.1%和29.2%;以MLM分析,发现8个与株高、分蘖数和小穗数相关的标记,各标记对表型变异的解释率分别为31.7%~49.8%、28.1%~37.2%、22.7%~32.7%。关联标记分布在基因组全部6个连锁群上。</p>
Magsci [本文引用: 2]
<P>采用内含子切接点引物(intron-splice junction primer)和长随机引物的PCR分子标记方法,对来自西藏主要农区的29个青稞育成品种和3个农家种的遗传多样性进行了分析。结果表明,选用的5个引物共扩增出稳定清晰的条带39条,其中26条(66.7%)为多态性条带。根据PCR扩增的条带建立[1,0]型数据矩阵,应用NTSYSpc2.1数据分析软件计算各品种间的遗传相似系数,并据此进行聚类分析,结果表明,西藏主要农区青稞品种间有一定的遗传差异,但总体遗传差异较小,遗传基础相对狭窄;聚类结果与各品种的实际生产利用情况较为接近,可以反映出这些青稞品种较为明显的区域特征。</P>
Magsci [本文引用: 2]
<P>采用内含子切接点引物(intron-splice junction primer)和长随机引物的PCR分子标记方法,对来自西藏主要农区的29个青稞育成品种和3个农家种的遗传多样性进行了分析。结果表明,选用的5个引物共扩增出稳定清晰的条带39条,其中26条(66.7%)为多态性条带。根据PCR扩增的条带建立[1,0]型数据矩阵,应用NTSYSpc2.1数据分析软件计算各品种间的遗传相似系数,并据此进行聚类分析,结果表明,西藏主要农区青稞品种间有一定的遗传差异,但总体遗传差异较小,遗传基础相对狭窄;聚类结果与各品种的实际生产利用情况较为接近,可以反映出这些青稞品种较为明显的区域特征。</P>
DOI:10.3724/SP.J.1005.2008.00115Magsci [本文引用: 2]
<P>利用SRAP(Sequence-related Amplified Polymorphism)分子标记技术, 对25份来自四川高原的青稞育成品种进行了遗传多样性研究。结果表明: 64对引物组合共检测出999条清晰条带, 62对可以获得多态性条带, 多态性引物组合占96.9%, 共产生225条多态性条带, 占总条带数的22.5%。64对引物组合共扩增出333种等位变异, 平均每个引物组合检测到5.20种等位变异。遗传多样性在0(me9/em14, me9/em15)~0.8928(me6/em18)之间, 平均为0.5126。聚类分析结果表明, 25份材料可分成A、B、C 3大类, 材料聚类与其来源地有明显的相关性。25份材料间的平均遗传距离较小(0.3240), 平均遗传多样性较低(0.5126), 遗传基础较为狭窄。</P>
DOI:10.3724/SP.J.1005.2008.00115Magsci [本文引用: 2]
<P>利用SRAP(Sequence-related Amplified Polymorphism)分子标记技术, 对25份来自四川高原的青稞育成品种进行了遗传多样性研究。结果表明: 64对引物组合共检测出999条清晰条带, 62对可以获得多态性条带, 多态性引物组合占96.9%, 共产生225条多态性条带, 占总条带数的22.5%。64对引物组合共扩增出333种等位变异, 平均每个引物组合检测到5.20种等位变异。遗传多样性在0(me9/em14, me9/em15)~0.8928(me6/em18)之间, 平均为0.5126。聚类分析结果表明, 25份材料可分成A、B、C 3大类, 材料聚类与其来源地有明显的相关性。25份材料间的平均遗传距离较小(0.3240), 平均遗传多样性较低(0.5126), 遗传基础较为狭窄。</P>
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
【目的】探讨山东省小麦品种的遗传多样性。【方法】对山东省建国以来历经9次更换的66个小麦品种的2145个组合进行了亲缘系数(coefficient of parentage, COP)分析。【结果】67.44%品种组合间存在亲缘关系,所有品种COP值变异范围为0.00~0.8203,平均为 0.06249,系谱亲缘系数高于平均值的占21.53%。随着时间的推移,各个年代之内品种间亲缘系数有逐渐增大的趋势,到90年代有所降低,然后又再增大。各年代内品种的亲缘系数普遍高于年代间品种的亲缘系数。早期的品种与以后年代的品种间的亲缘系数较小,中期和近期品种与以后年代的品种间的亲缘系数较大。同一个年代的品种,对后代的遗传贡献率,则随着年代的推移而逐渐减小。对品种间COP值按WAP法进行了聚类分析,除徐州438、齐大195、烟农78在COP为0处单独聚在一起外,其余63个品种可以聚为6个大类群,大多是同一年代的品种聚在一起,体现了不同年代的特点。山东省历史上年种植面积较大或推广时间较长的17个小麦品种,大都是较早利用引进亲本进行组配选育而成,亲本的亲缘系数较小,遗传差异较大。从系谱上看,山东省小麦的种质资源可基本上追溯到碧玉麦、Villa Glori(中农28)、洛夫林系、蚰子麦等四大骨干亲本体系。【结论】自50年代以来,品种间的遗传差异日趋狭窄,遗传多样性逐渐降低。
Magsci [本文引用: 1]
【目的】探讨山东省小麦品种的遗传多样性。【方法】对山东省建国以来历经9次更换的66个小麦品种的2145个组合进行了亲缘系数(coefficient of parentage, COP)分析。【结果】67.44%品种组合间存在亲缘关系,所有品种COP值变异范围为0.00~0.8203,平均为 0.06249,系谱亲缘系数高于平均值的占21.53%。随着时间的推移,各个年代之内品种间亲缘系数有逐渐增大的趋势,到90年代有所降低,然后又再增大。各年代内品种的亲缘系数普遍高于年代间品种的亲缘系数。早期的品种与以后年代的品种间的亲缘系数较小,中期和近期品种与以后年代的品种间的亲缘系数较大。同一个年代的品种,对后代的遗传贡献率,则随着年代的推移而逐渐减小。对品种间COP值按WAP法进行了聚类分析,除徐州438、齐大195、烟农78在COP为0处单独聚在一起外,其余63个品种可以聚为6个大类群,大多是同一年代的品种聚在一起,体现了不同年代的特点。山东省历史上年种植面积较大或推广时间较长的17个小麦品种,大都是较早利用引进亲本进行组配选育而成,亲本的亲缘系数较小,遗传差异较大。从系谱上看,山东省小麦的种质资源可基本上追溯到碧玉麦、Villa Glori(中农28)、洛夫林系、蚰子麦等四大骨干亲本体系。【结论】自50年代以来,品种间的遗传差异日趋狭窄,遗传多样性逐渐降低。
DOI:10.3724/SP.J.1006.2013.01693Magsci [本文引用: 1]
<p><span >追溯北京市</span><span >1983—2010</span><span >年</span><span >90</span><span >个大豆品种的系谱信息,计算其亲本间及品种组合间亲缘系数</span><span >(coefficient of parentage</span><span >,</span><span >COP)</span><span >并据此聚类,旨在探讨北京市育成大豆品种的遗传多样性特点。结果表明,</span><span >19</span><span >个品种的父母本间存在亲缘关系,占品种总数的</span><span >21.11%</span><span >;</span><span >90</span><span >个品种间共组成</span><span >4005</span><span >个组合,其中</span><span >63.62%</span><span >存在亲缘关系,</span><span >COP</span><span >平均值为</span><span >0.031</span><span >,表明大部分品种亲缘关系相对较远。随着时间的推移,品种间亲缘系数呈下降趋势,但从</span><span >2005</span><span >年</span><span >至</span><span >2010</span><span >年亲缘系数值增大。<span>除</span></span><span >12</span><span >个品种各自独为一类外,其余</span><span >78</span><span >个品种被聚成</span><span >10</span><span >类,从中可见不同年代的育种特征和品种的演化过程;中品</span><span >661</span><span >等</span><span >9</span><span >份种质对北京市大豆品种贡献较大,是骨干亲本。</span><span >北京市品种总体的遗传背景丰富,但随年代推移,遗传多样性有逐渐降低趋势。</span></p>
DOI:10.3724/SP.J.1006.2013.01693Magsci [本文引用: 1]
<p><span >追溯北京市</span><span >1983—2010</span><span >年</span><span >90</span><span >个大豆品种的系谱信息,计算其亲本间及品种组合间亲缘系数</span><span >(coefficient of parentage</span><span >,</span><span >COP)</span><span >并据此聚类,旨在探讨北京市育成大豆品种的遗传多样性特点。结果表明,</span><span >19</span><span >个品种的父母本间存在亲缘关系,占品种总数的</span><span >21.11%</span><span >;</span><span >90</span><span >个品种间共组成</span><span >4005</span><span >个组合,其中</span><span >63.62%</span><span >存在亲缘关系,</span><span >COP</span><span >平均值为</span><span >0.031</span><span >,表明大部分品种亲缘关系相对较远。随着时间的推移,品种间亲缘系数呈下降趋势,但从</span><span >2005</span><span >年</span><span >至</span><span >2010</span><span >年亲缘系数值增大。<span>除</span></span><span >12</span><span >个品种各自独为一类外,其余</span><span >78</span><span >个品种被聚成</span><span >10</span><span >类,从中可见不同年代的育种特征和品种的演化过程;中品</span><span >661</span><span >等</span><span >9</span><span >份种质对北京市大豆品种贡献较大,是骨干亲本。</span><span >北京市品种总体的遗传背景丰富,但随年代推移,遗传多样性有逐渐降低趋势。</span></p>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
<P><FONT face=Verdana>【目的】通过对大面积推广小麦品种和育种骨干亲本的全基因组多位点扫描分析,探讨两者之间的遗传关系及骨干亲本在小麦育种中的作用。【方法】利用ABI3730对中国66份大面积推广的小麦品种和13份育种骨干亲本的481个SSR位点进行基因型扫描分析。【结果】以基因型数据为基础的主坐标及NJ聚类分析,发现中国大面积推广品种可聚成6个大组,每一组中至少有一个骨干亲本。系谱分析表明,每一组中大面积推广品种和骨干亲本之间均存在密切的血缘关系,前者大多为骨干亲本的后裔;同一省份的品种多数聚在一起。来自同一组合的骨干亲本碧蚂4号含有的优势等位变异数多于大面积推广品种碧蚂1号,同样,郑引4号(St2422-464)多于郑引1号(St1472-506);新一代骨干亲本往往由上一代骨干亲本衍生而来;来源于同一组合的骨干亲本与大面积品种相比,前者与其组合中的骨干亲本间存在更大的遗传差异。【结论】只有对资源的创新、引进和利用给予足够的重视,育种工作才能取得大的突破。<BR></FONT></P>
Magsci [本文引用: 1]
<P><FONT face=Verdana>【目的】通过对大面积推广小麦品种和育种骨干亲本的全基因组多位点扫描分析,探讨两者之间的遗传关系及骨干亲本在小麦育种中的作用。【方法】利用ABI3730对中国66份大面积推广的小麦品种和13份育种骨干亲本的481个SSR位点进行基因型扫描分析。【结果】以基因型数据为基础的主坐标及NJ聚类分析,发现中国大面积推广品种可聚成6个大组,每一组中至少有一个骨干亲本。系谱分析表明,每一组中大面积推广品种和骨干亲本之间均存在密切的血缘关系,前者大多为骨干亲本的后裔;同一省份的品种多数聚在一起。来自同一组合的骨干亲本碧蚂4号含有的优势等位变异数多于大面积推广品种碧蚂1号,同样,郑引4号(St2422-464)多于郑引1号(St1472-506);新一代骨干亲本往往由上一代骨干亲本衍生而来;来源于同一组合的骨干亲本与大面积品种相比,前者与其组合中的骨干亲本间存在更大的遗传差异。【结论】只有对资源的创新、引进和利用给予足够的重视,育种工作才能取得大的突破。<BR></FONT></P>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}