 ,, 高志强山西农业大学农学院,山西太谷030801
,, 高志强山西农业大学农学院,山西太谷030801Effects of Rapeseed Green Manure on Soil Fertility and Bacterial Community in Dryland Wheat Field
LI WenGuang, YANG XiaoXiao, HUANG ChunGuo, XUE NaiWen, XIA Qing, LIU XiaoLi, ZHANG XiaoQi, YANG Si, YANG ZhenPing,, GAO ZhiQiangCollege of Agriculture, Shanxi Agricultural University, Taigu 030801, Shanxi通讯作者:
收稿日期:2019-03-15接受日期:2019-05-29网络出版日期:2019-08-01
| 基金资助: |
Received:2019-03-15Accepted:2019-05-29Online:2019-08-01
作者简介 About authors
李文广,Tel:18404983422;E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (1351KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
李文广, 杨晓晓, 黄春国, 薛乃雯, 夏清, 刘小丽, 张晓琪, 杨思, 杨珍平, 高志强. 饲料油菜作绿肥对后茬麦田土壤肥力及细菌群落的影响[J]. 中国农业科学, 2019, 52(15): 2664-2677 doi:10.3864/j.issn.0578-1752.2019.15.010
LI WenGuang, YANG XiaoXiao, HUANG ChunGuo, XUE NaiWen, XIA Qing, LIU XiaoLi, ZHANG XiaoQi, YANG Si, YANG ZhenPing, GAO ZhiQiang.
0 引言
【研究意义】黄土高原旱地是我国西北地区重要的耕地资源,但其土壤全氮含量和有机质含量相对较低,超过一半的土壤有机质含量低于1%[1]。为提高作物产量,往往过量化学氮肥被使用,最终导致土壤酸化板结,微生物多样性降低[2]。作为化学肥料零增长[3]计划和土壤有机质提升计划的重要替代肥源,还田后的绿肥在腐解过程中能释放大量养分[4],从而提高土壤肥力,增加作物产量[5,6],对农业可持续发展有着重要意义。土壤微生物作为农田生态系统的重要组成部分,对土壤生态变化十分敏感,易受气候、温度、土壤湿度及施肥等外界因素影响,是评价施肥和耕作制度改进土壤肥力和作物生产力的重要指标[7,8,9]。施化肥或绿肥均可改变土壤细菌群落结构及数量,进而导致土壤结构及肥力的改变[10]。饲料油菜作为绿肥优势明显,其植株生物量大,盛花期养分含量较其他绿肥高,其粗长的直根系能对土壤形成穿刺效应,破除犁底层,改善土壤通气状况,提高土壤有氧呼吸效率及酶活性[11,12]。因此,开展将饲料油菜作为绿肥还田提高土壤肥力、改善土壤细菌群落结构及其多样性研究具有重要意义。【前人研究进展】由于旱地小麦种植区年降水量不稳定,降水季节分布不均,过去常采用夏季休耕方式来稳定冬小麦产量[13],但该法会加重黄土高原地区土壤侵蚀。国内外一些研究均表明[14,15],夏闲期复种绿肥能有效提高旱地土壤蓄水效率,使小麦增产,可代替夏季休耕。李红燕等[16]研究发现,麦后复种长武怀豆和油菜并还田,均可显著提高黄土高原南部土壤有机质、速效磷含量及磷酸酶、蔗糖酶和脲酶活性。何红霞等[17]研究发现,休闲期种植长武怀豆能有效培肥黄土高原旱地土壤肥力。持续干旱条件可能使旱地小麦种植区土壤微生物生物量逐渐下降,而绿肥还田不仅可以增加土壤蓄水能力,还能有效增加土壤微生物活性[18,19]。LONGA等[20,21]发现,绿肥是土壤微生物多样性的重要来源,绿肥还田能显著改变土壤微生物群落组成及丰富度。【本研究切入点】本试验所在旱地小麦种植区,雨水多集中于麦后休闲期,该时期光热资源充足,但该区土壤肥力偏低,不合理复种会导致土壤肥力持续下降。利用饲料油菜作绿肥能充分利用麦后休闲期的水、热、土资源,提高土壤使用可持续性。目前,对绿肥的研究多集中于品种间比较及其对土壤理化性质的影响,鲜有研究将饲料油菜引入北方旱地麦田,探讨还田管理方式对土壤培肥及合理农作的影响。【拟解决的关键问题】本研究应用16S rRNA 基因Illumina MiSeq 高通量测序技术分析细菌群落及代谢功能变化情况,探讨后茬麦田土壤养分、酶活性变化,以期明确饲料油菜还田对土壤养分、酶活性和细菌群落的影响,确定最佳播量及还田时期,为提高旱地麦田土壤肥力,维护微生物生态系统和旱地农业可持续发展提供科学依据。1 材料与方法
1.1 研究区域与样地概况
试验于2015年6月至2016年6月在山西省运城市垣曲县长直乡鲁家坡村十倾园(111°43.3′E,35°14.4′N)进行。该区年均气温13.5℃,年均日照时数2 026.2 h,年积温4 900℃,全年无霜期236 d左右,降水集中在5—9月份,年降水量600—800 mm,年蒸发量1 200 mm,是典型旱作雨养农业区,属暖温带半湿润大陆性季风气候。供试土壤质地为中壤土,土壤类型属于褐土性红立黄土,pH为8.0。饲料油菜播前测得0—20 cm耕层土壤基本性质为:有机质11.51 g·kg-1,全氮0.87 g·kg-1,全磷0.81 g·kg-1,碱解氮41.13 mg·kg-1,速效磷14.58 mg·kg-1。1.2 供试品种
饲料油菜品种:华油杂62号,长势中等、含油量高,抗病抗倒伏能力强,由华中农业大学油菜研究室提供。小麦品种:烟农21号,具有较强的抗寒、抗旱和抗倒伏能力,由侯马市金色农田有限公司提供。
1.3 试验设计
前茬小麦收获后,复种饲料油菜,考察合理的种植密度;后茬小麦播种前,将饲料油菜还田,考察适宜的还田时间。试验采用二因素裂区设计:主区为播量(S),设置7.5 kg·hm-2(小播量,S1)、15 kg·hm-2(中播量,S2)和22.5 kg·hm-2(大播量,S3)3个播量水平。副区为还田时期(D),设置9月10日(早期还田,D1)、9月20日(中期还田,D2)和9月30日(晚期还田,D3)3个不同还田时期,以常规农户的麦后复种夏玉米模式(玉米收获后秸秆全量还田)作为对照。各处理3个小区重复,共计30个小区,各小区面积为15 m×44 m=660 m2。2016年6月10日麦收后按试验设计复种饲料油菜,条播,行距20 cm。于9月10日、9月20日、9月30日3次对设定小区分别进行饲料油菜翻压还田,10月9日采用施肥播种一体机种植冬小麦,条播,行距20 cm,种植密度112.5 kg·hm-2,所施氮磷钾复合肥用量分别为纯N 135 kg·hm-2、P2O5 165 kg·hm-2、K2O 37.5 kg·hm-2 ,施肥深度为20—40 cm。常规农户模式的麦后复种夏玉米种植采用机械化种肥同穴播种,肥料用量是氮磷钾复合肥750 kg·hm-2,肥料施于距种子10 cm处。
其他管理措施同一般大田生产。
1.4 土壤样品采集与处理
2017年6月1日小麦收割前采集土壤样品。每个试验小区取3个样方作为区内重复,每个样方内随机选取5个点,使用直径为4 cm的灭菌土钻采集0—20 cm的耕层土壤土样,将采集的5个样点的土样去除根系、动植物残体、石块等杂物后混合均匀迅速装入无菌自封袋,暂存于4℃冰盒,带回实验室后将混合土样分为3份,其中2份风干后分别用于土壤养分及酶活性测定,1份放置于-80℃冰箱保存用于土壤DNA提取。1.5 测定项目及方法
1.5.1 土壤养分及酶活性测定 采用常规方法[22]测定土壤化学性质:有机质采用重铬酸钾外加热法;碱解氮采用碱解扩散法;速效磷采用0.5 mol·L-1 NaHCO3浸提-钼锑抗比色法;速效钾采用1.0 mol·L-1 NH4OAc 浸提-火焰光度法测定;脲酶活性采用靛酚比色法测定;碱性磷酸酶活性采用磷酸苯二钠比色法测定;蔗糖酶活性采用3,5﹣二硝基水杨酸比色法测定[23]。1.5.2 饲料油菜还田生物量 还田当日,选取3个具有代表性的、面积为1 m2的样段,将收获鲜植株地上部分放入烘箱,于105℃杀青,30 min后再80℃烘至恒重,记录各样段干物质重。
1.5.3 细菌基因组DNA的提取及测序 采用Fast DNA SPIN Extraction Kits 试剂盒(MP Biomedicals,Santa Ana,CA,美国),提取土壤总DNA。使用NealDel- ND1000分光光度计(Thermo Fisher Scientific,Waltham,MA,USA)和琼脂糖凝胶电泳分别测定DNA浓度和纯度。
对所提取基因组DNA的16S rRNA V4区进行扩增,通用引物序列为:520F(AYTGGGYDTAAA GNG),802R(TACNVGGGTATCTAATCC)。PCR反应条件为:98℃预变性2 min;25个循环;98℃变性15 s;55℃退火30 s;72℃延伸30 s;72℃后延长5 min,于4℃保存。PCR扩增产物使用Agencourt AMPure Beads(Beckman Coulter,Indianapolis,IN)试剂盒进行纯化,使用PicoGreen dsDNA Assay Kit(Invitrogen,Carlsbad,CA,USA)检测试剂盒进行定量。样品委托上海派森诺生物科技有限公司采用Illumina MiSeq高通量测序技术平台进行序列测序和分析。
1.5.4 测序数据优化处理 对原始数据进行质量过滤和双端序列连接,运用QIIME(version 1.9.0,//qiime. org/)进行序列过滤,数据过滤标准为[24]:去除5′端引物错配碱基数>1的序列;去除含有N(模糊碱基)序列;去除含有连续相同碱基数>8的序列;去除长度≤150 bp的序列。运用MOTHUR软件(version1.31.2,http://www.mothur.org/) 中UCLHIME方法去除嵌合体序列,得到最终用于后续分析的优质序列。
1.5.5 OTU聚类分析及注释 在QIIME中调用UCLUST方法对优质序列按序列相似度97%进行聚类,选取每个类中最长的序列为代表序列。在QIIME中调用BLAST方法对序列数据库进行比对,获得每个OTU代表序列的分类学信息。注释数据库为:Greengenes(Release 13.8,http://greengenes. secondgenome.com/)。之后对OTU进行精简处理,去掉丰度值小于总序列数条数0.001%的OTU,得到后续分析使用的精简后的OTU列表。
1.5.6 PICRUSt功能预测 功能和代谢途径预测采用PICRUSt 软件进行分析[25],利用QIIME获得的closed OTU-table 与KEGG 数据库进行比对,获得不同细菌群落功能预测信息。
1.6 计算和统计方法
根据OTU列表中的各样品物种丰度情况,应用软件MOTHUR中的summary.Single命令,计算菌群Chao、ACE和Shannon指数[26,27]。使用QIIME软件,对Weighted的UniFrac距离矩阵进行UPGMA聚类分析,并使用R软件进行可视化。采用Excel 2003进行基本计算,并制作各水平群落分类学组成和差异分析图。采用SAS9.4软件的ANOVA过程进行二因素裂区设计方差分析及多重比较(文中多重比较含CK,方差分析不含CK),差异显著性水平为P<0.05;使用CANOCO4.5软件对土壤化学性质和细菌群落结构及多样性进行冗余分析(RDA)。2 结果
2.1 饲料油菜作绿肥还田对后茬麦田土壤养分及酶活性的影响
通过分析饲料油菜还田对后茬麦田土壤养分及酶活性变化情况(表1)可知。与常规种植方式相比,所有饲料油菜还田处理均改变了后茬麦田各样地土壤养分含量及酶活性,其中,土壤中速效磷、碱解氮和有机质含量的增加范围为3.15%—18.5%、3.33%—30.2%和11.73%—60.5%;脲酶、碱性磷酸酶和蔗糖酶活性的增加范围为3.12%—31.25%、4.84%—25.8%和1.4%—94.5%。各指标中,大播量晚期还田(S3D3)处理表现出明显优势,这可能与饲料油菜还田生物量(表1)及腐解速率有关。对9个饲料油菜还田处理作二因素方差分析,F检验表明,播量对土壤养分及酶活性的影响均达到极显著水平(P<0.01),还田时期仅对有机质及蔗糖酶的影响达到显著水平 (P<0.05),播量与还田时期互作则对土壤碱性磷酸酶活性的影响达到显著水平(P<0.05)。土壤养分及酶活性均随播量增加及还田时期推迟而增加,以S3与D3表现最优。Table 1
表1
表1饲料油菜播量(S)及还田时期(D)对后茬麦田土壤养分及酶活性的影响
Table 1
| 处理 Treatment | 自由度 DF | 饲料油菜生物量 Feed rape biomass (kg·hm-2) | 速效磷 Available P (mg·kg-1) | 碱解氮 Alkaline N (mg·kg-1) | 有机质 SOM (g·kg-1) | 脲酶 Urease (mg·g-1·24h-1) | 碱性磷酸酶 Alkaline phosphatase (mg·g-1·24h-1) | 蔗糖酶 Sucrase (mg·g-1·24h-1) | |
|---|---|---|---|---|---|---|---|---|---|
| S1 | D1 | 793.7c | 14.50b | 40.97d | 11.25cde | 0.29fg | 1.19ef | 0.74f | |
| D2 | 794.5c | 15.04b | 40.38d | 9.35e | 0.27g | 1.13f | 0.82f | ||
| D3 | 815.2c | 15.16b | 40.65d | 10.17de | 0.30efg | 1.30de | 0.84ef | ||
| S2 | D1 | 1367.4b | 15.49ab | 42.50cd | 12.86bcd | 0.35cd | 1.46ab | 0.98de | |
| D2 | 1379.0b | 16.03ab | 42.07d | 13.29bc | 0.33de | 1.42bc | 1.11cd | ||
| D3 | 1388.8b | 16.10ab | 44.28cd | 14.32b | 0.34cd | 1.33cd | 1.11cd | ||
| S3 | D1 | 1866.47a | 16.39ab | 46.88bc | 14.33b | 0.37bc | 1.47ab | 1.26bc | |
| D2 | 1854.3a | 16.60ab | 49.97ab | 15.05b | 0.40ab | 1.54ab | 1.37ab | ||
| D3 | 1938.7a | 17.28a | 53.55a | 18.47a | 0.42a | 1.56a | 1.42a | ||
| CK | 14.58b | 41.13d | 11.51cde | 0.32def | 1.24def | 0.73f | |||
| S1 | 1342.52a | 14.90b | 40.67b | 10.26c | 0.29c | 1.20c | 0.79c | ||
| S2 | 1349.51a | 15.87ab | 42.95b | 13.49b | 0.34b | 1.40b | 1.06b | ||
| S3 | 1374.01a | 16.76a | 50.13a | 15.95a | 0.39a | 1.52a | 1.35a | ||
| D1 | 801.16c | 15.46a | 43.45a | 12.81b | 0.34a | 1.37a | 0.99b | ||
| D2 | 1378.40b | 15.89a | 44.14a | 12.56b | 0.34a | 1.36a | 1.09a | ||
| D3 | 1886.48a | 16.18a | 46.06a | 14.32a | 0.35a | 1.39a | 1.12a | ||
| F值F value | |||||||||
| 播量S | 2 | 394.43** | 6.35** | 29.43** | 39.32** | 80.38** | 48.32** | 88.01** | |
| 还田时期D | 2 | 0.37 | 0.96 | 2.39 | 4.35* | 2.01 | 0.45 | 5.71* | |
| 播量×还田时期S×D | 4 | 0.34 | 0.73 | 1.34 | 2.93 | 2.25 | 4.02* | 0.12 | |
| 误差Error | 12 | ||||||||
| 总变异Total | 26 | ||||||||
新窗口打开|下载CSV
2.2 饲料油菜作绿肥还田对后茬麦田土壤细菌群落多样性的影响
饲料油菜还田提高了土壤细菌群落多样性(表2),播量、还田时期以及二者交互作用对Chao 1、ACE、Shannon指数及OTU的影响均达到显著(P<0.05)或极显著(P<0.01)水平。OTU及多样性指数均随饲料油菜播量增加而增加,S3显著高于S1与S2。随还田时期推后,OTU及多样性指数表现出先降低后增加的趋势。Table 2
表2
表2饲料油菜播量(S)及还田时期(D)对后茬麦田土壤细菌群落多样性的影响
Table 2
| 处理 Treatment | 自由度 DF | OTU | 多样性指数Diversity Index | ||||
|---|---|---|---|---|---|---|---|
| Chao 1 | ACE | Shannon | |||||
| S1 | D1 | 5490b | 6086b | 4875bc | 10.8abc | ||
| D2 | 4458g | 4994g | 3845f | 10.4d | |||
| D3 | 4708f | 5227f | 4188e | 10.6c | |||
| S2 | D1 | 5144d | 5782cd | 4518d | 10.7bc | ||
| D2 | 5016e | 5583de | 4515d | 10.6c | |||
| D3 | 4629f | 5141f | 4443d | 10.6c | |||
| S3 | D1 | 5291c | 5981bc | 4705c | 10.9ab | ||
| D2 | 5498b | 6194b | 4903b | 10.9ab | |||
| D3 | 5711a | 6443a | 5256a | 11.0a | |||
| CK | 5254c | 5491e | 4946b | 10.9ab | |||
| S1 | 4885c | 5435b | 4302c | 10.57b | |||
| S2 | 4929b | 5527b | 4492b | 10.63b | |||
| S3 | 5500a | 6206a | 4944a | 10.91a | |||
| D1 | 5308a | 5949a | 4699a | 10.76a | |||
| D2 | 4991c | 5590b | 4421b | 10.62b | |||
| D3 | 5016b | 5629b | 4619a | 10.73ab | |||
| F值F value | |||||||
| 播量S | 2 | 1952** | 88.82** | 79.04** | 20.63** | ||
| 还田时期D | 2 | 517** | 19.48** | 14.93** | 3.22* | ||
| 播量×还田时期S×D | 4 | 864** | 29.26** | 39.44** | 3.22* | ||
| 误差Error | 12 | ||||||
| 总变异Total | 26 | ||||||
新窗口打开|下载CSV
2.3 饲料油菜作绿肥还田对后茬麦田土壤细菌群落组成的影响及差异分析
2.3.1 门水平的细菌群落结构特征 各处理细菌群落在门分类水平上具有较高的多样性。共得到19个类群,其中相对丰度≥1%的有12个,分别为变形菌门(Proteobacteria,13.1%—28.5%)、放线菌门(Actinobacteria,15.2%—30.7%)、酸杆菌门(Acidobacteria,8.4%—21.4%)、厚壁菌门(Firmicutes,7.5%—21.6%)、芽单胞菌门(Gemmatimonadetes,4.7%—11.7%)、硝化螺旋菌门(Nitrospirae,0.8%—9.2%)、浮霉菌门(Planctomycetes,3.7%—6.1%)、绿弯菌门(Chloroflexi,3.6%—5.6%)、GAL15(0%—6.1%)、疣微菌门(Verrucomicrobia,1.5%—3.2%)、拟杆菌门(Bacteroidetes,0.7%—3.9%)、WS3(0.2%—2.4%),它们构成了土壤样品在门水平上的基本结构。其余7个门的平均占比均小于1%,共占1.9%。对门水平相对丰度前10的菌群进行方差分析(图1),结果表明,变形菌门相对丰度随播量增加而增加,S3D3显著高于其他样地(P<0.05);厚壁菌门、硝化螺旋菌门相对丰度随播量增加呈现降低趋势;CK的放线菌门相对丰度显著高于其他样地(P<0.05);饲料油菜还田显著提高土壤酸杆菌门、芽单胞菌门、硝化螺旋菌门、GAL15和疣微菌门的相对丰度(P<0.05)。
图1
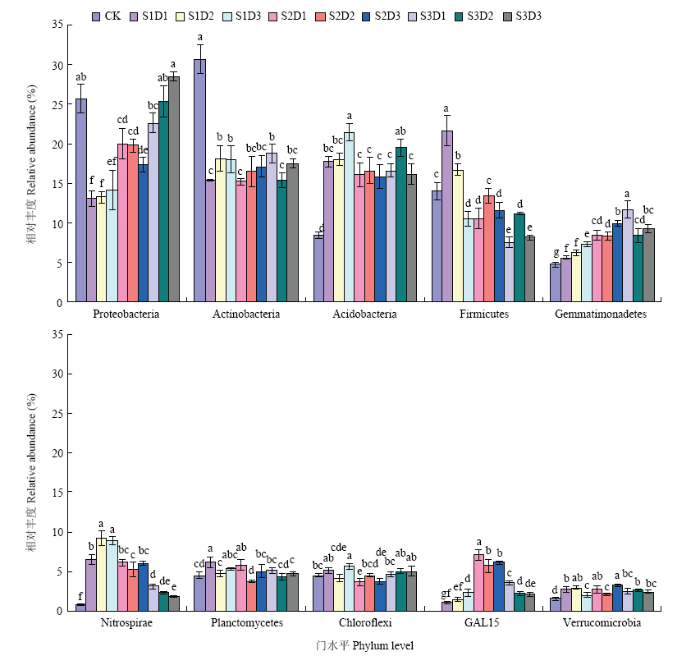 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同土壤样本相对丰度前10的菌群(门水平)
Fig. 1Relative abundances of the top 10 bacterial community for different soil samples (at the phylum level)
2.3.2 纲水平的微生物群落分布特征 各处理细菌群落隶属63个纲,相对丰度≥2%的有15个。其中相对丰度≥2%的有芽孢杆菌纲(Bacilli,7.5%—16.9%),酸杆菌纲(Acidobacteria,4.5%—14.7%),α-变形菌纲(Alphaproteobacteria,5.6%—12.4%),放线菌纲(Actinobacteria,5.2%—17.8%),嗜热油菌纲(Thermoleophilia,4%—8.7%),硝化螺旋菌纲(Nitrospira,0.8%—9.2%)和β-变形菌纲(Betaproteobacteria,2.5%—7.8%);其次为γ-变形菌纲(Gammaproteobacteria,1.5%—6.6%),芽单胞菌纲(Gemmatimonadetes,1.2%—6.2%),芽单孢菌门的Gemm-1纲(0.8%—5%)、GAL15纲(0—7%),δ 变形菌纲(Deltaproteobacteria,2.6%—3.8%),浮霉菌门的Planctomycetia纲(2.2%—3.8%),放线菌门的MB-A2-108纲(0.6%—4.1%)和酸杆菌门的Chloracidobacterial纲(1.3%—2.9%)。他们构成了纲水平上的基本结构。其余48纲平均相对丰度共占20%。
对纲水平相对丰度前10的细菌类群进行方差分析(图2)发现,饲料油菜还田显著增加了酸杆菌纲、硝化螺旋菌纲和芽单胞菌门下Gemm-1纲的相对丰度(P<0.05),显著降低了放线菌纲、嗜热油菌纲和β-变形菌纲的相对丰度(P<0.05);α-变形菌纲、β-变形菌纲、γ-变形菌纲和芽单孢菌纲的相对丰度随饲料油菜播量增加而增加,以S3D3处理的相对丰度最高。
图2
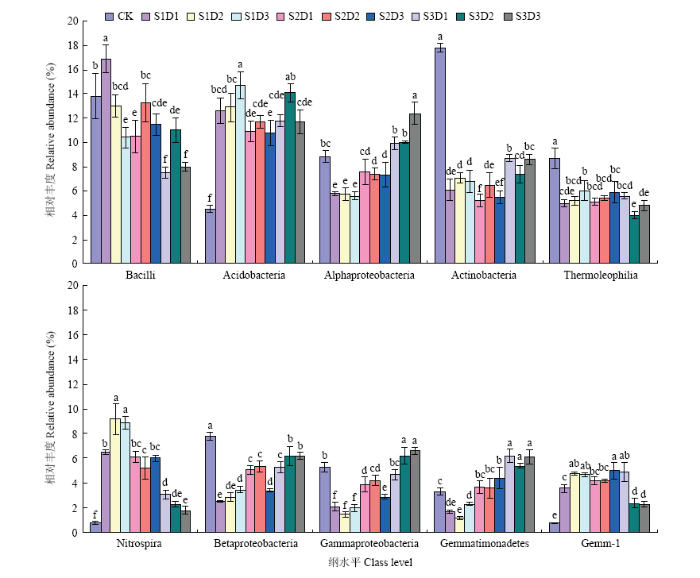 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同土壤样本前10个细菌类群的相对丰度(纲水平)
Fig. 2Relative abundances of the top 10 bacterial community for different soil samples (at the class level)
2.4 不同样本土壤细菌群落与土壤养分、酶活性之间的相关性
基于OTU层次的UPGMA聚类分析表明(图3),在0.075的相似性水平上,包括CK在内的不同样品可聚为4类,这表明,与CK相比,饲料油菜在不同播量及还田时期互作下还田改变了后茬麦田耕层土壤细菌群落结构。图3
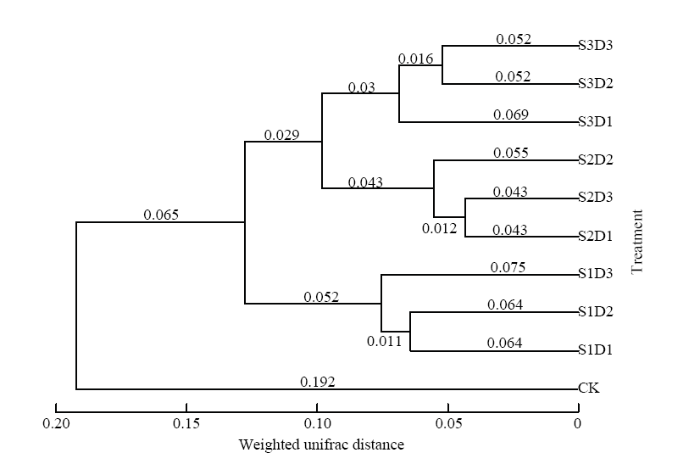 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同土壤样本基于UniFrac加权的细菌群落结构UPGMA聚类分析
Table 3The UPGMA cluster analysis based on weighted UniFrac for different soil samples bacterial community
通过对土壤养分、酶活性与细菌群落结构纲分类水平及多样性间的RDA冗余分析发现(图4),土壤养分和酶活性均与Betaproteobacteria、Gammaproteobacteria、Alphaproteobacteria、Gemmatimonadetes及细菌群落多样性间呈显著正相关;Acidobacteria与碱性磷酸酶、碱解氮、速效磷及蔗糖酶呈正相关;Actinobacteria 与脲酶及有机质呈正相关;Bacilli、Thermoleophilia、Nitrospira和Gemm-1则与土壤化学性质呈负相关。表明土壤养分、酶活性和细菌群落三者间密切相关,在土壤物质能量循环中均发挥着重要作用。饲料油菜作绿肥的合理还田模式有利于促进土壤酶活性提高、改善土壤细菌群落多样性,进而提高土壤肥力。
图4
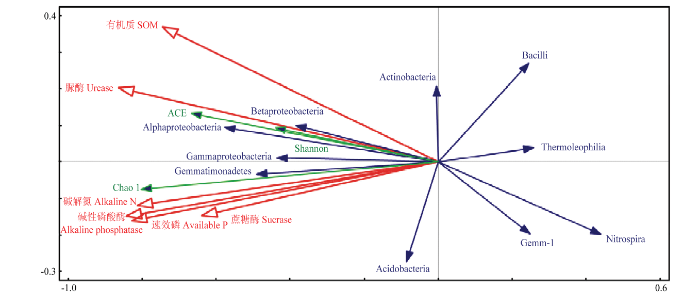 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4土壤养分、酶活性与细菌群落纲分类水平及多样性指数的冗余分析
Fig. 4The redundancy analysis (RDA) of soil nutrients, enzyme activities and soil bacterial community class level with diversity index
2.5 不同土壤样本细菌群落PICRUSt功能预测分析
2.5.1 功能基因家族组成 将预测得到的功能基因家族(图5)与KEGG数据库比对发现,第一等级上,所有菌群的基因序列注释到的功能可分为6类,其中前3类分别是:新陈代谢、环境信息加工和遗传信息加工。第二等级上,参与新陈代谢的菌群中,以参与氨基酸代谢、碳水化物代谢和能量代谢的为最大类群;参与环境信息加工的菌群中,参与膜运转类群具有明显优势,其次是参与信号转导的类群;参与遗传信息加工的菌群中,参与遗传信息复制和修复机制的类群具有明显优势。图5
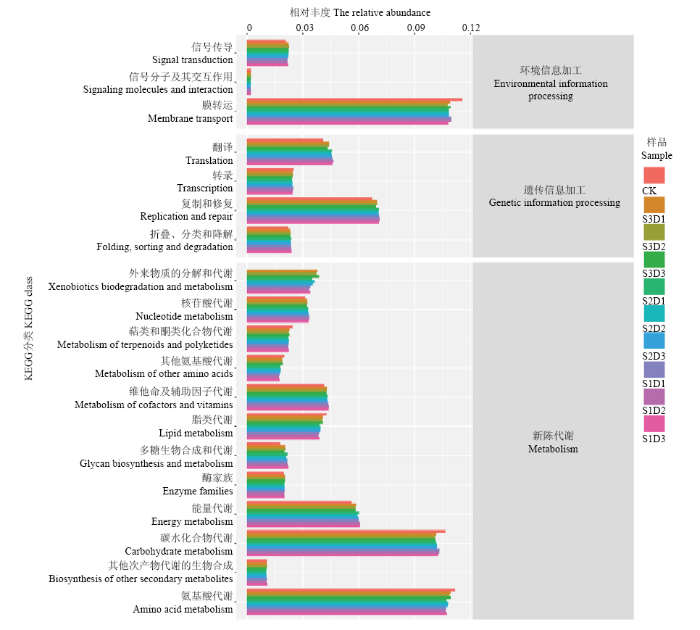 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同土壤样本细菌群落功能基因KEGG丰度图
Fig. 5The abundance chart of KEGG of functional genes for different soil samples bacterial community
2.5.2 代谢相关基因分析 图6为所测土壤样品KEGG直系同源基因簇(KEGG orthologous groups)丰度热图,横坐标表示所测土壤样本,纵坐标表示代谢相关基因所对应的酶。分析发现,氨基酸代谢、碳水化合物代谢和能量代谢通路上的相关基因表达差异较为显著;各土壤样品可大致聚为CK和饲料油菜还田(S2D3除外)两类,饲料油菜还田明显提高代谢相关基因丰度,其中S3D3在基因丰度上更具优势。对该处理下的优势基因所对应的酶进行筛选并归类,得到:与脂肪酸合成代谢相关的酰基载体蛋白还原酶(K00059),与能源物质代谢相关的重要中间代谢物乙酰CoA(K00626),具有特异催化蛋白质底物功能(丝氨酸、苏氨酸羟基磷酸化)的蛋白激酶(K08884),具有特异性识别天冬氨酸和谷氨酰胺并与相应tRNA形成氨酰tRNA的氨酰tRNA合成酶(K02433),作用于支链氨基酸(亮氨酸、缬氨酸和异亮氨酸)转运系统中的酶(K01995、K01998、K01999),参与脂肪酸β-氧化的烯酰CoA水合酶(K01692),参与脂肪酸合成与分解的活性代谢中间产物酰基CoA合成酶(K01897),以及参与肽/镍转运系统底物结合和渗透的酶蛋白(K02032、K02033、K02034、K02035)。
图6
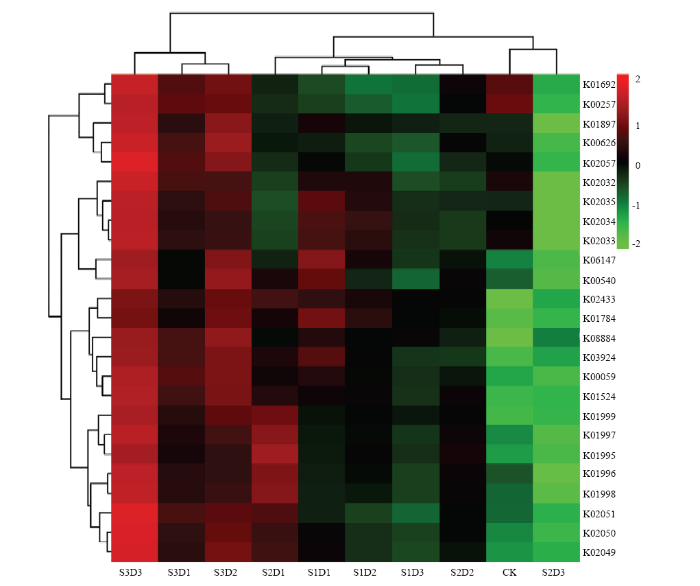 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同土壤样本细菌群落KEGG直系同源基因簇(KO)丰度热图
Fig. 6The abundance heatmap of KEGG orthologous groups (KO) cluster for different soil samples bacterial community
3 讨论
3.1 土壤养分及酶活性变化
油菜作绿肥翻压还田可增加后茬麦田耕层土壤酶活性[16,28],随绿肥还田量增大,土壤酶活性增加变缓[29]。本试验中,与常规农户模式相比,9个饲料油菜还田处理均增加了后茬麦田耕层土壤养分及酶活性,与前人研究结果一致,不同的是,随播量增加、还田时期推迟土壤养分及酶活性均呈逐渐增加趋势,以大播量晚期还田处理S3D3的土壤养分及酶活性最高。这可能与还田油菜生物量及腐解速率有关。本试验表明,随播量增加,还田生物量明显增加;随还田时期推后,还田生物量虽没有明显变化,但气温明显降低,可能会降低土壤微生物及土壤酶活性,进而导致还田油菜腐解速率变缓。有研究还指出,绿肥腐解速率表现为前期快、后期慢,腐解高峰均出现在还田后最初的1个月内[30]。本研究中,绿肥还田时期为9月10、20、30日,对应平均气温分别为15、11、7℃,因此,随气温降低,晚期还田油菜较前、中期还田油菜腐解速率变慢,有利于养分缓慢释放[31],进而促进了后茬麦田土壤养分及酶活性提高,尤其与碳代谢相关的土壤有机质和蔗糖酶活性显著提高。3.2 土壤细菌群落结构及多样性分析
LONGA等[20]研究指出,绿肥还田能增加土壤细菌Chao1、Shannon和Simpson指数。本研究结果表明,饲料油菜作绿肥时,其播量、还田时期及二者的互作效应均显著影响细菌群落多样性,以大播量晚期还田处理的多样性指数较高且高于CK,因此应合理规划播量及还田时期。本研究中,多样性指数在中低播量处理中出现低于CK的情况,可能的原因是该播量下还田油菜生物量低,对土壤微生物及土壤酶活性的促进作用低,向土壤释放养分低(表1)。而大播量晚期还田处理多样性指数较高,一方面缘于还田生物量高,有利于活化土壤微生物、酶活性,释放土壤养分;另一方面受低气温影响,还田油菜腐解速率变慢,其中未腐解油菜可能与后茬小麦构成了混作体系,进而增加了土壤细菌群落多样性及种群数量[31,32]。本研究进一步表明,饲料油菜还田改善了土壤细菌群落组成且有益细菌丰度显著增加。各样地优势菌门为变形菌门、放线菌门、酸杆菌门、厚壁菌门。前人研究表明,绿肥还田后,土壤中酸杆菌门、芽单胞菌门和硝化螺旋菌门相对丰度增加[33,34,35],这与本研究结果一致。变形菌门属需营养类群,其相对丰度在高氮及高有机质水平下通常增加[36,37]。本研究中,随播量增加变形菌门相对丰度逐渐增加,说明油菜还田为变形菌门提供了丰富的有机养分。酸杆菌门作为嗜酸菌具有降解植物残体多聚体、参与铁循环、增强光合作用及参与单碳化合物代谢等功能,其相对丰度也可反映土壤酸性条件[38,39]。本研究中,油菜还田显著增加了酸杆菌门的相对丰度,说明油菜还田可能降低了后茬麦田土壤pH。芽单胞菌门对土氮素循环中反硝化作用具有重要意义[40]。本研究中,芽单胞菌门相对丰度显著增加,这可能与油菜还田后土壤中具有合适的碳氮比有关。硝化螺旋菌门适合高pH环境条件生存[41],本研究中硝化螺旋菌门相对丰度随着绿肥还田量的增加而逐渐降低,可能也是由于绿肥还田导致土壤pH降低引起的,而这种改变有利于改良黄土高原碱性土壤环境。疣微菌门为寡营养性细菌,其分布广泛,兼性好氧,能够利用各种糖类,其相对丰度显著增加可能是由于油菜还田提供大量的营养物质[42,43]。另外,本研究发现油菜还田降低了放线菌门及具有致病性的厚壁菌门的相对丰度。这与林叶春等研究结果一致[34]。关于上述菌群在油菜还田后可能促进土壤碳氮循环及降低土壤pH值中发挥的作用,将在后续研究中进一步探讨。
RDA结果表明,土壤优势菌群中的α-变形菌纲、β-变形菌纲、γ-变形菌纲、酸杆菌纲和芽单孢菌纲与土壤养分、酶活性相关性较强,说明油菜还田后土壤养分的增加有益于改善土壤有益细菌群落的多样性[44]。而硝化螺旋菌纲、厚壁菌门中的芽孢杆菌纲、嗜热油菌纲则与之呈负相关,这也与上述门水平上细菌群落变化相吻合。
3.3 功能预测
LEBRUN[45]研究表明,麦田根际土壤中存在大量具有相对特定生态功能的微生物,并进行着诸多活跃的与新陈代谢相关的活动。本研究发现,油菜还田后,后茬麦田土壤中编码新陈代谢的相关基因具有明显优势,由此推论,土壤化学性质的改变导致相对特定生态功能微生物改变。KEGG丰度热图初步得出,S3D3处理的多种酶基因相对丰度具有明显优势。研究表明[46],酰基载体蛋白还原酶能从头合成脂肪酸,烯酰CoA水合酶催化脂肪酸氧化分解中β氧化循环,酰基CoA合成酶在脂肪酸合成和分解代谢中也扮演重要角色[47]。乙酰CoA是微生物碳代谢的关键分子,是体内能源物质代谢的枢纽性物质[48]。本研究上述酶基因丰度较CK明显增加,表明油菜还田能增加土壤中与碳代谢有关的细菌类群,增强了碳源利用能力。当土壤中氨含量较高时可形成谷氨酰胺或天冬酰胺,通过不同的氨基酸转移酶形成多种氨基酸[49]。氨基酸能促进土壤微生物活动,增加土壤微生物数量,有利于加速有机物矿化,促进营养元素释放[44]。其中,支链氨基酸是一类具有多种生理和代谢作用的必需氨基酸[50]。氨基酸合成需要的碳骨架来源于糖酵解、光合作用、氧化磷酸戊糖途径及三羧酸循环[51],因此,氨基酸的合成是碳代谢与氮代谢的枢纽。本研究显示,油菜还田增加了土壤中与碳代谢及氮代谢相关细菌,促进土壤碳、氮代谢。此外,镍元素能促进氮素转运,是脲酶和氢化酶活性表达所必需的金属辅基,缺乏会导致植物体内尿素的积累[52],钛元素对增加叶绿素含量、提高光合效率、提高植物体内多种酶的活性有积极作用[53]。本研究发现,S3D3下与钛、镍元素转运相关的酶基因丰度,有利于增加土壤中微量元素钛和有益元素镍的利用。
4 结论
相比常规农户麦后复种夏玉米的种植方式,黄土高原旱地小麦种植区夏闲期引入饲料油菜作绿肥能提高后茬麦田土壤肥力,有效改善土壤细菌群落结构及多样性,增强土壤细菌碳代谢及氮代谢能力,对旱地农业可持续发展及合理农作具有重要意义。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2013.15.010URLMagsci [本文引用: 1]

【目的】氮素是生命活动的必需元素,也是生态系统变化的主要因子。定量氮素的来源和贡献,并确定未来的发展方向是协调中国资源、环境与粮食安全的重要内容。【方法】通过食物链模型分析中国陆地生态系统中氮素来源的构成、特点和变化,以及氮素投入变化对农业生产、人体营养改善的贡献,并通过国内外综合比较提出了未来中国氮素管理的重点方向。【结果】中国已是世界上最大的氮肥生产和消费国,对近二十年全球氮肥产用量增长的贡献达61%和52%。氮肥已占中国陆地生态系统氮素输入量的72%,中国人蛋白质消费量中有56%来自于氮肥,氮肥等新技术的增产作用相当于将人均耕地面积从0.08 hm2提高到了0.52 hm2,扩大了6.5倍,这是中国以全球10%的土地资源、21%的灌溉面积养活20%的人口并不断提高生活水平的关键。但目前中国氮肥生产量超过了消费量、而消费量超过作物最高产量需求量,农业系统中的氮肥盈余量已经达到175 kg•hm-2,成为环境污染因子。【结论】中国需要尽快转变发展方式,从鼓励施肥转向控制施肥,以同时实现增产和环保;从满足化肥供应数量转向提高质量,加强节能和产品优化工作;从单一促进农业管理转向能源、农业和环境管理的同步。
DOI:10.3864/j.issn.0578-1752.2013.15.010URLMagsci [本文引用: 1]
【目的】氮素是生命活动的必需元素,也是生态系统变化的主要因子。定量氮素的来源和贡献,并确定未来的发展方向是协调中国资源、环境与粮食安全的重要内容。【方法】通过食物链模型分析中国陆地生态系统中氮素来源的构成、特点和变化,以及氮素投入变化对农业生产、人体营养改善的贡献,并通过国内外综合比较提出了未来中国氮素管理的重点方向。【结果】中国已是世界上最大的氮肥生产和消费国,对近二十年全球氮肥产用量增长的贡献达61%和52%。氮肥已占中国陆地生态系统氮素输入量的72%,中国人蛋白质消费量中有56%来自于氮肥,氮肥等新技术的增产作用相当于将人均耕地面积从0.08 hm2提高到了0.52 hm2,扩大了6.5倍,这是中国以全球10%的土地资源、21%的灌溉面积养活20%的人口并不断提高生活水平的关键。但目前中国氮肥生产量超过了消费量、而消费量超过作物最高产量需求量,农业系统中的氮肥盈余量已经达到175 kg•hm-2,成为环境污染因子。【结论】中国需要尽快转变发展方式,从鼓励施肥转向控制施肥,以同时实现增产和环保;从满足化肥供应数量转向提高质量,加强节能和产品优化工作;从单一促进农业管理转向能源、农业和环境管理的同步。
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
DOI:10.3864/j.issn.0578-1752.2014.10.011URLMagsci [本文引用: 1]
【目的】综合评价南方稻区不同冬季绿肥种类种植下水稻生长动态及稻田杂草群落效应,进一步优化稻田种植模式,大力发展冬季作物种植与开发。【方法】从2011年冬开始,在南方双季稻区江西农业大学科技园,采用田间试验,分别选取豆科、禾本科、十字花科冬季绿肥种类,比较冬季绿肥紫云英、油菜、黑麦草及混播绿肥(紫云英×油菜×肥田萝卜)对后作早稻产量及产量构成、干物质生产特性、稻田杂草群落的差异变化。【结果】紫云英前茬处理的早稻(紫云英—早稻—晚稻,MV-R-R)分蘖盛期、孕穗期、抽穗期和成熟期的群体干物重均明显高于其他处理,分别高出14.46%、10.99%、11.83%和7.23%,相应地其产量也分别比黑麦草—早稻—晚稻(RG-R-R)、油菜—早稻—晚稻(RP-R-R)、混播绿肥—早稻—晚稻(MS-R-R)处理高6.61%、3.29%及0.78%。其中产量构成中有效穗数、每穗粒数、结实率和千粒重也均以MV-R-R处理最高。水稻主要生育阶段的干物质积累,除抽穗期至成熟期MV-R-R低于RG-R-R处理外,在播种至分蘖盛期、分蘖盛期至孕穗期、孕穗期至抽穗期均是MV-R-R处理最高;与其他处理相比,增加幅度分别为11.38%—17.72%、4.50%—28.00%、1.85%—35.25%。水稻播种期至分蘖盛期和孕穗期至抽穗期两个生育阶段干物质积累量最大,是物质快速积累的阶段,各处理这2个生育阶段干物质积累量分别达到成熟期干物重的70.43%、60.50%、69.87%、69.08%;而抽穗期至成熟期各处理干物质积累所占比例均最低。干物质在茎鞘、叶片和穗分配情况,茎鞘的分配比例在孕穗期最大,并随着生育进程不断降低,在成熟期达到最低,各处理从58.30%—64.20%降低至27.95%—30.47%;叶片的分配比例也以孕穗期最大,并从该期的35.80%—41.70%降低至成熟期的13.07%—14.62%。另外,据调查,稻田杂草有7科11种,其中稗草(Echinochloa crusgalli)、鸭舌草(Monochoia vaginalis)、矮慈姑(Sagittaria pygmaea)和牛毛毡(Eleocharis yokoscensis)密度最大,群体数量为田间杂草的80.84%。MV-R-R处理对稻田杂草密度有显著影响,对杂草的抑制效果明显优于其他处理,RG-R-R处理的效果最差。通过分析杂草在群落中的相对优势度可以看出,MV-R-R和RG-R-R处理中杂草群落组成主要为稗草+鸭舌草+牛毛毡,而RP-R-R和MS-R-R处理为稗草+鸭舌草+矮慈姑。RG-R-R处理杂草物种丰富度最大,显著高于其他处理。从表征多样性的Shannon指数来看,RG-R-R冬种黑麦草处理的杂草物种多样性指数最高,而MV-R-R处理最低。从表征优势度的Simpson指数来看,MV-R-R处理的杂草优势度最高,RG-R-R处理最低。从Pielou均匀度指数来看,MV-R-R处理下的均匀度指数高于其他处理,但差异不显著,而其他冬季绿肥处理的稻田杂草均匀度有所下降,其中RG-R-R处理最低。【结论】紫云英—早稻—晚稻处理能保证较高的干物质积累量,具有较高的增产潜力,对杂草发生种类和密度有显著影响,并且有利于提高杂草均匀度,弱化稻田优势种杂草在田间的危害性。
DOI:10.3864/j.issn.0578-1752.2014.10.011URLMagsci [本文引用: 1]
【目的】综合评价南方稻区不同冬季绿肥种类种植下水稻生长动态及稻田杂草群落效应,进一步优化稻田种植模式,大力发展冬季作物种植与开发。【方法】从2011年冬开始,在南方双季稻区江西农业大学科技园,采用田间试验,分别选取豆科、禾本科、十字花科冬季绿肥种类,比较冬季绿肥紫云英、油菜、黑麦草及混播绿肥(紫云英×油菜×肥田萝卜)对后作早稻产量及产量构成、干物质生产特性、稻田杂草群落的差异变化。【结果】紫云英前茬处理的早稻(紫云英—早稻—晚稻,MV-R-R)分蘖盛期、孕穗期、抽穗期和成熟期的群体干物重均明显高于其他处理,分别高出14.46%、10.99%、11.83%和7.23%,相应地其产量也分别比黑麦草—早稻—晚稻(RG-R-R)、油菜—早稻—晚稻(RP-R-R)、混播绿肥—早稻—晚稻(MS-R-R)处理高6.61%、3.29%及0.78%。其中产量构成中有效穗数、每穗粒数、结实率和千粒重也均以MV-R-R处理最高。水稻主要生育阶段的干物质积累,除抽穗期至成熟期MV-R-R低于RG-R-R处理外,在播种至分蘖盛期、分蘖盛期至孕穗期、孕穗期至抽穗期均是MV-R-R处理最高;与其他处理相比,增加幅度分别为11.38%—17.72%、4.50%—28.00%、1.85%—35.25%。水稻播种期至分蘖盛期和孕穗期至抽穗期两个生育阶段干物质积累量最大,是物质快速积累的阶段,各处理这2个生育阶段干物质积累量分别达到成熟期干物重的70.43%、60.50%、69.87%、69.08%;而抽穗期至成熟期各处理干物质积累所占比例均最低。干物质在茎鞘、叶片和穗分配情况,茎鞘的分配比例在孕穗期最大,并随着生育进程不断降低,在成熟期达到最低,各处理从58.30%—64.20%降低至27.95%—30.47%;叶片的分配比例也以孕穗期最大,并从该期的35.80%—41.70%降低至成熟期的13.07%—14.62%。另外,据调查,稻田杂草有7科11种,其中稗草(Echinochloa crusgalli)、鸭舌草(Monochoia vaginalis)、矮慈姑(Sagittaria pygmaea)和牛毛毡(Eleocharis yokoscensis)密度最大,群体数量为田间杂草的80.84%。MV-R-R处理对稻田杂草密度有显著影响,对杂草的抑制效果明显优于其他处理,RG-R-R处理的效果最差。通过分析杂草在群落中的相对优势度可以看出,MV-R-R和RG-R-R处理中杂草群落组成主要为稗草+鸭舌草+牛毛毡,而RP-R-R和MS-R-R处理为稗草+鸭舌草+矮慈姑。RG-R-R处理杂草物种丰富度最大,显著高于其他处理。从表征多样性的Shannon指数来看,RG-R-R冬种黑麦草处理的杂草物种多样性指数最高,而MV-R-R处理最低。从表征优势度的Simpson指数来看,MV-R-R处理的杂草优势度最高,RG-R-R处理最低。从Pielou均匀度指数来看,MV-R-R处理下的均匀度指数高于其他处理,但差异不显著,而其他冬季绿肥处理的稻田杂草均匀度有所下降,其中RG-R-R处理最低。【结论】紫云英—早稻—晚稻处理能保证较高的干物质积累量,具有较高的增产潜力,对杂草发生种类和密度有显著影响,并且有利于提高杂草均匀度,弱化稻田优势种杂草在田间的危害性。
URLMagsci [本文引用: 1]
为探讨冬季种植绿肥作物改良土壤的生态作用, 以冬闲连作为对照, 通过连续8年的定位试验研究了冬种绿肥(紫云英、油菜和黑麦草)收获后翻压还田对后作水稻产量及土壤肥力的影响。结果表明: 连续8年冬种绿肥期间, 冬种绿肥各处理的水稻平均产量均高于冬闲对照, 但不同处理增产效果有所不同, 其中紫云英 早稻 晚稻处理增产效果最明显, 为10.48%, 其次是油菜 早稻 晚稻处理(7.31%), 黑麦草 早稻 晚稻处理的增产效果较差, 仅为5.76%。紫云英 早稻 晚稻处理的土壤pH和碱解氮含量最高, 与其他处理相比, pH增加0.58%~1.37%, 碱解氮增加6.59%~35.35%。黑麦草 早稻 晚稻处理的土壤有机质、有效磷、速效钾含量和C/N在各处理中最高, 分别较其他处理高2.99%~4.98%、19.25%~31.05%、16.97%~22.15%和0.97%~6.61%。相关性分析结果表明, 水稻产量与每穗颖花数、结实率、千粒重及土壤碱解氮、有效磷极显著相关(<i>P</i><0.01), 与每穗粒数、有机质、速效钾显著相关(<i>P</i><0.05), 而与穗长、有效穗数及土壤pH和C/N相关不显著; 水稻产量各构成要素与土壤养分之间的关系极为密切。综合来看, 稻田冬种绿肥有利于增加水稻产量和改善土壤肥力。
URLMagsci [本文引用: 1]
为探讨冬季种植绿肥作物改良土壤的生态作用, 以冬闲连作为对照, 通过连续8年的定位试验研究了冬种绿肥(紫云英、油菜和黑麦草)收获后翻压还田对后作水稻产量及土壤肥力的影响。结果表明: 连续8年冬种绿肥期间, 冬种绿肥各处理的水稻平均产量均高于冬闲对照, 但不同处理增产效果有所不同, 其中紫云英 早稻 晚稻处理增产效果最明显, 为10.48%, 其次是油菜 早稻 晚稻处理(7.31%), 黑麦草 早稻 晚稻处理的增产效果较差, 仅为5.76%。紫云英 早稻 晚稻处理的土壤pH和碱解氮含量最高, 与其他处理相比, pH增加0.58%~1.37%, 碱解氮增加6.59%~35.35%。黑麦草 早稻 晚稻处理的土壤有机质、有效磷、速效钾含量和C/N在各处理中最高, 分别较其他处理高2.99%~4.98%、19.25%~31.05%、16.97%~22.15%和0.97%~6.61%。相关性分析结果表明, 水稻产量与每穗颖花数、结实率、千粒重及土壤碱解氮、有效磷极显著相关(<i>P</i><0.01), 与每穗粒数、有机质、速效钾显著相关(<i>P</i><0.05), 而与穗长、有效穗数及土壤pH和C/N相关不显著; 水稻产量各构成要素与土壤养分之间的关系极为密切。综合来看, 稻田冬种绿肥有利于增加水稻产量和改善土壤肥力。
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLMagsci [本文引用: 1]
<div >以冬小麦收获后高留茬休闲地为对照,连续4年在陇东黄土旱塬设6个油菜播种期,研究了休闲期作物覆盖对土壤水分及后作冬小麦产量与水分利用效率的影响.结果表明:不同播种期夏休闲期土壤贮水量差异显著(<em>P</em><0.05),其中8月5日播种油菜的土壤蓄水效率为58.5%,产量和水分利用效率较对照提高7.5%和5.9%.平均而言,夏休闲期复种油菜后作小麦干旱年份增产16.1%,平水年份增产6.8%.夏休闲期复种油菜是西北旱地小麦抗旱增产的有益途径.</div><div > </div>
URLMagsci [本文引用: 1]
<div >以冬小麦收获后高留茬休闲地为对照,连续4年在陇东黄土旱塬设6个油菜播种期,研究了休闲期作物覆盖对土壤水分及后作冬小麦产量与水分利用效率的影响.结果表明:不同播种期夏休闲期土壤贮水量差异显著(<em>P</em><0.05),其中8月5日播种油菜的土壤蓄水效率为58.5%,产量和水分利用效率较对照提高7.5%和5.9%.平均而言,夏休闲期复种油菜后作小麦干旱年份增产16.1%,平水年份增产6.8%.夏休闲期复种油菜是西北旱地小麦抗旱增产的有益途径.</div><div > </div>
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
DOI:10.11674/zwyf.2013.0222URLMagsci [本文引用: 1]
<p> 通过田间试验,研究翻压绿肥对植烟土壤酶活性及土壤肥力的影响。结果表明, 翻压绿肥能够明显提高土壤酶活性和土壤肥力水平,当绿肥翻压量在15000 kg/hm<sup>2</sup>以上时,尤其在22500~30000 kg/hm<sup>2</sup>之间时对土壤各项指标的影响更加明显。与对照相比,翻压绿肥的各处理土壤脲酶、 酸性磷酸酶、 蔗糖酶、 过氧化氢酶增幅分别为13.10%2~3.81%、 12.92%~29.38%、 75.35%~234.51%、 29.17%~37.08%; 土壤有机质、 全氮、 碱解氮、 有效磷、 速效钾、 pH、 孔隙度增幅分别为13.01%~70.41%、 6.42%~27.52%、 1.14%~10.99%、 15.97%~34.99%、 10.28%~38.30%、 2.74%~7.05%、 0.19%~2.50%,土壤容重降幅为1.47%~5.15%。简单相关分析表明,脲酶、 酸性磷酸酶、 蔗糖酶、 过氧化氢酶4种酶之间以及4种酶与土壤理化因子之间均有极显著的相关关系,而土壤酶活性之间的相互关系表明,土壤酶在促进土壤有机物质转化中不仅显示其专性特性,同时也存在共性关系; 典型相关分析结果为,第一对典型变量线性函数反映了土壤酶活性和土壤养分因子对土壤综合肥力水平的影响,第二对典型变量线性函数反映了施入绿肥对土壤内部重要的生理生化过程变化的影响; 主成分分析结果显示,第一主成分反映了土壤的综合肥力水平,所有因子均对土壤肥力水平起到了正效应,土壤酶活性能够和土壤理化因子共同评价土壤综合肥力水平。以上结果说明,翻压绿肥后土壤生物过程活跃,有利于土壤有机物质的转化和烤烟正常生长所需的营养供应。</p>
DOI:10.11674/zwyf.2013.0222URLMagsci [本文引用: 1]
<p> 通过田间试验,研究翻压绿肥对植烟土壤酶活性及土壤肥力的影响。结果表明, 翻压绿肥能够明显提高土壤酶活性和土壤肥力水平,当绿肥翻压量在15000 kg/hm<sup>2</sup>以上时,尤其在22500~30000 kg/hm<sup>2</sup>之间时对土壤各项指标的影响更加明显。与对照相比,翻压绿肥的各处理土壤脲酶、 酸性磷酸酶、 蔗糖酶、 过氧化氢酶增幅分别为13.10%2~3.81%、 12.92%~29.38%、 75.35%~234.51%、 29.17%~37.08%; 土壤有机质、 全氮、 碱解氮、 有效磷、 速效钾、 pH、 孔隙度增幅分别为13.01%~70.41%、 6.42%~27.52%、 1.14%~10.99%、 15.97%~34.99%、 10.28%~38.30%、 2.74%~7.05%、 0.19%~2.50%,土壤容重降幅为1.47%~5.15%。简单相关分析表明,脲酶、 酸性磷酸酶、 蔗糖酶、 过氧化氢酶4种酶之间以及4种酶与土壤理化因子之间均有极显著的相关关系,而土壤酶活性之间的相互关系表明,土壤酶在促进土壤有机物质转化中不仅显示其专性特性,同时也存在共性关系; 典型相关分析结果为,第一对典型变量线性函数反映了土壤酶活性和土壤养分因子对土壤综合肥力水平的影响,第二对典型变量线性函数反映了施入绿肥对土壤内部重要的生理生化过程变化的影响; 主成分分析结果显示,第一主成分反映了土壤的综合肥力水平,所有因子均对土壤肥力水平起到了正效应,土壤酶活性能够和土壤理化因子共同评价土壤综合肥力水平。以上结果说明,翻压绿肥后土壤生物过程活跃,有利于土壤有机物质的转化和烤烟正常生长所需的营养供应。</p>
[本文引用: 1]
URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[D].
[本文引用: 1]
[D].
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLMagsci [本文引用: 1]
利用Illumina平台Miseq高通量测序技术对小麦分蘖期砂姜黑土耕层土壤细菌进行高通量测序, 结合相关生物信息学分析, 探讨了不施化肥秸秆不还田(CK)、施化肥秸秆不还田(F)以及不施化肥秸秆还田(W)3种处理土壤细菌群落组成、多样性和结构的变化。结果显示, 测序共获得14 873个OTUs, 计173 323条读数, 平均读长439 bp。砂姜黑土细菌优势门(相对丰度>10%)为变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、放线菌门(Actinobacteria)和拟杆菌门(Bacteroidetes); 优势纲(相对丰度>10%)为α-变形菌纲(Alphaproteobacteria)、β-变形菌纲(Betaproteobacteria)、酸杆菌纲(Acidobacteria)、鞘脂杆菌纲 (Sphingobacteriia)和γ-变形菌纲(Gammaproteobacteria); 优势属(相对丰度>1%)共47个, 3个处理中均有分布的优势属21个, F处理的细菌优势属的种类最多, 为39个。相对丰度最大的门、纲和属分别是变形菌门(38.7%~43.1%)、α-变形菌纲(14.5%~18.1%)和鞘氨醇单胞菌属(<i>Sphingomonas</i>) (4.6%~7.7%)。F处理细菌丰富度指数(Chao1指数和ACE指数)显著低于CK及W处理, W处理和CK处理土壤细菌丰富度指数无显著差异, 与CK处理相比, F处理ACE指数降低22.8%。W处理土壤细菌Shannon多样性指数显著大于CK及F处理, W处理Shannon指数较CK处理提高4.1%, 而F处理土壤细菌Shannon指数与CK处理无显著差异。F处理Simpson指数显著高于CK及W处理; F处理Simpson指数较CK处理提高38.1%, 而W处理细菌Simpson指数最小, 显著低于CK处理, 较CK降低23.8%。分层聚类图显示在属的水平上, W处理和CK处理土壤细菌群落结构相似性较高, F处理与CK处理及W处理细菌群落结构差异较大。施化肥对土壤细菌优势类群组成、相对丰度及群落结构的影响大于秸秆还田, 施化肥显著降低了土壤细菌丰富度, 秸秆还田显著提高了土壤细菌的多样性。
URLMagsci [本文引用: 1]
利用Illumina平台Miseq高通量测序技术对小麦分蘖期砂姜黑土耕层土壤细菌进行高通量测序, 结合相关生物信息学分析, 探讨了不施化肥秸秆不还田(CK)、施化肥秸秆不还田(F)以及不施化肥秸秆还田(W)3种处理土壤细菌群落组成、多样性和结构的变化。结果显示, 测序共获得14 873个OTUs, 计173 323条读数, 平均读长439 bp。砂姜黑土细菌优势门(相对丰度>10%)为变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、放线菌门(Actinobacteria)和拟杆菌门(Bacteroidetes); 优势纲(相对丰度>10%)为α-变形菌纲(Alphaproteobacteria)、β-变形菌纲(Betaproteobacteria)、酸杆菌纲(Acidobacteria)、鞘脂杆菌纲 (Sphingobacteriia)和γ-变形菌纲(Gammaproteobacteria); 优势属(相对丰度>1%)共47个, 3个处理中均有分布的优势属21个, F处理的细菌优势属的种类最多, 为39个。相对丰度最大的门、纲和属分别是变形菌门(38.7%~43.1%)、α-变形菌纲(14.5%~18.1%)和鞘氨醇单胞菌属(<i>Sphingomonas</i>) (4.6%~7.7%)。F处理细菌丰富度指数(Chao1指数和ACE指数)显著低于CK及W处理, W处理和CK处理土壤细菌丰富度指数无显著差异, 与CK处理相比, F处理ACE指数降低22.8%。W处理土壤细菌Shannon多样性指数显著大于CK及F处理, W处理Shannon指数较CK处理提高4.1%, 而F处理土壤细菌Shannon指数与CK处理无显著差异。F处理Simpson指数显著高于CK及W处理; F处理Simpson指数较CK处理提高38.1%, 而W处理细菌Simpson指数最小, 显著低于CK处理, 较CK降低23.8%。分层聚类图显示在属的水平上, W处理和CK处理土壤细菌群落结构相似性较高, F处理与CK处理及W处理细菌群落结构差异较大。施化肥对土壤细菌优势类群组成、相对丰度及群落结构的影响大于秸秆还田, 施化肥显著降低了土壤细菌丰富度, 秸秆还田显著提高了土壤细菌的多样性。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URLMagsci [本文引用: 1]
<p>生物体β-氧化循环是脂肪酸氧化分解的主要途径,许多代谢疾病都与其密切相关。线虫β-氧化循环与人类相似,但线虫β-氧化循环的研究报道却很少。线虫基因C32E8.9编码的蛋白被WormBase命名为烯脂酰CoA水合酶(WormBase ID:CE29693),被推测具有催化β-氧化循环第二步反应的功能。作者将C32E8.9基因克隆到原核表达载体上,在大肠杆菌中获得高效表达,并分别纯化了母体蛋白以及硒代蛋白衍生物。多角度静态光散射实验表明该蛋白的聚合状态为三聚体。该蛋白在沉淀剂2-甲基-2,4-戊二醇的作用下形成可供衍射分析的六棱柱形状晶体,空间群为P21,晶胞参数为a=79.0?,b=82.4?,c=79.2?,α=γ=90.0°,β=120°,数据分析表明该晶体非单晶,是一种罕见的蛋白质“三晶”——包含三套晶格。</p>
URLMagsci [本文引用: 1]
<p>生物体β-氧化循环是脂肪酸氧化分解的主要途径,许多代谢疾病都与其密切相关。线虫β-氧化循环与人类相似,但线虫β-氧化循环的研究报道却很少。线虫基因C32E8.9编码的蛋白被WormBase命名为烯脂酰CoA水合酶(WormBase ID:CE29693),被推测具有催化β-氧化循环第二步反应的功能。作者将C32E8.9基因克隆到原核表达载体上,在大肠杆菌中获得高效表达,并分别纯化了母体蛋白以及硒代蛋白衍生物。多角度静态光散射实验表明该蛋白的聚合状态为三聚体。该蛋白在沉淀剂2-甲基-2,4-戊二醇的作用下形成可供衍射分析的六棱柱形状晶体,空间群为P21,晶胞参数为a=79.0?,b=82.4?,c=79.2?,α=γ=90.0°,β=120°,数据分析表明该晶体非单晶,是一种罕见的蛋白质“三晶”——包含三套晶格。</p>
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}