Proposition, Development and Application of the Integrated Microbiome Agent (IMA)
LIU Bo1, CHEN QianQian1, WANG JiePing1, RUAN ChuanQing1, CHEN YanPing1, XIA JiangPing2, CHE JianMei1, CHEN Zheng1, PAN ZhiZhen1, WEN Xiao2, ZHU YuJing1, ZHANG HaiFeng1, ZHENG XueFang1���α༭: ��÷
�ո�����:2019-03-19��������:2019-05-9�����������:2019-07-16
| ��������: |
Received:2019-03-19Accepted:2019-05-9Online:2019-07-16
����� About authors
������ͨ�����ߣ�,E-mail: fzliubo@163.com

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (2603KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
����, ��ٻٻ, ����ƽ, ���, ����Ƽ, �Ľ�ƽ, ������, ���, ��־��, ��Ц, ����ݼ, �ź���, ֣ѩ��. ���������������������з���Ӧ��[J]. �й�ũҵ��ѧ, 2019, 52(14): 2450-2467 doi:10.3864/j.issn.0578-1752.2019.14.006
LIU Bo, CHEN QianQian, WANG JiePing, RUAN ChuanQing, CHEN YanPing, XIA JiangPing, CHE JianMei, CHEN Zheng, PAN ZhiZhen, WEN Xiao, ZHU YuJing, ZHANG HaiFeng, ZHENG XueFang.
0 ����
���о����塿�����ִ�ũҵ��ҵ�ṹ����,�����רҵ��������ģ�����������������,��������Դ����,ʹ����������ָ����������,�������ص����������ϰ���continuous cropping obstacle���ѳ�Ϊ����ũҵ�����ɳ�����չ��ƿ��֮һ�������ϰ���������㡢�漰�����������,�����¼�������,�������,������صľ�����ʧ��������****�����������ϰ����о��϶�,������[1]������[2,3]���˲�[4]����[5]���ػ�[6,7,8]���ƹ�[9]�����[10]������[11]��������[12]������[13]����[14]������[15]�ȡ�����о��ձ���Ϊ,��������������ϰ�����Ҫ���ذ���:��1����������������ṹʧ��,��ԭ��������,�����������[5];��2������������״�Ķ��,�����ữ�����ջ��ͷ����½�,Ϊ��ԭ����̳��ṩ������������,�ɼ������������ķ���[2,3];��3��ֲ���ϵ�Զ����������ϵ�������쳣,�����˸�ϵ������Բ���������,��������������ع̵�����,����ֲ��Ӫ��ȱ��[9,16]��Ŀǰ��������ϰ���ʩ��Ҫ�������ͼ���[2]������Ʒ��ɸѡ[10]����������[17]����ʩ�л�������������ʵʩ������ε�[5]������ֳ�������ũҵ������Ϊԭ��,�������������������integrated microbiome agent,IMA��,���������������,�������ϰ�������غ����ݷ����Դ�����þ�����Ҫ���塣��ǰ���о���չ��������������ũ�²�������������Ⱥ��仯��ϵ���С��˲Ρ������������ʹ����������е���������Ⱥ��仯���н�Ϊ������о�[4-5,12,14]�����������������ƽ����ƻ������������ϰ�����Ҫ���ء����������飨integrated microbiome��Ӱ�����������ܡ������������������ú������ȷ�����������������ļ���,���н϶��о�����,��YANG��[18]������������������Ⱥ��仯���̲���ݲ��Ĺ�ϵ;NOVELLO��[19]�����������������������������Ѳ����ۺ������Ĺ�ϵ;KALIVAS��[20]���ú�������ʾ���������������������������������ʵĻ��ơ�����������������������������,���������������������ĸ���,���ú�������ܼ�������������ļ��ϳ�֮Ϊ����������,��ͼͨ������������ĸ�Ԥ,�ı�ֲ���ϵ�����е�����ṹ,���������ڻ����еĹ���,��Ԫ�ؽ��⡢�������ơ����طֽ��,�����������������ϰ�,����о�δ�������������о�����㡿Χ�����������������������Ͼ�������Ʒ��������������ָ�ꡢ���Ͼ������ܱȽϡ����Ͼ����������������Ӱ�졢���Ͼ����������ϰ��������õȿ�չ�о�,������Ϊһ�������Ƽ�������������������������Ӧ�õĿ����ԡ��������Ĺؼ����⡿���������������������������,�о�������������ṹ,�ⶨ������������������ѿ�ʡ������ʡ��������ص�����,Ϊ�����ϰ�������ء����ݷ����Դ�����á�����������з���Ӧ���ṩ��ѧ���ݡ�1 �����뷽��
������2018��2019���ڸ���ʡũҵ��ѧԺ��ɡ�1.1 �����������������������
��������������������������ʹ���������ϵͳ���С��ص�Ϊ����ʡũҵ��ѧԺ�����ִ���ʩũҵ��������ʾ������,����ʹ����������Ὠ�����Ϊ2 100 m2����60 m,��35 m��,��ֳ���Ϊ1 600 m2,���ʹ��������80 cm,���ʹ�������33%Ҭ����33%�⿷��34%�ȿ���ɡ����ʹ�����1 600ͷ������,�������й�ڵ�����,���Ϲ���ÿ������1��,���Ϸ����³�ʱ�����µ��ϵ�ԭ���߶�;����ʹ��1���������ʹ���Ϊ����,�����������������ʱ,����30%������+70%�ӱ���������ϳ�����ԭ��,ƽ���ڷ��ʹ���10 cm,���ʹ����ʪ�ȿ�����50%��60%,���ʹ������¶�Լ24��26��,�ڲ��¶�40.5��45.5�档ÿ�����ű���Ϊ��������,���ж��ι����������,ÿ��������������1��,�������20 cm,��ֻ�Ϸ��ʹ�ԭ�е��ϡ����ӵ��ϡ�����,���ʪ�ȵ�����55%��65%,������������20 d;���ͽ��������Ʒ�ӹ�,������20 cm���ͺõĵ����ռ���Ϊ�������������ԭ��,������ɹ����5 d,�����Ͳ��ﺬˮ��<30%ʱ,�������ɡ����顢��ɸ�ȼӹ�,���顢��װ��Ϊ�ߺ����������������������Ʒ,��װ���Ϊ10 kg/������������������������հ�����ԭ�����á����ʹ����͡����Ϳ��ơ���Ʒ�ӹ�����Ʒ��װ�ȹ��̣�ͼ1����ͼ1�����������Ͼ�������װ�������͵��غ�Ʒ�ܼ��������ı�����ͼ1
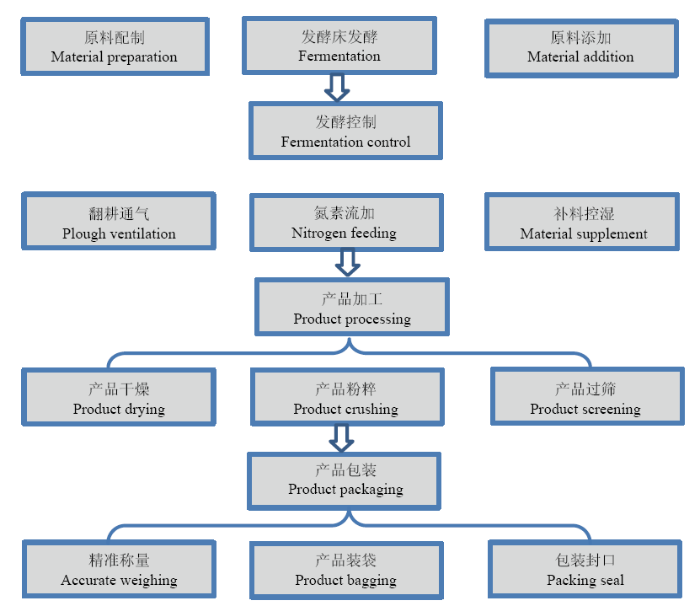 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ1�����������������������
Fig. 1Production process of the integrated microbiome agent
1.2 �������������Ӫ���ɷַ���
�Ӳ�Ʒ��ȡ��7��,��Ϊg1,g2,�� g7,�ͼ�����ʵ�λ����ʡũҵ��ѧԺ���������о�������,�ⶨ�������ʺ�Ӫ���ɷ�,����ˮ�֣�%����pH���л��ʣ�%����ȫ����%������ֳ�ᣨ%��������ά��%����,ͳ��ƽ��ֵ��1.3 �������������ѿ�߸˾���Bacillus spp.������ܺ����IJⶨ
�Բɼ���7����Ʒ����ѿ�߸˾��������,����MS������,����10 g��Ʒ,���ƺ�10%��Ʒˮ����Һ,������80��ά��20 min,����ѿ�߸˾�,���������Ӿ�,ͨ���ݶ�ϡ��Ϳ�����������,��Ǿ�����̬����,���о�������ͳ�Ƽ���;����ͨ��ϸ��16S rRNA����������������ȶԡ�����,ȷ����Ǿ���Ϊѿ�߸˾���,ͳ�Ƽ���ÿ�˲�Ʒ��ѿ�߸˾���������cfu/g��,ÿ����Ʒ�����ظ�3��,����ƽ��ֵ��1.4 ���ں����������Ͼ�����Ʒϸ��������ⶨ
�����������������Ʒ��ȡ��7����Ʒ,ÿ����ȡ100 g��Ͼ��Ⱥ��700 g��Ʒ,����ȡ50 g���ú���������ϸ�����������:��1����DNA����ȡ:������DNA��ȡ�Լ���FastDNA SPIN Kit for Soil�IJ���ָ��,�ֱ���ȡ��������������DNA,��-80����䶳�汸��;��2��16S rDNA�����Ŀ�Ĺ���:��������ԭ������16S rDNA��V3-V4����ͨ������U341F��U785R�Ը�������������DNA����PCR����,�������ϲ����ͷ,�Ӷ�������������������ϸ��16S rDNA V3-V4�������Ŀ�;��3����ͨ������:ʹ��Illumina MiSeq����ƽ̨,����PE300�������,ÿ���������ٻ��10����reads;ϸ��������ⶨ���Ϻ�������˾��ɡ�����������֪���ֵ�16S���ݿ⣨Silva,http://www.arb-silva.de���ȶ�,��������ע��,�ó�������Ϣ����õ���Ʒϸ�����������ݺ����:��1��ϸ����ˮƽ��������������reads��;��2��ϸ��������������Ԫ���ࣨOTU������;��3��ǰ10�ָߺ���ϸ������ⶨ;��4��ϸ����������ѿ�߸˾������ȣ�%����1.5 ������������������ӷ�ѿ��Ӱ��
��0.1 kg�����������������Ʒ������0.4 Lˮ��,�����ҹ��,��4��ɴ������,�ռ���Һ,��Ϊ25%�������������������Һ��ȡ������Һ20 mL,����ֱ��9 cm��������,������ײ�����һ����ֽ;ѡ���������溦���̶�����,�ý���Һ����30 min,ÿ���з���12���̶�����,�ظ�5��,����ˮ20 mL��Ϊ���գ�CK��,��������28������˹�������,����16 h:�ڰ�8 h��ÿ24 h��1 d���۲��̶��ķ�ѿ�������߸�����,ͳ�Ʒ�ѿ�ʡ���ѿָ���ͻ���ָ��;�Զ�����Ϊ����,���㴦������������߸����Ⱥͻ���ָ���ı�ֵ,�����������������25%����Һ���̶���ѿ��Ӱ�졣���㹫ʽ:��ѿ�ʣ�G,%��=����ѿ������/��������������100����ѿָ����GI��=����Gt/Dt��������,GtΪ��t��ķ�ѿ������,DtΪ��Ӧ��ѿ����������ָ����VI��=GI��S������,GIΪ��ѿָ��,SΪ��ѿtʱ�����߸���������ܳ��ȣ�cm����1.6 ��������������Է���Ѩ������׳�����õ�Ӱ��
�����������,ѡ�����������������Ʒ��Ҭ�����Ƴɲ�ͬ����������������,�������øߺ����鴦��1�ʹ���2,�������������ռ�ȷֱ�Ϊ30%��25%;�к����鴦��3�ʹ���4,ռ�ȷֱ�Ϊ20%��15%;�ͺ����鴦��5�ʹ���6,ռ�ȷֱ�Ϊ10%��5%;����������ƻ�Ϻ�,ƽ����Ѩ�̽��з�������,����Ʒ��Ϊ������ݲ��ġ�ũ��180��,ÿ����������25������,�ظ�2��,��Ҭ����������ʶ��գ�CK��;����30������,���չ��Ϊ��Դ,����16 h:�ڰ�8 h,ÿ�ն�����ˮ����;��10��20��30 d���ڹ۲췬�ѵij�����������,ͳ��30 d�ķ��ѳ����ʣ�%����������cm������ߣ�cm�������֣�cm����,�ȽϷ�����ͬ������������������Է���Ѩ������׳�����õ�Ӱ�졣1.7 ��������������Է���Ѩ��������ݲ����ص�Ӱ��
�����ϲ�ͬ�������Ѩ������30 d��ķ���Ѩ���磨��ũ��180��������������ݲ���������;ÿ�������������Ũ��Ϊ106 cfu/mLǿ�²���������Ͼ���Ralstonia solanacearum��FJAT-91����Һ200 mL,�����齽����ˮΪ����;���ֺ�ֱ��ڵ�4��7��10��۲���㷬������ݲ�������,ͳ����������������ߺ����飨25%��30%�����к����飨15%��20%�����ͺ����飨5%��10%���������飨��ˮ����ƽ��������,����10 dУ����Ч,���з����Ƚϡ�У����Ч��%��=[�������鷢����-�����鷢���ʣ�/�����鷢����]��100��2 ���
2.1 �������������Ӫ���ɷַ���
���������������ƷӪ���ɷֲⶨ�������1,ˮ�ֺ�����Χ27.60%��33.60%,ƽ��ֵ29.74%;pH��Χ6.2��9.5,ƽ��ֵ7.56;�л��ʺ�����Χ38.90%��48.50%,ƽ��ֵ44.46%;ȫ��������Χ2.00%��2.62%,ƽ��ֵ2.23%;��ֳ�Ậ����Χ9.31%��12.90%,ƽ��ֵ11.20%;����ά������Χ11.30%��17.80%,ƽ��ֵ14.06%��Table 1
��1
��1������������Ӫ���ɷַ���
Table 1
| ��Ʒ��� Sample number | ˮ�ֺ��� Water content (%) | pH | �л��� Organic material (%) | ȫ�� Total nitrogen (%) | ��ֳ�� Humic acid (%) | ����ά Coarse fiber (%) |
|---|---|---|---|---|---|---|
| g1 | 31.60 | 9.20 | 45.30 | 2.42 | 11.10 | 11.40 |
| g2 | 27.60 | 7.10 | 38.90 | 2.31 | 9.31 | 11.30 |
| g3 | 27.70 | 6.30 | 45.90 | 2.00 | 9.88 | 17.80 |
| g4 | 33.60 | 6.20 | 45.10 | 2.10 | 11.40 | 14.80 |
| g5 | 31.40 | 9.50 | 42.20 | 2.08 | 12.70 | 12.80 |
| g6 | 28.70 | 8.10 | 45.30 | 2.62 | 12.90 | 13.00 |
| g7 | 27.60 | 6.50 | 48.50 | 2.09 | 11.10 | 17.30 |
| ƽ��ֵAverage | 29.74 | 7.56 | 44.46 | 2.23 | 11.20 | 14.06 |
�´��ڴ�|����CSV
2.2 �������������ѿ�߸˾��ܻ�����ⶨ
�Ӳɼ���7�����������������Ʒ��Ʒ�з����ǵ�100����������,����16S rRNA����,���ƶ�>97%,��������Ϊ23�֣����֣�ѿ�߸˾���2���������ֵ�ѿ�߸˾�����2��;ѿ�߸˾�������ⶨ�������3,��Ʒ��ѿ�߸˾��������ƽ��ֵ��ΧΪ��17.67��26.67����107 cfu/g,���������������Ʒ�����ƽ��ֵ��20.62��107 cfu/g��Table 2
��2
��2���������������Ʒ��Ʒѿ�߸˾��������
Table 2
| ������ Colony number | ������ Strain number | ������ѧ�� Name of Bacillus-like species | 16S rDNA���ƶ� 16S rDNA similarity (%) |
|---|---|---|---|
| 1. | FJAT-46225 | [1] ѿ�߸˾�Bacillus sp. | 97.13 |
| 2. | FJAT-46301 | [2] ����ѿ�߸˾�Bacillus aerophilus 28KT | 100.00 |
| 3. | FJAT-46217 | 100.00 | |
| 4. | FJAT-46253 | [3] �ߵ�ѿ�߸˾�Bacillus altitudinis 41KF2bT | 100.00 |
| 5. | FJAT-46318 | [4] �����ѿ�߸˾�Bacillus amyloliquefaciens FZB42T | 99.72 |
| 6. | FJAT-46213 | [5] ����ѿ�߸˾�Bacillus cereus ATCC 14579T | 99.70 |
| 7. | FJAT-46201 | 99.64 | |
| 8. | FJAT-46307 | [6] ����ѿ�߸˾�Bacillus aryabhattai B8W22T | 100.00 |
| 9. | FJAT-46281 | 97.89 | |
| 10. | FJAT-46278 | 100.00 | |
| 11. | FJAT-46272 | 100.00 | |
| 12. | FJAT-46223 | 100.00 | |
| 13. | FJAT-46186 | 99.86 | |
| 14. | FJAT-46270 | [7] ������ѿ�߸˾�Bacillus clausii DSM 8716T | 99.50 |
| 15. | FJAT-46194 | 99.20 | |
| ������ Colony number | ������ Strain number | ������ѧ�� Name of Bacillus-like species | 16S rDNA���ƶ� 16S rDNA similarity (%) |
| 16. | FJAT-46263 | [8] ���ѿ�߸˾�Bacillus eiseniae A1-2T | 99.58 |
| 17. | FJAT-46178 | 99.72 | |
| 18. | FJAT-46289 | [9] ˿״ѿ�߸˾�Bacillus filamentosus SGD-14T | 100.00 |
| 19. | FJAT-46204 | 100.00 | |
| 20. | FJAT-46297 | [10] �º�ѿ�߸˾�Bacillus kochii WCC 4582T | 99.93 |
| 21. | FJAT-46258 | 99.79 | |
| 22. | FJAT-46246 | 100.00 | |
| 23. | FJAT-46230 | 99.65 | |
| 24. | FJAT-46221 | 99.65 | |
| 25. | FJAT-46208 | 99.72 | |
| 26. | FJAT-46202 | 99.86 | |
| 27. | FJAT-46176 | 98.86 | |
| 28. | FJAT-46175 | 99.72 | |
| 29. | FJAT-46313 | [11] ����ѿ�߸˾�Bacillus licheniformis ATCC 14580T | 99.10 |
| 30. | FJAT-46312 | 99.30 | |
| 31. | FJAT-46296 | 99.70 | |
| 32. | FJAT-46271 | 99.37 | |
| 33. | FJAT-46268 | 99.23 | |
| 34. | FJAT-46266 | 99.37 | |
| 35. | FJAT-46260 | 99.40 | |
| 36. | FJAT-46244 | 99.72 | |
| 37. | FJAT-46239 | 99.30 | |
| 38. | FJAT-46237 | 97.70 | |
| 39. | FJAT-46235 | 99.16 | |
| 40. | FJAT-46233 | 99.37 | |
| 41. | FJAT-46218 | 98.30 | |
| 42. | FJAT-46215 | 99.65 | |
| 43. | FJAT-46212 | 97.50 | |
| 44. | FJAT-46203 | 99.93 | |
| 45. | FJAT-46200 | 99.72 | |
| 46. | FJAT-46196 | 99.79 | |
| 47. | FJAT-46188 | 99.60 | |
| 48. | FJAT-46182 | 99.70 | |
| 49. | FJAT-46180 | 99.60 | |
| 50. | FJAT-46173 | 99.23 | |
| 51. | FJAT-46197 | [12] ��Ӫ����ѿ�߸˾�Bacillus methylotrophicus KACC 13105T | 100.00 |
| 52. | FJAT-46274 | [13] ����ѿ�߸˾�Bacillus rhizosphaerae SC-N012T | 99.09 |
| 53. | FJAT-46174 | [14] ɳ��ѿ�߸˾�Bacillus safensis FO-36bT | 100.00 |
| 54. | FJAT-46311 | [15] ��ŵ��ɳĮѿ�߸˾�Bacillus sonorensis NBRC 101234T | 99.44 |
| 55. | FJAT-46255 | 99.30 | |
| 56. | FJAT-46199 | 99.37 | |
| 57. | FJAT-46177 | 99.44 | |
| ������ Colony number | ������ Strain number | ������ѧ�� Name of Bacillus-like species | 16S rDNA���ƶ� 16S rDNA similarity (%) |
| 58. | FJAT-46316 | [16] �ݲ�ѿ�߸˾���������Bacillus subtilis subsp. inaquosorum KCTC 13429T | 99.72 |
| 59. | FJAT-46298 | 99.93 | |
| 60. | FJAT-46295 | 99.93 | |
| 61. | FJAT-46279 | 99.93 | |
| 62. | FJAT-46259 | 99.93 | |
| 63. | FJAT-46252 | 99.79 | |
| 64. | FJAT-46251 | 99.72 | |
| 65. | FJAT-46248 | 99.93 | |
| 66. | FJAT-46247 | 99.93 | |
| 67. | FJAT-46242 | 99.79 | |
| 68. | FJAT-46238 | 99.93 | |
| 69. | FJAT-46231 | 99.93 | |
| 70. | FJAT-46229 | 99.93 | |
| 71. | FJAT-46220 | 99.37 | |
| 72. | FJAT-46211 | 99.93 | |
| 73. | FJAT-46206 | 99.93 | |
| 74. | FJAT-46185 | 99.79 | |
| 75. | FJAT-46184 | 99.93 | |
| 76. | FJAT-46262 | [17] �ݲ�ѿ�߸˾��ݲ�����Bacillus subtilis subsp. subtilis NCIB 3610T | 99.93 |
| 77. | FJAT-46241 | 99.93 | |
| 78. | FJAT-46234 | 99.93 | |
| 79. | FJAT-46219 | 99.93 | |
| 80. | FJAT-46214 | 99.93 | |
| 81. | FJAT-46190 | 99.93 | |
| 82. | FJAT-46179 | 99.93 | |
| 83. | FJAT-46314 | [18] �ػ���ѿ�߸˾�Bacillus tequilensis KCTC 13622T | 100.00 |
| 84. | FJAT-46257 | 100.00 | |
| 85. | FJAT-46250 | 99.79 | |
| 86. | FJAT-46216 | 100.00 | |
| 87. | FJAT-46191 | 99.86 | |
| 88. | FJAT-46249 | [19] �˲ι������Ͼ�Bhargavaea ginsengige 14T | 99.79 |
| 89. | FJAT-46227 | 99.93 | |
| 90. | FJAT-46224 | 99.93 | |
| 91. | FJAT-46192 | 99.79 | |
| 92. | FJAT-46276 | [20] ������ѿ�߸˾�Lysinibacillus sp. | 97.76 |
| 93. | FJAT-46275 | [21] �������ѿ�߸˾�Oceanobacillus caeni S-11T | 99.30 |
| 94. | FJAT-46254 | [22] ʰ������ѿ�߸˾�Ornithinibacillus scapharcae TW25T | 98.47 |
| 95. | FJAT-46303 | [23] �η�����֦ѿ�߸˾�Virgibacillus halodenitrificans DSM10037T | 99.79 |
| 96. | FJAT-46195 | 99.86 | |
| 97. | FJAT-46309 | [24] ����֦ѿ�߸˾�Virgibacillus oceani MY11T | 98.70 |
| 98. | FJAT-46232 | 98.30 | |
| 99. | FJAT-46198 | 98.29 | |
| 100. | FJAT-46222 | [25] �κ�֦ѿ�߸˾�Virgibacillus salinus XH-22T | 98.48 |
�´��ڴ�|����CSV
Table 3
��3
��3�������������ѿ�߸˾��������
Table 3
| ������� Sample number | ѿ�߸˾��������Colony forming unit of Bacillus species (��107cfu/g) | ƽ��ֵ Average (��107cfu/g) | ||
|---|---|---|---|---|
| �ظ�I Rep1 | �ظ�II Rep2 | �ظ�III Rep3 | ||
| g1 | 20 | 31 | 29 | 26.67 |
| g2 | 23 | 18 | 22 | 21.00 |
| g3 | 19 | 16 | 18 | 17.67 |
| g4 | 26 | 18 | 21 | 21.67 |
| g5 | 18 | 19 | 18 | 18.33 |
| g6 | 21 | 15 | 19 | 18.33 |
| g7 | 19 | 22 | 21 | 20.67 |
| ƽ��ֵ Average (��107cfu/g) | 20.85 | 19.85 | 21.14 | 20.62 |
�´��ڴ�|����CSV
2.3 ���ں���������Ͼ�����Ʒϸ��������ⶨ
2.3.1 ���������������Ʒ���������� ���ں����������Ͼ���ϸ��������ⶨ�������4���Բɼ������������������,��Ʒ��������ƽ��ֵΪ99 701.75,�����������������Ԫ��operational taxonomic units,OTU��ƽ��ֵΪ1 469.29,����ϸ�������������;Aceָ����Chaoָ����������ָ��,ƽ��ֵ�ֱ�Ϊ1 726.42��1 757.57,ָʾ��ϸ�����ֵ�����,�����ࣨOTU��������У��ֵ;�������ָ����Coverage��ƽ��ֵΪ0.9972,������������Ѿ��������ǵ����������е�����;��ũָ����Shannon��Ϊ5.0686,����ϸ�����ֵĶ����Խϸ�,����ɭָ����Simpson��Ϊ0.0188,�����������ƶȽϵ�,����һ����ռ�о������ơ�Table 4
��4
��4���ں������������Ͼ���ϸ��������ⶨ
Table 4
| ���� Sample | ������ Reads | ���� OTUs | ��0.97�����ƶ���Under the similarity of 0.97 | ||||
|---|---|---|---|---|---|---|---|
| Aceָ�� Ace index | Chaoָ�� Chao index | �������ָ�� Coverage index | ��ũ������ָ�� Shannon index | ����ɭ���ƶ�ָ�� Simpson index | |||
| g1 | 102575 | 1608 | 1824 (1779, 1882) | 1879 (1809, 1973) | 0.997114 | 5.14 (5.13, 5.15) | 0.0212 (0.0208, 0.0216) |
| g2 | 129891 | 1698 | 1922 (1875, 1981) | 1973 (1902, 2068) | 0.997721 | 5.55 (5.54, 5.55) | 0.0101 (0.01, 0.0103) |
| g3 | 83279 | 1255 | 1604 (1536, 1689) | 1592 (1511, 1700) | 0.995845 | 4.68 (4.67, 4.70) | 0.0252 (0.0248, 0.0255) |
| g4 | 137195 | 1192 | 1381 (1337, 1438) | 1421 (1356, 1512) | 0.998287 | 4.85 (4.85, 4.86) | 0.0187 (0.0185, 0.0189) |
| g5 | 119811 | 1711 | 1997 (1942, 2067) | 2047 (1966, 2153) | 0.997087 | 5.11 (5.10, 5.12) | 0.0255 (0.0251, 0.026) |
| g6 | 111290 | 1257 | 1566 (1503, 1646) | 1574 (1493, 1683) | 0.997232 | 4.96 (4.95, 4.97) | 0.0153 (0.0151, 0.0155) |
| g7 | 113573 | 1564 | 1791 (1744, 1851) | 1817 (1752, 1904) | 0.997385 | 5.19 (5.17, 5.20) | 0.0159 (0.0157, 0.0161) |
| ƽ��ֵAverage | 99701.75 | 1469.29 | 1726.42 | 1757.57 | 0.997239 | 5.0686 | 0.0188 |
�´��ڴ�|����CSV
2.3.2 ���Ͼ�����ˮƽϸ����������������reads���ֲ� 7���������������������������������京��39����ϸ��;��ͬ������������,��������Χ��83 279��137 195,ƽ��ֵΪ113 944.85;ǰ7��ϸ����readsƽ��ֵ�ܺ�Ϊ108 242.43,ռ��95%;����С����7��ϸ���ŷֱ�Ϊ��˾��ţ�Bacteroidetes,reads=30 783.14�������ξ��ţ�Proteobacteria,27 106.29�������߾��ţ�Actinobacteria,19 740.00������ھ��ţ�Firmicutes,10 725.29�����ǻ�ϸ���ţ�Saccharibacteria,8 005.00�����쳣���-���Ⱦ��ţ�Deinococcus-Thermus,6 360.71����������ţ�Chloroflexi,5 522.00����ͼ2����
ͼ2
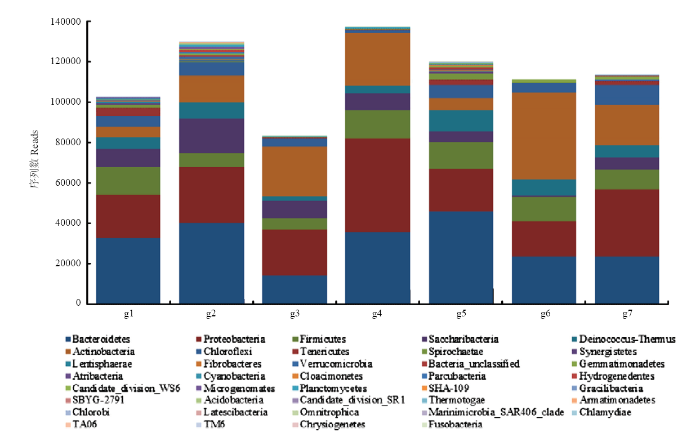 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2���Ͼ�����ˮƽϸ����������������reads��
Fig. 2Quantity reads of bacteria microbiome at the phylum level of the integrated microbiome agent
2.3.3 ���Ͼ���ϸ��������������Ԫ���ࣨOTU������ ���Ͼ���ϸ��������������Ԫ���ࣨOTU�������ֱ�Ϊϸ����39��,ϸ����96��,ϸ��Ŀ189��,ϸ����383��,ϸ����786��,ϸ����1 281��;��������������������зḻ��ϸ������,ÿ�˾�������ϸ��1 281�֡�
2.3.4 �������������ǰ10����������reads�������ߵ�ϸ������ �����������ϸߵ�ǰ10��ϸ���ֱ�����:�ٻ�ɫ�˾�����Luteibacter sp.,reads= 8 067.71�����Ǹ˾���ij����Saccharibacteria, 7 937.00���������������Truepera sp.,5 771.86����Į�Ӹ˾�����Moheibacter sp.,5 552.14�����Ƹ˾�����Flavobacterium sp.,4 261.14������֬�˾�����Crenotalea sp.,3 443.43����������������Saprospira sp,2 718.86�������˾�����Marinobacter sp.,2 324.71���������������Oceanococcus sp.,1 916.43������˾�����Ornithobacterium sp.,1 665.85��,ǰ10��ϸ�����������ܺ�Ϊ43 659.14,ռ���������������1 281��ϸ����������113 944.90��38.31%��
2.3.5 �������������ѿ�߸˾������� ��7�������й���46��ѿ�߸˾�,������ѿ�߸˾�����ֶȲ�������,ѿ�߸˾�����ܺͷ�Χ��0.4244%��3.5710%;ǰ10�ָ߷��ѿ�߸˾��ֱ�Ϊѿ�߸˾�2��Bacillus sp.2,���=0.4698%������˾�1��Lactobacillus sp.1,0.2557%����ʳ������˾���Lactobacillus amylovorus,0.2144%������ѿ�߸˾���Tuberibacillus sp.,0.1364%������ϸѿ�߸˾���Gracilibacillus sp.,0.057%������ѿ�߸˾�1��Geobacillus sp.1,0.0529%������������ѿ�߸˾���Halolactibacillus halophilus,0.0429%������ѿ�߸˾�2��Paenibacillus sp.2,0.031%������������������ѿ�߸˾���Ornithinibacillus californiensis,0.0152%��������ѿ�߸˾�3��Paucisali bacillus sp.3,0.013%��;46��ѿ�߸˾��ܷ��Ϊ1.4244%����5����
Table 5
��5
��5���������������Ʒ��ѿ�߸˾�������
Table 5
| ���� Species name | ���Species richness (%) | ƽ��ֵ Average (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| g1 | g2 | g3 | g4 | g5 | g6 | g7 | ||
| [1] ����֬��ѿ�߸˾� Ammoniibacillus agariperforans | 0.0000 | 0.0015 | 0.0000 | 0.0000 | 0.0000 | 0.0162 | 0.0000 | 0.0025 |
| [2] ����ѿ�߸˾�Amphibacillus sp. | 0.0000 | 0.0000 | 0.0060 | 0.0160 | 0.0000 | 0.0036 | 0.0018 | 0.0039 |
| [3] ������ѿ�߸˾�Aneurinibacillus sp. | 0.0000 | 0.0008 | 0.0048 | 0.0000 | 0.0000 | 0.0009 | 0.0000 | 0.0009 |
| [4] ����ѿ�߸˾�Bacillus azotoformans | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| [5] ��ƺѿ�߸˾� Bacillus graminis | 0.0010 | 0.0023 | 0.0036 | 0.0015 | 0.0008 | 0.0018 | 0.0000 | 0.0016 |
| [6] ����ѿ�߸˾�Bacillus humi | 0.0010 | 0.0008 | 0.0084 | 0.0015 | 0.0025 | 0.0234 | 0.0009 | 0.0055 |
| [7] ���ѿ�߸˾� Bacillus infernus | 0.0039 | 0.0054 | 0.0144 | 0.0036 | 0.0025 | 0.0036 | 0.0343 | 0.0097 |
| [8] ѿ�߸˾�1 Bacillus sp.1 | 0.0000 | 0.0108 | 0.0000 | 0.0000 | 0.0659 | 0.0009 | 0.0009 | 0.0112 |
| [9] ѿ�߸˾�2 Bacillus sp.2 | 0.0614 | 0.2079 | 1.2836 | 0.4665 | 0.1327 | 0.7332 | 0.4033 | 0.4698 |
| [10] ����ѿ�߸˾�Bacillus thermolactis | 0.0010 | 0.0062 | 0.0120 | 0.0036 | 0.0025 | 0.0108 | 0.0158 | 0.0074 |
| [11] �ȼ�ѿ�߸˾�CaldalkaLibacillus sp. | 0.0000 | 0.0015 | 0.0000 | 0.0000 | 0.0008 | 0.0099 | 0.0018 | 0.0020 |
| [12] ����ѿ�߸˾� Desulfuribacillus sp. | 0.0088 | 0.0000 | 0.0000 | 0.0000 | 0.0008 | 0.0000 | 0.0000 | 0.0014 |
| [13] ��ѿ�߸˾�1 Geobacillus sp.1 | 0.0000 | 0.0054 | 0.0012 | 0.0000 | 0.0017 | 0.3603 | 0.0018 | 0.0529 |
| [14] ������֬����ѿ�߸˾�Geobacillus stearothermophilus | 0.0000 | 0.0015 | 0.0012 | 0.0029 | 0.0017 | 0.0018 | 0.0009 | 0.0014 |
| [15] ��ϸѿ�߸˾�Gracilibacillus sp. | 0.0029 | 0.0100 | 0.0036 | 0.0058 | 0.0017 | 0.3423 | 0.0326 | 0.0570 |
| [16] ϲ��ѿ�߸˾�Halobacillus sp. | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| [17] ��������ѿ�߸˾� Halolactibacillus halophilus | 0.0556 | 0.0031 | 0.0108 | 0.1844 | 0.0225 | 0.0117 | 0.0123 | 0.0429 |
| [18] ����ѿ�߸˾�Halothiobacillus sp. | 0.0010 | 0.0000 | 0.0012 | 0.0146 | 0.0000 | 0.0018 | 0.0044 | 0.0033 |
| [19] ʳ������˾�Lactobacillus amylovorus | 0.1950 | 0.0254 | 0.0588 | 0.0722 | 0.0250 | 1.0145 | 0.1101 | 0.2144 |
| [20] ������˾�Lactobacillus animalis | 0.0010 | 0.0023 | 0.0000 | 0.0036 | 0.0000 | 0.0036 | 0.0000 | 0.0015 |
| [21] ��˾����� Lactobacillus equicursoris | 0.0010 | 0.0062 | 0.0000 | 0.0007 | 0.0000 | 0.0584 | 0.0044 | 0.0101 |
| [22] ճĤ��˾�Lactobacillus mucosae | 0.0000 | 0.0008 | 0.0000 | 0.0015 | 0.0000 | 0.0306 | 0.0000 | 0.0047 |
| [23] �����˾�Lactobacillus sakei | 0.0000 | 0.0008 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0001 |
| [24] ��˾�1 Lactobacillus sp.1 | 0.3246 | 0.0477 | 0.1465 | 0.4497 | 0.0209 | 0.6676 | 0.1330 | 0.2557 |
| [25] С�������ѿ�߸˾�Oceanobacillus oncorhynchi | 0.0000 | 0.0008 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0001 |
| [26] ����ѿ�߸˾�1 Oceanobacillus sp.1 | 0.0000 | 0.0000 | 0.0000 | 0.0066 | 0.0000 | 0.0045 | 0.0026 | 0.0020 |
| [27] ����ѿ�߸˾�2 Oceanobacillus sp.2 | 0.0049 | 0.0023 | 0.0024 | 0.0117 | 0.0000 | 0.0521 | 0.0070 | 0.0115 |
| [28] ��������������ѿ�߸˾� Ornithinibacillus californiensis | 0.0049 | 0.0154 | 0.0144 | 0.0117 | 0.0008 | 0.0279 | 0.0317 | 0.0152 |
| [29] ����ѿ�߸˾�DX-3 Ornithinibacillus sp. DX-3 | 0.0019 | 0.0008 | 0.0060 | 0.0022 | 0.0025 | 0.0350 | 0.0352 | 0.0120 |
| [30] ����ѿ�߸˾�GD05 Ornithinibacillus sp. GD05 | 0.0000 | 0.0008 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0026 | 0.0005 |
| [31] ����ѿ�߸˾�HME7715 Ornithinibacillus sp. HME7715 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0180 | 0.0000 | 0.0026 |
| [32] ��ѿ�߸˾�1 Paenibacillus sp.1 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0062 | 0.0009 |
| [33] ��ѿ�߸˾�2 Paenibacillus sp.2 | 0.0078 | 0.0331 | 0.0096 | 0.0204 | 0.0150 | 0.0494 | 0.0819 | 0.0310 |
| [34] ����ѿ�߸˾�1 Paucisalibacillus sp.1 | 0.0000 | 0.0000 | 0.0024 | 0.0000 | 0.0008 | 0.0009 | 0.0009 | 0.0007 |
| [35] ����ѿ�߸˾�2 Paucisalibacillus sp.2 | 0.0029 | 0.0115 | 0.0060 | 0.0087 | 0.0025 | 0.0099 | 0.0123 | 0.0077 |
| ���� Species name | ���Species richness (%) | ƽ��ֵ Average (%) | ||||||
| g1 | g2 | g3 | g4 | g5 | g6 | g7 | ||
| [36] ����ѿ�߸˾�3 Paucisalibacillus sp.3 | 0.0049 | 0.0000 | 0.0072 | 0.0095 | 0.0025 | 0.0584 | 0.0088 | 0.0130 |
| [37] ���³÷��ѿ�߸˾�Rummeliibacillus pycnus | 0.0146 | 0.0062 | 0.0048 | 0.0073 | 0.0000 | 0.0045 | 0.0018 | 0.0056 |
| [38] ɭ������ѿ�߸˾�Solibacillus silvestris | 0.0049 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0009 | 0.0000 | 0.0008 |
| [39] ��ѿ�߸˾�Sulfobacillus sp. | 0.0000 | 0.0023 | 0.0024 | 0.0000 | 0.0000 | 0.0000 | 0.0018 | 0.0009 |
| [40] ��ѿ�߸˾�1 Thermobacillus sp.1 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0008 | 0.0000 | 0.0114 | 0.0018 |
| [41] ��ѿ�߸˾�2 Thermobacillus sp.2 | 0.0000 | 0.0069 | 0.0024 | 0.0000 | 0.0008 | 0.0036 | 0.0018 | 0.0022 |
| [42] ��ѿ�߸˾�3 Thermobacillus sp.3 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 | 0.0000 |
| [43] ������ѿ�߸˾� Tuberibacillus calidus | 0.0000 | 0.0008 | 0.0024 | 0.0029 | 0.0000 | 0.0009 | 0.0106 | 0.0025 |
| [44] ��ѿ�߸˾�Tuberibacillus sp. | 0.0000 | 0.0000 | 0.0216 | 0.8951 | 0.0000 | 0.0000 | 0.0379 | 0.1364 |
| [45] ����ѿ�߸˾�Ureibacillus sp. | 0.0058 | 0.0023 | 0.0000 | 0.0000 | 0.0008 | 0.0054 | 0.0141 | 0.0041 |
| [46] ��ɽѿ�߸˾�Vulcanibacillus sp. | 0.0273 | 0.0008 | 0.0000 | 0.0153 | 0.0442 | 0.0027 | 0.0009 | 0.0130 |
| �ܺ�Total | 0.7381 | 0.4244 | 1.6377 | 2.2195 | 0.3527 | 3.5710 | 1.0278 | 1.4244 |
�´��ڴ�|����CSV
�ڼ���46��ѿ�߸˾�����9��ѿ�߸˾�δ�������о�����,�ֱ�Ϊ:��1������ѿ�߸˾���Bacillus aerophilus��,��ӡ��****SHIVAJI��[21]�ӿ����ɼ��������з������������;��2�����ѿ�߸˾���Bacillus eiseniae��,�ɺ���****HONG��[22]�������������������;��3��˿״ѿ�߸˾���Bacillus filamentosus��,��ӡ��****SONALKAR��[23]�Ӻ����������������;��4���º�ѿ�߸˾���Bacillus kochii��,�ɵ¹�****SEILER ��[24]��ʳƷҩ��ӹ������������;��5������ѿ�߸˾���Bacillus rhizosphaerae��,��ӡ��****MADHAIYAN��[25]�Ӹ�������������������;��6������������ѿ�߸˾���Lysinibacillus macroides��,�ɱ���ʱ****COOREVITS��[26]1947����ռ���ţ���з���ľ������������õ�����;��7���������ѿ�߸˾���Oceanobacillus caeni��,�ɺ���****NAM��[27]����ˮ����ϵͳ�з��������;��8��ʰ������ѿ�߸˾���Ornithinibacillus scapharcae��,�ɺ���****SHIN��[28]�Ӻ��������ı����з��������;��9������֦ѿ�߸˾���Virgibacillus oceani��,���й�****YIN��[29]����̫ƽ��������з�������֡�����9����Ϊ�й��¼�¼�֡�����2����,����10���κ�֦ѿ�߸˾���Virgibacillus salinus��,��������****CARRASCO��[30]���й����ɹ����ֺ����κ��з��������;��11����ݶ�ѿ�߸˾���Bacillus vanilla��,���й�****CHEN��[31]�Ӻ�����ݶ��з��������,�����ڹ��ڵ��о�δ������,���������ڹ��ڻ����ɼ�������,Ϊ�й��ֲ��֡�
2.4 ������������������ӷ�ѿ��Ӱ��
�������̶���ѿ�ʺͷ�ѿָ������ˮ�������������죨P>0.05��,���������̶��߸���ƽ��ֵΪ�������1.58��,���켫������P<0.01��;��ȶ�����,�������̶�����ָ�������56.26%,���켫������P<0.01������6����ͼ2�������,���������������Ʒ����Һ���̶���ѿ��Ӱ��,�������Դٽ��̶�����������,�����ָ����Table 6
��6
��6����������������̶���ѿ��Ӱ��
Table 6
| ���� Treatment | ��ѿ�� Germination rate (%) | �߸���Radical length (RL, cm) | ��ѿָ��Germination index (GI) | ����ָ��Vigor index (VI) | |||
|---|---|---|---|---|---|---|---|
| RL | ��ֵRatio | GI | ��ֵRatio | VI | ��ֵRatio | ||
| ���Ͼ���IMA | 96.67��2.11a | 2.64��0.77A | 1.5808 | 57.67a | 0.9885 | 152.25��13.29A | 1.5626 |
| ��ˮ����CK | 98.33��1.53a | 1.67��0.23B | 1.0000 | 58.34a | 1.0000 | 97.43��10.67B | 1.0000 |
�´��ڴ�|����CSV
2.5 ��������������Է���Ѩ������׳�����õ�Ӱ��
������������Ͼ�����������ʱ,�������Ʒ�����ij�����,����ճ�����86.20%���,�����ʽ���9.04%��46.10%;ͬʱ�����˷��Ѹ�����0.70��1.45 cm������ߣ�1.50��6.23 cm���;��֣�0.70��1.05 cm�������Ͼ�����Ũ������ʱ,���ѵij����ʣ�85.30%��86.03%����������4.45��4.85 cm������ߣ�9.67��9.78 cm�������֣�1.45��1.56 cm�������������������졣���Ͼ����ͺ�������ʱ,��������߷��ѳ�����3.0%,�����������25.1%,���Ը����;���Ӱ�첻��������7����Table 7
��7
��7��������������Է���Ѩ�������Ӱ��
Table 7
| ������ Treatment | ������� Treatment number | ���Ͼ�������IMA percent (%) | ������������״��Growth of tomato seedlings | |||
|---|---|---|---|---|---|---|
| ������ Emergence rate (%) | ���� Root length (cm) | ��� Plant height (cm) | ���� Stem diameter (cm) | |||
| �ߺ����� High content group | ����1 Treatment 1 | 30 | 9.04 | 0.70 | 1.50 | 0.70 |
| ����2 Treatment 2 | 25 | 46.10 | 1.45 | 6.23 | 1.05 | |
| ƽ��ֵAverage | �� | 27.57c | 1.08b | 3.87c | 0.88b | |
| ����� Middle content group | ����3 Treatment 3 | 20 | 85.30 | 4.45 | 9.78 | 1.56 |
| ����4 Treatment 4 | 15 | 86.03 | 4.85 | 9.67 | 1.45 | |
| ƽ��ֵAverage | �� | 85.67b | 4.65a | 9.73b | 1.51a | |
| �ͺ����� Low content group | ����5 Treatment 5 | 10 | 89.50 | 4.73 | 10.67 | 1.49 |
| ����6 Treatment 6 | 5 | 88.90 | 4.62 | 11.92 | 1.51 | |
| ƽ��ֵAverage | �� | 89.20a | 4.68a | 11.30a | 1.50a | |
| �հ���CK | ���� CK | 0 | 86.20b | 4.68a | 9.03b | 1.40a |
�´��ڴ�|����CSV
2.6 ��������������Է���Ѩ��������ݲ����ص�Ӱ��
�����Ͼ������Ƶ��������������������,�ڳ���30 d����������ַ�������,��ǿ�²���������Ͼ���Һ��106 cfu/mL���������,����������˸ߺ������Ͼ�������1������ʵ���ͳ��,���ദ���Ӿ���4 d��ʼ����,����ʱ����̷�����������,10 d�ﵽ�߷�;�ڷ����߷��ڣ�10 d��,�����飨CK�������ʴ�88.89%,�ߺ������Ͼ���������ƽ��������Ϊ66.67%,У����Ч��17.88%;�к������Ͼ���������ƽ��������Ϊ30.45%,У����ЧΪ62.49%,�ͺ������Ͼ���������ƽ��������Ϊ16.72%,У����ЧΪ79.41%;��ݲ�����Ч��Ϊ�ߺ�����<�к�����<�ͺ����飨��8��ͼ3����Table 8
��8
��8��������������Է���������ݲ����ص�Ӱ��
Table 8
| ������ Treatment | ������� Treatment number | ���Ͼ������� IMA percent (%) | ����30 d�������ݲ�ԭ��Һ Inoculation of R. solanacearum 30 days after emergence (106 cfu/mL) | 10 dУ����ЧCorrected control efficacy of 10 d (%) | ||
|---|---|---|---|---|---|---|
| 4 d���� Incidence after 4 d (%) | 7 d���� Incidence after 7 d (%) | 10 d���� Incidence after 10 d (%) | ||||
| �ߺ����� High content group | ����1 Treatment 1 | 30 | �� | �� | �� | |
| ����2 Treatment 2 | 25 | 6.25 | 23.00 | 66.67 | ||
| ƽ��ֵAverage | �� | 6.25b | 23.00a | 66.67a | 17.88 | |
| ����� Middle content group | ����3 Treatment 3 | 20 | 6.47 | 13.33 | 32.40 | |
| ����4 Treatment 4 | 15 | 8.33 | 16.57 | 28.50 | ||
| ƽ��ֵAverage | �� | 7.40ab | 14.95b | 30.45ab | 62.49 | |
| �ͺ����� Low content group | ����5 Treatment 5 | 10 | 6.67 | 14.47 | 18.30 | |
| ����6 Treatment 6 | 5 | 6.67 | 13.33 | 15.14 | ||
| ƽ��ֵAverage | �� | 6.67b | 13.90b | 16.72b | 79.41 | |
| �հ���CK | ���� CK | 0 | 11.11a | 33.33a | 88.89a | �� |
�´��ڴ�|����CSV
ͼ3
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3��������������Է���Ѩ�����缰��ݲ����ص�Ӱ��
Fig. 3Effect of the integrated microbiome agent on tomato plug seedling and biocontrol of bacterial wilt
3 ����
3.1 �������������������з���Ӧ��
������,��������о���Ϊ�ȵ�,�����ڻ����ж������������ʽ���ֺͷ������á�����������Ӱ���Ŷ��������Ľ���[32],����������ƽ����ˮ����̬ϵͳ[33],��������������������������ֲ�。��[34],�ȵȡ���������������������,ͨ�����ʹ������������Ͳ�������������,�ӹ��ɾ���,��������������������䵽��������,��������������Ľ���ˮƽ,Ӱ�����������������,ʹ֮���������������������ӡ�������������Լ�ֲ�。�������������,�����������������Ʋ�������,ͬʱ��������Ϻ�����ũҩ�����á���Ϊ��ũҩ����˫���ж������Ʒ������Ҫ��ֵ,Ҳ�����ݷ����Դ���ʻ����õ�һ����Ҫ;�������ݷ����ũҵ�ոѽ�ϵ�ũҵ���������������л��ʷ�ʽ�õ�����ķ�չ��ͨ���ѷʷ�ʽ,��ũҵ������ͨ������ת���ɸ�ֳ�ʣ��л��ʣ����о��н϶౨��[35,36,37]���[38]�����˶ѷʷ��ͽ��ּ�����ģ�������л���;�ŷ�����[39]���������ݷ�����ѷʲ�Ʒ���������ʼ�����ЧӦ;��D��[40]�����˽ո��������϶ѷʡ������о��Ӷѷ�ԭ�ϡ�Ӫ�����ӡ�ͨ�����ơ�����ɸѡ�ȽǶȽ��������л��ʵ�����,Ȼ��ũҵ�����ᆳ����ֵĶѷʸ���,��ʧ�˴�����������,��Ч��������������½�,���ղ�Ʒ��Ч�������Ȼ���������л���ũҵ��ҵ����NY884-2012��,������Ч���������2��107 cfu/g���ҡ����ݷ��Ķѷʷ���������������������ʹ����Žϴ����,�ѷ��ǰ�������ոѵ�̼�����䷽,һ�����ӻ�Ϸ�����,����ɱ���˴���������;����������������������÷��ʹ����з��۵���Ӫ����������,�����ӵ����ͽ���,�����������˷ḻ�������顣
����������������������������Ͼ������ĸ���,���ù�����������������ƣ���������ͨ�����������ݷ��۵����������Ӽ���,�ڱ��Ͼ�����Ʒ�л��ʺ����Ļ�����,ʹ�����ﺬ�����;���Ͼ���������������,���ʹ��������ڳ���Ӫ����ͨ����������Ѹ����������,����20 d����,ȡ���ϲ㷢�͵��ϼӹ��γ����Ͼ���,�����������,����˷��ʹ���������,��Ч������������Դ��������������ֵ�����Ͼ���;���Ͼ�����ˮ��29.74%,pH 7.56,�л��ʺ���44.46%,ȫ������2.23%,��ֳ�Ậ��11.20%,����ά����14.06%,ѿ�߸˾��������2.062��108 cfu/g;��Ʒ��̬�������л�������,���������л����й�ũҵ��ҵ����NY884-2012���л��ʺ���>40%,��ˮ��<30%,pH 5.5��7.5�Ĺ涨,��Ч���������ͳ��ѿ�߸˾����������л��ʵ�10������,�ﵽ���л��ʺ����������Ŀ��,���ٵ��������ɴٽ�ֲ�ѿ����,����������ͬʱ,���Ͼ������Ӳ�ͬ�ĵ��ϣ���������,�綹���ۡ����ǡ����۵�,��Ӱ���Ʒ����������,����о������ı�����
3.2 ��������������е�ϸ����������
���Ͼ���ÿ�˾�������ϸ����39��,ϸ����96��,ϸ��Ŀ189��,ϸ����383��,ϸ����786��,ϸ����1 281����ǰ7��ϸ���ź����ܺ�Ϊ108 242.43,ռ��95%��ǰ10�ָߺ���ϸ��ռ���������������1 281��ϸ��������38.31%,��Ϊ���Ͼ�����Ҫ�Ĺ���ϸ�������顣����****���л��ʵķ����ռ����еij���������������������о�[26,41-43],�ڷ����������ӹ����������,�ܹ��ḻ�л����еĹ�������,��ǿ��Ч��Ȼ��,�ѷʹ�������Ȼ�Ӿ��Ĺ���,������Ӫ��������,�������ϻ��Զ���ѡ���������,���ӽ�ȥ������Զѷʷ�����Ӱ�����,�������ɷֱ仯���ѷʷ����������ѷʸ���ʱ��ȵı仯Ӱ���Ŷѷʲ�Ʒ����������,���ӽ�ȥ������Ͳ�Ʒ�е��������������û�б�Ȼ��ϵ����ͳ�ѷʹ��̺��������麬��������½�,������������������������������麬����ߵ�ʱ���������,�����˲�Ʒ���������ߺ��������Ͼ�����Ʒ��ѿ�߸˾������Ϊ2.062��108 cfu/g,�������ⶨ��ʾѿ�߸˾��ܷ��Ϊ1.42%,���������������������Чϸ�������ɴ�1.45��1010 cfu/g;���������������Ʒ�ܺõؽ���������л��ʺ�������ϵ��ص�,Ϊ���������������Ӧ�ô��������õ�������3.3 ���������������ѿ�߸˾��ķ���
ͨ���������ͺ��������,�ó����������������Ʒ�к��д�����ѿ�߸˾�;����ѿ�߸˾��������������Լ����뺬�����Ȳ���,���ַ����������ѿ�߸˾����������нϴ����;�������������뵽ѿ�߸˾�23�֣����֣���2���������ֵ�ѿ�߸˾�,�ܺ���Ϊ2.062��108 cfu/g;�������ⶨ������ѿ�߸˾�46����,�ܷ��Ϊ1.42%,��ѿ�߸˾�ռ����ϸ���������1.42%������,9����δ�����ڷ�����о��ı�����ѿ�߸˾��ܲ���ѿ��,�������õı�������,ͬʱѿ�߸˾��ܽ���ء��ֽ��л�������������ٽ�ֲ�������ȡ�����****ͨ���ѷ����������䡢�����յ��ء��������ӵȷ������ٽ������л��ʸ���,����ѿ�߸˾�����,�Ӷ���������л��ʵ�Ӧ��Ч����������������������Ӧ�������л��ʷ�չ�ķ���,ͨ�����ι��巢��,�����˾����е�ѿ�߸˾����������,Ϊ����Ӧ�÷�Χ��Ч���������ṩ��������3.4 �������������������ԭ�����շ���
�����л��ʼ���ָ�����з��Ⱥ����ָ��,�������ƶ��ﲡԭ����������Ҫ���á���������������������������Ǻ������͵Ĺ���,������÷��ʹ�����͵��ϵ���������л��ʻ���,ͨ���������·���,Ϊ�����������������˽Ϻõ�����,�Ӷ���������������Ϊ�������ݲ�ԭ�ķ�չ;����Ҫ��ԭ������Ӫ��������Ժ�����������,ͨ�������Ͼ�����Ʒ�ĺ��������δ������Ҫ����ԭ��ϸ����,����Ĥ�����߸˾���Actinobacillus pleuropneumoniae����֧���ܰ�Ѫ���ϸ˾���Brodetella bronchiseptica���������ϸ˾���Brucella suis����������Ѫ�˾���Haemophilus parasuis����ϸ������ɭ�Ͼ���Lawsonia intracellularis������ɱ���ϸ˾���Pasteurella multocida����������ɳ���Ͼ���Salmonella cholerae�������ڵ�ϸ����;�ܼ���������ԭϸ����,���������Clostridium��������˿������Erysipelothrix������³�ȿ�������Globicatella�����ٵ����˾�����Pseudomonas����,����ⲻ����Ӧ������ԭ������Ĵ���,���ⲻ��������Ĥ�����Clostridium perfringens��������������Erysipelothrix rhusiopathiae����Ѫ��³�ȿ�����Globicatella sanguinis������ŧ�ٵ�������Pseudomonas aeruginosa����;�����������Ĵ˾���Escherichia coli��Ҳ��ⲻ��,�����ٻٻ��[44]�����ķ��ʹ����ͳ̶�II�����ϣ��еȷ��ͳ̶ȣ��ĵ��Ϸ��벻�����˾��Ľ����һ�µġ������Ͼ����м�ⲻ����ϸ���Բ�ԭ��ԭ��;����ĺ������͡�Ӫ���䷽�������ӹ��Ȳ�������һ���Ĺ�ϵ,ͬʱ�����Ͼ�����������Ⱥ�������ڹ�ϵ,���Ͼ�������������һ������������ԭЯ���ķ��ա�3.5 ������������������ӷ�ѿ�����������Ͳ������ص�Ӱ��
�ų����������������ֲ���Ƿ��ж����ĵ�һ�����Dzⶨ������ӷ�ѿ��Ӱ��,���̶���Ϊ���Ͻ��������ȷ�����,���㷺�����ڲⶨ�Ƽ������ӷ�ѿ��Ӱ��,��ⶨ��Ѫ���Ƽ�[45]������[46]����ù��[47]����������[48]�ȶ��̶���ѿ�ʵ�Ӱ�졣���о�����,�������Ͼ���25%����Һ�����̶������ܹ��ٽ��̶���������,����������������̶����ӷ�ѿ������ͬ�������Ӷ���������������ķ�Ӧ��ͬ,�д��ڽ�һ���о������ѻ��������DZ���׳��Ĺؼ������û������ʣ�Ҭ����,���Ӳ�ͬŨ�ȵ��������������,���Ƴ��������,�о��Է����������ݲ����ص�Ӱ��,����������������������������߷��ѳ�����,�ٽ��������,���ҿ�����Ч������ݲ�;����������е����Ͼ�������Ϊ5%��10%�ȽϺ��ʡ�3.6 ��������������������պ�����������ȷ��
���������������������,��������ʹ��1�����ϵ�����ʹ�,����һ��10 cm���30%������+70%�ӱ�����������,���ж��ι����������,ÿ�췭��1��,������������20 d��,ȡ���ϲ�20 cm�ĵ���,������ɹ�����顢��ɸ����װ,�ӹ��������������������������Ϊ:ԭ�����á����ʹ����͡��������Ϳ��ơ���Ʒ�ӹ�����Ʒ��װ�ȡ�����������������������ο�ũҵũ�岿�����л���ũҵ��ҵ����NY884-2012��,��Ʒ����ָ�����ȷ��Ϊ:�л��ʡ�40%,��ˮ����30%,pH 5.5��7.5,���Ⱥ����100��/g,�׳���������>95%,��Ч��>6����;�ؽ������������Ҫ��:��<15 mg��kg-1,�ӡ�15 mg��kg-1,Ǧ��15 mg��kg-1,����15 mg��kg-1,����15 mg��kg-1;��Ч���������Ϊ:ϸ���������30��108 cfu/g��ʵ�ʺ����ߵö�,���ڻ����Ʒ�����ں�������˥����,����ѿ�߸˾���2��108 cfu/g��4 ����
�������������������ĸ���,���������൪���������Ӻ����º������ͼ�������������;�����Ʒ����ָ��,��ũҵũ�岿�����л��ʵ��������ӹ�,�ؽ������������Ҫ��,ѿ�߸˾���Ч�������2��108 cfu/g�����������������������Ӱ�����ӷ�ѿ,�ܴٽ���������,���Ը���������������ݲ��������õķ���Ч�����ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 1]
[��������: 1]
DOI:10.1371/journal.pone.0175934URL [��������: 3]
DOI:10.1094/PHYTO-09-12-0243-RURL [��������: 2]
[��������: 2]
[��������: 2]
DOI:10.4238/2015.March.6.7URL [��������: 4]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
URLMagsci [��������: 2]

<div >�Իƹ�Ϊ���壬�Բ�ͬ����ЧӦ(�ٽ�/����)С��Ʒ��Ϊ���壬����PCR-DGGE�������о���С���ϵ�����P����С��Իƹ��������������Ⱥ��ṹ��Ӱ��.��������� �ڴ�����6��͵�12�죬���дٽ�ЧӦС���ϵ������ֱ���������˻ƹ�������ߺ;��֣��ڴ�����18�죬���дٽ�������ЧӦС���ϵ���������������˻ƹ�������ߣ��ڴ�����6�죬��ͬ����ЧӦС���ϵ����������������˻ƹ���������������Ⱥ����������Shannonָ�������ȶ�ָ��,�������(W)���������������(Wn)���ڴ�����18�죬�����������Ⱥ��ṹ��������Shannonָ�������ȶ�ָ�������������������(Wn).������������ЧӦС�����������˻ƹϸ����������Ⱥ��Shannonָ���;��ȶ�ָ����˵��С���ϵ�����P����С��ı����������Ⱥ��ṹ.DGGEͼ�������ɷַ������������������ͬ����ЧӦС����������Ⱥ��ṹӰ��ϴ�.</div><div > </div>
URLMagsci [��������: 2]
<div >�Իƹ�Ϊ���壬�Բ�ͬ����ЧӦ(�ٽ�/����)С��Ʒ��Ϊ���壬����PCR-DGGE�������о���С���ϵ�����P����С��Իƹ��������������Ⱥ��ṹ��Ӱ��.��������� �ڴ�����6��͵�12�죬���дٽ�ЧӦС���ϵ������ֱ���������˻ƹ�������ߺ;��֣��ڴ�����18�죬���дٽ�������ЧӦС���ϵ���������������˻ƹ�������ߣ��ڴ�����6�죬��ͬ����ЧӦС���ϵ����������������˻ƹ���������������Ⱥ����������Shannonָ�������ȶ�ָ��,�������(W)���������������(Wn)���ڴ�����18�죬�����������Ⱥ��ṹ��������Shannonָ�������ȶ�ָ�������������������(Wn).������������ЧӦС�����������˻ƹϸ����������Ⱥ��Shannonָ���;��ȶ�ָ����˵��С���ϵ�����P����С��ı����������Ⱥ��ṹ.DGGEͼ�������ɷַ������������������ͬ����ЧӦС����������Ⱥ��ṹӰ��ϴ�.</div><div > </div>
URLMagsci [��������: 2]
<p>����Ͽ�ή������ֲ�꼰����ʲ����з��뵽8������������Ʒ��“����1��”Ϊ���ϣ����ûؽ�����������ȷ�����ȷ��������������ϵ��²��Ժ������������ã�ͨ���ⶨ���������ϵ�յ�ø���ԡ��������ʺ�����ϸ��Ĥ����ԣ��о������к�����ֶ��ض��������Ļ������ã�������̬ѧ������ITS���з����������к�������г�������.��������� �����к����TF��HF�ֶ��ض���������ȷ��Լ����ᡢ�߸����������������ã��ɵ�����������ϵ����ȩ(MDA)�����������Ե�������ϸ��Ĥ��������ӣ�����TF�ֶ���ԭҺ��������������ϵMDA������ϸ��Ĥ����Խ϶��շֱ�����108.6%��40.6%.�����к�����ֶ��ؿ������������ϵ�������յ�ø���ԣ����У�TF�ֶ���10��ϡ��Һ�����¸�ϵ��������ⰱø(PAL)��������ø(POD)���Խ϶��շֱ�����25.6%��23.2%��HF�ֶ���ԭҺ�����¸�ϵPAL���Խ϶������30.0%.�����к����TF��HF��������Ϊľ�����߾�(<em>Fusarium equiseti</em>)�Ͳ�����߾�(<em>F. proliferatum</em>).�������߾���Ȼ����ͨ���ؽ���Ⱦ��ϣ�����ͨ�����ڶ���Ӱ�������������������������л��ͬʱ�����ϸ�ϵ�������յ�ø���ԣ������к�������˫�����ã��仯��Σ����������������ϰ�����Ҫԭ��֮һ.</p>
URLMagsci [��������: 2]
<p>����Ͽ�ή������ֲ�꼰����ʲ����з��뵽8������������Ʒ��“����1��”Ϊ���ϣ����ûؽ�����������ȷ�����ȷ��������������ϵ��²��Ժ������������ã�ͨ���ⶨ���������ϵ�յ�ø���ԡ��������ʺ�����ϸ��Ĥ����ԣ��о������к�����ֶ��ض��������Ļ������ã�������̬ѧ������ITS���з����������к�������г�������.��������� �����к����TF��HF�ֶ��ض���������ȷ��Լ����ᡢ�߸����������������ã��ɵ�����������ϵ����ȩ(MDA)�����������Ե�������ϸ��Ĥ��������ӣ�����TF�ֶ���ԭҺ��������������ϵMDA������ϸ��Ĥ����Խ϶��շֱ�����108.6%��40.6%.�����к�����ֶ��ؿ������������ϵ�������յ�ø���ԣ����У�TF�ֶ���10��ϡ��Һ�����¸�ϵ��������ⰱø(PAL)��������ø(POD)���Խ϶��շֱ�����25.6%��23.2%��HF�ֶ���ԭҺ�����¸�ϵPAL���Խ϶������30.0%.�����к����TF��HF��������Ϊľ�����߾�(<em>Fusarium equiseti</em>)�Ͳ�����߾�(<em>F. proliferatum</em>).�������߾���Ȼ����ͨ���ؽ���Ⱦ��ϣ�����ͨ�����ڶ���Ӱ�������������������������л��ͬʱ�����ϸ�ϵ�������յ�ø���ԣ������к�������˫�����ã��仯��Σ����������������ϰ�����Ҫԭ��֮һ.</p>
URLMagsci [��������: 1]
<p>���������������£��о���֦����(AM)����ر�����ù(<em>Glomus versiforme</em>)�����������������Ը���ͼ�����������ϵĤ�ԡ�����ȩ(MDA)�����ͷ�����ø���Ե�Ӱ��.��������� ����AM������������������Ը���ͼ��������������߸�ϵ����������ϵĤ�Ժ�MDA����.����AM������Ը�����ϲ������������ϲ���������ϵ�����ֱ�������57.6%��60.0%��142.1%��������AM����ļ���ֱ�������26.7%��28.0%��11.0%���Ը��磨C�������磨G��������AM����Ը��磨C+M���ͽ���AM������磨G+M���ĸ�ϵϸ��Ĥ��ΪC>G>C+M>G+M����ϵMDA����ΪC>G>G+M>C+M.����AM�������������Ը���ͼ����ϵ�ı�������ⰱø(PAL)����������ø(CAT)����������ø(POD)��������ø��β��1,3���Ͼ���ø���ԣ����ҽ���AM����������Ը���ͼ����ϵPOD��PAL��β-1,3-�Ͼ���ø���Եķ�ֵ�Ȳ����ֵ���ǰ2�ܳ���.����AM����ܼ��������Ը���ͼ����뿹�����йصķ�����ø��Ӧ��ʹ��ϵ���澳�������ٷ�Ӧ���Ӷ�����俹�����ϰ�������.</p>
URLMagsci [��������: 1]
<p>���������������£��о���֦����(AM)����ر�����ù(<em>Glomus versiforme</em>)�����������������Ը���ͼ�����������ϵĤ�ԡ�����ȩ(MDA)�����ͷ�����ø���Ե�Ӱ��.��������� ����AM������������������Ը���ͼ��������������߸�ϵ����������ϵĤ�Ժ�MDA����.����AM������Ը�����ϲ������������ϲ���������ϵ�����ֱ�������57.6%��60.0%��142.1%��������AM����ļ���ֱ�������26.7%��28.0%��11.0%���Ը��磨C�������磨G��������AM����Ը��磨C+M���ͽ���AM������磨G+M���ĸ�ϵϸ��Ĥ��ΪC>G>C+M>G+M����ϵMDA����ΪC>G>G+M>C+M.����AM�������������Ը���ͼ����ϵ�ı�������ⰱø(PAL)����������ø(CAT)����������ø(POD)��������ø��β��1,3���Ͼ���ø���ԣ����ҽ���AM����������Ը���ͼ����ϵPOD��PAL��β-1,3-�Ͼ���ø���Եķ�ֵ�Ȳ����ֵ���ǰ2�ܳ���.����AM����ܼ��������Ը���ͼ����뿹�����йصķ�����ø��Ӧ��ʹ��ϵ���澳�������ٷ�Ӧ���Ӷ�����俹�����ϰ�������.</p>
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
DOI:10.1007/s11356-010-0381-4URL [��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.3389/fmicb.2017.02179URL [��������: 1]
[��������: 1]
DOI:10.1016/j.gdata.2017.07.013URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 2]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
DOI:10.1126/science.1058709URL [��������: 1]
DOI:10.1073/pnas.1203849109URL [��������: 1]
DOI:10.1111/1574-6976.12028URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
.
[��������: 1]
URL [��������: 1]
.
URL [��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
[��������: 1]
.
[��������: 1]
URL [��������: 1]
.
URL [��������: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}