 ,1,*
,1,*Individual and combined effects of air temperature at filling stage and nitrogen application on storage protein accumulation and its different components in rice grains
HAN Zhan-Yu1, GUAN Xian-Yue1, ZHAO Qian1, WU Chun-Yan2, HUANG Fu-Deng2, PAN Gang1, CHENG Fang-Min,1,*通讯作者:
收稿日期:2019-11-28接受日期:2020-03-24网络出版日期:2020-07-12
| 基金资助: |
Received:2019-11-28Accepted:2020-03-24Online:2020-07-12
| Fund supported: |
作者简介 About authors
E-mail:21716125@zju.edu.cn。

摘要
关键词:
Abstract
Keywords:
PDF (3708KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
韩展誉, 管弦悦, 赵倩, 吴春艳, 黄福灯, 潘刚, 程方民. 灌浆温度和氮肥及其互作效应对稻米贮藏蛋白组分的影响[J]. 作物学报, 2020, 46(7): 1087-1098. doi:10.3724/SP.J.1006.2020.92062
HAN Zhan-Yu, GUAN Xian-Yue, ZHAO Qian, WU Chun-Yan, HUANG Fu-Deng, PAN Gang, CHENG Fang-Min.
稻米品质是品种基因型与环境生态因素共同作用的结果, 灌浆结实期温度与氮肥施用量是影响稻米品质的两个重要生态因子[1,2]。现已基本明确, 灌浆结实期高温会引起稻米整精米率下降、垩白度增加和稻米蒸煮食味品质变劣[2,3,4], 水稻生育后期氮素营养不足或氮肥施用过量均不利于稻米品质改良[5,6,7,8,9]。其中, 氮素营养不足会引起水稻生育后期功能叶片的提早衰老和光合同化物生产能力迅速下降[10], 导致籽粒灌浆充实不良、稻米垩白度增加、整精米率下降, 但氮素穗肥施用过大, 则会引起稻米蛋白含量偏高、米饭色泽和透明度变差、食味口感下降[11,12]。
贮藏蛋白是稻米的第二大类化学组分, 它不仅是评价稻米营养品质的一个重要指标, 而且与稻米外观、蒸煮食味品质也存在密切联系[13]。一般认为, 蛋白质含量过高会引起稻米食味品质变差[1,7-9,11], 但陈能等[14]认为, 两者的负相关性并不显著。据Champagne等[5]和姚姝等[12]报道, 尽管蛋白质含量对食味有一定程度的影响, 但品种类型不同, 其影响程度各异。稻米蛋白质对其蒸煮食味品质的负面效应, 只有在蛋白质含量超过一定的上限指标(6%~ 7%)后才会体现[14,15]。此外, 稻米蛋白含量对米饭色泽和食味口感的影响, 也与其贮藏蛋白组分差异及其亚基构成等因素有关。据Hamaker等[16,17]报道, 稻米蛋白质中的谷蛋白和球蛋白是由一些优良氨基酸组成, 属营养丰富且不影响米饭食味的蛋白质, 只有阻碍淀粉网眼状结构发展的醇溶蛋白才是导致米饭食味降低的贮藏蛋白组分。但Lin等[18]认为, 稻米谷蛋白对淀粉的糊化和膨胀具有抑制作用, 其含量高低与稻米糊化温度、胶稠度等品质指标呈显著负相关, 占稻米贮藏蛋白含量80%左右的谷蛋白含量增加会降低稻米蒸煮食味品质。近年来的水稻优质栽培实践表明, 灌浆结实期高温和氮肥施用过量均会引起稻米蛋白质含量增加和米饭食味下降[6,7,8,9,10,11,12], 但对于不同生态条件下稻米蛋白质含量与其蒸煮食味品质间的相互关系, 尤其是灌浆结实期高温对稻米贮藏蛋白组分的影响及其有别于氮肥效应的理化生态特点, 迄今尚缺乏较明确认识。为此, 本研究以多个水稻主栽品种为材料, 通过人工控温试验、大田氮肥试验、温氮两因素互作试验, 比较分析水稻贮藏蛋白含量及其组分的差异变化, 旨在揭示灌浆结实期高温与氮肥施用量对稻米贮藏蛋白组分的影响及其交互效应, 为稻米营养品质改良以及通过品种选育和栽培调控等措施来协调稻米营养品质与蒸煮食味品质间的关系提供理论依据。
1 材料与方法
1.1 温度处理试验
温度处理试验于2017年在浙江大学紫金港校区农业试验站进行。供试水稻品种为浙恢7954、黄华占、秀水134、秀水09、9311和甬优2640。其中, 浙恢7954、黄华占、9311为籼稻品种, 秀水134和秀水09为粳稻品种, 甬优2640是籼粳亚种间杂交稻品种。5月20日播种, 采用湿润育秧, 6月15日移栽, 各品种在田间随机排列, 每个品种在田间种植2个小区, 每小区面积9 m2, 栽插密度为18 cm × 20 cm, 单本栽插, 待各品种分别在田间生长至孕穗后期, 选取发育进程与长势基本一致的稻株10株, 带泥移入盆钵培育7~10 d至抽穗开花期, 选同日开花且发育良好的单穗挂牌, 并在花后第4天将稻株移入人工气候箱进行灌浆结实期的不同温度处理(每个品种在每个气候箱中各放3~4盆), 直至水稻成熟收获。使用2台CONVIRON人工气候箱, 设常温(日均温度23℃)和高温度(日均温度32℃) 2个处理, 由程序自动控温, 模拟自然气候日变化, 24 h连续运转。每日14:00为最高温、5:00为最低温, 期间温度以1℃为间隔呈线性增加。其中, 常温处理的最高温度和最低温度分别为26℃和20℃, 高温处理时的最高温度和最低温度分别为36℃和28℃, 2台气候箱中的其他气候因子均保持完全一致。其中, 光照时间为6:00—19:00, 光照强度为120~150 J m-2 s-1, 相对湿度为75%~80%, 风速0.5 m s-1。
1.2 氮素(肥)处理试验
试验于2018年在浙江省农业科学院海宁试验基地进行。供试水稻品种为浙恢7954、黄华占、秀水134、秀水09。设不施氮肥(N0)、180 kg hm-2(MN)和300 kg hm-2(HN) 3个氮处理水平, 氮肥为主区, 品种为裂区, 小区面积为6.0 m × 4.0 m, 设3次重复, 共12个小区。氮肥依据含氮率折合成尿素按基肥﹕分蘖肥﹕穗肥为4:3:3施用, 并依此比例来确定各处理不同生育时期的氮素施用量。试验地前茬为油菜, 土壤含有机质2.41%、全氮0.15%、碱解氮104.55 mg kg-1、全磷0.14%、速效磷107.18 mg kg-1、全钾1.38%、速效钾110.26 mg kg-1。5月25日播种, 6月20日移栽, 栽插密度为20 cm × 20 cm, 每穴2苗, 移栽前施过磷酸钙(含P2O5 13.5%) 250 kg hm-2和氯化钾(含K2O 52%) 200 kg hm-2, 不同氮处理的磷肥与钾肥施用量相等。大田常规水作栽培管理, 不同施氮处理间做埂隔开, 用塑料薄膜覆盖埂体, 保证各处理单独排灌, 成熟后收获籽粒用于稻米品质指标测定。1.3 温度与氮肥(素)两因素处理试验
试验于2018年在浙江大学紫金港校区农业试验站进行, 供水稻材料为浙恢7954。5月15日播种, 大田湿润育秧, 6月12日移栽至盆钵, 盆钵直径25 cm、高30 cm, 内装过筛土约15 kg。盆钵土壤2017年取自浙江省农业科学院海宁试验基地, 土壤含有机质2.34%、全氮0.13%、速效氮96.35 mg kg-1、全磷0.17%、速效磷120.41 mg kg-1、全钾1.45%、速效钾110.26 mg kg-1。土壤经自然风干、充分浸泡和充分混匀后, 分别装入不同盆钵(共48盆)。移栽前每盆施2 g尿素和0.5 g KH2PO4做基肥, 移栽后7 d每盆施0.5 g尿素。在水稻生长至穗分化期(叶龄余数为1.5~1.2时)实施氮肥处理。其中, 低氮处理水平(LN)每盆施0.5 g尿素, 高氮处理水平(HN)每盆施2.0 g尿素。每个氮处理水平24盆, 盆钵水稻放置于大型智能温室(28℃白天/22℃夜间)生长, 温室内自然光照, 每3 d左右浇1次水, 及时防控病虫。待盆钵水稻在大型智能温室生长发育至齐穗期, 选同日开花且发育良好的单穗挂牌, 并将每个氮肥处理的盆钵进一步分成3组(每组8盆), 在挂牌标记稻穗开花后的第4天, 分别将其盆钵移入3台CONVIRON人工气候箱实施不同温度处理, 待水稻在气候箱中生长至成熟时, 收获挂牌标记的稻穗。3台CONVIR ON人工气候箱的温度处理设计为常温(日均温度23℃, 日最高温度和最低温度分别为26℃和20℃)、高温I (日均温度30℃, 日最高温度和最低温度分别为34℃和26℃)、高温II (日均温度34℃, 日最高温度和最低温度分别为38℃和30℃)。温度日变化模拟自然气候特征, 每日14:00为最高温、5:00为最低温, 期间温度以1℃为间隔呈线性增加。3台气候箱中的其他气候因子均保持完全一致。其中, 光照时间为6:00—19:00, 光照强度为120~150 J m-2 s-1, 相对湿度为75%~80%, 风速0.5 m s-1。1.4 籽粒总蛋白含量测定
将成熟稻谷放在通风处2个月后, 去壳、磨粉和过筛(0.25 mm)。参照Luthe等[18]报道的方法略作修改, 采用凯氏定氮法测定籽粒糙米米粉总蛋白含量。称取糙米粉 200 mg, 倒入硝煮管, 加3 g硫酸钾:硫酸铜粉末(10:1, w/w), 再加8 mL浓硫酸, 在硝煮仪(Tecator Digestor Auto, Foss)上360℃煮150 min, 将冷却后样品注入凯氏定氮仪, 蒸馏和滴定过程自动运行, 记录下空白和样品消耗盐酸的体积, 利用公式计算出总氮含量, 总蛋白含量等于总氮含量×5.95。1.5 4种蛋白组分的提取与测定
参照Luthe等[19]和Liu等[20]方法对0.5 g糙米米粉中的4种蛋白组分进行分步提取。1)去离子水提取清蛋白; 2) 2%氯化钠提取球蛋白; 3) 55% (v/v)正丙醇提取醇溶蛋白; 4) 0.24%五水合硫酸铜、1.68% KOH、0.5%酒石酸钾钠和50% (v/v)异丙醇混合溶液提取谷蛋白。每次提取液用量为25 mL, 室温下2 h, 期间不断振荡, 其后4000×g离心10 min, 重复提取3次。采用Bradford法[19]测定清蛋白、球蛋白和醇溶蛋白含量, 以牛血清蛋白为对照; 采用双缩脲法测定谷蛋白含量[20], 仍以牛血清蛋白为对照, 并用凯式法加以标定。1.6 蛋白亚基组成的SDS-PAGE分析
参照Yamagata等[21]方法进行稻米蛋白的SDS-PAGE分析。准确称取30 mg糙米粉于2 mL离心管中, 加入1 mL的SDS-urea提取液(4%SDS、5%硫基乙醇、20%甘油和8 mol L-1尿素, pH 6.8), 室温下过夜, 将混合液在20℃ 15,000×g离心20 min, 上清液用于点样。取10 μL总蛋白提取液上样进行SDS-PAGE电泳。采用分离胶浓度为12%, 浓缩胶浓度均为5%。电极缓冲液为Tris-HCl体系, 含0.1% SDS。室温下恒流20~30 mA (起始15~20 mA)电泳2~3 h至溴酚蓝泳动到胶板下缘停止, 取出凝胶板进行冲洗、固定、染色、脱色等。最后将凝胶在Bio-Rad GS-800扫描仪扫描, 以ImageJ对染色凝胶中谷蛋白及其前驱物的相对光密度进行估算。
2 结果与分析
2.1 灌浆温度对水稻籽粒蛋白含量和组分的影响
由表1可见, 高温处理会引起水稻籽粒总蛋白含量(%)的显著上升, 在4种蛋白组分中, 清蛋白、球蛋白和谷蛋白组分含量(%)在高温处理下呈上升趋势, 而多数品种的醇溶蛋白组分含量(%)在高温处理下呈下降趋势。其中, 9311在高温处理下的醇溶蛋白含量(%)虽略有上升, 但与其常温处理间的差异未达统计显著水平。从单位籽粒蛋白绝对含量(mg grain-1)来看, 浙恢7954和秀水134在高温处理下有所上升(mg grain-1), 但黄华占、秀水09、9311和甬优2640在不同温度处理下的含量(mg grain-1)变化不明显, 甚至略有降低。此外, 供试6个品种的谷蛋白/醇溶蛋白比值在高温处理下均呈不同程度的上升趋势(表1)。上述现象说明, 高温处理虽然会引起水稻籽粒总蛋白和谷蛋白组分的相对含量(%)有所提高, 并显著提高水稻籽粒中的谷蛋白与醇溶蛋白比值, 但单位籽粒中的谷蛋白积累量(mg grain-1)并不一定会出现显著增加, 且醇溶蛋白占籽粒的百分相对含量(%)和单位籽粒的醇溶蛋白绝对量(mg grain-1)均有所下降。Table 1
表1
表1不同温度处理下水稻籽粒蛋白含量及其组分差异
Table 1
| 品种 Cultivar | 处理 Treatment | 相对含量Relative content (%) | 绝对含量Accumulation amount (mg grain-1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 清蛋白 Albumin | 球蛋白 Globulin | 醇溶蛋白 Prolamin | 谷蛋白 Glutelin | 总蛋白 Total protein | 清蛋白 Albumin | 球蛋白 Globulin | 醇溶蛋白 Prolamin | 谷蛋白 Glutelin | 谷/醇比 Glu/Prol | |||
| 浙恢7954 Zhehui 7954 | NT | 0.72 b | 0.80 b | 0.61 a | 6.92 b | 9.17 b | 0.171 b | 0.190 a | 0.145 a | 1.644 b | 11.34 b | |
| HT | 1.06 a | 0.95 a | 0.57 b | 7.83 a | 10.29 a | 0.221 a | 0.199 a | 0.122 b | 1.678 a | 13.74 a | ||
| 黄华占 Huanghuazhan | NT | 0.95 b | 0.93 b | 0.72 a | 7.93 b | 10.46 b | 0.214 b | 0.209 b | 0.162 a | 1.785 a | 11.01 a | |
| HT | 1.15 a | 1.02 a | 0.65 b | 8.21 a | 10.83 a | 0.253 a | 0.225 a | 0.143 b | 1.810 a | 12.63 a | ||
| 秀水134 Xiushui 134 | NT | 1.06 a | 0.96 a | 0.56 a | 6.53 b | 8.97 b | 0.233 a | 0.211 a | 0.123 a | 1.442 b | 11.71 b | |
| HT | 0.92 b | 0.97 a | 0.52 b | 7.26 a | 9.61 a | 0.186 b | 0.196 b | 0.103 b | 1.466 a | 14.23 a | ||
| 秀水09 Xiushui 09 | NT | 1.01 a | 1.07 b | 0.64 a | 6.47 b | 9.38 b | 0.224 a | 0.237 a | 0.142 a | 1.433 a | 10.11 b | |
| HT | 0.98 a | 1.12 a | 0.57 b | 7.24 a | 10.15 a | 0.195 b | 0.221 b | 0.113 b | 1.439 a | 12.70 a | ||
| 9311 | NT | 0.89 a | 0.97 a | 0.57 a | 7.32 b | 10.67 b | 0.209 b | 0.228 a | 0.130 a | 1.724 a | 12.84 b | |
| HT | 0.88 a | 1.02 a | 0.58 a | 8.01 a | 11.02 a | 0.224 a | 0.216 b | 0.120 a | 1.695 a | 13.81 a | ||
| 甬优2640 Yongyou 2640 | NT | 0.92 b | 1.09 a | 0.65 a | 7.13 b | 9.96 b | 0.212 a | 0.251 a | 0.150 a | 1.642 a | 10.97 b | |
| HT | 0.99 a | 1.18 a | 0.54 b | 7.46 a | 10.35 a | 0.216 a | 0.258 a | 0.123 b | 1.632 a | 13.81 a | ||
新窗口打开|下载CSV
利用SDS-PAGE对不同温度处理下籽粒贮藏蛋白亚基差异的检测表明(图1), 水稻籽粒贮藏蛋白主要由57 kD、37~39 kD、22~23 kD和醇溶蛋白亚基(prolamin)组成。其中, 57 kD、37 kD和22 kD是3个谷蛋白亚基, 分别对应其pro-glutelin、α-glutelin和β-glutelin。同一品种的不同温度处理间相比, 57 kD谷蛋白前体亚基(pro-glutelin)在高温处理下的条带亮度相对较高, 而醇溶蛋白亚基在高温处理下的条带亮度明显变暗。光密度扫描结果表明, 浙恢7954在高温处理下的57 kD谷蛋白前体亚基(pro-glutelin)、37 kD谷蛋白酸性亚基(α-glutelin)和22 kD谷蛋白碱性亚基(β-glutelin)含量均高于其常温对照, 而黄华占和秀水134在不同温度处理下的α-glutelin亚基含量差异不明显。上述现象说明, 高温处理在引起水稻籽粒醇溶蛋白积累量降低的同时, 会引起谷蛋白前体亚基(pro-glutelin)量的较明显增加。
图1
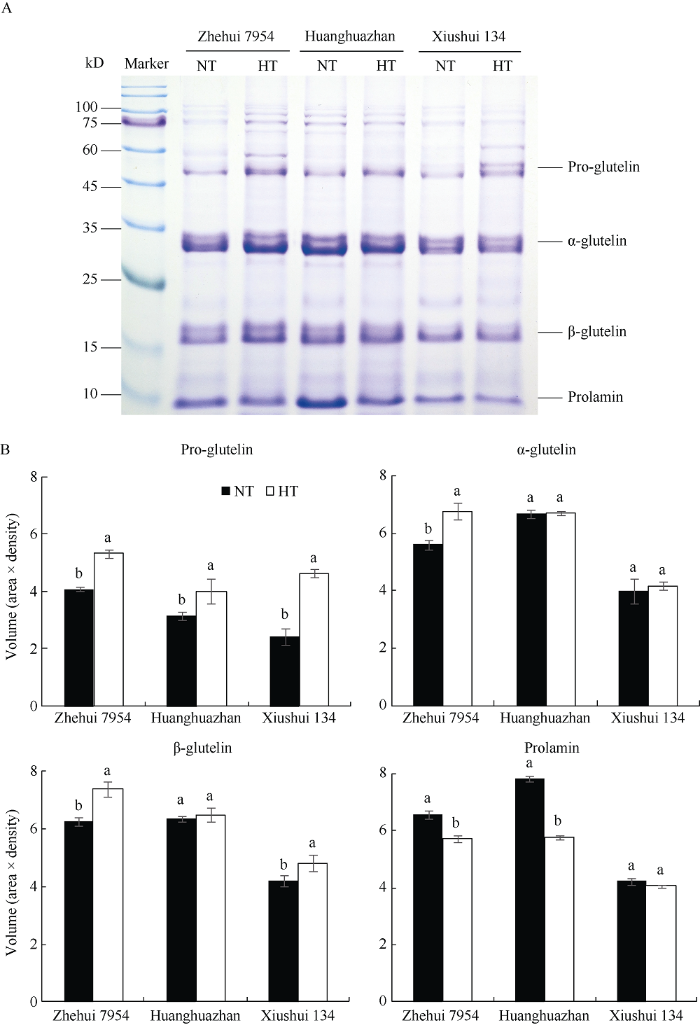 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同温度处理下水稻籽粒贮藏蛋白SDS-PAGE及其主要蛋白条带的相对光密度差异
NT和HT分别表示灌浆结实期的常温处理和高温处理; A和B分别表示水稻籽粒贮藏蛋白SDS-PAGE图和主要蛋白条带的相对光密度图; 同一品种中的2个温度处理间相比, 小写字母相同者表示2个温度处理间(NT和HT)的差异未达到统计显著水平(P < 0.05); 垂直棒代
Fig. 1Differences in SDS-PAGE and relative optical density of storage protein between two temperature treatments
NT and HT mean normal temperature regime and high temperature regime during grain filling, respectively. A and B mean SDS-PAGE and relative optical density of storage protein, respectively. For a same rice cultivar, bars indexed with the same small letters are not significantly different (P < 0.05) between NT and HT. Error bars represent SD values of three biological replicates.
2.2 氮肥处理对水稻籽粒蛋白含量和组分的影响
表2显示, 增施氮肥引起水稻籽粒总蛋白及其4种蛋白组分含量(%)的显著上升, 且高氮处理(HN)下籽粒总蛋白、清蛋白、球蛋白、醇溶蛋白和谷蛋白含量(%)通常高于中氮处理(MN), 但部分品种的总蛋白、谷蛋白和醇溶蛋白在高氮处理(HN)与中氮处理(MN)间差异未到达统计显著水平。氮素穗肥处理对水稻单位籽粒蛋白绝对含量(mg grain-1)的影响与其百分相对含量(%)相似, 总体趋势表现为HN > MN >LN, 但HN与MN两个处理之间的差异幅度相对较小。其中, 部分品种在HN和MN两个处理间的单位籽粒醇溶蛋白和谷蛋白含量(mg grain-1)差异未达统计显著水平(表2)。此外, 与不施氮对照(LN) 相比, 增施氮肥(HN和MN)引起稻米醇溶蛋白含量(包括百分相对含量与单位籽粒绝对含量)的显著提高, 这与醇溶蛋白含量受高温处理影响所表现出的下降趋势有明显差别。与高温处理相似, 增施氮肥引起水稻籽粒中的谷蛋白/醇溶蛋白比值上升(表2)。但从变化幅度看, 同一品种的谷蛋白/醇溶蛋白比值在不同氮素处理下的差异范围大致在0.46~0.95之间(表2), 远低于其在不同温度处理(HT与NT)间的差异幅度(表1)。这说明氮素穗肥对水稻籽粒谷蛋白/醇溶蛋白比值的影响程度相对小于高温处理。Table 2
表2
表2不同施氮水平下水稻籽粒蛋白含量及其组分差异
Table 2
| 品种 Cultivar | 处理 Treatment | 相对含量Relative content (%) | 绝对含量Accumulation amount (mg grain-1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 清蛋白Albumin | 球蛋白Globulin | 醇溶 蛋白Prolamin | 谷蛋白Glutelin | 总蛋白Total protein | 清蛋白Albumin | 球蛋白Globulin | 醇溶蛋白Prolamin | 谷蛋白Glutelin | 谷/醇比Glu/Prol | |||
| 浙恢7954 Zhehui7954 | LN | 0.87 b | 0.83 b | 0.53 b | 6.71 c | 8.87 c | 0.197 b | 0.188 c | 0.120 b | 1.524 c | 12.66 b | |
| MN | 0.89 b | 1.06 a | 0.58 a | 7.39 b | 10.57 a | 0.206 ab | 0.246 a | 0.137 a | 1.714 b | 12.53 b | ||
| HN | 1.03 a | 0.97 b | 0.61 a | 8.06 a | 10.81 a | 0.228 a | 0.215 b | 0.135 a | 1.786 a | 13.21 a | ||
| 黄华占 Huanghuazhan | LN | 0.86 b | 0.92 b | 0.66 b | 7.47 b | 9.43 b | 0.187 c | 0.200 b | 0.144 b | 1.626 b | 11.57 b | |
| MN | 1.02 a | 1.11 a | 0.68 ab | 8.08 a | 11.06 a | 0.226 a | 0.245 a | 0.151 a | 1.790 a | 11.93 a | ||
| HN | 0.98 a | 1.06 a | 0.71 a | 8.12 a | 11.18 a | 0.211 b | 0.228 ab | 0.153 a | 1.747 a | 11.66 b | ||
| 秀水 134 Xiushui 134 | LN | 0.87 b | 0.91 b | 0.56 b | 5.75 c | 8.57 b | 0.184 b | 0.192 b | 0.110 b | 1.320 b | 11.16 b | |
| MN | 0.91 b | 1.07 a | 0.63 a | 6.91 b | 10.24 a | 0.189 b | 0.222 a | 0.131 a | 1.538 a | 11.76 a | ||
| HN | 1.12 a | 1.08 a | 0.66 a | 7.17 a | 10.53 a | 0.223 a | 0.215 ab | 0.132 a | 1.530 a | 11.62 a | ||
| 秀水09 Xiushui 09 | LN | 0.92 b | 0.93 c | 0.51 c | 6.32 c | 9.09 c | 0.204 b | 0.206 b | 0.113 b | 1.384 c | 12.21 b | |
| MN | 0.90 b | 1.07 b | 0.57 b | 7.04 b | 9.85 b | 0.201 b | 0.239 a | 0.127 a | 1.663 b | 13.05 a | ||
| HN | 1.06 a | 1.14 a | 0.64 a | 7.76 a | 10.98 a | 0.223 a | 0.240 a | 0.130 a | 1.716 a | 13.16 a | ||
新窗口打开|下载CSV
利用SDS-PAGE对不同氮处理水平下籽粒贮藏蛋白亚基组成的检测表明(图2), 低氮处理下几个主要蛋白亚基条带(包括pro-glutelin、α-glutelin、β-glutelin和prolamin)的相对亮度均弱于其同一品种的高氮(HN)和中氮(MN)处理(图2-A)。光密度扫描结果表明, 施氮处理(HT和MN)对谷蛋白亚基组成的影响不明显, 谷蛋白的57 kD前体亚基(pro- glutelin)、37 kD酸性亚基(α-glutelin)和22 kD碱性亚基(β-glutelin)在高氮(HN)和中氮(MN)的含量均显著高于其低氮处理(LN)。与此同时, 醇溶蛋白亚基含量在高氮处理(HN)下也呈上升趋势。说明高氮处理虽然会引起水稻籽粒谷蛋白和醇溶蛋白含量的上升, 但对谷蛋白亚基的积累形态影响并不明显。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同施氮水平下水稻籽粒贮藏蛋白SDS-PAGE及其主要蛋白条带的相对光密度差异
LN、MN和HN分别表示低氮(0 kg hm-2)、中氮(180 kg hm-2)和高氮(300 kg hm-2)处理水平; A和B分别表示水稻籽粒贮藏蛋白SDS-PAGE图和主要蛋白条带的相对光密度图; 相同水稻品种, 小写字母相同者表示不同氮处理水平间的差异未达到统计显著水平(P < 0.05); 垂直棒代
Fig. 2Differences in SDS-PAGE and relative optical density of storage protein among three N treatments
LN, MN, and HN mean low nitrogen (0 kg hm-2), medium nitrogen (180 kg hm-2), and high nitrogen (300 kg hm-2) levels, respectively. A and B mean SDS-PAGE and relative optical density of storage protein, respectively. Bars indexed with the same small letters are not significantly different (P < 0.05) between different N levels for the same rice cultivar; Error bars represented SD values of three biological replicates.
2.3 温氮两因素处理对水稻籽粒蛋白含量和组分的影响
对不同温氮处理下水稻籽粒4种蛋白组分和总蛋白含量的比较可知(表3), 高氮(HN)+高温(HT2)处理组合下的籽粒谷蛋白和总蛋白含量(%)明显高于高氮(HN)+常温(NT)和低氮(LN)+高温(HT2)等处理组合, 但高温(HT2)+高氮(HN)处理组合下的单位籽粒谷蛋白的绝对含量(mg grain-1)却低于高氮(HN)+常温(NT)处理组合, 这说明高氮+高温处理(HN+HT2)并不会引起谷蛋白绝对含量(mg grain-1)的进一步提升。此外, 在低氮(LN)和高氮(HN)水平下, 高温处理均会引起水稻籽粒醇溶蛋白含量(相对百分含量和单位籽粒绝对含量)的较明显下降和谷蛋白/醇溶蛋白的比值上升, 且其绝对含量(mg grain-1)在HT2处理下的下降或上升幅度大于HT1处理。同一温度处理下的不同氮水平相比, 籽粒总蛋白、清蛋白、球蛋白、醇溶蛋白和谷蛋白在高氮水平(HN)下的相对含量(%)和绝对含量(mg grain-1)均显著高于其相应的低氮(LN)处理(表3)。Table 3
表3
表3温氮处理下水稻籽粒蛋白及其组分的含量差异
Table 3
| 氮水平 N level | 温度处理 Temperature treatment | 相对含量Relative content (%) | 绝对含量Accumulation amount (mg grain-1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 清蛋白 Albumin | 球蛋白 Globulin | 醇溶蛋白 Prolamin | 谷蛋白 Glutelin | 总蛋白 Total protein | 清蛋白 Albumin | 球蛋白 Globulin | 醇溶蛋白 Prolamin | 谷蛋白 Glutelin | 谷/醇比 Glu/prol | |||
| LN | NT | 0.81 b | 0.84 b | 0.53 a | 6.45 b | 8.52 c | 0.19 a | 0.19 a | 0.12 a | 1.49 a | 12.17 c | |
| HT1 | 0.87 b | 0.95 a | 0.51 a | 7.03 a | 9.29 b | 0.19 a | 0.21 a | 0.11 a | 1.52 a | 13.48 b | ||
| HT2 | 1.09 a | 1.03 b | 0.46 b | 6.98 a | 9.41 a | 0.20 a | 0.20 a | 0.09 b | 1.38 b | 15.17 a | ||
| 平均 Mean | 0.92 B | 0.94 B | 0.50 B | 6.80 B | 9.04 B | 0.19 B | 0.20 B | 0.11 B | 1.47 B | 13.63 A | ||
| HN | NT | 0.93 b | 1.02 b | 0.61 a | 7.15 b | 9.77 b | 0.21 a | 0.23 ab | 0.14 a | 1.64 a | 11.72 b | |
| HT1 | 0.91 a | 0.97 a | 0.56 b | 7.23 b | 10.24 a | 0.21 a | 0.22 b | 0.13 ab | 1.66 a | 13.15 a | ||
| HT2 | 1.07 a | 1.18 a | 0.58 ab | 7.84 a | 10.59 a | 0.22 a | 0.24 a | 0.12 b | 1.58 b | 13.75 a | ||
| 平均 Mean | 0.97 A | 1.06 A | 0.58 A | 7.41 A | 10.20 A | 0.21 A | 0.23 A | 0.13 A | 1.63 A | 12.84 B | ||
| 氮肥效应 Nitrogen | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ||
| 温度效应 Temperature | ** | ** | ** | ** | ** | ns | * | ** | ** | ** | ||
| 氮肥×温度 Nitrogen × temperature | ** | ** | ** | ** | ** | ns | ns | ns | ** | ** | ||
新窗口打开|下载CSV
对不同氮(素)×温(度)处理下籽粒贮藏蛋白亚基组成的SDS-PAGE检测结果(图3)表明, 在低氮水平下, 高温处理(HT1和HT2)会引起籽粒谷蛋白的57 kD前体亚基(pro-glutelin)、37 kD酸性亚基(α- glutelin)和22 kD碱性亚基(β-glutelin)含量显著升高, 但在高氮水平下, 高温处理(HT1和HT2)的37 kD酸性亚基(α-glutelin)和22 kD碱性亚基(β-glutelin)含量却低于常温处理(NT)。这说明灌浆温度对籽粒谷蛋白含量的影响可能与氮营养供应状况有关, 温度与氮肥对稻米谷蛋白及其亚基组分的影响存在一定程度的互作效应。对醇溶蛋白而言, 在低氮水平(LN)和高氮水平(HN)下, 高温处理均会引起其含量的明显降低(图3-B)。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3温氮处理下水稻籽粒贮藏蛋白的SDS-PAGE及其主要蛋白条带的相对光密度差异
LN和HN分别表示高氮和低氮水平; NT、HT1和HT2分别表示常温处理(日均温度23℃)、高温I处理(日均温度30℃)和高温II处理(日均温度34℃)。A和B分别表示水稻籽粒贮藏蛋白SDS-PAGE图和主要蛋白条带的相对光密度图。小写字母相同者表示不同处理水平间的差异未达到统计显著水平(P < 0.05); 垂直棒代
Fig. 3Differences in SDS-PAGE and relative optical density of storage protein treated under different temperature regimes and different N levels
LN and HN mean low nitrogen and high nitrogen levels, respectively; NT, HT1, and HT2 mean normal temperature regime (23℃), HT1 (30℃) and HT2 (34℃), respectively. A and B mean SDS-PAGE and relative optical density of storage protein, respectively. Bars indexed with the same small letters are not significantly different (P < 0.05) among different treatments; Error bars represent SD values of three biological replicates.
3 讨论
关于灌浆结实期温度对稻米蛋白质含量影响, 现已不乏报道, 但研究结论不一[15,18,24-25]。多数研究报道认为, 灌浆结实高温或夜间增温会引起稻米粗蛋白含量提高[15,22-24], 但周广洽等[26]认为, 灌浆结实期温度过高或过低都会引起稻米蛋白质含量下降。据Krishman等[15]报道, 灌浆温度对稻米蛋白质含量的影响因品种而异, 大多数品种蛋白质含量与结实期温度正相关, 但也有部分品种的蛋白质含量与灌浆温度相关不显著或呈负相关。Lin等[18]认为, 稻米蛋白质含量在高温胁迫下增加的原因主要是籽粒粗蛋白占籽粒总干重的相对比值增加, 且高温胁迫会导致籽粒灌浆后期贮藏蛋白积累量减少[27]。本研究中, 稻米谷蛋白和总蛋白的相对含量(%)在高温处理下呈显著上升趋势, 这与前人的大多数研究结果基本一致。但从单位籽粒中积累的蛋白绝对量看, 除浙恢7954和秀水134之外, 其余4个品种单位籽粒中所积累的谷蛋白的绝对含量在高温处理下并没有表现出十分明显的上升趋势(表1)。据前人研究报道, 水稻籽粒淀粉合成积累量在高温胁迫下显著降低、糊粉层占淀粉层的相对比例明显提高[15,22], 高温胁迫对灌浆籽粒中糖转运和淀粉合成代谢相关基因转录表达水平的影响幅度比贮藏蛋白合成代谢相关基因明显[28]。Yamakawa等[29]研究表明, 水稻胚乳中的谷蛋白、醇溶蛋白和球蛋白类基因在高温处理下多呈下调表达特征。由此笔者推测, 灌浆结实期高温引起稻米蛋白含量(%)上升的原因, 在很大程度上与高温胁迫下的淀粉积累量下降和贮藏蛋白占籽粒干重的相对比例增加所引起的“浓缩效应”有关, 籽粒干重和淀粉积累量的下降, 是高温处理下水稻籽粒贮藏蛋白的相对含量(%)增加的一个重要因素。此外, 高温胁迫处理不仅影响水稻籽粒的贮藏蛋白总量, 而且影响贮藏蛋白的组成及其积累形态。本文人工气候箱控温试验表明, 不同水稻品种籽粒中的醇溶蛋白含量在高温处理下均一定程度下降, 这与马启林等[25]的研究结果基本一致。本文进一步揭示, 高温处理所引起水稻籽粒醇溶蛋白含量的下降现象, 不仅表现在醇溶蛋白占籽粒干重的相对含量(%)上, 而且也表现在单位籽粒中的醇溶蛋白的绝对含量(mg grain-1)上(表1)。高温处理在引起水稻籽粒醇溶蛋白及其13 kD亚基组分含量下降的同时, 籽粒谷蛋白的57 kD前体亚基(pro-glutelin)组分含量有所增加, 而37 kD酸性亚基(α-glutelin)和22 kD碱性亚基(β-glutelin)含量在不同温度处理下的差异变化因品种而异(图1)。与之相比, Dou等[23]的大田增温试验结果表明, 夜间增温会引起稻米粗蛋白含量增加, 但对稻米各种氨基酸组分的相对比例影响不大。因此, 灌浆温度对稻米谷蛋白含量的影响相对较复杂, 可能既与水稻品种本身的谷蛋白含量高低及其各个亚基组分的比例差异等遗传因素有关, 也与高温范围或增温幅度等生态因素存在一定联系。氮肥水平是影响稻米粗蛋白含量的重要栽培因素之一。大量研究表明, 增施氮肥或前氮后移会引起稻米贮藏蛋白含量的较明显增加[6,7,8,9,10]。在一定施氮量范围内, 稻米蛋白含量随氮肥(尤其是氮素穗肥)施用量的增加呈上升趋势, 但不同水稻品种的稻米蛋白含量对氮肥的敏感程度不同, 氮肥施用量过高会引起部分水稻品种籽粒饱满度下降[8,10-11]。在本文研究结果中, 增施氮肥使水稻籽粒总蛋白及其4种蛋白组分含量(%)显著上升, 这与前人研究结果一致。本研究利用SDS-PAGE对不同氮处理水平下籽粒贮藏蛋白亚基组成的检测表明, 谷蛋白的57 kD前体亚基(pro-glutelin)、37 kD酸性亚基(α-glutelin)和22 kD碱性亚基(β-glutelin), 以及醇溶蛋白的13 kD亚基在高氮处理下含量均明显高于其相同品种的低氮处理(LN)(图2), 且增施氮肥引起水稻籽粒谷蛋白/醇溶蛋白的比值有所上升(表1), 但与灌浆结实期温度处理相比, 氮肥处理对谷蛋白各个亚基的积累形态及其相对比例影响不甚明显, 且同一品种在不同氮肥处理的谷蛋白/醇溶蛋白比值的变化幅度也没有高温处理对两者比值的影响程度大。值得关注的是, 增施氮肥(HN和MN)会引起稻米醇溶蛋白含量的显著提高, 这与醇溶蛋白受高温处理影响所表现出的下降趋势有明显不同, 这也是高温处理有别于高氮处理对稻米贮藏蛋白组分影响的一个重要特点。据前人研究报道, 醇溶蛋白存在于蛋白体I (PB-I), 球蛋白和谷蛋白储存于蛋白体II (PB-II)中[13,21,30]。其中, 富含谷蛋白的PB-II蛋白体易于消化, 对食味负面效应小, 而以醇溶蛋白为主的PB-I对蛋白分解酶具有较强的抵抗力, 可能是稻米蛋白质影响米饭食味的一个重要因素[30,31]。但在本研究中, 增施氮肥会引起稻米中的醇溶蛋白含量上升, 而稻米醇溶蛋白含量在高温处理下呈下降趋势, 且同一品种在高氮处理(HN)下的谷蛋白/醇溶蛋白比值还略高于其低氮处理(LN)和中氮处理(MN)(表1和表2)。因此, 尽管稻米蛋白含量及其组分变化会对稻米蒸煮食味品质产生明显影响, 但稻米食味品质与贮藏蛋白含量及组分间的关系可能较复杂, 尤其是灌浆结实期高温造成的稻米食味品质下降现象, 实际上并不是高温胁迫下籽粒醇溶蛋白含量升高所引起的。
随全球气候的变暖, 水稻高温危害及其栽培措施缓解问题正日益引起人们关注[32]。据前人研究报道, 合理氮素穗粒肥可减轻或缓解高温对水稻灌浆结实的胁迫伤害[33,34,35]。段骅等[36]认为, 在高温胁迫下适当增施氮肥可提高水稻的结实率和千粒重, 减轻高温对产量和稻米品质的不利影响。吴翠平等[37]研究表明, 在基追比例1:1的基础上增大拔节期氮素追施比例, 能显著提高小麦常温和高温胁迫下籽粒产量, 缓解高温胁迫对小麦千粒重和籽粒产量的不良影响, 但对小麦品质影响不明显。Ito等[38]研究认为, 高温胁迫会引起小麦籽粒中的淀粉合成受阻, 而适度施氮能通过维持淀粉合成相关酶活性来减轻高温危害。从而降低高温下稻米垩白发生、提高稻米食味和整精米率。Tang等[35]研究表明, 氮素粒肥可通过影响稻米淀粉组分及其有关理化特性来缓解高温胁迫对稻米食味不利效应, 但对于灌浆温度与氮素穗肥施用对稻米蛋白含量及其组分的影响效应目前尚不清楚。在本研究中, 高氮(HN)+高温(HT2)处理组合下的稻米谷蛋白和总蛋白含量明显高于高氮(HN)+常温(NT)和低氮(LN)+高温(HT2)等处理组合, 这说明高温和高氮对稻米谷蛋白和总蛋白含量的影响存在着一定程度的“累加”效应, 但在高氮处理(HN)下高温引起稻米醇溶蛋白含量的降低幅度相对小于其相应的低氮处理(LN), 有利于稻米醇溶蛋白含量在不同温度处理下的相对稳定(表3和图3)。此外, 本文温氮两因素试验结果表明, 在低氮水平下, 高温会引起籽粒谷蛋白的57 kD前体亚基、37 kD酸性亚基和22 kD碱性亚基含量显著升高, 而在高氮水平下, 谷蛋白的上述亚基组分随温度处理变化却呈相反趋势, 说明灌浆温度对籽粒谷蛋白亚基组分的影响与氮处理水平有关。对于温度与氮肥对稻米谷蛋白及其亚基组分影响的互作效应等问题, 尚需今后进一步深入研究。
4 结论
高温处理和增施氮肥虽然均会引起水稻籽粒总蛋白和谷蛋白等组分占籽粒干重相对含量(%)的显著提升, 但高温处理对单位籽粒中总蛋白与谷蛋白的绝对含量(mg grain-1)影响却远没有增施氮肥的效应明显, 因此灌浆结实期高温所引起的稻米蛋白相对含量(%)上升现象, 在很大程度上与高温胁迫处理下的淀粉积累量下降和贮藏蛋白占籽粒干重的相对比例增加所引起的“浓缩效应”有关。此外, 高温处理与增施氮肥对稻米醇溶蛋白组分的影响存在明显差别, 高温处理会引起稻米醇溶蛋白在相对含量(%)和绝对含量(mg grain-1)上均呈现明显下降, 并显著提高稻米中的谷蛋白/醇溶蛋白比值, 而增施氮肥在引起稻米谷蛋白含量提高的同时, 稻米醇溶蛋白含量也呈现上升趋势, 氮肥处理对谷蛋白/醇溶蛋白比值的影响相对小于温度处理, 说明灌浆结实期高温造成的稻米食味品质下降现象, 并不是其在高温胁迫下籽粒醇溶蛋白含量升高所引起的。高氮+高温处理组合对稻米总蛋白与谷蛋白含量的影响程度要显著大于单一高温或高氮处理, 两因素对稻米总蛋白含量和谷蛋白组分的影响存在一定程度的“累加”效应, 但增施氮肥可缓解稻米醇溶蛋白在高温胁迫下的降低幅度, 有利于水稻在遭遇高温胁迫气候下稻米醇溶蛋白含量的相对稳定。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.tplants.2008.12.004URL [本文引用: 2]
URL [本文引用: 2]

就食用稻米组分、稻米品质研究进展进行了概述。剖析了传统的品质研究思路,分析了食用优质稻米研究与品种改良中存在的问题,即稻米品质性状的静态研究较多,动态形成的研究较少;对直链淀粉、蛋白质含量作了大量的研究,而对淀粉的结构研究较少;淀粉代谢和稻米品质形成过程中关键酶的动态研究较少。为此提出稻米品质形成规律研究中急需开展的研究内容及与其配套的研究新方法和新手段。
URL [本文引用: 2]
就食用稻米组分、稻米品质研究进展进行了概述。剖析了传统的品质研究思路,分析了食用优质稻米研究与品种改良中存在的问题,即稻米品质性状的静态研究较多,动态形成的研究较少;对直链淀粉、蛋白质含量作了大量的研究,而对淀粉的结构研究较少;淀粉代谢和稻米品质形成过程中关键酶的动态研究较少。为此提出稻米品质形成规律研究中急需开展的研究内容及与其配套的研究新方法和新手段。
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 3]
[本文引用: 3]
[本文引用: 4]
[本文引用: 4]
[本文引用: 4]
[本文引用: 4]
[本文引用: 4]
[本文引用: 4]
[本文引用: 4]
[本文引用: 4]
[本文引用: 4]
[本文引用: 4]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 5]
[本文引用: 1]
[本文引用: 1]
DOI:10.1021/jf101575jURLPMID:20839801 [本文引用: 4]
The present study was performed to understand the effects of high temperature (HT) during filling on the expression of storage proteins and the quality of rice grains. HT (35/30 degrees C day/night) reduced the weight, amylose content, and flour gel consistency of grains. It increased the accumulation of all classes of storage proteins at early filling stage but decreased the accumulation of prolamins at maturation. For albumins, the expressions of cyclophilin 2, peroxiredoxin, and HSP16.9 were differentially enhanced by HT. For globulins, HT decreased the accumulation of globulin but increased that of glyoxalase I and peroxiredoxin. HT enhanced the transcription of genes for glutelins, prolamins, globulins, and protein disulfide isomerase at early filling stage but decreased the expression of these genes at a later stage. Low amounts of prolamins and globulins, as well as low pH value, were found in sound, immature, and dead kernels grown under HT. The relationships among HT, storage proteins, and grain quality are discussed.
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:17384160 [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
URLPMID:15226500 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.foodchem.2018.10.137URLPMID:30502170 [本文引用: 2]
Relevant evidence of rice quality responses to increased temperatures under field condition has been obtained in our previous study. Understanding the intrinsic mechanism of increased temperatures and fertilizer treatment in regulating rice quality formation will be contributed to the development of mitigation measures suitable for actual field operations and to cope with climate warming. This study investigated the potential role of nitrogen fertilizer in regulating the rice quality under field warming through identifying starch-related property indicators. Results showed application of nitrogen fertilizer at heading stage effectively delayed the significantly increased accumulation of total starch, amylose and amylopectin, and reduced the starch particle size, chain length distribution, and crystal structure induced by high temperature, which further compensated the deterioration of rice quality caused by elevated temperature.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}