 ,1,*, ������,1,*
,1,*, ������,1,*Development and validation of markers linked to genes resistant to Sitodiplosis mosellana in wheat
HAO Zhi-Ming1, GENG Miao-Miao1, WEN Shu-Min1, YAN Gui-Jun2, WANG Rui-Hui,1,*, LIU Gui-Ru,1,*ͨѶ����:
�ո�����:2019-04-8��������:2019-09-26�����������:2019-10-16
| ��������: |
Received:2019-04-8Accepted:2019-09-26Online:2019-10-16
| Fund supported: |
����� About authors
E-mail:haozhiming0730@hotmail.com��

ժҪ
�ؼ��ʣ�
Abstract
Keywords��
PDF (3375KB)Ԫ������ά�����������������EndNote|Ris|Bibtex�ղر���
�������ø�ʽ
��־��, ������, ������, �ƹ��, ���, ������. С���������������ǵĿ�������֤[J]. ����ѧ��, 2020, 46(2): 179-193. doi:10.3724/SP.J.1006.2020.91029
HAO Zhi-Ming, GENG Miao-Miao, WEN Shu-Min, YAN Gui-Jun, WANG Rui-Hui, LIU Gui-Ru.
���������(Sitodiplosis mosellana G��hin)��������вС����������Ҫ����, ���������׳���˱�ཬ��С�������ں���, ʹ�������ݡ����ؽ���, �����ܲ�ԭ����Ⱦ, ����С������Ʒ���½�, ��Ʒ���ܵ�����Ӱ�졣�ܺ����������ʧ��10%~60%֮��, ����ʱ������������[1,2]���ڼ��ô�������Ӣ�����������¹���С��������, ����������Ϊ���ı���[3,4,5,6,7,8]����20����50�����80���, �ҹ�С��������������������������Ϊ��, ��ʧ��[9,10]��21��������, �������Σ����Ȼ���ء�2013��2018��, С������������溦�����������2.0��106 hm2 (http://www. agri.gov.cn/), ��ΪӰ�컪�����ƻ��������ȵ�С����������Ҫ�溦(https://www.natesc.org.cn/)��������, ��ͨ��ҩ��(ũҩ)��ũ�մ�ʩ(���������)������(�����ռ�����е�)�Ϳ���Ʒ�ַ������������[4,11-12], ���Կ���Ʒ�ֵ�ѡ����������Ϊ������Ч[13]����20����, ��ũ6028�ȿ���Ʒ�ֵ�ʹ�ö����Ƴ溦���˹ؼ�����, ������Ʒ�ֵ�ȱ��Ҳ�ǵ�ǰ�ҹ�����������Ϊ������Ӹ߲��µ���Ҫԭ��[9]����������Σ���������ԡ��������Լ�С��������濹���Ŵ��ĸ�����, ʹ��С�����ֽ�չ����[14,15]����ͳ�Ŀ������ֵ��������ڱ��ͼ������, �����ܻ�������Ϊ���ص�Ӱ��, ���ҷ�ʱ����, ����Ʒ�ֵ�ѡ��Ч��ʮ�ֵ��¡����ӱ�Ǹ���ѡ��(marker-assisted selection, MAS)������С�����Ҫ����������е�Ӧ��, �ܹ���������������ضԿ����������ĸ���, Ҳ������ʵ�����������ũ����״��ͬ������, �����������Ч��[16,17]�������뿹������������/QTL���������ķ��ӱ��, �����Ǹ��ݿ����ѡ�������������ϵ�ͻ��λ�㿪���ɵ�Ŀ�Ļ�����(gene-targeted markers, GTMs)���ܱ��(functional markers, FMs)[18,19,20,21,22], �����ܹ���߿���Ʒ��ɸѡ��������ѡ���Ч��, ����Ҳ�����ڸ���ʵ�ֲ�ͬ��Դ�������ľۺ�, ʹС��Ŀ����Ը��־ú��ȶ�[23]��
Sm1�����緢�ֵ�С��������������[24], λ��2BS��SSR���Xgwm210��Xbarc35��SCAR���XWM1֮��2.5 cM���Ŵ�������[3]��Xbarc35��XWM1���ڱ�����Ӣ����С����MAS�����е���Ӧ��[25]��Kassa��[26]���ú���Sm1�����С���ױ���Ϲ�����DHȺ��, �ַ�����7���ɵ��������̬��(SNP)λ�㿪����KASP����뿹���Խ�������, �����ڼ�����/�г浥���͡�Sm1Ҳ�ǵ�ǰ���ô�Ӣ���������Ŀ���С��Ʒ����Ψһ���㷺���õĿ������[27](http://midgetolerantwheat.ca/; http://www. bcpc.org/; https://www.usda.gov/)������������һ�������, ����������¶��������������ͳ��ֵķ���, Ҳ��ʹ����Ѱ���µĿ��������Դ[28]��
�й�С��Ʒ�ֿ���Я���˲�ͬ����������濹�Ի����ҹ���������С��Ʒ�ֶ�����ϴ�2419����ũ6028�������10�ŵȵ�ѪԵ[9,14,29-32], ����31����ũ151����ũ99��֣��8�š�����23������24����ũ215������66�ȿ���Ʒ�ֵ�ϵ���ж�������Я��������Ʒ�ֵ�ѪԵ[9,14,30-31], ����⿹��С��Ʒ�ֵ�ϵ�ײ�ͬ[3,24,27-28,33-34]�������ȵ�С��Ŀ�������QTL����λ��2B (Sm1)[24]��1A (QSm.mst-1A)[28]Ⱦɫ����, ��Sm1Ϊ��Ҫ��Դ�����Ƿ���, �����ҹ�С��Ʒ�ֵĿ�������Ҫ����λ��4AȾɫ��(QSm.hbau- 4A)[14,35]��Sm1������������(Xgwm210��Xbarc35��XWM1)���ѱ�Ӧ���ڵ���С��Ʒ�ֵ�����, ����Щ������ҹ�С��Ʒ����Ҫô���߶�̬��, Ҫô������뿹���Բ��Ǻϡ���κ��ȶ��������������Ϊ��С����������Ҫ�Ŀ���ɷ�[36,37], ��Sm1����Ŀ����й�[3,26]��Ȼ��, ���Dzⶨ��4A��ЧQTL���Ȼ���ϵ�İ�κ�Ậ��, δ�ܷ������ֲ���Ĵ��ڡ�������-O-��ת��ø(COMT)�ǰ�κ��ϳ�;���еĹؼ�ø[38] (https://www.kegg.jp/)�����Ǵ�������в���µ�С��ת¼�������м�����3��COMTͬԴ����, ���ڿ�����С���ױ����������SNP�������о�����, �ҹ�С��Ʒ�ֿ��ܴ����µ������濹���Ŵ����ơ����ڴ�, �����ҹ�С���������뿹��λ�������������������ķ��ӱ�Ǻ��ܱ��, �����ܸ��õ������ҹ�������MAS���ֵ���Ҫ, Ҳ�����ڶ��ҹ�С��������Դ�����Ե�ȷ������
���Ų������ĸĽ����ɱ��Ľ���, �Լ�������ֲο�����������(�����й���С��ο�����������)�Ĺ���, �Ի������д������б���ͽṹ������ھ��Ϊ����, ��ֱ������ת¼�����ݻ��ϼ�Ⱥ���������(bulked segregant analysis, BSA)��˼·���б�ǿ�����ø��ӱ���[39]����������[40]�Կ�����ػ���TaPP2Aa�������м��ͷ���, ���ݸû����ڲ�ͬС��Ʒ���д��ڵĶ�̬��λ�㿪���˹��ܱ�ǡ�Wu��[41]��ת¼���оƬ�����������ھ�235��Ⱦɫ��������SNPλ��, ��������KASP���, �ɹ�����Yr26����ľ�ϸ��λ��������[42]����BSR-Seq����������SNP���, ����2��SNP��Ƕ�PmYBL��PmSGD������Ч�ʴ�97%��100%��
��ǰ���о���, ���Dz���BSR-Seq[43]������QTL-Seq����[44]��˼·, ��ϻ�����ע����Ϣ��qRT-PCR��������, �ӿ����г�С������(��˫�ס����˻�ء��������ϵ�����Ȼ���ϵ)��ת¼��������, �����λ��4AL��ЧQTL[14,35]�����6����������ز������, ���л���TraesCS4A01G436100��TraesCS4A01G437800������ȫ��������RIL��ϵ��NIL��ϵ��ı���ˮƽ��������, ���������������뿹������ص����ͪ����ϳɴ�лͨ·(isoflavonoid biosynthesis pathway)��[45,46]�����, ���о�������6��������ػ�����SNP�Ͳ���ȱʧλ��(InDel)Ϊ����, ��ƺͿ���Ŀ�Ļ�����, �����ÿ����г�С��RILϵ��С��Ʒ����֤����Щ�����С�������濹�Լ����еĿ����ԡ�
1 �����뷽��
1.1 ֲ����ϼ�DNA��ȡ
���ڱ����֤��ֲ����ϰ���С���ױ�6218 (�г�)������24 (����), ��6218/����24�������ϵ(RIL)Ⱥ����ѡ���92����������������ϵ�Լ����в�ͬ����ˮƽ��95��С��Ʒ�֡�����, RIL��ϵ����С���ױ�6218�뼽��24��F2��Ⱥ��ĵ������Խ����, �����꿹�����, �����ȶ�������С����Ϸֱ���2014��2015��ȡ�2015��2016��Ⱥ�2016��2017�����ֲ�ںӱ�ũҵ��ѧ�������ij��ԡ�ÿ��������ֲ1��, ÿ��40��, �г�20 cm, �о�20 cm[14]����ȫ����������, 3���ظ������Բ��ϵ����Ƽ����ͼ���1������CTAB��[47]��ȡ�����Բ��ϵĻ�����DNA��Table 1
��1
��1����С���ױ���RIL��ϵ�ı���
Table 1
| С���ױ�����ϵ Wheat parent and line | ����ָ��(RI) Resistance index | ����ȼ� Classification | С����ϵ Wheat line | ����ָ��(RI) Resistance index | ����ȼ� Classification |
|---|---|---|---|---|---|
| ����24 Jimai 24 | 0.0128-0.1233 | 1 | RIL-264 | 0.1007-0.2659 | 2 |
| 6218 | 2.9543-7.2714 | 5 | RIL-285 | 0.0064-0.2105 | 2 |
| RIL-7 | 0.0575-0.0622 | 1 | RIL-16 | 0.7300-1.3397 | 4 |
| RIL-12 | 0.0133-0.0336 | 1 | RIL-21 | 0.9122-1.4373 | 4 |
| RIL-18 | 0.0745-0.0817 | 1 | RIL-60 | 1.2009-1.2429 | 4 |
| RIL-23 | 0.0029-0.0330 | 1 | RIL-106 | 1.2142-1.2325 | 4 |
| RIL-38 | 0.1441-0.1886 | 1 | RIL-251 | 0.9603-1.4106 | 4 |
| RIL-64 | 0.0188-0.0380 | 1 | RIL-19 | 2.7085-2.8602 | 5 |
| RIL-73 | 0.0784-0.1409 | 1 | RIL-20 | 2.6327-3.1797 | 5 |
| RIL-91 | 0.0205-0.0818 | 1 | RIL-25 | 3.0783-4.3757 | 5 |
| RIL-115 | 0.0503-0.1358 | 1 | RIL-29 | 2.1227-2.5619 | 5 |
| RIL-134 | 0.0104-0.0394 | 1 | RIL-45 | 2.2811-2.7601 | 5 |
| RIL-156 | 0.0244-0.0253 | 1 | RIL-46 | 2.3158-6.3697 | 5 |
| RIL-169 | 0.0434-0.0569 | 1 | RIL-49 | 2.0398-2.6978 | 5 |
| RIL-170 | 0.0539-0.1697 | 1 | RIL-62 | 1.2193-2.6308 | 5 |
| RIL-175 | 0.0123-0.0731 | 1 | RIL-63 | 2.0683-2.5504 | 5 |
| RIL-186 | 0.0231-0.0467 | 1 | RIL-68 | 2.2674-4.9395 | 5 |
| RIL-194 | 0.1535-0.1781 | 1 | RIL-72 | 2.8236-3.1560 | 5 |
| RIL-214 | 0.0778-0.1384 | 1 | RIL-84 | 2.0781-2.8191 | 5 |
| RIL-223 | 0.0460-0.1956 | 1 | RIL-92 | 2.1872-4.3090 | 5 |
| RIL-249 | 0.0403-0.0848 | 1 | RIL-95 | 2.6088-2.6854 | 5 |
| RIL-253 | 0.0037-0.0109 | 1 | RIL-97 | 1.6879-2.1144 | 5 |
| RIL-259 | 0.0406-0.0534 | 1 | RIL-102 | 2.7594-2.9148 | 5 |
| RIL-274 | 0.0293-0.0953 | 1 | RIL-113 | 1.4552-2.8464 | 5 |
| RIL-283 | 0.0091-0.0658 | 1 | RIL-119 | 1.2420-3.1363 | 5 |
| RIL-13 | 0.1109-0.2648 | 2 | RIL-122 | 2.4698-2.6681 | 5 |
| RIL-28 | 0.1087-0.4394 | 2 | RIL-125 | 1.6834-2.1306 | 5 |
| RIL-39 | 0.2354-0.2983 | 2 | RIL-139 | 2.5142-3.2524 | 5 |
| RIL-44 | 0.2961-0.3166 | 2 | RIL-148 | 1.8636-5.5925 | 5 |
| RIL-54 | 0.0331-0.2245 | 2 | RIL-150 | 2.4328-3.1815 | 5 |
| RIL-56 | 0.0064-0.3427 | 2 | RIL-167 | 2.5747-2.9479 | 5 |
| RIL-69 | 0.1050-0.2124 | 2 | RIL-168 | 1.9348-3.3892 | 5 |
| RIL-78 | 0.0586-0.2337 | 2 | RIL-174 | 1.1562-2.6991 | 5 |
| RIL-107 | 0.0039-0.3020 | 2 | RIL-182 | 2.7128-3.3572 | 5 |
| RIL-155 | 0.0188-0.2217 | 2 | RIL-185 | 3.1085-4.9160 | 5 |
| RIL-158 | 0.2659-0.4615 | 2 | RIL-212 | 2.0868-2.6547 | 5 |
| RIL-164 | 0.1272-0.3710 | 2 | RIL-213 | 2.0094-3.0880 | 5 |
| RIL-180 | 0.3315-0.3891 | 2 | RIL-216 | 1.1651-1.7900 | 5 |
| RIL-183 | 0.0392-0.3963 | 2 | RIL-233 | 2.4937-2.6506 | 5 |
| RIL-196 | 0.0518-0.2096 | 2 | RIL-238 | 1.1102-3.0323 | 5 |
| RIL-197 | 0.1203-0.2525 | 2 | RIL-240 | 3.0399-6.1309 | 5 |
| RIL-218 | 0.1467-0.2391 | 2 | RIL-247 | 1.5939-2.1915 | 5 |
| RIL-219 | 0.1613-0.2189 | 2 | RIL-248 | 2.1621-4.1447 | 5 |
| RIL-226 | 0.3513-0.4673 | 2 | RIL-265 | 2.6932-2.9406 | 5 |
| RIL-241 | 0.2658-0.3099 | 2 | RIL-267 | 2.7879-4.7565 | 5 |
| RIL-244 | 0.0933-0.2738 | 2 | RIL-275 | 1.5796-2.3936 | 5 |
| RIL-260 | 0.0977-0.2581 | 2 | RIL-276 | 1.3032-2.7370 | 5 |
�´��ڴ�|����CSV
1.2 ����С����ϵĿ����Լ���
���ÿ���ָ��(resistance index, RI)���۹���С����ϵĿ�����[14]����С��ཬ��������, ȡÿ�ظ���ÿ��ϵ10~15������, �ܷ���ֽ����, �������ڼ��������ϵij��ӡ����ݴ��ڵ���������Ŀ, ��������С���������Է�Ϊ5����0��Ϊ��������; 1��Ϊ�������г�1ͷ; 2��Ϊ�������г�2ͷ; 3��Ϊ�������г�3ͷ; 4��Ϊ�����ϵij�����4ͷ���������¹�ʽ����ÿ����ϵ�Ĺ�����ʧ��(L)[2]��L(%) = ��(xf) / 4��f �� 100%
ʽ��, xΪ��Ӧ�����Ŀ��Լ���, fΪ�������������Ŀ��
��ÿ����ϵ�Ĺ�����ʧ��(L)����ȫ��������ϵ(Ʒ��)��ƽ��������ʧ��(ML), ���������ָ��(RI)��ȷ����ϵ�Ŀ��Էּ�����ͬһ��ϵ�ڲ�ͬ�ظ���ļ���������ڽϴ����ʱ, �������ظ����ݽ���L��RIֵ�Ĺ���[2]������RIֵ������С����ϵĿ��Է�Ϊ����(RI=0) (immune, I)���߿�(0.01��RI<0.19) (highly resistant, HR)���п�(0.20��RI<0.49)(moderately resistant, MR)���Ϳ�(0.50��RI<0.99) (lowly resistant, LR)���г�(1.00��RI<1.50) (susceptible, S)�߸�(RI��1.50)(highly susceptible, HS)��
1.3 ��������������
��ǰ���о��л�õ�6��������ػ���TraesCS4A01G436000��TraesCS4A01G436100��TraesCS4A01G436500��TraesCS4A01G437300��TraesCS4A01G437400��TraesCS4A01G437800���п����ǵĿ������������й����ο�����(IWGSC RefSeq v1.0, https://wheat-urgi.versailles.inra.fr/)�ıȶԽ��, ��ȡ��������IJ������С���Primer5�����������ø߱���ø��С���ױ�����24��6218������Ŀ�Ļ���10 ��L PCR��ϵ����ģ��DNA 50 ng�����������0.4 ��mol L-1��1�� Pfu PCR MasterMix (��������Ƽ�(����)����˾)��PCR����Ϊ: (1) 94��, Ԥ����3 min; (2) 94��, 30 s; 55��, 30 s; 72��, 2 min; ��35��ѭ��; (3) 72��������5 min���������ᆳ1%��֬��������Ӿ����, ����Ŀ������, ���������﹤��(�Ϻ�)�ɷ�����˾����˫��������������й���(IWGSCv1.0)�Ͱ�ũAK58 (v4.24)�Ļ��������н��бȶԡ�1.4 �����ǵĿ���
1.4.1 EST (expressed sequence tag, �������б�ǩ)��� ���ݲ�����, ����Ŀ�������С���ױ����InDel, ������InDel��������200 bp������������������ͨDNA�ۺ�ø(2�� Es Taq MasterMix)����PCR������10 ��L PCR��ϵ����ģ��DNA 50 ng�����������0.4 ��mol L-1��1�� Es Taq MasterMix (������Ϊ��������Ƽ�����˾)����Ӧ����Ϊ: (1) 94��, Ԥ����5 min; (2) 94��, 30 s; 57��, 30 s; 72��, 1 min; ��35��ѭ��; (3) 72��������7 min������������30%�ľ۱�ϩ���������ϵ�Ӿ, ��Ⱦ��ɫ��1.4.2 KASP (kompetitive allele-specific PCR, �����Ե�λ����PCR)��� �Կ�����ػ����ڼ���24 (�߿�������)��6218 (�߸�������)����ũAK58 (�߸�������)������ֳ���̬�Ե�SNPλ�㼰�����������С��ο�����(IWGSCv1.0)���бȶ�, ��ѡ���ڻ������о����������л�����һ��SNP��λ��(ͻ������)�ڲο��������в����ڵ�SNP, ����KASP��ǵ�����뿪������ȡѡ��SNP�ڲο��������еIJ�������(>200 bp), �ɰ������Ƽ�(�Ϻ�)����˾(LGC Science Shanghai Ltd.)��������������ϳ�, ������KASP�����������е�5��˼���FAM (6-carboxy-fluorescein)��HEX (Hexachlorofluorescein)ӫ����ŵĽ�ͷ���С����չ�˾�ṩ�ij������KASP��ǵķ������顣��Ӧ��ϵ�����Ϊ3 ��L, ��KASP (V4.0) 2�� Mastermix 1.5 ��L, ����mix (72��) 0.0417 ��L, DNAģ��50~100 ng���ҡ�PCRΪTouchdown��Ӧ����, 94�����15 min; 94�����20 s, 68���˻�60 s, ÿѭ��һ���½�0.6��, 10��ѭ��; 94�����20 s, 62���˻�60 s, 26��ѭ����PCR������Ϻ�, ����LGCϵͳ��SNPlineƽ̨����SNP���ͼ��, ����Kraken (v16.3.16.16288)��������ɨ�����ݡ�
2 ��������
2.1 ���������
6��������ػ����ڿ�����С���ױ�����24��6218�о���������Ŀ��Ƭ�Ρ���TraesCS4A01G436500��, ����5�������ڿ�����С���ױ��е�������ο�����������(�й����Ͱ�ũAK58)���; TraesCS4A01G436500�ڿ����ױ�����24��������ο��������������, ���ڸг��ױ�6218��������ο����������в���ϴ����������ڿ�����С���ױ��������SNP, �Ҷ�����ʹ�����ᷢ���ı�ķ�ͬ��ͻ��λ��(ͼ1)������6218�Ͱ�ũAK58���߸����������, �����Щͬʱ�����ڼ���24��6218֮�䡢����24���ũAK58֮��Ķ�̬��SNP, ����Ϊ�߿��Ŷ�SNP��TraesCS4A01G436100�����д���4����ͬ��ͻ��SNP, ����3��Ϊ���ŶȽϸߵ�SNP��TraesCS4A01G437400�����д���6����ͬ��ͻ��SNP, ����4��Ϊ���ŶȽϸߵ�SNP��TraesCS4A01G437800����1���߿��Ŷȵķ�ͬ��ͻ���SNP��TraesCS4A01G437300����SNP�Ͳ���/ȱʧ(InDel)��1��, ���п���Ʒ��(����24)�д���һ��12 bp�IJ���, ���г�Ʒ��(ϵ)(6218�Ͱ�ũAK58)��ȱʧ����12 bp������δ�ܻ��TraesCS4A01G436500�ڸг��ױ�(6218)�е���������, �ʸ��ݸû����ڿ����ױ�����24��г�Ʒ�ְ�ũAK58����ڵ�2��SNPλ����б�ǿ�����TraesCS4A01G436000�ڿ�����С���ױ��зֱ����1��4 bp��1 bp�IJ���, �������ֲ�����ܵ������Ķ�������ı�(����ͻ��), �������Ԥ���˸û����ڿ����г�С���ױ��е����п����Ķ���(https://web.expasy.org/), �ֱ����ڿ�����С���ױ���ʹ���뵰���Ծ���˿���ᵰ��ø���Ƽ�(SPI)�ṹ��ı�������, �����ڱ����������ϴ���11����ͬ��ͻ��λ��, ����1��Ϊ���ŶȽϸߵ�SNP; ����, �û����ڼ���24�Ļ������δ�����һ��29 bp�IJ���, ���ڸг��ױ���ȱʧ����29 bp�����С�
ͼ1
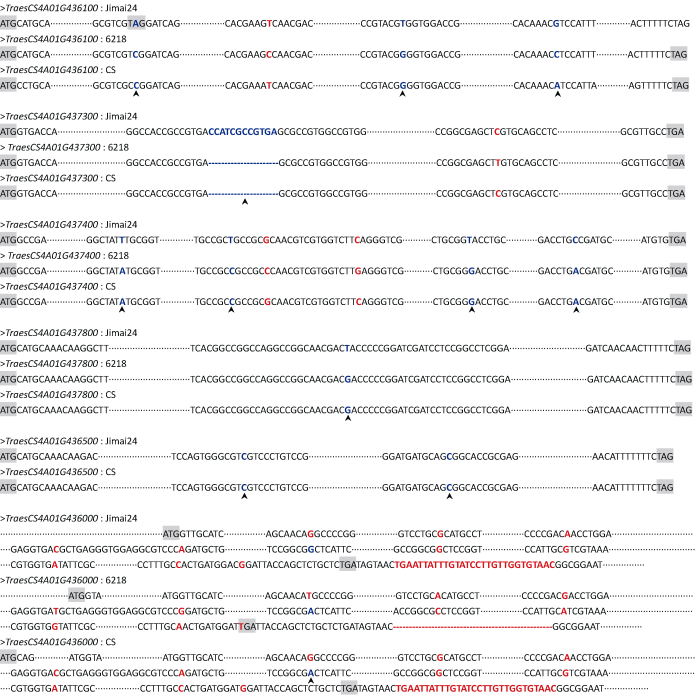 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ16����������ڿ�(����24)����(6218)С���ױ����й����ο�����֮��ıȶԽ��
��ɫ���ɫ���������ͬ��ͻ���SNPλ������ȱʧ, ������ɫ���Ϊ���ŶȽϸߵ�SNPs, ���Լ�ͷ��ע��
Fig. 1Sequence alignments of the six differentially expressed genes from wheat parents, ��6218�� (susceptible) and ��Jimai 24�� (resistant) and the Chinese Spring reference genome
Red or blue font letters represent non-synonymous SNPs and InDels present in the genes. Blue letters with arrowheads represent the SNPs with high confidence.
����Щ�߿��Ŷȵ�SNPλ�㼰�����������С��ο�����������(IWGSCv1.0)�ıȶԽ��, �Ի����������Ի�ͻ���λ��������(������Ŀ�Ļ��������е��ض����λ���Ͼ��п����ױ���ͻ������, ���ڲο���������ȫ��ͬԴ���ж�Ӧ�ļ��λ���Ͼ������ڸ�ͻ������)��SNP, ����KASP��ǿ��������ݱȶԽ��, ѡ�����TraesCS4A01G436100�ϴ��ڵ�2��������λ��, TraesCS4A01G437400��TraesCS4A01G437800��TraesCS4A01G436000��TraesCS4A01G436500�ϴ��ڵĸ�1��������λ��, ��6�����ŶȽϸߵ�������SNP����KASP��ǵĿ���, ѡ�����TraesCS4A01G437300��TraesCS4A01G436000�����д��ڵ�InDel, ����EST��ǵĿ�����
2.2 �����ǵĿ�������̬�Լ��
�������������д��ڵ�SNP��InDel, �ֱ���Ƴ�6��KASP���(K3-1-1��K3-7-1��K3-7-3��K3-16-1��K10-10-6��K10-13-x)��2��EST���(E1-2��E10-10), ����Щ��Ǽ���˿����г�С���ױ���RIL��ϵ��С��Ʒ�֡�2��EST�����С���ױ�������ֳ���̬��, ����E1-2�ڿ����г��ױ��зֱ�������173 bp��161 bp��Ƭ��, E10-10�ֱ�������211 bp��182 bp��Ƭ��(ͼ2)��6��KASP����ڿ����г�С���ױ�������ж�̬��(ͼ3)�����K3-7-1��K3-7-3��K3-16-1��K10-10-6��С���ױ��о�Ϊ���Ϸ���, �ڿ����ױ��зֱ�ΪA:A��T:T��T:T��G:G, �ڸг��ױ��зֱ�ΪC:C��G:G��A:A��A:A��K3-1-1��K10-13-x�ڿ���С���ױ���Ϊ�ӺϷ���(�ֱ�ΪT:G��C:G), �ڸг��ױ��б��ִ��Ϸ���(��ΪG:G)����K3-1-1��K10-13-x��2����������������С��ο�����������(IWGSCv1.0)�ȶԺ���, �г�λ���Ӧ�����ڲο��������о����ڶ��ƥ��(����δ�г�)�����, �ڿ����г�С���ױ���, ��2����Ǿ��ܼ����г��λ��; ������λ���Ӧ������ο�������䲻����3��ĩ��ƥ��, �������ڿ���С��Ʒ�ּ���24�е�ͻ����λ���ڲο��������в�����, ����ڲ����и�ͻ��IJ���������������λ�㡣����, ��Щ��ǿ����ڼ��С��Ʒ�ֻ���ϵ��4AȾɫ�忹��QTL��ͼ2
 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ2EST���E1-2 (A)��E10-10 (B)�ڿ����г�С���ױ�������RILϵ�е��������
M: ������marker (pBR322/Mst Iø��Ƭ��); 1: �г�С���ױ�6218; 2: ����С���ױ�����24; 3~14: �г���ϵ(RIL-16��RIL-19��RIL-20��RIL-21��RIL-25��RIL-29��RIL-45��RIL-102��RIL-113��RIL-119��RIL-125��RIL-139); 15~26: ������ϵ(RIL-7��RIL-12��RIL-18��RIL-23��RIL-64��RIL-73��RIL-91��RIL-115��RIL-134��RIL-156��RIL-169��RIL-170)��
Fig. 2Polyacrylamide gel electrophoresis profiles for EST markers E1-2 (A) and E10-10 (B) in susceptible, or resistant-wheat parent, and selected RIL lines
M: Molecular marker (restriction fragments of plasmid pBR322 digested with Mst I endonuclease); 1: Susceptible wheat parent ��6218��; 2: Resistant parent ��Jimai 24��; 3-14: Susceptible lines (RIL-16, RIL-19, RIL-20, RIL-21, RIL-25, RIL-29, RIL-45, RIL-102, RIL-113, RIL-119, RIL-125, and RIL-139); 15-26: Resistant lines (RIL-7, RIL-12, RIL-18, RIL-23, RIL-64, RIL-73, RIL-91, RIL-115, RIL-134, RIL-156, RIL-169, and RIL-170).
ͼ3
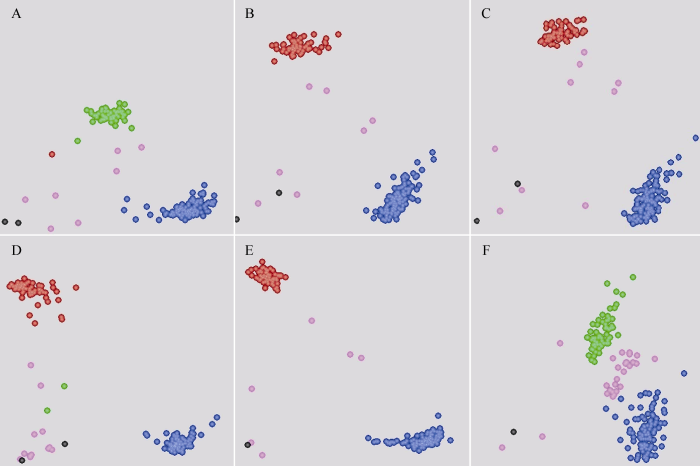 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ3KASP����ڿ����г�С���ױ���92��RILϵ��95��С��Ʒ���еķ��ͽ��
��ɫ��, HEX������; ��ɫ��, FAM������; ��ɫ��, �Ӻϻ�����; ��ɫ��, ȱʧ; ��ɫ��, ����. ͼ��A~F�ֱ�ΪKASP���K3-1-1��K3-7-1��K3-7-3��K3-16-1��K10-10-6��K10-13-x�ķ��ͽ����
Fig. 3KASP assays in wheat parents, 92 RIL lines, and 95 wheat cultivars
Red dots represent the HEX-type allele; blue dots, the FAM-type allele; green dots, the heterozygous-type allele; pink dots, undetected; black dots, the NTC (non-template control); A to F refer to genotypes for K3-1-1, K3-7-1, K3-7-3, K3-16-1, K10-10-6, and K10-13-x, respectively.
2.3 EST��KASP�����С���г�RILϵ�еļ����
����Щ��Ǽ����47������(RI = 0.0330~ 0.4673)��45���г�(RI = 1.2325~6.3697) RILϵ(��1)������KASP��Ǻ�EST����ڿ����г�RIL��ϵ�еļ����(���ͻ����)����Ӧ������С���ױ����Ǻ϶Ƚϸߡ�KASP����ܹ���85%���ϵĿ�����ϵ�м���������λ��(���б��K3-1-1��K10-13-xΪ�ӺϷ���), ��95%���ϵĸг���ϵ�м����г���λ��(��2)������EST���, ���ܹ���85%���ϵĿ�����ϵ���������뿹���ױ�һ�µĴ���, �ڳ���90%�ĸг���ϵ����������г��ױ�һ�µĴ���(��2)��������Ը������ڸ�����ϵ�е�ȱʧ���(���K3-1-1��K3-7-1��K3-7-3��K10-10-6���ֱ���2��RIL��ϵ��ȱʧ, ���K10-13-x��4��RIL��ϵ�д���ȱʧ), KASP����������RIL��ϵ��Ŀ����з��ͽ����ȫһ��, ˵����Щ��Ǿ�Ӧλ��ͬһ��Ⱦɫ�彻��������, ���ҿ����뿹���������������롣Table 2
��2
��2EST��Ǻ�KASP����ڹ���RIL��ϵ�еı�ǻ����ͼ�����
Table 2
| ��� Marker | ����λ��a Physical location a (bp) | ����RILs����ǻ����ͼ����� Marker-based genotypes and their ratio for resistant RIL lines (%) | �г�RILs����ǻ����ͼ����� Marker-based genotypes and their ratio for susceptible RIL lines (%) | |||||
|---|---|---|---|---|---|---|---|---|
| A | H | B | A | H | B | |||
| E1-2 | 707248439 | 40 (85.1) | 0 (0.0) | 7 (14.9) | 2 (4.4) | 0 (0.0) | 43 (95.6) | |
| E10-10 | 705763221 | 40 (85.1) | 0 (0.0) | 7 (14.9) | 4 (8.9) | 0 (0.0) | 41 (91.1) | |
| K3-7-1 | 705889579 | 39 (86.7) | 0 (0.0) | 6 (13.3) | 2 (4.4) | 0 (0.0) | 43 (95.6) | |
| K3-7-3 | 705890140 | 39 (86.7) | 0 (0.0) | 6 (13.3) | 2 (4.4) | 0 (0.0) | 43 (95.6) | |
| K10-10-6 | 705763509 | 39 (86.7) | 0 (0.0) | 6 (13.3) | 2 (4.4) | 0 (0.0) | 43 (95.6) | |
| K3-16-1 | 707251252 | 41 (87.2) | 0 (0.0) | 6 (12.8) | 2 (4.4) | 0 (0.0) | 43 (95.6) | |
| K3-1-1 | 707477512 | 0 (0.0) | 39 (86.7) | 6 (13.3) | 0 (0.0) | 2 (4.4) | 43 (95.6) | |
| K10-13-x | 706095841 | 0 (0.0) | 39 (88.6) | 5 (11.4) | 0 (0.0) | 2 (4.6) | 42 (95.5) | |
�´��ڴ�|����CSV
2.4 EST��KASP����ڲ�ͬ����С��Ʒ���еļ����
����95����ͬ���Ե�С��Ʒ��Ϊ����(��3), ��������ǵ������Խ����˼�⡣���E1-2�ܹ���37.2%�Ŀ���С��Ʒ�������������Ͽ������, ��80%�ĸг�Ʒ�������������ϸг����(��4)������E1-2�ڸг�С��Ʒ���еļ����Ч�ʽϸ�(Ϊ80.0%), ��˸ñ�ǶԾ��п�����λ��С��Ʒ�ֵļ���ȷ��Ϊ80%���ҡ��ڶ�С��Ʒ�ֽ��м��ʱ, ��ֻҪ�ڲ���Ʒ���м����������, ��ô��Ʒ��Ϊ����Ʒ��(������QSm.hbau-4A����λ��)�Ŀ�������80%����, ����Ϊɸѡ����������Դ�ı�ǡ������E10-10�ڿ����г�С��Ʒ�����������Ŀ����л�������ռ�������(��4), �������ֿ����г�С��Ʒ��, ��˸ñ�Dz������ڼ�������С��Ʒ�ֵĿ����ԡ�Table 3
��3
��3����С��Ʒ�ֵı���
Table 3
| С��Ʒ�� Wheat cultivar | ����ָ��(RI) Resistance index | ����ȼ� Classification | С��Ʒ�� Wheat cultivar | ����ָ��(RI) Resistance index | ����ȼ� Classification |
|---|---|---|---|---|---|
| ��81 Xiaoyan 81 | 0.0039-0.0073 | 1 | ����12�� Hanmai 12 | 0.3599-0.8138 | 3 |
| ����47 Jinmai 47 | 0.0026-0.0155 | 1 | ��ũ6425 Henong 6425 | 0.1632-0.8449 | 3 |
| ��ũ6049 Henong 6049 | 0.0000-0.0235 | 1 | �ܺ���1�� Zhouheimai 1 | 0.6180-0.8651 | 3 |
| ʯ��12�� Shimai 12 | 0.0197-0.0468 | 1 | ����12 Zhongmai 12 | 0.3425-0.8812 | 3 |
| ����18 Hengyou 18 | 0.0442-0.0513 | 1 | ��9618 Gaoyou 9618 | 0.0978-0.9218 | 3 |
| ��ũ58-3 Henong 58-3 | 0.0300-0.0770 | 1 | ʯ��14�� Shimai 14 | 0.9019-0.9743 | 3 |
| ʯ��828 Shixin 828 | 0.0013-0.0862 | 1 | ��ũ199 Kenong 199 | 0.8349-1.0414 | 4 |
| ��ũ4198 Henong 4198 | 0.0602-0.1087 | 1 | ����99 Liangxing 99 | 0.3466-1.1006 | 4 |
| ��ũ6028 Xinong 6028 | 0.0015-0.1089 | 1 | ʯ��ׯ10�� Shijiazhuang 10 | 0.0074-1.1150 | 4 |
| ��ũ215 Henong 215 | 0.0900-0.1214 | 1 | ʯ4185 Shi 4185 | 0.3530-1.1329 | 4 |
| PH82-2-2 | 0.0423-0.1223 | 1 | ����7�� Xingmai 7 | 0.6757-1.1334 | 4 |
| ��ũ1093 Kenong 1093 | 0.0678-0.1338 | 1 | ��35 Guan 35 | 0.7848-1.1727 | 4 |
| ����1�� Aifeng 1 | 0.0009-0.1345 | 1 | ��6878 Chang 6878 | 0.1776-1.1764 | 4 |
| ����33 Jinmai 33 | 0.0632-0.1395 | 1 | ʯ��ׯ8�� Shijiazhuang 8 | 1.1628-1.2792 | 4 |
| ��ũ28 Zhongnong 28 | 0.1412-0.1509 | 1 | ��9908 Gaoyou 9908 | 0.1786-1.2974 | 4 |
| ���2�� Fengchan 2 | 0.0302-0.1567 | 1 | ����20 Jimai 20 | 0.6224-1.3508 | 4 |
| �ϴ�2419 Nanda 2419 | 0.0012-0.1645 | 1 | ��0628 Heng 0628 | 1.1138-1.3548 | 4 |
| ����22 Jimai 22 | 0.0636-0.1806 | 1 | ��ũ826 Henong 826 | 1.0058-1.3817 | 4 |
| ����155 Zhongmai 155 | 0.1302-0.2029 | 2 | ��ѡ061 Lunxuan 061 | 0.8530-1.5171 | 5 |
| ����79 Jinmai 79 | 0.0115-0.2049 | 2 | ����0045 Beijing 0045 | 0.7773-1.7354 | 5 |
| ��ũ822 Henong 822 | 0.0567-0.2088 | 2 | ũ��399 Nongda 399 | 0.6830-1.7594 | 5 |
| ��229 Shaan 299 | 0.1115-0.2167 | 2 | ��95��26 Heng 95 guan 26 | 1.2248-1.7931 | 5 |
| ��ũ14 Wennong 14 | 0.1723-0.2213 | 2 | ��4444 Heng 4444 | 1.6026-1.8759 | 5 |
| ����9�� Hanmai No.9 | 0.0259-0.2524 | 2 | ��7228 Heng 7228 | 0.8897-1.8930 | 5 |
| �ٷ�3050 Linfen 3050 | 0.0146-0.2557 | 2 | ����6�� Xingmai 6 | 1.6227-1.9706 | 5 |
| ��Ӳ��2�� Baiyingdong 2 | 0.2241-0.2673 | 2 | ����23 Zhoumai 23 | 1.1381-1.9960 | 5 |
| ����66 Liangxing 66 | 0.1759-0.2731 | 2 | ʯ��618 Shixin 618 | 0.8939-1.9989 | 5 |
| Ӥ��700 Yingbo 700 | 0.1775-0.2770 | 2 | NC2 | 1.4307-2.0183 | 5 |
| ��ũ9206 Henong 9206 | 0.2792-0.2844 | 2 | ��4399 Heng 4399 | 1.7993-2.1132 | 5 |
| ����23 Jimai 23 | 0.0296-0.2923 | 2 | ��4338 Heng 4338 | 2.0068-2.1404 | 5 |
| ʦ��02-1 Shiluan 02-1 | 0.0307-0.3112 | 2 | ����22 Zhoumai 22 | 1.6367-2.2851 | 5 |
| ��5579 Ji 5579 | 0.0469-0.3598 | 2 | ��Ŵ200 Jiru 200 | 0.0244-2.2972 | 5 |
| ��225 Shaan 225 | 0.1824-0.3881 | 2 | ��ũ7106 Henong 7106 | 0.3960-2.4059 | 5 |
| ʯ��21�� Shimai 21 | 0.3801-0.4025 | 2 | ����14 Hanmai 14 | 2.4645-2.6210 | 5 |
| ����175 Zhongmai 175 | 0.0873-0.4178 | 2 | ����6005 Cangmai 6005 | 0.5190-2.6750 | 5 |
| ��5265 Ji 5265 | 0.2441-0.4259 | 2 | ֣��9694 Zhengmai 9694 | 0.4956-2.7112 | 5 |
| ��ũ213 Kenong 213 | 0.2667-0.4381 | 2 | ����119 Cangmai 119 | 1.0727-3.1996 | 5 |
| ʯ��539 Shixin 539 | 0.1862-0.4457 | 2 | ��ũAK58 Bainong AK58 | 3.0456-3.2033 | 5 |
| ����10�� Baomai 10 | 0.1057-0.4679 | 2 | ����028 Cangmai 028 | 2.5945-3.3725 | 5 |
| ��ũ23 Yannong 23 | 0.0583-0.4747 | 2 | ����16 Zhoumai 16 | 1.5595-3.9194 | 5 |
| ʯ��ׯ11�� Shijiazhuang 11 | 0.3940-0.4798 | 2 | ����361 Yanyou 361 | 4.1998-4.7174 | 5 |
| ��ѡ987 Lunxuan 987 | 0.4651-0.4932 | 2 | �ٷ�6035 Linfen 6035 | 1.5158-4.9971 | 5 |
| ����21 Luomai 21 | 0.5366-0.5377 | 3 | ��6358 Ji 6358 | 2.8190-5.1615 | 5 |
| ũ��3432 Nongda 3432 | 0.1076-0.5399 | 3 | ��6003 Cang 6003 | 2.9203-5.2570 | 5 |
| ʯ��16�� Shimai 16 | 0.5887-0.6046 | 3 | ����18 Zhoumai 18 | 5.9047-19.5165 | 5 |
| ������1�� Jingbaimai 1 | 0.3560-0.6230 | 3 | ��� Chinese Spring | �� | �� |
| ��ũ827 Henong 827 | 0.1498-0.6978 | 3 | ��ũ39 Xiannong 39 | �� | �� |
| ��ũ5290 Henong 5290 | 0.6108-0.7991 | 3 |
�´��ڴ�|����CSV
Table 4
��4
��4EST��Ǻ�KASP����ڹ���С��Ʒ���еı�ǻ����ͼ�����
Table 4
| ��� Marker | ����Ʒ�ֵı�ǻ����ͼ����� Marker-based genotypes and their ratios for resistant wheat cultivars | �м���Ʒ�ֵı�ǻ����ͼ����� Marker-based genotypes and their ratios for lowly resistant wheat cultivars | �г�Ʒ�ֵı�ǻ����ͼ����� Marker-based genotypes and their ratios for susceptible wheat cultivars | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| A | H | B | A | H | B | A | H | B | |||
| E1-2 | 16 (37.2) | 7 (16.3) | 20 (46.5) | 3 (25.0) | 0 (0.0) | 9 (75.0) | 5 (12.5) | 3 (7.5) | 32 (80.0) | ||
| E10-10 | 37 (84.1) | 0 (0.0) | 7 (15.9) | 9 (75.0) | 1 (8.3) | 2 (16.7) | 30 (75.0) | 1 (2.5) | 9 (22.5) | ||
| K3-7-1 | 12 (30.8) | 0 (0.0) | 27 (69.2) | 1 (8.3) | 0 (0.0) | 11 (91.7) | 3 (7.7) | 0 (0.0) | 36 (92.3) | ||
| K3-7-3 | 12 (31.6) | 0 (0.0) | 26 (68.4) | 1 (9.1) | 0 (0.0) | 10 (90.9) | 3 (7.7) | 0 (0.0) | 36 (92.3) | ||
| K10-10-6 | 15 (37.5) | 0 (0.0) | 25 (62.5) | 2 (16.7) | 0 (0.0) | 10 (83.3) | 5 (12.8) | 0 (0.0) | 34 (87.2) | ||
| K3-16-1 | 20 (62.5) | 1 (3.1) | 11 (34.4) | 3 (25.0) | 0 (0.0) | 9 (75.0) | 8 (20.5) | 1 (2.6) | 30 (76.9) | ||
| K3-1-1 | 1 (2.5) | 13 (32.5) | 26 (65.0) | 0 (0.0) | 1 (8.3) | 11 (91.7) | 0 (0.0) | 3 (7.7) | 36 (92.3) | ||
| K10-13-x | 0 (0.0) | 15 (38.5) | 24 (61.5) | 0 (0.0) | 1 (11.1) | 8 (88.9) | 0 (0.0) | 5 (20.8) | 19 (79.2) | ||
�´��ڴ�|����CSV
KASP���K3-7-1��K3-7-3��K10-10-6�ڳ���30% (�ֱ�Ϊ30.8%��31.6%��37.5%)�Ŀ���С��Ʒ���м����Ͽ�����λ��, ��85%����(�ֱ�Ϊ92.3%��92.3%��87.2%)�ĸг�С��Ʒ���м����ϸг���λ��(��4)����˵��, �������Ʒ���м���Щ������λ��, ��ù���Ʒ��Ϊ����Ʒ��(������QSm.hbau-4A����λ��)�Ŀ����Ը���85%��Ϊ�г�Ʒ��(����������Ͳ����)�ĸ��ʵ���15%, �����3����ǿ��ԽϺõ�ɸѡ����QSm.hbau-4Aλ��Ŀ���С�����ʡ�
���K3-16-1, �ܹ���62.5%�Ŀ���Ʒ���м������Ͽ�����λ��, ��76.9%�ĸг�Ʒ���м������ϸг���λ��(��4), �����Ч��Ϊ70.4%, ��˸ñ�������ΪƷ�ֿ����Ե�ɸѡ��ǡ�
���K3-1-1��K10-13-x�ڿ���С���ױ���Ϊ�ӺϷ��͡����ͽ������, 2������ܹ��ڳ���30% (�ֱ�Ϊ35.0%��38.5%)�Ŀ���С��Ʒ���м�������λ��, �ڳ���75% (�ֱ�Ϊ92.3%��79.2%)�ĸг�Ʒ���м����ϸг���λ��(��4), ��, ���K3-1-1��K10-13-x��������λ��, ��ù���Ʒ��Ϊ����Ʒ��(������QSm.hbau-4A����λ��)�Ŀ�������75%����, ��Ϊ�г�Ʒ��(����������Ͳ����)�ĸ��ʲ���10%��25%����˱��K3-1-1��K10-13-x�ܸ��õؼ�������QSm.hbau-4Aλ��Ŀ���С�����ʡ�
��һ������7�����E1-2��K3-1-1��K3-7-1��K3-7-3��K3-16-1��K10-10-6��K10-13-x�ڸ���ͬ����С��Ʒ�ֵķ��ͽ��(���Ƽ��ȱʧ��Ʒ��)����, ��Щ����ڸ߿�С��Ʒ���е�ƽ�������Ч��Ϊ64.7% (56.3%~86.7%), ���п�Ʒ���е�ƽ�������Ч��Ϊ19.1% (13.0%~41.2%), �ڸг�Ʒ����Ϊ66.7% (50.0%~75.0%), �ڸ߸�Ʒ����Ϊ94.1% (85.7%~100.0%) (ͼ4)��ͬ��, ���ֻ���Ǹ߿��߸����༫������Ʒ��, ����7����ǶԿ���Ʒ�ֵ�ƽ�������Ч��Ϊ64.7% (56.3%~86.7%), ���ڶԿ���RILϵ�ļ����Ч��(ƽ��86.8%, ��Χ85.1%~ 88.6%); �Ըг�Ʒ�ֵ�ƽ�������Ч��Ϊ94.1% (85.7%~100.0%), ��Ըг�RILϵ�ļ����Ч�ʽӽ�(ƽ��95.5%, ��Χ95.5%~95.6%)(ͼ4)�������RILϵ�����ʽϸ�, ��RILϵ���н�Ϊһ�µ��Ŵ������ܲ��ɷ�, ������ڸ߿�RILϵ�䡢�߿�Ʒ�ּ�ļ����Ч�ʵ��ڶ�Ӧ�߸�RILϵ�߸�Ʒ�ֵ���ʵ, �ٴ�˵��С�����������濹�Ե������Ŵ�������
ͼ4
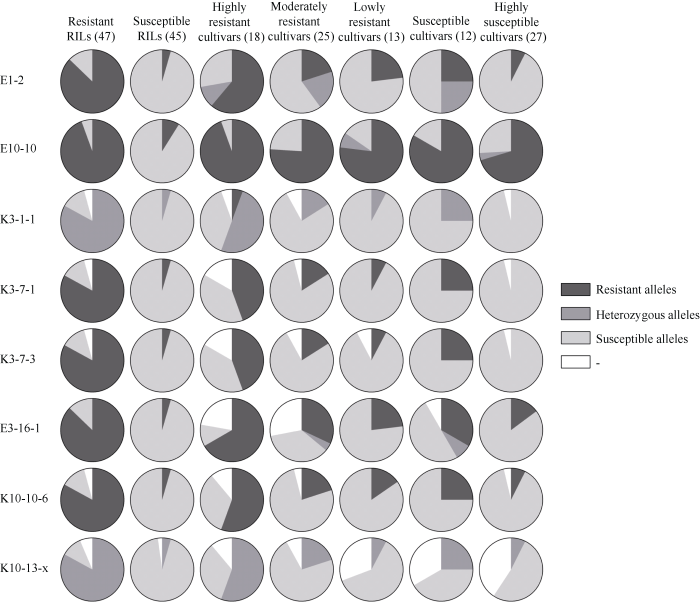 �´��ڴ�|����ԭͼZIP|����PPT
�´��ڴ�|����ԭͼZIP|����PPTͼ4��(47��)����(45��)��С��RIL��ϵ���߿�Ʒ��(19��)���п�Ʒ��(24��)���Ϳ�Ʒ��(12��)���г�Ʒ��(12��)�߸�Ʒ��(28��)�еı�ǻ�������ռ����
Fig. 4Marker-based genotypes and their proportions for resistant- (47) or susceptible- (45) RIL lines, highly resistant- (19), moderately resistant- (24), lowly resistant- (12), susceptible- (12), and highly susceptible- (28) wheat cultivars
����3������ʽϸߵı��K3-1-1��K3-7-1��K3-7-3��95��С��Ʒ���еķ��ͽ�����Կ���, ��3�������11������С��Ʒ��(RI = 0.0235~0.4798)�еķ����뿹���ױ�����24�ķ�����ȫһ��(��5), ��11��Ʒ�ֱַ�����ũ28 (1932����������)����ũ6028 (1956����, ��ͬ)�����2��(1968)������33 (1985)������23 (1986)����ũ215 (�뼽��23������24��Ϊ���ϵ)������9��(2003)��ʯ��ׯ11��(2003)��ʯ��12��(2004)����ũ4198(2005)�ͺ�ũ6049 (2008)��
Table 5
��5
��5��7�����Ա��λ������ȫһ�µ�11��С��Ʒ�ֵĿ����Լ����ǻ�����
Table 5
| С��Ʒ������ Wheat cultivar name | ����ָ��(RI) Resistance index | ����ȼ� Classification | E1-2a | K10-10-6 | K3-7-1 | K3-7-3 | K10-13-x | K3-16-1 | K3-1-1 |
|---|---|---|---|---|---|---|---|---|---|
| ��ũ6049 Henong 6049 | 0.0000-0.0235 | 1 | a | G:G | A:A | T:T | C:G | T:T | T:G |
| ʯ��12�� Shimai 12 | 0.0197-0.0468 | 1 | a | G:G | A:A | T:T | C:G | T:T | T:G |
| ��ũ4198 Henong 4198 | 0.0602-0.1087 | 1 | a | G:G | A:A | T:T | C:G | T:T | T:G |
| ��ũ6028 Xinong 6028 | 0.0015-0.1089 | 1 | a | G:G | A:A | T:T | C:G | T:T | T:G |
| ����33 Jinmai 33 | 0.0632-0.1395 | 1 | a | G:G | A:A | T:T | C:G | T:T | T:G |
| ��ũ28 Zhongnong 28 | 0.1412-0.1509 | 1 | a | G:G | A:A | T:T | C:G | T:T | T:G |
| ���2�� Fengchan 2 | 0.0302-0.1567 | 1 | a | G:G | A:A | T:T | C:G | T:T | T:T |
| ��ũ215 Henong 215 | 0.0502-0.2069 | 2 | a | G:G | A:A | T:T | C:G | T:T | T:G |
| ����9�� Hanmai 9 | 0.0259-0.2524 | 2 | a | G:G | A:A | T:T | C:G | T:T | T:G |
| ����23 Jimai 23 | 0.0296-0.2923 | 2 | a | G:G | A:A | T:T | C:G | T:T | T:G |
| ʯ��ׯ11�� Shijiazhuang 11 | 0.3940-0.4798 | 2 | a | G:G | A:A | T:T | C:G | T:T | T:G |
�´��ڴ�|����CSV
�ۺ�EST��KASP��ǵļ����, ����20��С��Ʒ��[����14������Ʒ��, 5���г���м���(�Ϳ�)Ʒ��, 1���ޱ��ͼ��������Ʒ��(��ũ39)]�����������н϶�Ŀ�����λ��(���������ͬ�����ױ�����24), ����20��Ʒ�����κα��λ���Ͼ�δ�������г���λ��, ���˵����Щ������4AL������λ��QSm.hbau-4A�����Ļ��������ο�������ͬ�ġ�����һЩ�г�С��Ʒ�ֱ����������������п���λ�������, Ҳ���˵���˻������ڿ�������4AL������Чλ��QSm.hbau-4A�����Ļ���(������QSm.hbau-4AЧӦ����λ�����)���ڡ�
3 ����
3.1 ���ܱ�ǵļ����Ч��
���ܱ����һ���ͨ���Ը�������͵ļ�Ӽ������ӦĿ����״(����)�ķ��ӱ��, �����϶Ա��͵ļ����Ӧ�ﵽ��ӽ�100%, �ܹ�ȷ�������������Դ�Ƿ���Ŀ�Ļ���[18,20,22]��������Ӱ����ͱ���Ļ������λ����ܲ�ֹһ��, ʵ�ʿ������Ĺ��ܱ�Dz�һ��������ȫ�����Բ����л�û���������Ӧ�ı�����ȫһ�µĽ��, ��Ը�����״��Ϊ���ԡ��������Lr34/Yr18/Pm38λ��Ĺ��ܱ��csLV34[48]��cssfr1��cssfr2��cssfr3��cssfr4��cssfr5[49], ��273��CIMMYT��Դ��С������л�øߴ�96.7%�ı�Ǽ����Ч��; ������Щ������ڼ���й�С��ط�Ʒ��ʱȴ����, 25.8%�ı������Ʒ��ȴ������ȫ�в�[50]���ŷ����Կ�����ػ���TaNRX��������4�����Ի������, ��Ȼ�ܹ����ҹ�150��С��Ʒ�ֵ������ȷ��ڿ����Խ��м��, ������6%���������Բ��ϱ����Ϊ����������, 2%�Ŀ����Խ�ǿ���ϱ����Ϊ������������[51]�����ù��ܱ�Ǽ��С��������ø(PPO)����ۻ�ɫ�غ���ʱ, Ҳ���ֹ��ܱ�Ǽ������Ͳ���ȫһ�µ�����[52,53,54,55]����Щ����ɹ�����Ŀ��λ������������Ĵ���(��������λ��)Ӱ�쵽Ŀ�Ļ���ı���[50,51], ͬʱҲ�����ų�Ʒ���Ŵ���������ͻ������ص�Ӱ��[56]�����о�����ת¼�������ھĿ�������������, ������8�������Ի�����(2��EST��6��KASP���)����Щ������Ŵ������Ͻ���RILϵ���ܹ��ﵽ�ܸߵļ����Ч��(~90%), ���Ŵ���������ϴ��С��Ʒ����, ��Щ���(������E10-10)�ڸг�Ʒ���еļ����Ч��(76.9%~92.3%)Զ�����ڿ���Ʒ���еļ����Ч��(30.8%~62.5%), ��˵������λ��(QSm.hbau- 4A)֮��������������������[50,51]���Ŵ������IJ�������ܶԱ�ǵļ��Ч�ʲ���Ӱ�졣���, ���о���������7����ǿ�Ӧ����ѡ��С��������ԴQSm.hbau-4Aλ��Ŀ����λ������, Ϊ���С�������濹�Ե���Ҫ�ֶΡ�ͬʱ, ����������С��������д��ڵ������Ŵ����Ӽ����, ���ڽ�ʾС����������ӻ��ƺ����С����ӱ�ǵļ��Ч��Ҳ��ʮ����Ҫ��3.2 ������ر�ǵ�Ӧ��ǰ����չ��
���о���RNA-Seq������BSA����˼·Ӧ����С��������������ǿ�������, �Ӷ��鿹����������С�����ת¼�������з���IJ������, ��qRT-PCR��KEGG Pathway��������ע�͵ȷ�������, ����֤ʵ��6�������������뿹�������; ��������Щ������С�����ױ�֮����ڵ����в���, ������8�����, �ܹ��Ϻõطֱ濹/��RIL��ϵ������7�����K3-1-1��K3-7-1��K3-7-3��K10-10-6��E1-2��K10-13-x��K3-16-1�ڹ��Ըг�С��Ʒ���еļ����Ч�ʽϸ�(76.9%~92.3%, ��4), ���ֻ���Ǹ߸�Ʒ��, �����Ч�������(85.7%~ 100.0%, ͼ4)��������˵, �г�Ʒ�ֲ�ӦЯ���κο����λ����, �����ijЩ�г�Ʒ���д��ڿ���λ��, ��ô��λ��Ͳ�����ȫȷԤ��Ʒ�ֵĿ����ԡ����, �ܹ�ȷԤ��С��Ʒ�ֿ����ԡ�����Ӧ����MAS�����еı��, Ӧ�ڸг�Ʒ���еļ����Ч�ʸ�(�����ܴﵽ100%)�����о���, ���K3-1-1��K3-7-1��K3-7-3�ڹ��Կ���С��Ʒ���еļ����Ч��Ϊ30%����(30.8%~35.0%), �ڹ��Ըг�С��Ʒ���еļ����Ч�ʳ���90% (Ϊ92.3%), �������3����Ǵ�С��Ʒ���м���ȫ��������λ��, ���Ʒ��Ϊ����Ʒ��(����QSm.hbau-4A)�Ŀ�����Ӧ����90%, �����3����ǿɽϺõ���Ϊɸѡ����QSm.hbau-4A����λ��С�����ʵı�ǡ�ͬʱ, ��ͼ4Ҳ�ɿ���, С��Ʒ�ֵ������濹��Խ��, ���ñ�ǵĿ����λ��(resistant alleles)��ռ�������Խ��, �����3����ǿ��Ժܺõ�����Ʒ���Ƿ����QSm.hbau-4A����λ��ļ��, ������Ǽ��������Լ�������ֲ��ϵĿ����Ծ�����Ҫ���塣���о���, ����С��Ʒ������11������Ʒ�ֱ���Я������3����ǵ�ȫ�������λ����(��5), ��ЩƷ�ֶ�Ϊ�������Ѿ�ֹͣ�ƹ����Ʒ��, �ȽϽ���Ʒ������ʱ��Ҳ��10������, �����θ���Ч��ʹ�ñȽ��ϵ�С�����ʴ���С����Դ, ����ʮ�����С����ֻ��ڻ��ת¼�����(BSR-Seq)�����������Ϣѧ���ߡ�qRT-PCR�����������Ŵ���λ��Ϣ�ȵķ���, �����ɿ����ھ����ڿ�����ǵĺ�ѡ������λ��, ����DZ�ڹ��ܱ�ǿ����ٶ�, ������С�������Χ�����������Ŀ����״������صļ��ʡ���Ȼ����Ŀǰ�Ĺ�����δʵ�ֶ�С����ЧQTL��ͼλ��¡, �����ø÷��������DZ�ں�ѡ����Ŀ�����ر��, �ڲ�ͬ��ͼȺ���б�֤�������õ�, �����ܱ����ڶ�С��������Դ�����Եĸ�������, ͬʱ�ɶ�С����������ЧQTL�ľ�ϸ��λ��ͼλ��¡�ṩ��Ҫ������
4 ����
����6��С����������ػ��������д��ڵIJ���λ��, �ɹ���Ʋ�������8��������, ��Щ����ڿ�����С��RIL��ϵ��ķ��ͽ�����Ӧ��ϵ�Ŀ�����ˮƽ�������90%����, �ڲ�ͬС��Ʒ���еļ����Ч�ʱ���ϴ��K3-1-1��K3-7-1��K3-7-3�ܹ��ӳ���30% (30.8%~35.0%)����С��Ʒ���м������λ��, ��90% (92.3%)���ϵĸг�Ʒ���м��г��λ��, ������Щ���ܼ������ǵ�λ�͵�С��Ʒ��, �����4AL������Чλ��QSm.hbau-4A�Ŀ����Գ���90% (Ϊ92.3%), ������С����������Դɸѡ�ͷ��ӱ�Ǹ������֡�����, ����ȫ�������λ��ǵ�11������С��Ʒ��, ���ѻ�ֹͣ�ƹ�, ��Ϊ�����Ч������Ʒ�ֵĿ����Դ���С�������������ʮ�����е�Ҫ����л:
��л�й�ũҵ��ѧԺ�������о�Ա�ṩ�˲���С������, ��л�й�ũҵ��ѧԺ�����ѧ�о����ּ����о�Ա���Թ�ҫ�о�Ա�ṩ�˰�ũAK58 (v4.24)�ο����������в����е���ط���, ��л������������ݼ�����õ��ڿ�������ȷ���Ĵ����������ο����� ԭ��˳��
������ȵ���
������������
�����ڿ�Ӱ������
[��������: 1]
[��������: 1]
[��������: 3]
[��������: 3]
DOI:10.1007/s11032-004-5041-2URL [��������: 4]
DOI:10.1002/ps.1307URLPMID:17078014 [��������: 2]

Field-trapping experiments with synthetic 2,7-nonadiyl dibutyrate, the female-produced sex pheromone of the orange wheat blossom midge, Sitodiplosis mosellana (G��hin), demonstrated that pheromone traps were highly attractive to males and caught very few non-target organisms. Different formulations of pheromone were tested to identify the optimum release rate and dispenser type for use in pheromone traps in the UK. Key findings were that racemic pheromone was as effective as enantiomerically pure (2S,7R)-2,7-nonadiyl dibutyrate, that release rates higher than 0.5 microg day(-1) were not necessary and that the optimal formulation was a 1 mg pheromone loading in a rubber septum. Pheromone traps gave a reliable indication of peak midge emergence, onset of flight and abundance of midges throughout the season. A strong correlation between maximum trap catch and crop infestation levels was obtained.
[��������: 1]
[��������: 1]
DOI:10.1007/s10343-010-0227-5URL [��������: 1]
The intensity of thrips and wheat blossom midges (WBM) infestations in twelve wheat cultivars was evaluated at the Plant Breeding Station, Silstedt, central Germany in 2008 & 2009 growing crop seasons. The research aimed at selecting the least infested cultivar to be profitably used in the forthcoming cultivation. Infestation levels were studied in flowering and milky stages (GS 65 and 73) of each cultivar in every single-spikelet in sample of 10 ears in both years.
There were significant differences in thrips and (WBM) densities among different cultivars in both years. Thrips numbers were the highest in Turkis, Global and Esket cultivars, while the lowest values were recorded in Robigus, Brompton and Carenius. The results showed that the highest WBM infestation was observed in Turkis, Tommi and Potenzial; on the other hand the lowest WBM infestation was found in some insect resistant cultivars (Brompton, Skalmeje, Robigus, Welford and Glasgow). The infested ears were positively correlated with the numbers of WBM among cultivars. The obtained results would give a good guide for choosing the proper cultivars which proved highly resistant to their specific pests.
[��������: 1]
[��������: 1]
DOI:10.7606/j.issn.1009-1041.2003.02.064URL [��������: 4]
����������20���ͺ�50���������������й�С��Ʒ�ֶ���������濹���Լ����ij��÷�������ѧģ�ͼ������Լ������õĶ����������������˱ܺ��ԡ���̬�����Ժ���������������������о������ָ�����Ρ��ۺ��Կ����Ŵ��͵��״��������յ��������йأ��������뿹����������ص���״��������Ϊ�Ŵ������������Ŵ����������ǵ������Ŵ��������ǹѻ����Ŵ��������˿����������������Դ����Ӧ��ȡ˫�ֻ�ѡ���ӽ�ϵ�����Ŀ�ĵؽ���ͬ�������������ۣ�ѡ����һ�������ױ�����Ʒ�֡�
DOI:10.7606/j.issn.1009-1041.2003.02.064URL [��������: 4]
����������20���ͺ�50���������������й�С��Ʒ�ֶ���������濹���Լ����ij��÷�������ѧģ�ͼ������Լ������õĶ����������������˱ܺ��ԡ���̬�����Ժ���������������������о������ָ�����Ρ��ۺ��Կ����Ŵ��͵��״��������յ��������йأ��������뿹����������ص���״��������Ϊ�Ŵ������������Ŵ����������ǵ������Ŵ��������ǹѻ����Ŵ��������˿����������������Դ����Ӧ��ȡ˫�ֻ�ѡ���ӽ�ϵ�����Ŀ�ĵؽ���ͬ�������������ۣ�ѡ����һ�������ױ�����Ʒ�֡�
URL [��������: 1]
���������Sitodiplosis mosellana���ҹ���һ����Ҫũҵ���棬 ���׳�Σ��С�����ڷ����������� �����С�����ؼ����� �������ա��ú�����г���С�� ����ʱ�䳤�� Ϊ�����ε��ص㡣��Щ������ ��ȫ������仯�� �����ƶȸı䡢 С��Ʒ�ָ����� �����ȶ������ص�Ӱ�죬 ������������ҹ��ķ���Σ����������˺ܴ�仯�� �����˱������Ƶ����������������Ҫ�ֲ����ҹ��ı��������� ����Ϊ�����������ԡ� ��Ъ�ԡ� �ֲ��Ժͱ����Ե��ص㡣���ֺ���ķ���Σ���ܳ�Դ������ ��̬���ӡ� ũҵ������ʩ�������ȶ������ص�Ӱ�졣����21���ͺ� ������������ҹ��ķ�����Χ�����˺ܴ�ı仯�� ����Ҫ�ֲ���43°N���ϵ�27°N�Ա��Ķ�С�����������й���������������Ķ�̬�ԡ� С������������Ŀ��Ի����� ����Ʒ�ֵ�ѡ���������Դ�Ŀ����ȷ�����о����ǽ�����Ҫ�о����� δ�������ǿ����������������ķ��ӻ��ơ� ����Σ�����ɡ� Ԥ��Ԥ���� �ۺϷ��κͼ���ֲ�者��������檲�������Ӫ����ϵ�ȷ����о�����������Ϊ����˽�������������ҹ��ķ���Σ�����ɡ� Ԥ��Ԥ�����ۺϷ��ε��ṩ�ο���
URL [��������: 1]
���������Sitodiplosis mosellana���ҹ���һ����Ҫũҵ���棬 ���׳�Σ��С�����ڷ����������� �����С�����ؼ����� �������ա��ú�����г���С�� ����ʱ�䳤�� Ϊ�����ε��ص㡣��Щ������ ��ȫ������仯�� �����ƶȸı䡢 С��Ʒ�ָ����� �����ȶ������ص�Ӱ�죬 ������������ҹ��ķ���Σ����������˺ܴ�仯�� �����˱������Ƶ����������������Ҫ�ֲ����ҹ��ı��������� ����Ϊ�����������ԡ� ��Ъ�ԡ� �ֲ��Ժͱ����Ե��ص㡣���ֺ���ķ���Σ���ܳ�Դ������ ��̬���ӡ� ũҵ������ʩ�������ȶ������ص�Ӱ�졣����21���ͺ� ������������ҹ��ķ�����Χ�����˺ܴ�ı仯�� ����Ҫ�ֲ���43°N���ϵ�27°N�Ա��Ķ�С�����������й���������������Ķ�̬�ԡ� С������������Ŀ��Ի����� ����Ʒ�ֵ�ѡ���������Դ�Ŀ����ȷ�����о����ǽ�����Ҫ�о����� δ�������ǿ����������������ķ��ӻ��ơ� ����Σ�����ɡ� Ԥ��Ԥ���� �ۺϷ��κͼ���ֲ�者��������檲�������Ӫ����ϵ�ȷ����о�����������Ϊ����˽�������������ҹ��ķ���Σ�����ɡ� Ԥ��Ԥ�����ۺϷ��ε��ṩ�ο���
DOI:10.1007/s10530-008-9324-0URL [��������: 1]
Wheat midge, Sitodiplosis mosellana (G��hin) (Diptera: Cecidomyiidae), was first detected as early as 1901 in western Canada. The first major outbreak in Saskatchewan was recorded in 1983. Today wheat midge infests much of the wheat-growing area of Manitoba, Saskatchewan and North Dakota (USA), and is beginning to invade Alberta and Montana (USA). In 1984, Saskatchewan wheat midge populations were found to be parasitized by the egg-larval parasitoid, Macroglenes penetrans (Kirby) (Hymenoptera). Through the successful implementation of conservation techniques, this parasitoid now controls an average of 31.5% of the wheat midge across Saskatchewan. Estimated value of the parasitoid, due to reduction in insecticide costs in Saskatchewan alone, was estimated to be in excess of $248.3 million in the 1990s. The environmental benefits of not having to apply this amount of chemical insecticide are a bonus. To minimize the economic and ecological impact of S. mosellana today, wheat producers in western Canada have access to one of the most comprehensive management programs of any insect pest of field crops. Forecasts and risk warnings, monitoring tools, cultural control, agronomic practices, chemical control, biological control and plant resistance are all available for producers to manage wheat midge.
DOI:10.1007/s10340-010-0325-2URL [��������: 1]
Population densities of wheat ear insects infesting different winter wheat varieties (n = 50) were estimated during 2008 and 2009 seasons near Halle, central Germany. The research was aimed at identifying wheat varieties most resistant to wheat ear insect pests. Two methods were used to evaluate the degree of insect infestations in different wheat ear varieties. Wheat ears were dissected when kernels were in Zadoks stage 73 and examined using a binocular microscope to count the number of spikelets and infested kernels, and to identify the insect pests present. In addition, white water traps were placed on the soil underneath each variety to collect mature larvae of wheat blossom midges (WBMs) as an indicator of potential crop risk for the next year. There were significant differences in the number of thrips and WBM infesting wheat ears among varieties in both years. Thrips numbers were the highest in Akratos, Limes and Ritmo varieties in 2008 and in Michigan Amber, Elegant and Kontrast in 2009. Thrips were the lowest in Thuareg in 2008 and Robigus varieties in 2009. The results showed that the highest WBM infestation level was observed in Michigan Amber in both years. The lowest WBM infestations were found in Turkis, Cubus, Capo, Welford and Robigus in both years. The number of infested kernels was positively correlated with WBM among varieties. In the water traps, the highest numbers of WBM larvae were recorded in Saladin and Bussard in 2008 and Orlando, Julius and Glasgow varieties in 2009. The lowest values were recorded in Victo, Enorm, Robigus and Welford varieties in both years. The results provide a guide for selecting winter wheat varieties with resistance to these wheat ear pests.
[��������: 1]
[��������: 1]
[��������: 7]
[��������: 7]
[��������: 1]
[��������: 1]
DOI:10.7606/j.issn.1009-1041.2011.06.010URL [��������: 1]
Ϊ������ؽ�ʾС�����������Ʒ�֣�ϵ�����Ŵ������ԣ��Ӷ�Ϊ��һ��ѡ������Ʒ���ṩ���ݣ��ڶ�������1 562��С��Ʒ��(ϵ)��ʧ�ʼ����Ļ����ϣ�ȡ47����ȼ�������Խ����Ϊһ�µIJ��ϣ����ñ��ͺ�SSR��ǣ������Ŵ������Է�������Щ�����������Ʒ��ũ����״���ֳ��ϴ�IJ��죬���;������Ŵ�����Ϊ0.68�������Բ��Ϸ�Ϊ6����Ⱥ��19��SSR�����47�ݲ�ͬ����Ʒ���м�104����λ�����ܹ�������Ʒ�����ֿ�����ÿ��������Լ�3��8����λ����ƽ��5.47����47��С��Ʒ�ּ��Ŵ�����Ϊ0.40��0.95��ƽ��Ϊ0.71��SSR��Ǿ���������Ŵ�����Ϊ0.74�������Բ��Ϸ�Ϊ6����Ⱥ��Mental����������������ͬ�����;�������������������(r=0.76,P<0.05)������Ʒ�ֽ���65�ŵ�����Ϊһ�࣬ͬ����Ʒ�־��н�Զ����Ե��ϵ������Ϊ�µĿ�Դ���ڿ������֣����������淢���ؿ��ƹ���ֲ��
DOI:10.7606/j.issn.1009-1041.2011.06.010URL [��������: 1]
Ϊ������ؽ�ʾС�����������Ʒ�֣�ϵ�����Ŵ������ԣ��Ӷ�Ϊ��һ��ѡ������Ʒ���ṩ���ݣ��ڶ�������1 562��С��Ʒ��(ϵ)��ʧ�ʼ����Ļ����ϣ�ȡ47����ȼ�������Խ����Ϊһ�µIJ��ϣ����ñ��ͺ�SSR��ǣ������Ŵ������Է�������Щ�����������Ʒ��ũ����״���ֳ��ϴ�IJ��죬���;������Ŵ�����Ϊ0.68�������Բ��Ϸ�Ϊ6����Ⱥ��19��SSR�����47�ݲ�ͬ����Ʒ���м�104����λ�����ܹ�������Ʒ�����ֿ�����ÿ��������Լ�3��8����λ����ƽ��5.47����47��С��Ʒ�ּ��Ŵ�����Ϊ0.40��0.95��ƽ��Ϊ0.71��SSR��Ǿ���������Ŵ�����Ϊ0.74�������Բ��Ϸ�Ϊ6����Ⱥ��Mental����������������ͬ�����;�������������������(r=0.76,P<0.05)������Ʒ�ֽ���65�ŵ�����Ϊһ�࣬ͬ����Ʒ�־��н�Զ����Ե��ϵ������Ϊ�µĿ�Դ���ڿ������֣����������淢���ؿ��ƹ���ֲ��
[��������: 1]
[��������: 1]
DOI:10.11869/j.issn.100-8551.2014.11.1963URL [��������: 2]
���ܱ���Ǹ�������ͽ�����صĹ��ܻ����ڲ��ض������̬�Ի���������һ�����ͷ��ӱ��.���ڹ��ܱ��ֱ����Դ�ڻ����ڲ��Ĺ����Ի���,���,�����ǿ��ԶԲ�ͬ�Ŵ�������Ŀ���λ�����������ֱ�ӡ����ٵ��ж�.������3��DNA���ӱ�ǻ���ֲ���еĿ�����Ӧ���ص�Ƚ������Ļ�����,�ص�����˹��ܱ�ǵĿ����ص�,���̽�ֹ��ܱ����Ϊһ�ָ��������ֶ��ں̹������ﳣ�������е�Ӧ�ü��俪��ǰ��.
DOI:10.11869/j.issn.100-8551.2014.11.1963URL [��������: 2]
���ܱ���Ǹ�������ͽ�����صĹ��ܻ����ڲ��ض������̬�Ի���������һ�����ͷ��ӱ��.���ڹ��ܱ��ֱ����Դ�ڻ����ڲ��Ĺ����Ի���,���,�����ǿ��ԶԲ�ͬ�Ŵ�������Ŀ���λ�����������ֱ�ӡ����ٵ��ж�.������3��DNA���ӱ�ǻ���ֲ���еĿ�����Ӧ���ص�Ƚ������Ļ�����,�ص�����˹��ܱ�ǵĿ����ص�,���̽�ֹ��ܱ����Ϊһ�ָ��������ֶ��ں̹������ﳣ�������е�Ӧ�ü��俪��ǰ��.
DOI:10.1007/s001220100570URL [��������: 1]
We developed a simple marker technique called sequence-related amplified polymorphism (SRAP) aimed for the amplification of open reading frames (ORFs). It is based on two-primer amplification. The primers are 17 or 18 nucleotides long and consist of the following elements. Core sequences, which are 13 to 14 bases long, where the first 10 or 11 bases starting at the 5′ end, are sequences of no specific constitution (”filler” sequences), followed by the sequence CCGG in the forward primer and AATT in the reverse primer. The core is followed by three selective nucleotides at the 3′ end. The filler sequences of the forward and reverse primers must be different from each other and can be 10 or 11 bases long. For the first five cycles the annealing temperature is set at 35��C. The following 35 cycles are run at 50��C. The amplified DNA fragments are separated by denaturing acrylamide gels and detected by autoradiography. We tested the marker technique in a series of recombinant inbred and doubled-haploid lines of Brassica oleracea L. After sequencing, approximately 45% of the gel-isolated bands matched known genes in the Genbank database. Twenty percent of the SRAP markers were co-dominant, which was demonstrated by sequencing. Construction of a linkage map revealed an even distribution of the SRAP markers in nine major linkage groups, not differing in this regard to AFLP markers. We successfully tagged the glucosinolate desaturation gene BoGLS-ALK with these markers. SRAPs were also easily amplified in other crops such as potato, rice, lettuce, Chinese cabbage (Brassica rapa L.), rapeseed (Brassica napus L.), garlic, apple, citrus, and celery. We also amplified cDNA isolated from different tissues of Chinese cabbage, allowing the fingerprinting of these sequences.
DOI:10.13560/j.cnki.biotech.bull.1985.2016.11.003URL [��������: 2]
���ܱ���Ǹ�������ͽ�����صĹ��ܻ����ڲ��ض�����Ķ�̬������,���ù�������������������RNA���ź�QTL��ͼ�ȷ�������������һ���������Է��ӱ��,�����ǿ��ԶԲ�ͬ�Ŵ�������Ŀ���λ�����������ֱ�ӡ����ٵ��ж��������˹��ܱ�ǵĸ���ص�,����̽�ֹ��ܱ����Ϊһ�ָ��������ֶ���С�������е�Ӧ�ü��俪��ǰ��,����Ϊ��ط��ӱ�ǵĿ����ṩ�ο���
DOI:10.13560/j.cnki.biotech.bull.1985.2016.11.003URL [��������: 2]
���ܱ���Ǹ�������ͽ�����صĹ��ܻ����ڲ��ض�����Ķ�̬������,���ù�������������������RNA���ź�QTL��ͼ�ȷ�������������һ���������Է��ӱ��,�����ǿ��ԶԲ�ͬ�Ŵ�������Ŀ���λ�����������ֱ�ӡ����ٵ��ж��������˹��ܱ�ǵĸ���ص�,����̽�ֹ��ܱ����Ϊһ�ָ��������ֶ���С�������е�Ӧ�ü��俪��ǰ��,����Ϊ��ط��ӱ�ǵĿ����ṩ�ο���
DOI:10.1016/j.tplants.2003.09.010URLPMID:14607101 [��������: 1]
Different approaches (including association studies) have recently been adopted for the functional characterization of allelic variation in plants and to identify sequence motifs affecting phenotypic variation. We propose the term 'functional markers' for DNA markers derived from such functionally characterized sequence motifs. Functional markers are superior to random DNA markers such as RFLPs, SSRs and AFLPs owing to complete linkage with trait locus alleles. We outline the definition, development, application and prospects of functional markers.
[��������: 2]
[��������: 2]
[��������: 1]
[��������: 1]
DOI:10.1046/j.1439-0523.2002.745267.xURL [��������: 3]
DOI:10.1094/PDIS-10-16-1421-REURLPMID:30682944 [��������: 1]
Genetic control of resistance to Fusarium head blight (FHB) is quantitative, making phenotypic selection difficult. Genetic markers to resistance are helpful to select favorable genotypes. This study was conducted to determine if Fhb1 and Fhb5 present in the Sumai 3 source of FHB resistance occur in Sumai 3-derived North American spring wheat cultivars and to understand the appropriateness of using markers to select for the favorable alleles at these loci in breeding. Sumai 3-derived parents Alsen, ND3085, ND744, Carberry, and Glenn were used in crosses to generate 14 doubled haploid breeding populations. The parents and progeny were genotyped with five Fhb1 and three Fhb5 microsatellite markers. Progeny were selected based on performance relative to parents and other control cultivars in FHB nurseries near Portage la Prairie and Carman, MB. ��2 and t test analyses were performed on marker and FHB data. The ��2 test frequently determined the proportion of lines carrying molecular variants associated with FHB resistance increased following nursery selection for FHB. Similarly, the t test regularly demonstrated that selection for FHB resistance lowered the mean level of disease associated with resistant marker haplotypes. The study affirmed FHB resistance sources Alsen, Carberry, ND3085, and ND744 have Fhb1 and Fhb5 loci like Sumai 3, but no evidence was found that Glenn carries Fhb1 and Fhb5 resistance alleles. The results justified use of Fhb1 and Fhb5 markers for marker assisted selection in populations derived from Alsen, Carberry, ND3085, and ND744, but not Glenn. Combined or individual application of Xgwm493 and Xgwm533 in selection of genotypes carrying Fhb1, and Xgwm150, Xgwm304, and Xgwm595 for Fhb5 will enhance FHB resistance in wheat.
DOI:10.1007/s00122-016-2720-4URLPMID:27160855 [��������: 2]
SNP markers were developed for the OWBM resistance gene Sm1 that will be useful for MAS. The wheat Sm1 region is collinear with an inverted syntenic interval in B. distachyon. Orange wheat blossom midge (OWBM, Sitodiplosis mosellana G��hin) is an important insect pest of wheat (Triticum aestivum) in many growing regions. Sm1 is the only described OWBM resistance gene and is the foundation of managing OWBM through host genetics. Sm1 was previously mapped to wheat chromosome arm 2BS relative to simple sequence repeat (SSR) markers and the dominant, sequence characterized amplified region (SCAR) marker WM1. The objectives of this research were to saturate the Sm1 region with markers, develop improved markers for marker-assisted selection (MAS), and examine the synteny between wheat, Brachypodium distachyon, and rice (Oryza sativa) in the Sm1 region. The present study mapped Sm1 in four populations relative to single nucleotide polymorphisms (SNPs), SSRs, Diversity Array Technology (DArT) markers, single strand conformation polymorphisms (SSCPs), and the SCAR WM1. Numerous high quality SNP assays were designed that mapped near Sm1. BLAST delineated the syntenic intervals in B. distachyon and rice using gene-based SNPs as query sequences. The Sm1 region in wheat was inverted relative to B. distachyon and rice, which suggests a chromosomal rearrangement within the Triticeae lineage. Seven SNPs were tested on a collection of wheat lines known to carry Sm1 and not to carry Sm1. Sm1-flanking SNPs were identified that were useful for predicting the presence or absence of Sm1 based upon haplotype. These SNPs will be a major improvement for MAS of Sm1 in wheat breeding programs.
DOI:10.3198/jpr2011.06.0329crcURL [��������: 2]
'Fieldstar' (Reg. No. CV-1067, PI 663950) (Registration. No. 6328 by the Plant Variety Registration Office, Plant Production Division, Seed Section, Canadian Food Inspection Agency, Agriculture and Agri-Food Canada [AAFC]) hard red spring wheat (Triticum aestivum L.) was developed and released by the Cereal Research Centre of AAFC and meets the end-use quality specifications of the Canada western red spring wheat market class. Fieldstar is a partial backcross derivative of 'McKenzie', with 'Clark' as the donor of the Sm1 gene for midge resistance, which produces a product that reduces the palatability of developing seeds to wheat midge larvae (Sitodiplosis mosellana Gehin). Fieldstar is adapted to the eastern wheat growing regions of the Canadian prairies as determined by the Central Bread Wheat Cooperative Registration Test in 2004, 2005, and 2006. For registration testing, the performance of Fieldstar was estimated using the varietal blend Fieldstar VB, which consisted of 90% Fieldstar and 10% 'Waskada'. The grain yield of Fieldstar was similar to that of the highest-yielding checks, McKenzie and 'Superb', and expressed resistance to leaf rust (caused by Puccinia triticina Eriks.) and stem rust (caused by P. graminis Pers.:Pers. f. sp. tritici Eriks. E. Henn.) and intermediate resistance to common bunt [caused by Tilletia tritici (Bjerk.) R. Wolff and T. laevis Kuhn in Rabenh.], loose smut [caused by Ustilago tritici (Pers.) Rostr.], and Fusarium head blight [FHB; caused by Fusarium graminearum Schwabe; teleomorph Gibberella zeae (Schwein.) Petch]. Fieldstar was released because of its combination of high yield, resistance to wheat midge, and intermediate resistance to FHB.
DOI:10.1111/pbr.2011.130.issue-1URL [��������: 3]
[��������: 1]
[��������: 1]
DOI:10.7606/j.issn.1009-1041.2003.03.106URL [��������: 1]
��С��Ʒ�ֽ�����������濹�Լ��������Ի����о�����������Ʒ�֡������������ʵ���ۺ������Ļ�������������������20����50��������������С��Ʒ�ֽ�����������濹�Լ����Ĵ�����Ȼ�г�������������˹��ӳ�����������ڼ���������ѧģ�ͼ��������Լ�Ŀǰ���Լ�����Ӧ�õĵȼ����۱�����ȡ�õĿ��Լ����������ϸ������С��Ʒ�ֿ�������������̬������ơ��ܳ�����Լ��������������Ӫ�����ʺʹ�����л������Ʒ�ֿ����ԵĹ�ϵ�������о����������С��Ʒ�ֶ����������Ŀ�����Ҫ�����ڿ���Ʒ��������ص������������ܺ�������ֹ����ѻ�ȡʳ����ͨ�������ж��Ĵ������ʻ�Ӫ���ϵ�Ƿȱʹ��ȡʳ�����������������������Ļ��Խ����о����ƽ�����չ����
DOI:10.7606/j.issn.1009-1041.2003.03.106URL [��������: 1]
��С��Ʒ�ֽ�����������濹�Լ��������Ի����о�����������Ʒ�֡������������ʵ���ۺ������Ļ�������������������20����50��������������С��Ʒ�ֽ�����������濹�Լ����Ĵ�����Ȼ�г�������������˹��ӳ�����������ڼ���������ѧģ�ͼ��������Լ�Ŀǰ���Լ�����Ӧ�õĵȼ����۱�����ȡ�õĿ��Լ����������ϸ������С��Ʒ�ֿ�������������̬������ơ��ܳ�����Լ��������������Ӫ�����ʺʹ�����л������Ʒ�ֿ����ԵĹ�ϵ�������о����������С��Ʒ�ֶ����������Ŀ�����Ҫ�����ڿ���Ʒ��������ص������������ܺ�������ֹ����ѻ�ȡʳ����ͨ�������ж��Ĵ������ʻ�Ӫ���ϵ�Ƿȱʹ��ȡʳ�����������������������Ļ��Խ����о����ƽ�����չ����
[��������: 1]
[��������: 1]
DOI:10.1007/BF00051118URL [��������: 1]
DOI:10.4039/Ent132591-5URL [��������: 1]
DOI:10.1007/s12298-019-00662-8URLPMID:31168238 [��������: 1]
Sunn pest is one of the most destructive insects in the western and central parts of Asia causing severe reductions of wheat yield and flour quality. Therefore, an effort was undertaken to find effective resistance by analyzing genetic variation among 25 wheat genotypes artificially infested in field-cages as well as using start codon targeted (SCoT) polymorphism and inter- retrotransposon amplified polymorphism (IRAP) markers. High variation was revealed amongst genotypes with Sunn pest resistance characteristics including Bayat, Bezostaya, Sayson, Line93, Line120, Rashagol, Golsepi and AarasGolsoor, which were classified as resistant to moderately resistant. SCoTs and IRAPs were determined as efficient markers for studying genetic diversity. The non-parametric Kruskal-Wallis test was conducted to evaluate the effect of specific SCoT and IRAP amplicons on Sunn pest resistance characteristics for wheat genotypes. The stepwise regression analysis exhibited seven informative SCoTs and IRAPs explaining the highest resistance characteristics variation ranging from 25.7-50.1 to 17.6-40.1, respectively. The relationship between resistance of genotypes and informative SCoT and IRAP amplicons was found based on canonical discriminant analysis showing the capacity of informative markers for functional marker selection method and screening the wheat germplasms for Sunn pest resistance characteristics.
[��������: 2]
[��������: 2]
DOI:10.1023/A:1005412309735URL [��������: 1]
Larvae of the wheat midge, Sitodiplosis mosellana (Géhin) feed on the surface of wheat seeds for about 10 days beginning when pollination occurs. A few wheats have a high level of antibiotic resistance to the larvae, which suppresses their growth and development. Nearly all larvae develop successfully on susceptible wheats. Analysis by HPLC of seed extracts produced by alkaline hydrolysis revealed rapid changes in the levels of p-coumaric and ferulic acids during early seed development. Seed infested by wheat midge larvae showed induced changes in the dynamics of these phenolic acids. The most resistant wheats had a higher constitutive level and a more rapid induction of ferulic acid than susceptible wheats. Levels of ferulic acid exceeding 0.35
 g/g fresh weight were associated with a high mortality of newly hatched larvae. In one wheat line, resistance also was associated with induced production of p-coumaric acid. The induction of ferulic acid was similar in wheat from the laboratory and field, except in one resistant wheat that produced higher levels in the field. In ripe seeds, resistant and susceptible wheats had similar levels of phenolic acids.
g/g fresh weight were associated with a high mortality of newly hatched larvae. In one wheat line, resistance also was associated with induced production of p-coumaric acid. The induction of ferulic acid was similar in wheat from the laboratory and field, except in one resistant wheat that produced higher levels in the field. In ripe seeds, resistant and susceptible wheats had similar levels of phenolic acids.DOI:10.1021/jf010027hURLPMID:11513628 [��������: 1]
The concentration of ferulic acid (FA), the major phenolic acid in the wheat kernel, was found to differ significantly in the mature grain of six wheat cultivars known to have a range of tolerance to the orange wheat blossom midge (Sitodiplosis mosellana). Differences in FA content were correlated with floret infestation level of the cultivars. The wheat cultivars ranked similarly in FA content at the four locations where they were tested, despite a significant effect of environment. Ferulic acid was synthesized mainly during the early stages of grain filling but at different rates among cultivars. Ferulic acid was concentrated primarily in the shorts and bran fractions in an insoluble-bound form. A high correlation was obtained between FA contents as determined by GLC, fluorometry, UV, and colorimetry. The colorimetric procedure was modified as a qualitative, simple, and rapid test for identifying midge-resistant wheat and evaluated in several field trials. The method should provide a rapid tool in the preliminary screening of experimental lines in the development of midge-resistant wheat cultivars.
DOI:10.1155/2013/423189URLPMID:23431288 [��������: 1]
Caffeic acid o-methyltransferase (COMT) is one of the important enzymes controlling lignin monomer production in plant cell wall synthesis. Analysis of the genome sequence of the new grass model Brachypodium distachyon identified four COMT gene homologs, designated as BdCOMT1, BdCOMT2, BdCOMT3, and BdCOMT4. Phylogenetic analysis suggested that they belong to the COMT gene family, whereas syntenic analysis through comparisons with rice and sorghum revealed that BdCOMT4 on Chromosome 3 is the orthologous copy of the COMT genes well characterized in other grass species. The other three COMT genes are unique to Brachypodium since orthologous copies are not found in the collinear regions of rice and sorghum genomes. Expression studies indicated that all four Brachypodium COMT genes are transcribed but with distinct patterns of tissue specificity. Full-length cDNAs were cloned in frame into the pQE-T7 expression vector for the purification of recombinant Brachypodium COMT proteins. Biochemical characterization of enzyme activity and substrate specificity showed that BdCOMT4 has significant effect on a broad range of substrates with the highest preference for caffeic acid. The other three COMTs had low or no effect on these substrates, suggesting that a diversified evolution occurred on these duplicate genes that not only impacted their pattern of expression, but also altered their biochemical properties.
[��������: 1]
[��������: 1]
DOI:10.3864/j.issn.0578-1752.2011.12.001URL [��������: 1]
��Ŀ�ġ�����С����ص�������ø�ṹ�ǻ�����TaPP2Aa�Ĺ��ܱ�Dz���ͼ��Ϊ���ӱ�Ǹ���ѡ���������ṩ���ݡ�������������õ���ͨС����Ұ����Ե��TaPP2Aa�Ļ������У�������SNPλ����죬���3�Ի��������������6�Ի������ڵ�λ����������������й���ȱ�����TaPP2Aa����Ⱦɫ�嶨λ������RILȺ�壨Opata 85��W7984����DHȺ�壨��ѡ10�š�³��14�����иû���Ĺ��ܱ����ͼ���������TaPP2Aa��λ��С���5ͬԴȾɫ��Ⱥ�ϣ�TaPP2Aa-Bλ��RILȺ��5B�ı������Xwg909��Xgwm67����2����ǵ��Ŵ�����ֱ�Ϊ4.0 cM��3.6 cM����DHȺ��5BȾɫ��ı������Xgwm234��WMC363����WMC363���Ŵ�����Ϊ7.5 cM����2���Ŵ�Ⱥ��������Xgwm67�ľ���ֱ�Ϊ3.6 cM��11.4 cM��TaPP2Aa-Dλ��RILȺ��Ⱦɫ��5D�ı������Xcmwg770��Xbarc205���Ŵ�����ֱ�Ϊ9.8 cM��10.0 cM�������ۡ�ȷ����TaPP2Aa���ڵ�Ⱦɫ��λ�ã�ͨ����DH��RIL 2���Ŵ���ͼȺ�������еĿ�����ЧQTL���жԱȷ�������ȷ��TaPP2Aa��С������״QTL�����Ŵ�������ϵ�������Ĺ��ܱ�ǿ�����С������״�ķ��ӱ�Ǹ���ѡ��
DOI:10.3864/j.issn.0578-1752.2011.12.001URL [��������: 1]
��Ŀ�ġ�����С����ص�������ø�ṹ�ǻ�����TaPP2Aa�Ĺ��ܱ�Dz���ͼ��Ϊ���ӱ�Ǹ���ѡ���������ṩ���ݡ�������������õ���ͨС����Ұ����Ե��TaPP2Aa�Ļ������У�������SNPλ����죬���3�Ի��������������6�Ի������ڵ�λ����������������й���ȱ�����TaPP2Aa����Ⱦɫ�嶨λ������RILȺ�壨Opata 85��W7984����DHȺ�壨��ѡ10�š�³��14�����иû���Ĺ��ܱ����ͼ���������TaPP2Aa��λ��С���5ͬԴȾɫ��Ⱥ�ϣ�TaPP2Aa-Bλ��RILȺ��5B�ı������Xwg909��Xgwm67����2����ǵ��Ŵ�����ֱ�Ϊ4.0 cM��3.6 cM����DHȺ��5BȾɫ��ı������Xgwm234��WMC363����WMC363���Ŵ�����Ϊ7.5 cM����2���Ŵ�Ⱥ��������Xgwm67�ľ���ֱ�Ϊ3.6 cM��11.4 cM��TaPP2Aa-Dλ��RILȺ��Ⱦɫ��5D�ı������Xcmwg770��Xbarc205���Ŵ�����ֱ�Ϊ9.8 cM��10.0 cM�������ۡ�ȷ����TaPP2Aa���ڵ�Ⱦɫ��λ�ã�ͨ����DH��RIL 2���Ŵ���ͼȺ�������еĿ�����ЧQTL���жԱȷ�������ȷ��TaPP2Aa��С������״QTL�����Ŵ�������ϵ�������Ĺ��ܱ�ǿ�����С������״�ķ��ӱ�Ǹ���ѡ��
DOI:10.1007/s00122-018-3092-8URLPMID:29666883 [��������: 1]
NGS-assisted super pooling emerging as powerful tool to accelerate gene mapping and haplotype association analysis within target region uncovering specific linkage SNPs or alleles for marker-assisted gene pyramiding. Conventional gene mapping methods to identify genes associated with important agronomic traits require significant amounts of financial support and time. Here, a single nucleotide polymorphism (SNP)-based mapping approach, RNA-Seq and SNP array assisted super pooling analysis, was used for rapid mining of a candidate genomic region for stripe rust resistance gene Yr26 that has been widely used in wheat breeding programs in China. Large DNA and RNA super-pools were genotyped by Wheat SNP Array and sequenced by Illumina HiSeq, respectively. Hundreds of thousands of SNPs were identified and then filtered by multiple filtering criteria. Among selected SNPs, over 900 were found within an overlapping interval of less than 30?Mb as the Yr26 candidate genomic region in the centromeric region of chromosome arm 1BL. The 235 chromosome-specific SNPs were converted into KASP assays to validate the Yr26 interval in different genetic populations. Using a high-resolution mapping population (&gt;?30,000 gametes), we confined Yr26 to a 0.003-cM interval. The Yr26 target region was anchored to the common wheat IWGSC RefSeq v1.0 and wild emmer WEWSeq v.1.0 sequences, from which 488 and 454?kb fragments were obtained. Several candidate genes were identified in the target genomic region, but there was no typical resistance gene in either genome region. Haplotype analysis identified specific SNPs linked to Yr26 and developed robust and breeder-friendly KASP markers. This integration strategy can be applied to accelerate generating many markers closely linked to target genes/QTL for a trait of interest in wheat and other polyploid species.
[��������: 1]
[��������: 1]
DOI:10.1371/journal.pone.0036406URLPMID:22586469 [��������: 1]
Bulked segregant analysis (BSA) is an efficient method to rapidly and efficiently map genes responsible for mutant phenotypes. BSA requires access to quantitative genetic markers that are polymorphic in the mapping population. We have developed a modification of BSA (BSR-Seq) that makes use of RNA-Seq reads to efficiently map genes even in populations for which no polymorphic markers have been previously identified. Because of the digital nature of next-generation sequencing (NGS) data, it is possible to conduct de novo SNP discovery and quantitatively genotype BSA samples by analyzing the same RNA-Seq data using an empirical Bayesian approach. In addition, analysis of the RNA-Seq data provides information on the effects of the mutant on global patterns of gene expression at no extra cost. In combination these results greatly simplify gene cloning experiments. To demonstrate the utility of this strategy BSR-Seq was used to clone the glossy3 (gl3) gene of maize. Mutants of the glossy loci exhibit altered accumulation of epicuticular waxes on juvenile leaves. By subjecting the reference allele of gl3 to BSR-Seq, we were able to map the gl3 locus to an �� 2 Mb interval. The single gene located in the �� 2 Mb mapping interval whose expression was down-regulated in the mutant pool was subsequently demonstrated to be the gl3 gene via the analysis of multiple independent transposon induced mutant alleles. The gl3 gene encodes a putative myb transcription factor, which directly or indirectly affects the expression of a number of genes involved in the biosynthesis of very-long-chain fatty acids.
DOI:10.1111/tpj.12105URL [��������: 1]
The majority of agronomically important crop traits are quantitative, meaning that they are controlled by multiple genes each with a small effect (quantitative trait loci, QTLs). Mapping and isolation of QTLs is important for efficient crop breeding by marker-assisted selection (MAS) and for a better understanding of the molecular mechanisms underlying the traits. However, since it requires the development and selection of DNA markers for linkage analysis, QTL analysis has been time-consuming and labor-intensive. Here we report the rapid identification of plant QTLs by whole-genome resequencing of DNAs from two populations each composed of 2050 individuals showing extreme opposite trait values for a given phenotype in a segregating progeny. We propose to name this approach QTL-seq as applied to plant species. We applied QTL-seq to rice recombinant inbred lines and F2 populations and successfully identified QTLs for important agronomic traits, such as partial resistance to the fungal rice blast disease and seedling vigor. Simulation study showed that QTL-seq is able to detect QTLs over wide ranges of experimental variables, and the method can be generally applied in population genomics studies to rapidly identify genomic regions that underwent artificial or natural selective sweeps.
DOI:10.1016/0305-1978(80)90029-0URL [��������: 1]
DOI:10.1021/np50048a030URL [��������: 1]
DOI:10.1073/pnas.81.24.8014URLPMID:6096873 [��������: 1]
Spacer-length (sl) variation in ribosomal RNA gene clusters (rDNA) was surveyed in 502 individual barley plants, including samples from 50 accessions of cultivated barley, 25 accessions of its wild ancestor, and five generations of composite cross II (CCII), an experimental population of barley. In total, 17 rDNA sl phenotypes, made up of 15 different rDNA sl variants, were observed. The 15 rDNA sl variants comprise a complete ladder in which each variant differs in length from adjacent variants by approximately equal to 115 nucleotide pairs. Studies of four rDNA sl variants in an F2 population showed that these variants are located at two unlinked loci, Rrn1 and Rrn2, each with two codominant alleles. Using wheat-barley addition lines, we determined that Rrn1 and Rrn2 are located on chromosomes 6 and 7, respectively. The nonrandom distribution of sl variants between loci suggests that genetic exchange occurs much less frequently between than within the two loci, which demonstrates that Rrn1 and Rrn2 are useful as new genetic markers. Frequencies of rDNA sl phenotypes and variants were monitored over 54 generations in CCII. A phenotype that was originally infrequent in CCII ultimately became predominant, whereas the originally most frequent phenotype decreased drastically in frequency, and all other phenotypes originally present disappeared from the population. We conclude that the sl variants and/or associated loci are under selection in CCII.
DOI:10.1007/s00122-006-0406-zURL [��������: 1]
Wheat expressed sequence tags (wESTs) were identified in a genomic interval predicted to span the Lr34/Yr18 slow rusting region on chromosome 7DS and that corresponded to genes located in the syntenic region of rice chromosome 6 (between 2.02 and 2.38Mb). A subset of the wESTs was also used to identify corresponding bacterial artificial chromosome (BAC) clones from the diploid D genome of wheat (Aegilops tauschii). Conservation and deviation of micro-colinearity within blocks of genes were found in the D genome BACs relative to the orthologous sequences in rice. Extensive RFLP analysis using the wEST derived clones as probes on a panel of wheat genetic stocks with or without Lr34/Yr18 revealed monomorphic patterns as the norm in this region of the wheat genome. A similar pattern was observed with single nucleotide polymorphism analysis on a subset of the wEST derived clones and subclones from corresponding D genome BACs. One exception was a wEST derived clone that produced a consistent RFLP pattern that distinguished the Lr34/Yr18 genetic stocks and well-established cultivars known either to possess or lack Lr34/Yr18. Conversion of the RFLP to a codominant sequence tagged site (csLV34) revealed a bi-allelic locus, where a variant size of 79bp insertion in an intron sequence was associated with lines or cultivars that lacked Lr34/Yr18. This association with Lr34/Yr18 was validated in wheat cultivars from diverse backgrounds. Genetic linkage between csLV34 and Lr34/Yr18 was estimated at 0.4cM
DOI:10.1007/s00122-009-1097-zURL [��������: 1]
The locus Lr34/Yr18/Pm38 confers partial and durable resistance against the devastating fungal pathogens leaf rust, stripe rust, and powdery mildew. In previous studies, this broad-spectrum resistance was shown to be controlled by a single gene which encodes a putative ATP-binding cassette transporter. Alleles of resistant and susceptible cultivars differed by only three sequence polymorphisms and the same resistance haplotype was found in the three independent breeding lineages of Lr34/Yr18/Pm38. Hence, we used these conserved sequence polymorphisms as templates to develop diagnostic molecular markers that will assist selection for durable multi-pathogen resistance in breeding programs. Five allele-specific markers (cssfr1–cssfr5) were developed based on a 3bp deletion in exon 11 of the Lr34-gene, and one marker (cssfr6) was derived from a single nucleotide polymorphism in exon 12. Validation of reference genotypes, well characterized for the presence or absence of the Lr34/Yr18/Pm38 resistance locus, demonstrated perfect diagnostic values for the newly developed markers. By testing the new markers on a larger set of wheat cultivars, a third Lr34 haplotype, not described so far, was discovered in some European winter wheat and spelt material. Some cultivars with uncertain Lr34 status were re-assessed using the newly derived markers. Unambiguous identification of the Lr34 gene aided by the new markers has revealed that some wheat cultivars incorrectly postulated as having Lr34 may possess as yet uncharacterised loci for adult plant leaf and stripe rust resistance.
URL [��������: 3]
��Ŀ�ġ���ȷCIMMYT�۲���273��С��Ʒ��(ϵ)����������Lr34/Yr18/Pm38λ��ĵ�λ�������ͼ�������ⲡ��Ҷ�ⲡ�Ͱ۲��Ŀ���,��һ����֤���������ܱ�ǵ���Ч�ԡ�������������Lr34/Yr18/Pm38����������STS���csLV34�ͻ��ڸû����11������(exon 11)��λ���쿪����5�Թ��ܱ��cssfr1��cssfr5�����������273��CIMMYTС��Ʒ��(ϵ),ͬʱ�ڳɶ��ͱ����ֱ�������������ⲡ�Ͱ۲��Ŀ��Լ���,�����CIMMYT��������ⲡ��Ҷ�ⲡ���Լ���������з������������STS���csLV34��5�Թ��ܱ��cssfr1��cssfr5�������һ����Ϊ96.7%;��273��CIMMYT��������43�ݲ��Ϻ���Lr34/Yr18/Pm38,�ڲ�ͬ�ص�����ⲡ��Ҷ�ⲡ�Ͱ۲����в�ͬ�̶ȵĿ����ԡ� �����ۡ����ܱ��cssfr1��cssfr5��ȷ����Lr34/Yr18/Pm38λ��exon 11�еĵ�λ����,cssfr3��cssfr4��cssfr5�����ں���Lr34/Yr18/Pm38�����ӽ�����ķ��ӱ�Ǹ���ѡ��
URL [��������: 3]
��Ŀ�ġ���ȷCIMMYT�۲���273��С��Ʒ��(ϵ)����������Lr34/Yr18/Pm38λ��ĵ�λ�������ͼ�������ⲡ��Ҷ�ⲡ�Ͱ۲��Ŀ���,��һ����֤���������ܱ�ǵ���Ч�ԡ�������������Lr34/Yr18/Pm38����������STS���csLV34�ͻ��ڸû����11������(exon 11)��λ���쿪����5�Թ��ܱ��cssfr1��cssfr5�����������273��CIMMYTС��Ʒ��(ϵ),ͬʱ�ڳɶ��ͱ����ֱ�������������ⲡ�Ͱ۲��Ŀ��Լ���,�����CIMMYT��������ⲡ��Ҷ�ⲡ���Լ���������з������������STS���csLV34��5�Թ��ܱ��cssfr1��cssfr5�������һ����Ϊ96.7%;��273��CIMMYT��������43�ݲ��Ϻ���Lr34/Yr18/Pm38,�ڲ�ͬ�ص�����ⲡ��Ҷ�ⲡ�Ͱ۲����в�ͬ�̶ȵĿ����ԡ� �����ۡ����ܱ��cssfr1��cssfr5��ȷ����Lr34/Yr18/Pm38λ��exon 11�еĵ�λ����,cssfr3��cssfr4��cssfr5�����ں���Lr34/Yr18/Pm38�����ӽ�����ķ��ӱ�Ǹ���ѡ��
DOI:10.3724/SP.J.1006.2014.00029URL [��������: 3]
������ػ�����ھ�ͷ��ӱ�ǿ�����ѡ������С��Ʒ������Ҫ���塣����ͬԴ��¡�����ӿ�¡��RACE������������Ϣѧ�����ֶΣ��������ͨС������������(Trx)������һ���»���(TaNRX)��ȫ��cDNA����(GenBank��¼��ΪKC890769)������2015 bp�����п����Ķ���1734 bp��Ԥ�����577�������ᣬ������Ϊ63.79 kD����3��Trx���Թ�����������2�����ڵ��͵�Cys-X1-X2-Cys�ṹ�����д�������ԭ��Ӧ�Ļ��ԡ�TaNRX����λ��С���5BȾɫ��̱��ϣ�����4�������Ӻ�3���ں��ӡ��Ƚϸû��������鼫����Է�ѿ��Ʒ���е����в��죬���ֵ�1���ں��Ӳ������ԡ����ڲ���λ�㿪����4�����Ի�����ǣ���������150��С��Ʒ��(ϵ)����TaNRX��������ͨС�������ٴ���2���뿹����صĵ�λ���죬�ֱ���TaNRX-a��TaNRX-b��TaNRX-a�����͵�Ʒ��(ϵ)ƽ����Է�ѿ����������TaNRX-b������(P<0.01)��˵�������ı�ǿɱ�����С���Լ���ɸѡ��
DOI:10.3724/SP.J.1006.2014.00029URL [��������: 3]
������ػ�����ھ�ͷ��ӱ�ǿ�����ѡ������С��Ʒ������Ҫ���塣����ͬԴ��¡�����ӿ�¡��RACE������������Ϣѧ�����ֶΣ��������ͨС������������(Trx)������һ���»���(TaNRX)��ȫ��cDNA����(GenBank��¼��ΪKC890769)������2015 bp�����п����Ķ���1734 bp��Ԥ�����577�������ᣬ������Ϊ63.79 kD����3��Trx���Թ�����������2�����ڵ��͵�Cys-X1-X2-Cys�ṹ�����д�������ԭ��Ӧ�Ļ��ԡ�TaNRX����λ��С���5BȾɫ��̱��ϣ�����4�������Ӻ�3���ں��ӡ��Ƚϸû��������鼫����Է�ѿ��Ʒ���е����в��죬���ֵ�1���ں��Ӳ������ԡ����ڲ���λ�㿪����4�����Ի�����ǣ���������150��С��Ʒ��(ϵ)����TaNRX��������ͨС�������ٴ���2���뿹����صĵ�λ���죬�ֱ���TaNRX-a��TaNRX-b��TaNRX-a�����͵�Ʒ��(ϵ)ƽ����Է�ѿ����������TaNRX-b������(P<0.01)��˵�������ı�ǿɱ�����С���Լ���ɸѡ��
URL [��������: 1]
��Ŀ�ġ����÷��ӱ���о��й���С��Ʒ��PPO����ĵ�λ���켰����PPO���ԵĹ�ϵ��ΪС��PPO���Եķ��ӱ�Ǹ���ѡ��MAS���춨������������������PPO����Ĺ��ܱ��PPO18��PPO29��PPO16���й�4�������Ķ�С��Ʒ�֣�ϵ���������ɽ��ʡ�����ױ��������311�ݽ���2A��2DȾɫ����PPO��λ����Ppo-A1a��Ppo-A1b��Ppo-D1a��Ppo-D1b�ļ�⡣�������PPO18�ڵ�λ����Ppo-A1a����PPO����Ppo-A1b����PPO���зֱ�����685 bp��876 bp��Ƭ�Σ�PPO16��PPO29��Ppo-D1a����PPO����Ppo-D1b����PPO��������λ�����зֱ�����713 bp��490 bp��Ƭ�Ρ�311��Ʒ�֣�ϵ����Ppo-A1a��Ppo-A1b��Ppo-D1a��Ppo-D1b��λ�����Ƶ�ʷֱ�Ϊ41.8%��58.2%��59.8%��40.2%��PPO������ͬһ�����������λ���������������ˮƽ��P��0.05��������PPO����ĵ�λ����������ͷֲ�Ƶ��Ϊ��Ppo-A1a/Ppo-D1a��27.3%����Ppo-A1a/Ppo-D1b��14.5%����Ppo-A1b/Ppo-D1a��32.5%����Ppo-A1b/Ppo-D1b��25.7%�����˴˲����������ˮƽ��P��0.05������������PPO���Ի���ֲ��������Բ��죬Ppo-A1a�����ڳ��������ζ����������϶������ֲ��϶࣬Ƶ�ʷֱ�Ϊ60.7%��56.0%��Ppo-A1b�����ڱ����������ͻƻ��������ֲ��ϸߣ�Ƶ�ʷֱ�Ϊ65.6%��60.3%����Ppo-D1a�����ڱ������������ƻ������������������ζ����������϶������ֲ����Ը���Ppo-D1b����Ƶ�ʣ��ֱ�Ϊ65.6%��53.6%��75.0%��76.0%�������ۡ�PPO18��PPO16��PPO29��ǵļ�ⷽ�����ȶ��Ժã������ĵ�λ������������Ч��ӳƷ�ֵ�PPO����ֵ���й�������Ʒ�ֵ�PPO���Ի���ֲ�Ƶ����Խ϶ࡣ��ˣ�����PPO�������������ױ���������PPO�����ܱ�����������ɸѡ��PPO���ԵĻ����ͣ��ɴٽ�����Ʒɫ��ĸ�����
URL [��������: 1]
��Ŀ�ġ����÷��ӱ���о��й���С��Ʒ��PPO����ĵ�λ���켰����PPO���ԵĹ�ϵ��ΪС��PPO���Եķ��ӱ�Ǹ���ѡ��MAS���춨������������������PPO����Ĺ��ܱ��PPO18��PPO29��PPO16���й�4�������Ķ�С��Ʒ�֣�ϵ���������ɽ��ʡ�����ױ��������311�ݽ���2A��2DȾɫ����PPO��λ����Ppo-A1a��Ppo-A1b��Ppo-D1a��Ppo-D1b�ļ�⡣�������PPO18�ڵ�λ����Ppo-A1a����PPO����Ppo-A1b����PPO���зֱ�����685 bp��876 bp��Ƭ�Σ�PPO16��PPO29��Ppo-D1a����PPO����Ppo-D1b����PPO��������λ�����зֱ�����713 bp��490 bp��Ƭ�Ρ�311��Ʒ�֣�ϵ����Ppo-A1a��Ppo-A1b��Ppo-D1a��Ppo-D1b��λ�����Ƶ�ʷֱ�Ϊ41.8%��58.2%��59.8%��40.2%��PPO������ͬһ�����������λ���������������ˮƽ��P��0.05��������PPO����ĵ�λ����������ͷֲ�Ƶ��Ϊ��Ppo-A1a/Ppo-D1a��27.3%����Ppo-A1a/Ppo-D1b��14.5%����Ppo-A1b/Ppo-D1a��32.5%����Ppo-A1b/Ppo-D1b��25.7%�����˴˲����������ˮƽ��P��0.05������������PPO���Ի���ֲ��������Բ��죬Ppo-A1a�����ڳ��������ζ����������϶������ֲ��϶࣬Ƶ�ʷֱ�Ϊ60.7%��56.0%��Ppo-A1b�����ڱ����������ͻƻ��������ֲ��ϸߣ�Ƶ�ʷֱ�Ϊ65.6%��60.3%����Ppo-D1a�����ڱ������������ƻ������������������ζ����������϶������ֲ����Ը���Ppo-D1b����Ƶ�ʣ��ֱ�Ϊ65.6%��53.6%��75.0%��76.0%�������ۡ�PPO18��PPO16��PPO29��ǵļ�ⷽ�����ȶ��Ժã������ĵ�λ������������Ч��ӳƷ�ֵ�PPO����ֵ���й�������Ʒ�ֵ�PPO���Ի���ֲ�Ƶ����Խ϶ࡣ��ˣ�����PPO�������������ױ���������PPO�����ܱ�����������ɸѡ��PPO���ԵĻ����ͣ��ɴٽ�����Ʒɫ��ĸ�����
DOI:10.1007/s00122-007-0660-8URL [��������: 1]
Phytoene synthase (Psy), a critical enzyme in the carotenoid biosynthetic pathway, demonstrated high association with the yellow pigment (YP) content in wheat grain. Characterization of Psy genes and the development of functional markers for them are of importance for marker-assisted selection in wheat breeding. In this study, the full-length genomic DNA sequence of a Psy gene (Psy-A1) located on chromosome 7A, was characterized by in silico cloning and experimental validation. The cloned Psy-A1 comprises six exons and five introns, 4,175bp in total, and an ORF of 1,284bp. A co-dominant marker, YP7A, was developed based on polymorphisms of two haplotypes of Psy-A1, yielding 194 and 231-bp fragments in cultivars with high and low YP content, respectively. The marker YP7A was mapped on chromosome 7AL using an RIL population from cross PH82-2/Neixing 188, and a set of Chinese Spring nullisomic–tetrasomic lines and ditelosomic line 7AS. Psy-A1, co-segregating with the STS marker YP7A, was linked to SSR marker Xwmc809 on chromosome 7AL with a genetic distance of 5.8cM, and explained 20–28% of the phenotypic variance for YP content across three environments. A total of 217 Chinese wheat cultivars and advanced lines were used to validate the association between the polymorphic band pattern and grain YP content. The results showed that the functional marker YP7A was closely related to grain YP content and, therefore, could be used in wheat breeding programs targeting of YP content for various wheat-based products.
DOI:10.7606/j.issn.1009-1041.2009.05.007URL [��������: 1]
Ϊ�˸��½�С��Ʒ�����ɫ������ṩ�������ݣ����û�ɫ�غ�������(Psy A1)�Ĺ��ܱ��YP7A�����247���½�С��Ʒ�֣�����ũ��Ʒ�֡�����������Ʒ�ֺ�20����60�������������Ʒ�֣�Psy A1��λ����Psy A1a����ɫ�غ�������Psy A1b���ͻ�ɫ�غ������ķֲ������̽����Psy A1��λ��������ۻ�ɫ�غ����ͻƶ�b*ֵ�Ĺ�ϵ������������½�С��Ʒ����Psy A1a��Psy A1b�����͵�Ƶ�ʷֱ�Ϊ91.9%��8.1%���Ը�ɫ�غ�����Psy A1a������Ϊ�����½�С��ũ��Ʒ�֡�����Ʒ�ֺ�����Ʒ��Psy A1a�����͵ķֲ�Ƶ�ʷֱ�Ϊ100%��81.3%��95.3%������Ʒ�ֵ����������öԽ����½�С��Ʒ�ֻ�ɫ�غ������˻������á��½�С��Ʒ����ۻ�ɫ�غ����ͻƶ�b*ֵ�ձ�ƫ�ߣ���С��Ʒ����ۻ�ɫ�غ������ƶ�b*ֵ���ڴ�С��Ʒ�֡���������Psy A1a��Psy A1b���ֻ����͵���ۻ�ɫ�غ����ͻƶ�b*ֵƽ��ֵ��IJ���ﵽ������ˮƽ����֮���½�С��Ʒ�����ɫ�����Ӧ���������ͻ�ɫ�غ�����Psy A1b������Ϊ�ص㣬ͬʱ����ֱ�����û�ɫ�غ�������(Psy A1)�Ĺ��ܱ��YP7A���������Ч�ʡ�
DOI:10.7606/j.issn.1009-1041.2009.05.007URL [��������: 1]
Ϊ�˸��½�С��Ʒ�����ɫ������ṩ�������ݣ����û�ɫ�غ�������(Psy A1)�Ĺ��ܱ��YP7A�����247���½�С��Ʒ�֣�����ũ��Ʒ�֡�����������Ʒ�ֺ�20����60�������������Ʒ�֣�Psy A1��λ����Psy A1a����ɫ�غ�������Psy A1b���ͻ�ɫ�غ������ķֲ������̽����Psy A1��λ��������ۻ�ɫ�غ����ͻƶ�b*ֵ�Ĺ�ϵ������������½�С��Ʒ����Psy A1a��Psy A1b�����͵�Ƶ�ʷֱ�Ϊ91.9%��8.1%���Ը�ɫ�غ�����Psy A1a������Ϊ�����½�С��ũ��Ʒ�֡�����Ʒ�ֺ�����Ʒ��Psy A1a�����͵ķֲ�Ƶ�ʷֱ�Ϊ100%��81.3%��95.3%������Ʒ�ֵ����������öԽ����½�С��Ʒ�ֻ�ɫ�غ������˻������á��½�С��Ʒ����ۻ�ɫ�غ����ͻƶ�b*ֵ�ձ�ƫ�ߣ���С��Ʒ����ۻ�ɫ�غ������ƶ�b*ֵ���ڴ�С��Ʒ�֡���������Psy A1a��Psy A1b���ֻ����͵���ۻ�ɫ�غ����ͻƶ�b*ֵƽ��ֵ��IJ���ﵽ������ˮƽ����֮���½�С��Ʒ�����ɫ�����Ӧ���������ͻ�ɫ�غ�����Psy A1b������Ϊ�ص㣬ͬʱ����ֱ�����û�ɫ�غ�������(Psy A1)�Ĺ��ܱ��YP7A���������Ч�ʡ�
DOI:10.7606/j.issn.1009-1041.2011.01.009URL [��������: 1]
Ϊ�˸���С��ӹ�Ʒ�ʡ����С������Ч�ʣ�����λ��7AL��7BLȾɫ�������ɫ�غ�����صİ��ⷬ�Ѻ��غ�ø��Phytoene synthase��PSY������Psy A1�ı��YP7A��YP7A 2������Psy B1�ı��YP7B 1��YP7B 2��YP7B 3��YP7B 4���Լ�λ��2ALȾɫ���ϵĶ������ø��Polyphenol oxidase��PPO�����Ի���Ppo A1 �ı��PPO18����221�ݶ�С��Ʒ�֣�ϵ�����л�ɫ�غ����Ͷ������ø���Ի���ĵ�λ�����⡣�������: (1) ��������С��Ʒ�֣�ϵ���У����ͻ�ɫ�غ�����λ����Psy A1b�IJ���78�ݣ�Ƶ��Ϊ35.3%�����ͻ�ɫ�غ�����λ����Psy B1b �IJ���117�ݣ�Ƶ��Ϊ52.9%������ɫ�غ�����λ����Psy B1c�IJ���31�ݣ�Ƶ��Ϊ14.0%����Psy B1d��λ����IJ���4�ݣ�Ƶ��Ϊ1.8%��δ����Я��Psy B1e�IJ��ϡ���2������PPO���Ե�λ����Ppo A1b�IJ���119�ݣ�Ƶ��Ϊ53.8%��(3) ��221�ݲ����У���ɫ�غ����Ͷ������ø���Ի����ͽԷ����й���������ͷ�ӹ�Ʒ��Ҫ���Ʒ�ֽ�25�ݣ��ۺ϶�����������С��Ʒ�ʸ������������ǿ����ʵ��ʹ�õı�Ǿ�Ϊ���������Թ��ܱ�ǣ��ظ��Ժã�ȷ�ʸߣ�����Ч��Ӧ����С��Ʒ�ʸ����ķ��ӱ�Ǹ���ѡ��
DOI:10.7606/j.issn.1009-1041.2011.01.009URL [��������: 1]
Ϊ�˸���С��ӹ�Ʒ�ʡ����С������Ч�ʣ�����λ��7AL��7BLȾɫ�������ɫ�غ�����صİ��ⷬ�Ѻ��غ�ø��Phytoene synthase��PSY������Psy A1�ı��YP7A��YP7A 2������Psy B1�ı��YP7B 1��YP7B 2��YP7B 3��YP7B 4���Լ�λ��2ALȾɫ���ϵĶ������ø��Polyphenol oxidase��PPO�����Ի���Ppo A1 �ı��PPO18����221�ݶ�С��Ʒ�֣�ϵ�����л�ɫ�غ����Ͷ������ø���Ի���ĵ�λ�����⡣�������: (1) ��������С��Ʒ�֣�ϵ���У����ͻ�ɫ�غ�����λ����Psy A1b�IJ���78�ݣ�Ƶ��Ϊ35.3%�����ͻ�ɫ�غ�����λ����Psy B1b �IJ���117�ݣ�Ƶ��Ϊ52.9%������ɫ�غ�����λ����Psy B1c�IJ���31�ݣ�Ƶ��Ϊ14.0%����Psy B1d��λ����IJ���4�ݣ�Ƶ��Ϊ1.8%��δ����Я��Psy B1e�IJ��ϡ���2������PPO���Ե�λ����Ppo A1b�IJ���119�ݣ�Ƶ��Ϊ53.8%��(3) ��221�ݲ����У���ɫ�غ����Ͷ������ø���Ի����ͽԷ����й���������ͷ�ӹ�Ʒ��Ҫ���Ʒ�ֽ�25�ݣ��ۺ϶�����������С��Ʒ�ʸ������������ǿ����ʵ��ʹ�õı�Ǿ�Ϊ���������Թ��ܱ�ǣ��ظ��Ժã�ȷ�ʸߣ�����Ч��Ӧ����С��Ʒ�ʸ����ķ��ӱ�Ǹ���ѡ��
DOI:10.7606/j.issn.1009-1041.2010.02.012URL [��������: 1]
Ϊ����ȷ�Ŵ��ͻ������ض�С��Ʒ�ʵ�Ӱ�죬ѡ��6���д����ԵĶ�С��Ʒ�֣�ԥ��34����8901��ԥ��49��ԥ��70������8716��ԥ��50���ں���ʡ���γ�ȵ㣨32��N~36��N����ֲ���о���Ʒ���Ŵ����ء����������붬С��Ʒ����״�Ĺ�ϵ������������Ŵ����ض�Ӳ�ȡ������ʡ�����ָ��������ʱ���Ӱ��ϴ��������ضԻҷ֡�����ֵ������������Ӱ�졣����������γ�ȵ����ߣ��ҷ֡�����ֵ�ʼ������ƣ����س��������ơ�С��ͬƷ����״���������ӵ�Ӱ��̶Ȳ�ͬ������Ӳ�ȡ��ҷ֡�����ֵ�����ؾ���С����������5�·ݵ���Ҫ�������ӳ�������ء���ˣ�С���������ڵ�������Ӧ��ֿ����������أ��Ը���С��Ʒ�ʡ�
DOI:10.7606/j.issn.1009-1041.2010.02.012URL [��������: 1]
Ϊ����ȷ�Ŵ��ͻ������ض�С��Ʒ�ʵ�Ӱ�죬ѡ��6���д����ԵĶ�С��Ʒ�֣�ԥ��34����8901��ԥ��49��ԥ��70������8716��ԥ��50���ں���ʡ���γ�ȵ㣨32��N~36��N����ֲ���о���Ʒ���Ŵ����ء����������붬С��Ʒ����״�Ĺ�ϵ������������Ŵ����ض�Ӳ�ȡ������ʡ�����ָ��������ʱ���Ӱ��ϴ��������ضԻҷ֡�����ֵ������������Ӱ�졣����������γ�ȵ����ߣ��ҷ֡�����ֵ�ʼ������ƣ����س��������ơ�С��ͬƷ����״���������ӵ�Ӱ��̶Ȳ�ͬ������Ӳ�ȡ��ҷ֡�����ֵ�����ؾ���С����������5�·ݵ���Ҫ�������ӳ�������ء���ˣ�С���������ڵ�������Ӧ��ֿ����������أ��Ը���С��Ʒ�ʡ�

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}