 ,*西南大学水稻研究所 / 转基因植物与安全控制重庆市重点实验室, 重庆 400715
,*西南大学水稻研究所 / 转基因植物与安全控制重庆市重点实验室, 重庆 400715Phenotype characterization and gene mapping of the semi-outcurved leaf mutant sol1 in rice (Oryza sativa L.)
XIE Yuan-Hua**, LI Feng-Fei**, MA Xiao-Hui, TAN Jia, XIA Sai-Sai, SANG Xian-Chun, YANG Zheng-Lin, LING Ying-Hua,*Rice Research Institute of Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Chongqing 400715, China通讯作者:
第一联系人:
收稿日期:2019-04-9接受日期:2019-08-9网络出版日期:2019-09-03
| 基金资助: |
Received:2019-04-9Accepted:2019-08-9Online:2019-09-03
| Fund supported: |
作者简介 About authors
谢园华,E-mail:402307420@qq.com。
李凤菲,E-mail:1071655475@qq.com。

摘要
关键词:
Abstract
Keywords:
PDF (3095KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
谢园华, 李凤菲, 马晓慧, 谭佳, 夏赛赛, 桑贤春, 杨正林, 凌英华. 水稻半外卷叶突变体sol1的表型分析与基因定位[J]. 作物学报, 2020, 46(2): 204-213. doi:10.3724/SP.J.1006.2020.92020
XIE Yuan-Hua, LI Feng-Fei, MA Xiao-Hui, TAN Jia, XIA Sai-Sai, SANG Xian-Chun, YANG Zheng-Lin, LING Ying-Hua.
水稻株型改良对水稻理想株型的形态建成以及提高水稻产量表现具有重要作用[1]。叶型为株型改良性状的重要组成部分, 在水稻植株营养生长的中后阶段, 叶片的适度卷曲可降低群体消光系数, 提高植株的光能利用率, 促进光合产物的积累, 从而增加稻谷产量[2]。
在叶的形态建成中, 叶原基从顶端分生组织中的周边区分化发育后, 进一步沿3个不同的方向极性分化, 即基-顶轴分化(proximo-distal axis polarization), 中-边轴分化(centrol-lateral axis polarization), 近-远轴分化(adaxial-abaxial axis polarization), 从而构成叶片三维体轴, 其中对叶片近-远轴分化发育相关的基因报道很多[3]。水稻叶片卷曲是一个复杂的生物学过程, 受多基因调控, 包括HD-ZipIII家族成员、KANADI家族成员等。HD-ZipIII (Class III Homenodomain-leucine Zipper)基因家族编码由同源异型框-亮氨酸拉链所组成的一典型转录因子家族, 水稻中有OSHB1、OSHB2、OSHB3、OSHB4、OSHB5[4]。水稻中的KANADI (KAN)基因家族成员——SLL1基因通过调控细胞程序性死亡(PCD)过程进而影响近-远轴面的发育, 编码一类MYB类家族转录因子, 属于GARP蛋白家族[2]。KANADI基因家族与HD-ZipIII在侧生器官近远轴面极性建立方面起重要协调作用, 主要是通过miRNA来构成相互抑制的调控网络[5]。
目前在水稻中已定位或克隆的卷叶基因超过30个, 在12条染色体上均有分布, 其中经典的卷叶基因有rl1、rl2、rl3、rl4、rl5、rl6等[6]。已报道基因如NRL1[7,8]、OsHox32[9]、ADL1[10]、OsZHD1[11]、OsARVL4[12]、 OsLBD3-7[13]等的研究结果显示, 泡状细胞是造成叶片卷曲的重要原因。此外, 其他因素如叶肉细胞的排布及角质层的形成等也会引起叶片卷曲。如SLL1基因通过影响厚壁组织和叶肉细胞调控远轴面的发育, 进而改变叶型[2]。rl9内卷突变体是60Coγ射线诱变中花11粳稻品种导致正常株厚壁细胞的位置被占据并影响叶绿体片层基粒排布而形成的[14]。CFL1基因是通过抑制角质层发育的关键基因BDG和FDH的表达, 导致cfl1突变体中叶片角质层发育受到影响, 叶片表现卷曲[15]。
本研究从籼型水稻保持系西农1B的EMS突变体库中, 鉴定到一个半外卷叶突变体sol1 (semi- outcurved leaf 1)。初步表型鉴定表明, sol1从十叶期至成熟期, 叶片向远轴面逐渐卷曲, 且各叶片出现不同程度的披垂。本研究对sol1的表型鉴定、光合特性分析、细胞学分析、遗传特性分析和基因精细定位等, 为后续基因的克隆和功能研究奠定了良好基础。
1 材料与方法
1.1 研究材料
经EMS诱变自育籼型保持系西农1B, 构建了相应的突变体库。通过筛选和连续多代自交, 从该突变体库中鉴定到性状稳定遗传的水稻半外卷叶突变体sol1。将实验室自育恢复系缙恢10号与sol1杂交, 利用F1和F2群体进行遗传分析, 明确目标性状的遗传特性; 并利用F2群体进行目标基因的精细定位。1.2 表型分析
大田种植sol1和西农1B (WT), 鉴定其全生育期植株表型。于成熟期, 随机选取长势一致的sol1与WT 各10株, 考察其叶长、叶宽、有效穗数、一次枝梗、二次枝梗、千粒重、每穗粒数、倒一、倒二、倒三、倒四节节间长度、穗长等性状。于拔节期, 随机选取长势一致的sol1和WT各10株, 分别测其倒一(the 1st leaf from the top, Top 1)、倒二(the 2nd leaf from the top, Top 2)、倒三(the 3rd leaf from the top, Top 3)叶的叶夹角(茎秆与主叶脉之间的角度), 以及倒一、倒二、倒三叶叶片最宽处卷曲时的宽度(Ln)和展开后的宽度(Lw), 计算卷曲指数[LRI = (Lw - Ln)/Lw × 100%][16]。
1.3 细胞学分析
随机选抽穗期sol1和WT各10株, 分别取其倒一叶片相同部位, 于FAA固定液(50%乙醇∶0.9 mol L-1冰乙酸∶甲醛 = 90∶5∶5)中固定, 依次经乙醇脱水, 二甲苯透明, 石蜡包埋, 切片, 番红-固绿对染, 最后在NIKON E6000显微镜下照相观察[17]。1.4 光合效率分析
大田期间, 于乳熟期随机选取sol1与WT各3株, 用Li-6400型便携式光合测定仪测定净光合速率(Pn, μmol m-2 s-1)、蒸腾速率(Tr, mmol m-2 s-1)、胞间CO2浓度(Ci, μmol mol-1)与气孔导度(Gs, mmol m-2 s-1)等光合参数, 分析sol1与WT植株的光合效率。1.5 基因定位
参照Michelmore等[18]的BSA法筛选连锁标记, 从F2植株中分别选取10株正常株和10株突变株剪取等量叶片, 构建正常基因池和突变基因池, 采用改良的CTAB法分别提取亲本和F2群体的基因组DNA, 利用本实验室筛选的均匀分布于水稻12条染色体上的SSR和InDel分子标记, 同时结合数据库(http://www.gramene.org/microsat)和Vector NTI Advance 10 软件开发新的SSR和InDel分子标记, 进行目标基因定位。1.6 RNA提取与qRT-PCR分析
选取长势一致的野生型和突变体各3株, 提取植株叶片的总RNA (RNA prep pure Plant Kit, Tiangen Co. Ltd., China), 并以之为模板反转录为cDNA (Prime Script 1st Strand cDNA Synthesis Kit, TaKaRa)。采用quantitative Real-time PCR (qRT-PCR)方法测量泡状细胞相关的内卷基因和外卷相关基因在野生型和突变体sol1植株中的表达情况。2 结果与分析
2.1 表型鉴定
与WT相比, sol1从十叶期开始叶片表现出轻微的向外卷曲, 随着生育进程其叶片外卷逐渐明显, 且在叶片未完全抽出时叶片基部就表现向远轴面卷曲和褶皱, 叶中上部位则只表现向外卷曲, 同时从苗期至成熟期出现叶片披垂和茎秆半矮化(图1-A~H)。拔节期sol1倒一、倒二、倒三叶的卷曲指数分别为34.83%、40.66%、31.75%, 均极显著高于野生型(5%~6%) (图1-I)。成熟期sol1的倒一、倒二、倒三、倒四节的节间长度和穗长较WT分别缩短13.68%、25.82%、34.00%、29.77%和30.77% (P<0.01)(图1-J, K)。拔节期sol1的倒一、倒二、倒三叶的叶夹角较WT分别增大了74.82%、19.20%、39.86% (图1-L)。图1
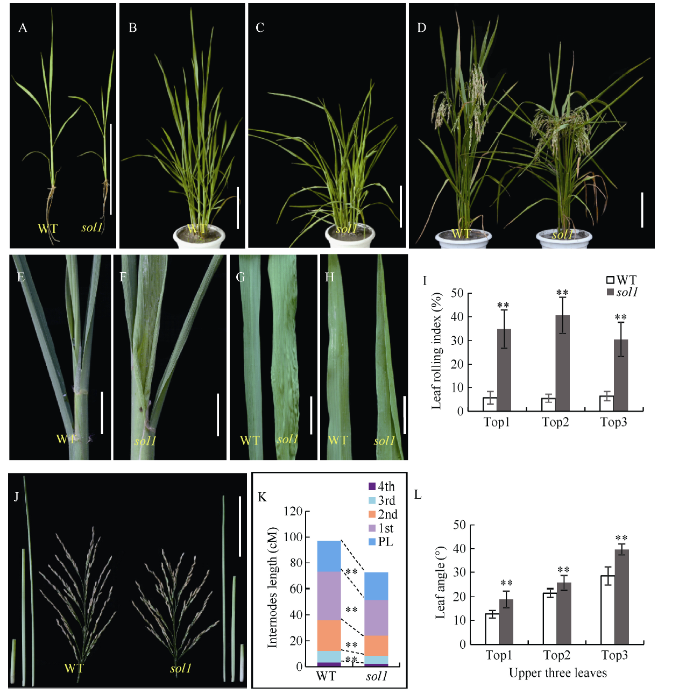 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1野生型(WT)与突变体sol1的表型鉴定
A、B、C、D: 分别代表WT与sol1的五至六叶期、分蘖期、成熟期表型; E、F: 分别为WT与sol1未完全抽出时的叶基部; G: WT与sol1倒一叶近轴面基部; H: WT与sol1倒一叶远轴面叶尖; I: WT与sol1拔节期叶片卷曲指数; J: WT与sol1穗长及各节间长; K: 成熟期WT与sol1的各节间比较; L: WT与sol1拔节期叶夹角。图A、B、C、D、J中, Bar = 10 cm; 图E、F、G、H中, Bar = 2 cm。Top 1: 叶片顶端往下第一片叶; Top 2: 叶片顶端往下第二片叶; Top 3: 叶片顶端往下第三片叶。*: 在0.01 ≤ P < 0.05区间差异显著, **: P < 0.01差异极显著。
Fig. 1Phenotypic identification of the wild type(WT) and the sol1 mutant
A, B, C, D: plant phenotypes of WT and sol1 at the 5-6 leaf stage, the tillering stage and the mature stage, respectively; E and F: incompletely extracted leaf base of WT and sol1, respectively; G: bases of the abaxial top 1 of WT and sol1; H: tips of the adaxial top 1 of WT and sol1; I: leaf curl indexes of WT and sol1 at jointing stage; J: lengths of the spike and internodes of WT and sol1; K: comparison of the internodes of WT and sol1 at mature period; L: leaf angles of WT and sol1 at the shooting period. Bar = 10 cm, in A, B, C, D, J diagrams; Bar = 2 cm, in E, F, G, H diagrams. Top1: the 1st leaf from the top; Top2: the 2nd leaf from the top; Top3: the 3rd leaf from the top. * means significant difference at 0.01 ≤ P < 0.05 by t-test; ** means significant difference at P < 0.01 by t-test.
与WT相比, 孕穗期突变体sol1倒一叶叶长无明显变化、叶宽较WT显著增加7.27%; 倒二叶叶宽无变化, 叶长显著缩短9.17%; 而倒三叶的叶长、叶宽则显著或极显著变短、变窄, 较WT分别降低35.76%和11.56% (图2-A, B)。与WT相比, 突变体sol1除每穗粒数、二次枝梗数无显著差异外, 其有效穗数、千粒重、结实率均显著或极显著减少, 一次枝梗数则显著增多(图2-C~H)。
图2
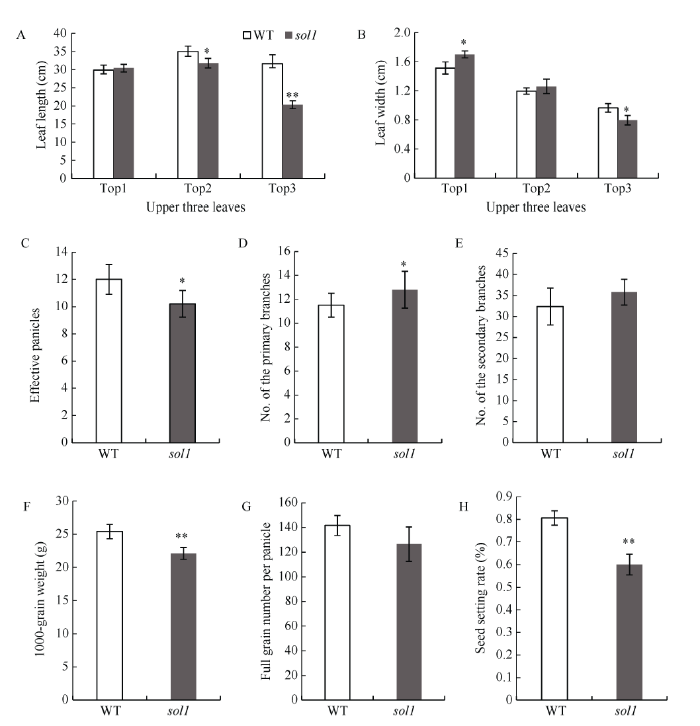 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2野生型(WT)与突变体sol1的农艺性状
A、B: 孕穗期倒一、倒二、倒三叶叶长及叶宽(cm); C: 有效穗数; D: 一次枝梗; E: 二次枝梗; F: 千粒重(g); G: 每穗实粒数; H: 结实率(%)。倒一叶: 叶片顶端往下第一片叶; 倒二叶: 叶片顶端往下第二片叶; 倒三叶: 叶片顶端往下第三片叶。*: 在0.01 ≤ P < 0.05区间差异显著, **: P<0.01差异极显著。
Fig. 2Agronomic characters of the wild type (WT) and the sol1 mutant
A, B: length and width of tops 1, 2, 3 at booting stage (cm); C: numbers of effective panicles; D: primary branches; E: secondary branches; F: 1000-grain weights (g); G: filled grain numbers per panicle; H: Seed setting rate (%); Top 1: the 1st leaf from the top; Top 2: the 2nd leaf from the top; Top 3: the 3rd leaf from the top. * means significant difference at 0.01 ≤ P < 0.05 by t-test; ** means significant difference at P < 0.01 by t-test.
2.2 sol1的光合效率分析
突变体sol1的净光合速率除倒二叶与野生型的差异无统计学意义外, 倒一、倒三叶均极显著下降; sol1的蒸腾速率、胞间CO2浓度、气孔导度均显著或极显著高于野生型。Tr较WT分别增加了161.52%、211.12%、82.08%, 差异均达到极显著水平; Ci均极显著上升; 倒一、倒二、倒三叶叶尖、叶基部Gs均极显著或显著增加(图3-A~D)。由此说明可能是上三叶叶型的改变引起光合特性参数的变化, 进而影响光能利用率和光合作用效率。图3
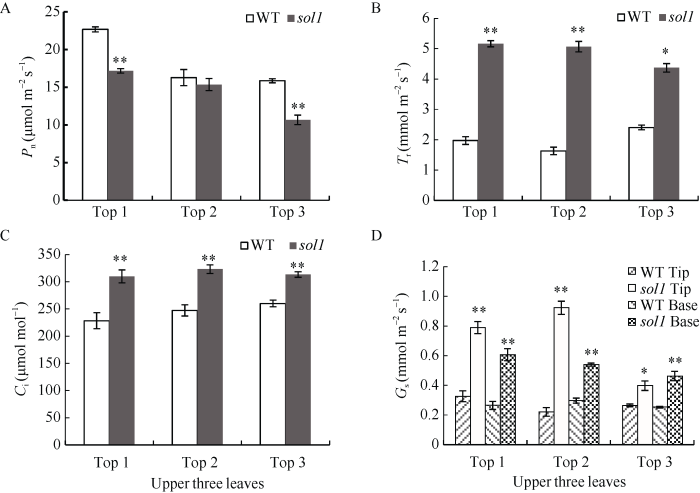 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3乳熟期野生型(WT)与突变体sol1叶片光合效率分析
A: 倒一、倒二、倒三叶净光合速率(Pn); B: 倒一、倒二、倒三叶蒸腾速率(Tr); C: 倒一、倒二、倒三叶胞间CO2浓度(Ci); D: 倒一、倒二、倒三叶叶尖、叶基气孔导度(Gs)。Top 1: 叶片顶端往下第一片叶; Top 2: 叶片顶端往下第二片叶; Top 3: 叶片顶端往下第三片叶。*: 在P < 0.05水平上差异显著, **: 在P < 0.01水平上差异显著。
Fig. 3Analysis of photosynthetic efficiency of blade at maturation period
A: net photosynthetic rate (Pn) of tops 1, 2, 3; B: transpiration rate (Tr) of tops 1, 2, 3; C: intercellular CO2 concentration (Ci) of tops 1, 2, 3; D: stomatal conductance (Gs) of the tip and base of tops 1, 2, 3. Top 1: the 1st leaf from the top; Top 2: the 2nd leaf from the top; Top 3: the 3rd leaf from the top. * significant difference at P < 0.05; ** significant difference at P < 0.01.
2.3 sol1叶片的细胞学分析
石蜡切片观察抽穗期倒一叶横切面, 发现sol1叶片向远轴面呈不对称卷曲, 而WT则基本平直展开(图4-A, B)。进一步放大观察, 发现sol1相邻维管束之间的泡状细胞向内凹陷并变小, 其数量和总面积较WT分别增加了38.88%和34.83%, 差异达显著水平(图4-C~H)。此外, 叶片上下表皮细胞略微增大, 泡状细胞下的叶肉细胞厚度略变薄。因此, 突变体sol1叶片外卷可能是其表皮泡状细胞异常所致。图4
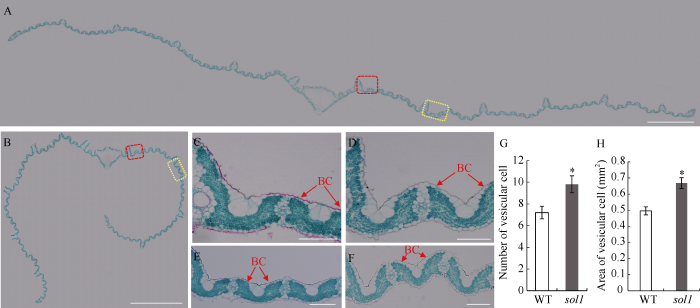 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4叶片横切面石蜡切片分析
A、B: 分别为抽穗期WT和sol1倒一叶横切图, Bar = 1 mm; C、D: 分别为A、B图中红色线框的放大图; E、F: 分别为A、B图中黄色线框的放大图, Bar = 0.1 mm; G: 两小维管束之间的泡状细胞数量; H: 两小维管束之间的泡状细胞面积。BC: 泡状细胞, *: 在0.01 ≤ P < 0.05区间差异显著。
Fig. 4Paraffin-cut section analysis of cross section of leaf
A, B: cross cutting diagrams of top 1 of WT and sol1 at the heading stage, respectively, Bar = 1 mm; C, D: magnified diagrams of a red box in picture A and B, respectively; E, F: magnified diagrams of a yellow box in picture A and B, respectively, Bar = 0.1 mm; G: number of bulliform cells between two small vein; H: area of bulliform cells between two small vein. BC: bulliform cell. * means significant difference at 0.01 ≤ P < 0.05 by t-test.
2.4 遗传分析
以缙恢10号为父本, sol1为母本, 杂交后F1代表型正常; F2单株则出现正常表型和半外卷表型。在调查的4251个F2单株中, 正常表型3226株, 突变表型1025株。χ2检验结果显示, 该F2群体中正常株与突变株分离比为3.25, 符合3﹕1的理论比例(χ2 = 1.74<χ20.05, 1 = 3.84), 因此可以推断出sol1的目标性状受1对隐性核基因控制。2.5 基因定位
利用本实验室筛选的、均匀分布在水稻12条染色体上的400多个SSR标记和98个InDel标记, 筛选西农1B和缙恢10号的多态性。筛选出96个多态性标记用于扩增亲本及正常表型基因池和突变表型基因池, 发现6号染色体短臂上的RM204标记在这2个基因池之间存在稳定差异, 预测该标记可能与SOL1基因连锁。进一步利用F2单株验证, 确定了RM204与SOL1的连锁。在RM204附近查找并开发新标记, 最终将SOL1基因初步定位在RM204和ZTQ54之间, 遗传距离分别为13.07 cM和19.28 cM (图5)。图5
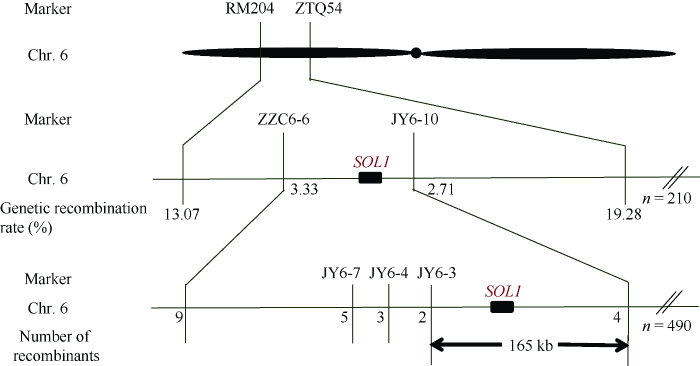 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5SOL1候选基因定位
Fig. 5Location of candidate gene SOL1
进一步开发初步定位区间的InDel标记, 筛选出ZZC6-6、JY6-7、JY6-4、JY6-3、JY6-10等标记(表1)在sol1与缙恢10号之间存在多态性。利用这些新开发的多态性标记对F2突变单株进行基因型鉴定, 结果显示ZZC6-6、JY6-7、JY6-4、JY6-3、JY6-10这5个标记在490个随机选取的F2单株中, 前4个标记分别检测到9、5、3、2个交换单株, 后1个标记检测到4个交换单株, 且两侧的交换单株不同, 从而最终将SOL1基因定位在JY6-3和JY6-10之间165 kb的物理距离内(图5)。
Table 1
表1
表1第6号染色体上新开发的连锁标记引物
Table 1
| 引物名称 Primer | 正向引物 Forward primer (5′-3′) | 反向引物 Reverse primer (5′-3′) |
|---|---|---|
| RM204 | CTAGCTAGCCATGCTCTCGTACC | CTGTGACTGACTTGGTCATAGGG |
| ZTQ54 | AAATAGATAGTAACGGCTCTAACATG | CTTGCACAACTGCTGTTCATG |
| ZZC6-6 | TCCATTGCTCCCAAACACTC | CGTGGAACCAAATACAGCCT |
| JY6-7 | GTGAGTGCTAAGGATGATGTGTTG | TGCAGGAAGGCATTGTATCC |
| JY6-4 | GCAGCAAAGATCCATACGAGGT | AATCTTGGCGGCTCGTGTAG |
| JY6-3 | CGAACACATGCCACACACACA | GGGCTAGACAAACAATAATGTGC |
| JY6-10 | TGGCAGCGGTGGTGGTATAT | CAATGAGAAGTTGGTGGTGTTCT |
新窗口打开|下载CSV
检索结果显示, 精细定位区间内共含有15个注释基因。其中7个编码表达蛋白, 其余8个编码反转录转座子蛋白、叶片衰老相关蛋白、环核苷酸门控离子通道、同源异型结构域和START结构域、转录延伸因子复合物SPT5、LysM-GPI锚定蛋白前体、天冬氨酸蛋白酶的猪笼草蛋白前体(表2)。
Table 2
表2
表2定位区间内的候选基因注解
Table 2
| 基因命名 Gene nomenclature | 基因注释 Gene annotation |
|---|---|
| LOC_Os06g10540 | 表达蛋白 Expressed protein |
| LOC_Os06g10550 | 反转录转座子 Retrotransposon protein, putative, unclassified, expressed |
| LOC_Os06g10560 | 叶片衰老相关蛋白 Leaf senescence related protein, putative, expressed |
| LOC_Os06g10570 | 表达蛋白 Expressed protein |
| LOC_Os06g10580 | 环核苷酸门控离子通道 Cyclic nucleotide-gated ion channel, putative, expressed |
| LOC_Os06g10590 | 表达蛋白 Expressed protein |
| LOC_Os06g10600 | 同源异型结构域和START结构域 Homeobox and START domains containing protein, putative, expressed |
| LOC_Os06g10610 | 表达蛋白 Expressed protein |
| LOC_Os06g10620 | 转录延伸因子复合物SPT5 Transcription elongation factor SPT5 homolog, putative, expressed |
| LOC_Os06g10650 | 蛋白质酪氨酸磷酸酶家族 Tyrosine phosphatase family protein, putative, expressed |
| LOC_Os06g10660 | 锚定蛋白前体LysM Domain-containing GPI-anchored protein 1 precursor, putative, expressed LysM-GPI |
| LOC_Os06g10670 | 天冬氨酸蛋白酶的猪笼草蛋白前体 Aspartic proteinase nepenthesin-1 precursor, putative, expressed |
| LOC_Os06g10680 | 表达蛋白 Expressed protein |
| LOC_Os06g10710 | 表达蛋白 Expressed protein |
| LOC_Os06g10720 | 表达蛋白 Expressed protein |
新窗口打开|下载CSV
2.6 相关基因表达量分析
突变体叶片的细胞学结果表明, 叶片外卷表型主要由泡状细胞的变化引起, 因此选取了水稻中与泡状细胞相关的内卷基因和外卷相关基因进行qRT-PCR分析。结果发现, 与WT相比, 在突变体sol1中RL14、Roc5、REL1的表达量呈不同程度的上调, 而NRL、BRD1、OsHox32、ADL1、LC2则呈不同程度的下调(图6), 表明泡状细胞相关的内卷基因和外卷相关基因可能共同参与调控基因SOL1的外卷表型。图6
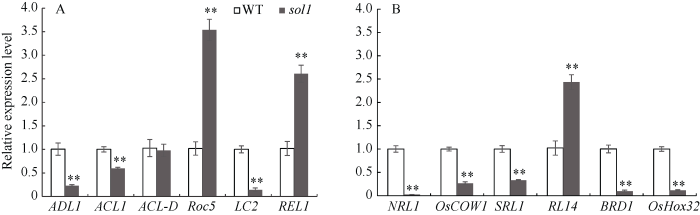 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6叶片卷曲相关基因的qRT-PCR
A: 泡状细胞发育相关的卷叶基因; B: 远轴面卷曲的相关基因。
Fig. 6qRT-PCR of genes related to curled leaf and dwarf internode
A: some genes in bulliform cell development-related rolling leaf; B: genes about some abaxially curled leaf.
3 讨论
叶片是水稻进行光合作用以及气体和水分交换的重要器官, 是塑造理想株型的重要性状之一。水稻叶片的适度卷曲, 有利于保持叶片挺立状态, 改善植株上层叶片的光合速率, 整体提升其光合产物总量的积累[2]。利用突变体研究水稻叶片的形态发育机制, 有助于丰富水稻卷叶基因的遗传资源, 对理想高产株型的改良有重要意义[19]。在水稻叶片的近轴面一般含有4~5个左右泡状细胞, 并呈中间大两头小类似扇形的结构。泡状细胞是一种特化的表皮细胞, 也是调节水分的高度液泡化的薄壁细胞[20]。一般而言, 泡状细胞失水时叶片易向近轴面卷曲, 吸水后易向远轴面卷曲[21]。研究发现, 泡状细胞数量或体积是水稻叶片形态的关键调控因素之一, 如NRL1、OsHox32、ADL1、ACL-D、ACL1、Roc5、RL14、BRD1、REL1、SRL1等。NRL1编码类纤维素合成蛋白酶, 其突变体的叶片近轴面泡状细胞变小, 导致叶片卷曲[7,8]。OsHox32与OsLBD3-7过表达时叶片内卷并变窄, 其泡状细胞数量减少[9,13]。ADL1定位在2号染色体上, 其突变体adl1叶片远轴面表皮细胞近轴化, 即在远轴面出现泡状细胞[10]。突变体ACL-D叶片因泡状细胞数目增加及排列异常而造成外卷并下垂[11], 其蛋白OsZHD1能结合OsDREB1B启动子[22]。ACL1基因突变后表现叶片外卷, 是由其泡状细胞变大和数目增加及表皮细胞扩大所引起[23]。Roc5负向调控泡状细胞的发育, 过表达Roc5时, 泡状细胞数量和大小都减少, 造成叶片内卷[24]。RL14编码2OG-Fe(II)氧化酶, 其突变体次生细胞壁组分减少, 从导管到叶片的水分运输速率降低, 导致泡状细胞失水, 表现出内卷[25]。BRD1基因除影响泡状细胞的数量外, 还影响叶片的其他细胞结构, 导致其突变体植株矮化且叶片严重卷曲[26]。REL1通过油菜素内酯信号的传导, 使叶片泡状细胞数目和大小改变, 进而调控叶片形态[27]。SRL1编码糖基磷脂酰肌醇蛋白, 能够特异性调控液泡H+-焦磷酸酶和H+-ATPase亚基编码基因的表达, 抑制液泡形成, 使泡状细胞数量减少; 其突变体泡状细胞数目增加, 致使叶片半卷[28]。前人报道结果与RT-PCR中基因表达量趋势一致, sol1叶片的泡状细胞体积略微减小, 数量显著增加, 泡状细胞下的叶肉细胞厚度略微变薄, 同时叶片气孔导度增大, 蒸腾速率升高, 这可能是叶片卷曲的主要因素之一。
SOL1基因被定位于6号染色体标记JY6-3和JY6-10之间165 kb的区间内。此前在6号染色体上已经报道了2个与水稻卷叶相关的基因sd-sl与RL13, 其中sd-sl调控的突变体是由辐射诱变粳稻9522所获得, 其主要表型为植株矮化, 其次还伴随叶片短且微卷。目标基因sd-sl位于水稻6号染色体PAC克隆AP003490与AP005619上118 kb的物理范围内[29]。与SOL1相似, RL13同样位于水稻6号染色体短臂SSR标记RM276附近。研究结果表明, rl13突变体叶绿素含量显著高于野生型, 叶肉细胞层数变薄; 并且在野生型细胞内的一个较大的泡状细胞, 在rl13细胞内有2个大小相近的泡状细胞, 而该突变有可能是导致rl13叶片筒状卷曲的主要原因[30]。从sol1的表型鉴定结果来看, 与rl13表现出一定的相似性, 叶肉细胞厚度变薄、泡状细胞变小等。但是sol1与rl13也存在显著不同, 即sol1的突出表型为叶片外卷(图1-H, 图4-B), 而rl13则表现为内卷; 此外, sol1细胞内较大的泡状细胞与WT相似(图4-C~F), 其外卷的原因有可能是因为表皮泡状细胞的显著增多, 也明显不同于rl13。从比较结果来看, sol1明显不同于sd-sl与rl13, 且其定位结果与这2个已经报道的卷叶相关基因不一致, 是一个新的调控水稻叶片卷曲的基因。对该基因的后续深入研究, 包括目标基因分离、功能验证、机制解析等, 有可能为水稻叶片发育等相关研究提供新的参考。
4 结论
通过EMS诱变籼稻西农1B获得半外卷突变体sol1, 从十叶期开始功能叶片呈现出轻微的卷曲, 且随生育进程卷曲程度加深, 叶片半外卷和披垂, 茎秆半矮化, 叶夹角显著大于野生型。叶片的泡状细胞略微减小但数量较野生型显著增加。sol1的蒸腾速率、胞间CO2浓度、气孔导度显著或极显著高于野生型。该性状受1对隐性核基因控制, SOL1为一新的基因, 被定位在6号染色体短臂165 kb区间内。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1146/annurev-arplant-042817-040422URLPMID:29553800 [本文引用: 1]

Shoot architecture is determined by the organization and activities of apical, axillary, intercalary, secondary, and inflorescence meristems and by the subsequent development of stems, leaves, shoot branches, and inflorescences. In this review, we discuss the unifying principles of hormonal and genetic control of shoot architecture including advances in our understanding of lateral branch outgrowth; control of stem elongation, thickness, and angle; and regulation of inflorescence development. We focus on recent progress made mainly in Arabidopsis thaliana, rice, pea, maize, and tomato, including the identification of new genes and mechanisms controlling shoot architecture. Key advances include elucidation of mechanisms by which strigolactones, auxins, and genes such as IDEAL PLANT ARCHITECTURE1 and TEOSINTE BRANCHED1 control shoot architecture. Knowledge now available provides a foundation for rational approaches to crop breeding and the generation of ideotypes with defined architectural features to improve performance and productivity.
DOI:10.1105/tpc.108.061457URLPMID:19304938 [本文引用: 4]
As an important agronomic trait, rice (Oryza sativa L.) leaf rolling has attracted much attention from plant biologists and breeders. Moderate leaf rolling increases the photosynthesis of cultivars and hence raises grain yield. However, the relevant molecular mechanism remains unclear. Here, we show the isolation and functional characterization of SHALLOT-LIKE1 (SLL1), a key gene controlling rice leaf rolling. sll1 mutant plants have extremely incurved leaves due to the defective development of sclerenchymatous cells on the abaxial side. Defective development can be functionally rescued by expression of SLL1. SLL1 is transcribed in various tissues and accumulates in the abaxial epidermis throughout leaf development. SLL1 encodes a SHAQKYF class MYB family transcription factor belonging to the KANADI family. SLL1 deficiency leads to defective programmed cell death of abaxial mesophyll cells and suppresses the development of abaxial features. By contrast, enhanced SLL1 expression stimulates phloem development on the abaxial side and suppresses bulliform cell and sclerenchyma development on the adaxial side. Additionally, SLL1 deficiency results in increased chlorophyll and photosynthesis. Our findings identify the role of SLL1 in the modulation of leaf abaxial cell development and in sustaining abaxial characteristics during leaf development. These results should facilitate attempts to use molecular breeding to increase the photosynthetic capacity of rice, as well as other crops, by modulating leaf development and rolling.
DOI:10.1111/nph.14142URLPMID:27545518 [本文引用: 1]
To understand the molecular mechanisms of rice aerial organ development, we identified a mutant gene that caused a significant decrease in the width of aerial organs, termed ABNORMAL VASCULAR BUNDLES (AVB). Histological analysis showed that the slender aerial organs were caused by cell number reduction. In avb, the number of vascular bundles in aerial organs was reduced, whereas the area of the vascular bundles was increased. Ploidy analysis and the in?situ expression patterns of histone H4 confirmed that cell proliferation was impaired during lateral primordia development, whereas procambium cells showed a greater ability to undergo cell division in avb. RNA sequencing (RNA-seq) showed that the development process was affected in avb. Map-based cloning and genetic complementation demonstrated that AVB encodes a land plant conserved protein with unknown functions. Our research shows that AVB is involved in the maintenance of the normal cell division pattern in lateral primordia development and that the AVB gene is required for procambium establishment following auxin signaling.
DOI:10.1104/pp.108.118679URLPMID:18567825 [本文引用: 1]
Members of the Class III homeodomain leucine zipper (Class III HD-Zip) gene family are central regulators of crucial aspects of plant development. To better understand the roles of five Class III HD-Zip genes in rice (Oryza sativa) development, we investigated their expression patterns, ectopic expression phenotypes, and auxin responsiveness. Four genes, OSHB1 to OSHB4, were expressed in a localized domain of the shoot apical meristem (SAM), the adaxial cells of leaf primordia, the leaf margins, and the xylem tissue of vascular bundles. In contrast, expression of OSHB5 was observed only in phloem tissue. Plants ectopically expressing microRNA166-resistant versions of the OSHB3 gene exhibited severe defects, including the ectopic production of leaf margins, shoots, and radialized leaves. The treatment of seedlings with auxin quickly induced ectopic OSHB3 expression in the entire region of the SAM, but not in other tissues. Furthermore, this ectopic expression of OSHB3 was correlated with leaf initiation defects. Our findings suggest that rice Class III HD-Zip genes have conserved functions with their homologs in Arabidopsis (Arabidopsis thaliana), but have also acquired specific developmental roles in grasses or monocots. In addition, some Class III HD-Zip genes may regulate the leaf initiation process in the SAM in an auxin-dependent manner.
DOI:10.1105/tpc.106.047472URLPMID:17307928 [本文引用: 1]
Embryo patterning in Arabidopsis thaliana is highly affected when KANADI or Class III HD-Zip genes are compromised. Triple loss-of-function kan1 kan2 kan4 embryos exhibit striking defects in the peripheral-central axis, developing lateral leaf-like organs from the hypocotyls, whereas loss of Class III HD-Zip gene activity results in a loss of bilateral symmetry. Loss of KANADI activity in a Class III HD-Zip mutant background mitigates the defects in bilateral symmetry, implying that the two gene families act antagonistically during embryonic pattern formation. Dynamic patterns of auxin concentration and flux contribute to embryo patterning. Polar cellular distribution of PIN-FORMED1 (PIN1) mediates auxin flow throughout embryogenesis and is required for establishment of the apical-basal axis and bilateral symmetry. Defects in the pattern of PIN1 expression are evident when members of either the KANADI or Class III HD-Zip gene families are compromised. Abnormal expression patterns of PIN1 in KANADI or Class III HD-Zip multiple mutants and the phenotype of plants in which members of both gene families are mutated suggest that pattern formation along the central-peripheral axis results from interplay between auxin and the KANADI and Class III HD-Zip transcription factors, whose defined spatial and temporal expression patterns may also be influenced by auxin.
DOI:10.1007/s00425-006-0472-0URL [本文引用: 1]
High-yield cultivars are characterized by erect leaf canopies that optimize photosynthesis and thus favor increased biomass. Upward curling of the leaf blade (called rolled leaf) can result in enhanced erect-leaf habit, increase erect duration and promote an overall erect leaf canopy. The rice mutant R05, induced through transferred DNA (T-DNA) insertion, had the rolled-leaf trait. The leaves in the wild type demonstrated natural drooping tendencies, resulting in decreasing leaf erection indices (LEIs) during senescence at the 20th day after flowering. Conversely, LEIs of the leaves in R05 remained high, even 20-day post-flowering. We applied T-DNA tagging and isolated a rolled-leaf gene from rice which, when over-expressed, could induce upward curling of the leaf blade. This gene encodes for a protein of 1,048 amino acids including the PAZ and PIWI conserved domains, belonging to the Argonaute (AGO) family. There are at least 18 members of the AGO family in rice. According to high-sequence conservation, the rolled-leaf gene in rice could be orthologous to the Arabidopsis ZIP/Ago7 gene, so we called it OsAGO7. These results provide a possible opportunity for implementing OsAGO7 gene in crop improvement.
DOI:10.1007/s11103-010-9614-7URL [本文引用: 2]
Leaf morphology is an important agronomic trait in rice breeding. We isolated three allelic mutants of NARROW AND ROLLED LEAF 1 (nrl1) which showed phenotypes of reduced leaf width and semi-rolled leaves and different degrees of dwarfism. Microscopic analysis indicated that the nrl1-1 mutant had fewer longitudinal veins and smaller adaxial bulliform cells compared with the wild-type. The NRL1 gene was mapped to the chromosome 12 and encodes the cellulose synthase-like protein D4 (OsCslD4). Sequence analyses revealed single base substitutions in the three allelic mutants. Genetic complementation and over-expression of the OsCslD4 gene confirmed the identity of NRL1. The gene was expressed in all tested organs of rice at the heading stage and expression level was higher in vigorously growing organs, such as roots, sheaths and panicles than in elsewhere. In the mutant leaves, however, the expression level was lower than that in the wild-type. We conclude that OsCslD4 encoded by NRL1 plays a critical role in leaf morphogenesis and vegetative development in rice.
DOI:10.1007/s00425-010-1180-3URL [本文引用: 2]
Appropriate leaf shape has proved to be useful in improving photosynthesis and increasing grain yield. To understand the molecular mechanism of leaf morphogenesis, we identified a rice mutant nrl1, which was characterized by a phenotype of narrow and rolled leaves. Microscopic observation showed that the mutation significantly decreased the number of vascular bundles of leaf and stem. Genetic analysis revealed that the mutation was controlled by a single nuclear-encoded recessive gene. To isolate the nrl1 gene, 756 F2 and F3 mutant individuals from a cross of the nrl1 mutant with Longtepu were used and a high-resolution physical map of the chromosomal region around the nrl1 gene was made. Finally, the gene was mapped in 16.5kb region between marker RL21 and marker RL36 within the BAC clone OSJNBa0027H05. Cloning and sequencing of the target region from the mutant showed that there was a 58bp deletion within the second exon of the cellulose synthase-like D4 gene (TIGR locus Os12g36890). The nrl1 mutation was rescued by transformation with the wild-type cellulose synthase-like D4 gene. Accordingly, the cellulose synthase-like D4 gene was identified as the NRL1 gene. NRL1 was transcribed in various tissues and was mainly expressed in panicles and internodes. NAL7 and SLL1 were found to be upregulated, whereas OsAGO7 were downregulated in the nrl1 mutant. These findings suggested that there might be a functional association between these genes in regulating leaf development.
DOI:10.1186/s12284-016-0118-1URLPMID:27624698 [本文引用: 2]
The Class III homeodomain Leu zipper (HD-Zip III) gene family plays important roles in plant growth and development. Here, we analyze the function of OsHox32, an HD-Zip III family member, and show that it exhibits pleiotropic effects on regulating plant type architecture and leaf development in rice.
DOI:10.1016/j.ydbio.2009.07.042URLPMID:19665012 [本文引用: 2]
The adaxial-abaxial axis in leaf primordia is thought to be established first and is necessary for the expansion of the leaf lamina along the mediolateral axis. To understand axis information in leaf development, we isolated the adaxialized leaf1 (adl1) mutant in rice, which forms abaxially rolled leaves. adl1 leaves are covered with bulliform-like cells, which are normally distributed only on the adaxial surface. An adl1 double mutant with the adaxially snowy leaf mutant, which has albino cells that specifically appear in the abaxial mesophyll tissue, indicated that adl1 leaves show adaxialization in both epidermal and mesophyll tissues. The expression of HD-ZIPIII genes in adl1 mutant increased in mature leaves, but not in the young primordia or the SAM. This indicated that ADL1 may not be directly involved in determining initial leaf polarity, but rather is associated with the maintenance of axis information. ADL1 encodes a plant-specific calpain-like cysteine proteinase orthologous to maize DEFECTIVE KERNEL1. Furthermore, we identified intermediate and strong alleles of the adl1 mutant that generate shootless embryos and globular-arrested embryos with aleurone layer loss, respectively. We propose that ADL1 plays an important role in pattern formation of the leaf and embryo by promoting proper epidermal development.
DOI:10.1007/s00425-013-2009-7URL [本文引用: 2]
Leaf rolling is receiving considerable attention as an important agronomic trait in rice (Oryza sativa L.). However, little has been known on the molecular mechanism of rice leaf rolling, especially the abaxial rolling. We identified a novel abaxially curled and drooping leaf-dominant mutant from a T-1 transgenic rice line. The abaxially curled leaf phenotypes, co-segregating with the inserted transferred DNA, were caused by overexpression of a zinc finger homeodomain class homeobox transcription factor (OsZHD1). OsZHD1 exhibited a constitutive expression pattern in wild-type plants and accumulated in the developing leaves and panicles. Artificial overexpression of OsZHD1 or its closest homolog OsZHD2 induced the abaxial leaf curling. Histological analysis indicated that both the increased number and the abnormal arrangement of bulliform cells in leaf were responsible for the abaxially curled leaves. We herein reported OsZHD1 with key roles in rice morphogenesis, especially in the modulating of leaf rolling, which provided a novel insight into the molecular mechanism of leaf development in rice.
DOI:10.1007/s10725-015-0097-zURL [本文引用: 1]
DOI:10.1371/journal.pone.0156413URLPMID:27258066 [本文引用: 2]
Appropriate leaf rolling enhances erect-leaf habits and photosynthetic efficiency, which consequently improves grain yield. Here, we reported the novel lateral organ boundaries domain (LBD) gene OsLBD3-7, which is involved in the regulation of leaf rolling. OsLBD3-7 works as a transcription activator and its protein is located on the plasma membrane and in the nucleus. Overexpression of OsLBD3-7 leads to narrow and adaxially rolled leaves. Microscopy of flag leaf cross-sections indicated that overexpression of OsLBD3-7 led to a decrease in both bulliform cell size and number. Transcriptional analysis showed that key genes that had been reported to be negative regulators of bulliform cell development were up-regulated in transgenic plants. These results indicated that OsLBD3-7 might acts as an upstream regulatory gene of bulliform cell development to regulate leaf rolling, which will give more insights on the leaf rolling regulation mechanism.
DOI:10.1007/s11103-008-9365-xURL [本文引用: 1]
Leaves, the collective organ produced by the shoot apical meristem (SAM), are polarized along their adaxial–abaxial axis. In this study, we characterized two rice (Oryza sativa) allelic rolled-leaf mutants, rolled leaf 9-1 (rl9-1) and rl9-2, which display very similar phenotypes with completely adaxialized leaves and malformed spikelets. We cloned the RL9 gene by way of a map-based cloning strategy. Molecular studies have revealed that RL9 encodes a GARP protein, an orthologue of Arabidopsis KANADIs. RL9 is mainly expressed in roots, leaves, and flowers. The transient expression of a RL9–GFP (green fluorescent protein) fusion protein has indicated that RL9 protein is localized in the nucleus, suggesting that RL9 acts as a putative transcription factor.
DOI:10.1105/tpc.111.088625URL [本文引用: 1]
Plants have a chemically heterogeneous lipophilic layer, the cuticle, which protects them from biotic and abiotic stresses. The mechanisms that regulate cuticle development are poorly understood. We identified a rice (Oryza sativa) dominant curly leaf mutant, curly flag leaf1 (cfl1), and cloned CFL1, which encodes a WW domain protein. We overexpressed both rice and Arabidopsis CFL1 in Arabidopsis thaliana; these transgenic plants showed severely impaired cuticle development, similar to that in cfl1 rice. Reduced expression of At CFL1 resulted in reinforcement of cuticle structure. At CFL1 was predominantly expressed in specialized epidermal cells and in regions where dehiscence and abscission occur. Biochemical evidence showed that At CFL1 interacts with HDG1, a class IV homeodomain-leucine zipper transcription factor. Suppression of HDG1 function resulted in similar defective cuticle phenotypes in wild-type Arabidopsis but much alleviated phenotypes in At cfl1-1 mutants. The expression of two cuticle development-associated genes, BDG and FDH, was downregulated in At CFL1 overexpressor and HDG1 suppression plants. HDG1 binds to the cis-element L1 box, which exists in the regulatory regions of BDG and FDH. Our results suggest that rice and Arabidopsis CFL1 negatively regulate cuticle development by affecting the function of HDG1, which regulates the downstream genes BDG and FDH.
[本文引用: 1]
[本文引用: 1]
DOI:10.1104/pp.112.200980URLPMID:22891238 [本文引用: 1]
The control of floral organ identity by homeotic MADS box genes is well established in eudicots. However, grasses have highly specialized outer floral organs, and the identities of the genes that regulate the highly specialized outer floral organs of grasses remain unclear. In this study, we characterized a MIKC-type MADS box gene, CHIMERIC FLORAL ORGANS (CFO1), which plays a key role in the regulation of floral organ identity in rice (Oryza sativa). The cfo1 mutant displayed defective marginal regions of the palea, chimeric floral organs, and ectopic floral organs. Map-based cloning demonstrated that CFO1 encoded the OsMADS32 protein. Phylogenetic analysis revealed that CFO1/OsMADS32 belonged to a monocot-specific clade in the MIKC-type MADS box gene family. The expression domains of CFO1 were mainly restricted to the marginal region of the palea and inner floral organs. The floral organ identity gene DROOPING LEAF (DL) was expressed ectopically in all defective organs of cfo1 flowers. Double mutant analysis revealed that loss of DL function mitigated some of the defects of floral organs in cfo1 flowers. We propose that the CFO1 gene plays a pivotal role in maintaining floral organ identity through negative regulation of DL expression.
DOI:10.1073/pnas.88.21.9828URLPMID:1682921 [本文引用: 1]
We developed bulked segregant analysis as a method for rapidly identifying markers linked to any specific gene or genomic region. Two bulked DNA samples are generated from a segregating population from a single cross. Each pool, or bulk, contains individuals that are identical for a particular trait or genomic region but arbitrary at all unlinked regions. The two bulks are therefore genetically dissimilar in the selected region but seemingly heterozygous at all other regions. The two bulks can be made for any genomic region and from any segregating population. The bulks are screened for differences using restriction fragment length polymorphism probes or random amplified polymorphic DNA primers. We have used bulked segregant analysis to identify three random amplified polymorphic DNA markers in lettuce linked to a gene for resistance to downy mildew. We showed that markers can be reliably identified in a 25-centimorgan window on either side of the targeted locus. Bulked segregant analysis has several advantages over the use of near-isogenic lines to identify markers in specific regions of the genome. Genetic walking will be possible by multiple rounds of bulked segregation analysis; each new pair of bulks will differ at a locus identified in the previous round of analysis. This approach will have widespread application both in those species where selfing is possible and in those that are obligatorily outbreeding.
DOI:10.1104/pp.007179URLPMID:12427982 [本文引用: 1]
We have isolated a new recessive dwarf mutant of rice (Oryza sativa L. cv Nipponbare). Under normal growth conditions, the mutant has very short leaf sheaths; has short, curled, and frizzled leaf blades; has few tillers; and is sterile. Longitudinal sections of the leaf sheaths revealed that the cell length along the longitudinal axis is reduced, which explains the short leaf sheaths. Transverse sections of the leaf blades revealed enlargement of the motor cells along the dorsal-ventral axis, which explains the curled and frizzled leaf blades. In addition, the number of crown roots was smaller and the growth of branch roots was weaker than those in the wild-type plant. Because exogenously supplied brassinolide considerably restored the normal phenotypes, we designated the mutant brassinosteroid-dependent 1 (brd1). Further, under darkness, brd1 showed constitutive photomorphogenesis. Quantitative analyses of endogenous sterols and brassinosteroids (BRs) indicated that BR-6-oxidase, a BR biosynthesis enzyme, would be defective. In fact, a 0.2-kb deletion was detected in the genomic region of OsBR6ox (a rice BR-6-oxidase gene) in the brd1 mutant. These results indicate that BRs are involved in many morphological and physiological processes in rice, including the elongation and unrolling of leaves, development of tillers, skotomorphogenesis, root differentiation, and reproductive growth, and that the defect of BR-6-oxidase caused the brd1 phenotype.
DOI:10.1016/j.pbi.2006.11.004URLPMID:17140837 [本文引用: 1]
Plants have evolved epidermal cells that have specialized functions as adaptations to life on land. Many of the functions of these specialized cells are dependent, to a significant extent, on their arrangement within the aerial epidermis. Considerable progress has been made over the past two years in understanding the patterning mechanisms of trichomes and stomata in Arabidopsis leaves at the molecular level. How universal are these patterning programmes, and how are they adjusted to meet the changing functions of specialized epidermal cells in different plant organs? In this review, we compare the patterning of stomata and trichomes in different plant species, describe environmental and developmental factors that alter cell patterning, and discuss how changes in patterning might relate to cell function. Patterning is an important aspect to the functioning of aerial epidermal cells, and a greater understanding of the processes that are involved will significantly enhance our understanding of how cellular activities are integrated in multicellular plants.
DOI:10.3724/SP.J.1006.2013.00767URL [本文引用: 1]
叶片形态是水稻“理想株型”的重要组成部分,是当前水稻高产育种关注的重点。本文通过对已克隆多个叶形相关调控基因综述了水稻叶片形态(叶片卷曲度、倾角、披散程度以及叶片宽度)建成的分子遗传学研究进展。综合分析认为,水稻叶片的卷曲主要是通过卷叶基因调控叶片近轴/远轴间的发育、泡状细胞的发育及其膨胀和渗透压、厚壁组织的形成以及叶片角质层的发育等来实现。影响植株空间伸展姿态的叶倾角主要通过叶角基因调控油菜素内酯的信号传导来影响叶枕细胞的生长发育;唯一被克隆的影响叶片披垂度的披叶基因DL1是通过控制叶片中脉发育而改变叶片形态的;而窄叶基因则主要通过调控生长素的合成与极性运输、维管组织的发育和分布,影响叶片维管束数目及宽度。但到目前为止,所有已克隆的叶形调控基因间相互调控关系的研究还不够深入,还不能完整清晰地勾勒水稻叶形建成和发育的分子调控网络。因此,在已有的研究基础上更深入地探索水稻叶片形态建成的分子调控机制,对进一步构建相关的调控网络,塑造水稻理想株型具有重要意义。
DOI:10.3724/SP.J.1006.2013.00767URL [本文引用: 1]
叶片形态是水稻“理想株型”的重要组成部分,是当前水稻高产育种关注的重点。本文通过对已克隆多个叶形相关调控基因综述了水稻叶片形态(叶片卷曲度、倾角、披散程度以及叶片宽度)建成的分子遗传学研究进展。综合分析认为,水稻叶片的卷曲主要是通过卷叶基因调控叶片近轴/远轴间的发育、泡状细胞的发育及其膨胀和渗透压、厚壁组织的形成以及叶片角质层的发育等来实现。影响植株空间伸展姿态的叶倾角主要通过叶角基因调控油菜素内酯的信号传导来影响叶枕细胞的生长发育;唯一被克隆的影响叶片披垂度的披叶基因DL1是通过控制叶片中脉发育而改变叶片形态的;而窄叶基因则主要通过调控生长素的合成与极性运输、维管组织的发育和分布,影响叶片维管束数目及宽度。但到目前为止,所有已克隆的叶形调控基因间相互调控关系的研究还不够深入,还不能完整清晰地勾勒水稻叶形建成和发育的分子调控网络。因此,在已有的研究基础上更深入地探索水稻叶片形态建成的分子调控机制,对进一步构建相关的调控网络,塑造水稻理想株型具有重要意义。
DOI:10.1093/jxb/ers035URL [本文引用: 1]
Plants have evolved several mechanisms in order to cope with adverse environmental conditions. The transcription factors (TFs) belonging to the DREB1/CBF subfamily have been described as major regulators of the plant responses to different abiotic stresses. This study focused on the rice gene OsDREB1B, initially described as highly and specifically induced by cold. However, here it is shown that OsDREB1B is not only induced by low temperatures, but also by drought and mechanical stress. In order to identify novel TFs that bind to its promoter, a yeast one-hybrid system was used to screen a cold-induced cDNA expression library. Thereby seven novel Zn-finger TFs were identified that bind to the promoter of OsDREB1B. Among them, there were four Zn-finger homeodomain (ZF-HD) and three C2H2-type Zn-finger TFs. Gene expression studies showed that these TFs are differentially regulated at transcriptional level by different abiotic stress conditions, which is illustrative of the crosstalk between stress signalling pathways. Protein-protein interaction studies revealed the formation of homo- and heterodimers among the ZF-HD TFs identified, but not for the C2H2-type. Using a transactivation assay in Arabidopsis protoplasts, all the TFs identified repressed the expression of the reporter gene, driven by the promoter of OsDREB1B. This assay also showed that the dimerization observed between the ZF-HD TFs may play a role on their transactivation activity. The results here presented suggest a prominent role of Zn-finger TFs in the regulation of OsDREB1B.
DOI:10.1093/mp/ssq022URLPMID:20494951 [本文引用: 1]
Understanding the genetic mechanism underlying rice leaf-shape development is crucial for optimizing rice configuration and achieving high yields; however, little is known about leaf abaxial curling. We isolated a rice transferred DNA (T-DNA) insertion mutant, BY240, which exhibited an abaxial leaf curling phenotype that co-segregated with the inserted T-DNA. The T-DNA was inserted in the promoter of a novel gene, ACL1 (Abaxially Curled Leaf 1), and led to overexpression of this gene in BY240. Overexpression of ACL1 in wild-type rice also resulted in abaxial leaf curling. ACL1 encodes a protein of 116 amino acids with no known conserved functional domains. Overexpression of ACL2, the only homolog of ACL1 in rice, also induced abaxial leaf curling. RT-PCR analysis revealed high expressions of ACLs in leaf sheaths and leaf blades, suggesting a role for these genes in leaf development. In situ hybridization revealed non-tissue-specific expression of the ACLs in the shoot apical meristem, leaf primordium, and young leaf. Histological analysis showed increased number and exaggeration of bulliform cells and expansion of epidermal cells in the leaves of BY240, which caused developmental discoordination of the abaxial and adaxial sides, resulting in abaxially curled leaves. These results revealed an important mechanism in rice leaf development and provided the genetic basis for agricultural improvement.
DOI:10.1104/pp.111.176016URL [本文引用: 1]
Leaf rolling is considered an important agronomic trait in rice (Oryza sativa) breeding. To understand the molecular mechanism controlling leaf rolling, we screened a rice T-DNA insertion population and isolated the outcurved leaf1 (oul1) mutant showing abaxial leaf rolling. The phenotypes were caused by knockout of Rice outermost cell-specific gene5 (Roc5), an ortholog of the Arabidopsis (Arabidopsis thaliana) homeodomain leucine zipper class IV gene GLABRA2. Interestingly, overexpression of Roc5 led to adaxially rolled leaves, whereas cosuppression of Roc5 resulted in abaxial leaf rolling. Bulliform cell number and size increased in oul1 and Roc5 cosuppression plants but were reduced in Roc5-overexpressing lines. The data indicate that Roc5 negatively regulates bulliform cell fate and development. Gene expression profiling, quantitative polymerase chain reaction, and RNA interference (RNAi) analyses revealed that Protodermal Factor Like (PFL) was probably down-regulated in oul1. The mRNA level of PFL was increased in Roc5-overexpressing lines, and PFL-RNAi transgenic plants exhibit reversely rolling leaves by reason of increases of bulliform cell number and size, indicating that Roc5 may have a conserved function. These are, to our knowledge, the first functional data for a gene encoding a homeodomain leucine zipper class IV transcriptional factor in rice that modulates leaf rolling.
DOI:10.1111/j.1467-7652.2012.00679.xURL [本文引用: 1]
As an important agronomic trait, leaf rolling in rice (Oryza sativa L.) has attracted much attention from plant biologists and breeders. Moderate leaf rolling increases the amount of photosynthesis in cultivars and hence raises grain yield. Here, we describe the map-based cloning of the gene RL14, which was found to encode a 2OG-Fe (II) oxygenase of unknown function. rl14 mutant plants had incurved leaves because of the shrinkage of bulliform cells on the adaxial side. In addition, rl14 mutant plants displayed smaller stomatal complexes and decreased transpiration rates, as compared with the wild type. Defective development could be rescued functionally by the expression of wild-type RL14. RL14 was transcribed in sclerenchymatous cells in leaves that remained wrapped inside the sheath. In mature leaves, RL14 accumulated mainly in the mesophyll cells that surround the vasculature. Expression of genes related to secondary cell wall formation was affected in rl14-1 mutants, and cellulose and lignin content were altered in rl14-1 leaves. These results reveal that the RL14 gene affects water transport in leaves by affecting the composition of the secondary cell wall. This change in water transport results in water deficiency, which is the major reason for the abnormal shape of the bulliform cells.
DOI:10.1046/j.1365-313x.2002.01438.xURLPMID:12445121 [本文引用: 1]
Molecular genetic and physiological studies on brassinosteroid (BR)-related mutants of dicot plants have revealed that BRs play important roles in normal plant growth and development. However, little is known about the function of BR in monocots (grasses), except for the phenotypic analysis of a rice mutant partially insensitive to BR signaling. To investigate the function of BR in monocots, we identified and characterized BR-deficient mutants of rice, BR-deficient dwarf1 (brd1). The brd1 mutants showed a range of abnormalities in organ development and growth, the most striking of which were defects in the elongation of the stem and leaves. Light microscopic observations revealed that this abnormality was primarily owing to a failure in the organization and polar elongation of the leaf and stem cells. The accumulation profile of BR compounds in the brd1 mutants suggested that these plants may be deficient in the activity of BR C-6 oxidase. Therefore, we cloned a rice gene, OsDWARF, which has a high sequence similarity to the tomato C-6 oxidase gene, DWARF. Introduction of the wild-type OsDWARF gene into brd1 rescued the abnormal phenotype of the mutants. The OsDWARF gene was expressed at a low level in all of the examined tissues, with preferential expression in the leaf sheath, and the expression was negatively regulated by brassinolide treatment. On the basis of these findings, we discuss the biological function of BRs in rice plants.
DOI:10.1093/jxb/erv319URLPMID:26142419 [本文引用: 1]
Leaf morphology, particularly in crop, is one of the most important agronomic traits because it influences the yield through the manipulation of photosynthetic capacity and transpiration. To understand the regulatory mechanism of leaf morphogenesis, an Oryza sativa dominant mutant, rolled and erect leaf 1 (rel1) has been characterized. This mutant has a predominant rolled leaf, increased leaf angle, and reduced plant height phenotype that results in a reduction in grain yield. Electron microscope observations indicated that the leaf incurvations of rel1 dominant mutants result from the alteration of the size and number of bulliform cells. Molecular cloning revealed that the rel1 dominant mutant phenotype is caused by the activation of the REL1 gene, which encodes a novel unknown protein, despite its high degree of conservation among monocot plants. Moreover, the downregulation of the REL1 gene in the rel1 dominant mutant restored the phenotype of this dominant mutant. Alternatively, overexpression of REL1 in wild-type plants induced a phenotype similar to that of the dominant rel1 mutant, indicating that REL1 plays a positive role in leaf rolling and bending. Consistent with the observed rel1 phenotype, the REL1 gene was predominantly expressed in the meristem of various tissues during plant growth and development. Nevertheless, the responsiveness of both rel1 dominant mutants and REL1-overexpressing plants to exogenous brassinosteroid (BR) was reduced. Moreover, transcript levels of BR response genes in the rel1 dominant mutants and REL1-overexpressing lines were significantly altered. Additionally, seven REL1-interacting proteins were also identified from a yeast two-hybrid screen. Taken together, these findings suggest that REL1 regulates leaf morphology, particularly in leaf rolling and bending, through the coordination of BR signalling transduction.
DOI:10.1104/pp.112.199968URL [本文引用: 1]
Leaf rolling is an important agronomic trait in rice (Oryza sativa) breeding and moderate leaf rolling maintains the erectness of leaves and minimizes shadowing between leaves, leading to improved photosynthetic efficiency and grain yields. Although a few rolled-leaf mutants have been identified and some genes controlling leaf rolling have been isolated, the molecular mechanisms of leaf rolling still need to be elucidated. Here we report the isolation and characterization of SEMI-ROLLED LEAF1 (SRL1), a gene involved in the regulation of leaf rolling. Mutants srl1-1 (point mutation) and srl1-2 (transferred DNA insertion) exhibit adaxially rolled leaves due to the increased numbers of bulliform cells at the adaxial cell layers, which could be rescued by complementary expression of SRL1. SRL1 is expressed in various tissues and is expressed at low levels in bulliform cells. SRL1 protein is located at the plasma membrane and predicted to be a putative glycosylphosphatidylinositol-anchored protein. Moreover, analysis of the gene expression profile of cells that will become epidermal cells in wild type but probably bulliform cells in srl1-1 by laser-captured microdissection revealed that the expression of genes encoding vacuolar H+-ATPase (subunits A, B, C, and D) and H+-pyrophosphatase, which are increased during the formation of bulliform cells, were up-regulated in srl1-1. These results provide the transcript profile of rice leaf cells that will become bulliform cells and demonstrate that SRL1 regulates leaf rolling through inhibiting the formation of bulliform cells by negatively regulating the expression of genes encoding vacuolar H+-ATPase subunits and H+-pyrophosphatase, which will help to understand the mechanism regulating leaf rolling.
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2012.00423URL [本文引用: 1]
叶片形态是理想株型的重要指标之一,叶片适度卷曲有利于理想株型的建成,是水稻超高产育种的重要材料。在EMS诱变籼稻缙恢10号群体中发现一个卷叶突变体,表现叶片筒状卷曲,经过多代连续自交,性状稳定,命名为rl13 (rolled leaf 13)。rl13的叶绿素a、叶绿素b和总叶绿素含量均显著高于野生型对照缙恢10号,类胡萝卜素含量在苗期、孕穗期与野生型相比有显著提高,而抽穗期和成熟期则差异不显著。rl13的三片功能叶的卷曲度与野生型相比均达到极显著差异,但rl13的三片功能叶之间差异不显著。通过石蜡切片分析,突变体叶肉细胞层数变薄,野生型含有的一个较大泡状细胞转变为卷叶突变体的两个大小相近的泡状细胞,导致了叶片弯曲。以该突变体为父本,西农1A为母本配制杂交组合构建F2遗传群体,结果表明,该卷叶性状由一对隐性核基因控制。选用F2代分离群体中的1 215个隐性单株作为定位群体,将RL13定位在第6染色体短臂上分子标记RM276和SWU6-1之间,遗传距离分别为1.1 cM和0.2 cM。
DOI:10.3724/SP.J.1006.2012.00423URL [本文引用: 1]
叶片形态是理想株型的重要指标之一,叶片适度卷曲有利于理想株型的建成,是水稻超高产育种的重要材料。在EMS诱变籼稻缙恢10号群体中发现一个卷叶突变体,表现叶片筒状卷曲,经过多代连续自交,性状稳定,命名为rl13 (rolled leaf 13)。rl13的叶绿素a、叶绿素b和总叶绿素含量均显著高于野生型对照缙恢10号,类胡萝卜素含量在苗期、孕穗期与野生型相比有显著提高,而抽穗期和成熟期则差异不显著。rl13的三片功能叶的卷曲度与野生型相比均达到极显著差异,但rl13的三片功能叶之间差异不显著。通过石蜡切片分析,突变体叶肉细胞层数变薄,野生型含有的一个较大泡状细胞转变为卷叶突变体的两个大小相近的泡状细胞,导致了叶片弯曲。以该突变体为父本,西农1A为母本配制杂交组合构建F2遗传群体,结果表明,该卷叶性状由一对隐性核基因控制。选用F2代分离群体中的1 215个隐性单株作为定位群体,将RL13定位在第6染色体短臂上分子标记RM276和SWU6-1之间,遗传距离分别为1.1 cM和0.2 cM。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}