Analysis of drought resistance and DNA methylation level of resynthesized Brassica napus
YUAN Yi, ZHU Shuang, FANG Ting-Ting, JIANG Jin-Jin,*, WANG You-Ping*College of Bioscience and Biotechnology, Yangzhou University, Yangzhou 225009, Jiangsu, China
This study was supported by the National Natural Science Foundation of China.31771824 This study was supported by the National Natural Science Foundation of China.31771825 China and Jiangsu Postdoctoral Science Foundation.2014M561719 China and Jiangsu Postdoctoral Science Foundation.2015T80591 China and Jiangsu Postdoctoral Science Foundation.1401078B
Abstract Brassica napus, as one of the important resources of edible plant oil and forage protein, is a polyploid species with great economic value. However, it is sensitive to drought stress throughout whole lifecycle due to short domestication history and narrow genetic background. Thus, it is a main purpose to breed B. napus cultivar with high yield and drought resistance. In the present study, we compared the drought resistance among S1-S4 generations of resynthesized B. napus and diploid parents under different time periods of 15% PEG-6000 treatment. The different drought tolerance levels were assessed based on phenotype observation, leaf physiological indexes (MDA, soluble protein, SOD and POD). Accompanying with water content analysis, we found the drought tolerance showed a trend of B. oleracea > Bn-S3 > Bn-S4 > Bn-S1 > Bn-S2 > B. rapa. Under drought stress, POD and SOD activities in Bn-S3 and Bn-S4 were higher than these in other plants tested, and MDA content was decreased, indicating that Bn-S3 and Bn-S4 have better ability in clearing ROS, and defending from peroxidation damage. On the basis of HPLC analysis, the methylation level in all materials was the highest under drought stress of 12 h. And the methylation level in B. rapa was higher than that in others, that in Bn-S1 and Bn-S4 was between that in parents, while that in Bn-S2 and Bn-S3 was lower than that in parents. Methylation sensitive amplification polymorphism analysis also revealed multiple changes in methylation and demethylation level of resynthesized B. napus under drought stress, indicating methylation changes might be involved in plant drought tolerance. Keywords:resynthesized Brassica napus; B. rapa; B. oleracea;drought stress;DNA methylation
PDF (1343KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 袁溢, 朱双, 方婷婷, 蒋金金, 王幼平. 人工合成甘蓝型油菜抗旱性及DNA甲基化水平分析[J]. 作物学报, 2019, 45(5): 693-704. doi:10.3724/SP.J.1006.2019.84120 YUAN Yi, ZHU Shuang, FANG Ting-Ting, JIANG Jin-Jin, WANG You-Ping. Analysis of drought resistance and DNA methylation level of resynthesized Brassica napus[J]. Acta Agronomica Sinica, 2019, 45(5): 693-704. doi:10.3724/SP.J.1006.2019.84120
CK: 对照; DT: 干旱胁迫; Br: 白菜型油菜; Bo: 甘蓝; S1~S4: 人工合成甘蓝型油菜自交后代。 Fig. 2Relative water content and relative water retention in plants under drought stress of 12 h
CK: control; DT: drought stress; Br: B. rapa; Bo: B. oleracea; S1-S4: progenies of resynthesized B. napus.
A: 过氧化物酶活力; B: 丙二醛含量; C: 超氧化物歧化酶活力; D: 可溶性蛋白含量。平均值±标准误, n = 3。Br: 白菜型油菜; Bo: 甘蓝; S1~S4: 人工合成甘蓝型油菜自交后代。 Fig. 3Physiological indexes of plants under drought stress
A: POD activity; B: MDA content; C: SOD activity; D: soluble protein. Average value ± SD, n = 3. Br: B. rapa; Bo: B. oleracea; S1-S4: progenies of resynthesized B. napus.
Wang HZ, YinY . Analysis and strategy for oil crop industry in China Chin J Oil Crop Sci, 2014,36:414-421 (in Chinese with English abstract). [本文引用: 1]
ZhuM, Assmann SM . Metabolic signatures in response to abscisic acid (ABA) treatment in Brassica napus guard cells revealed by metabolomics Sci Rep, 2017,7:12875-12890. [本文引用: 1]
TardieuF . Plant response to environmental conditions: assessing potential production, water demand, and negative effects of water deficit Front Physiol, 2013,4:17-27. [本文引用: 1]
Ji RP, Che YS, Zhu YN, LiangT, FengR, Yu WY, Zhang YS . Impacts of drought stress on the growth and development and grain yield of spring maize in northeast China Chin J Appl Ecol, 2012,23:3021-3026 (in Chinese with English abstract). [本文引用: 1]
Luo LJ . Breeding for water-saving and drought-resistance rice (WDR) in China J Exp Bot, 2010,61:3509-3517. DOI:10.1093/jxb/erq185URL [本文引用: 1]
Fang YJ, Xiong LZ . General mechanisms of drought response and their application in drought resistance improvement in plants Cell Mol Life Sci, 2015,72:673-689. DOI:10.1007/s00018-014-1767-0URL [本文引用: 1]
Jiao JZ, Xie LL, Li XL, Tian ZH, Xu BB . Drought tolerance in rapeseed ( Brassica napus L.) oil Hubei Agric Sci, 2015,54:773-777 (in Chinese with English abstract). [本文引用: 1]
Mailer RJ, Cornish PS . Effects of water stress on glucosinolate and oil concentrations in the seeds of rape ( Brassica napus L.) and turnip rape( Brassica rapa L. var. silvestris [Lam.] Briggs) Aust J Exp Agric, 1987,27:707-711. [本文引用: 2]
Tesfamariam EH, Annandale JG, Steyn JM . Water stress effects on winter canola growth and yield Agron J, 2010,102:658-666. DOI:10.2134/agronj2008.0043URL [本文引用: 1]
MaQ, Turner DW . Osmotic adjustment segregates with and is positively related to seed yield in F3 lines of crosses between Brassica napus and B. juncea subjected to water deficit . Aust J Exp Agric, 2006,46:1621-1627. [本文引用: 1]
DanquahA, Zelicourt AD, ColcombetJ, HirtH . The role of ABA and MAPK signaling pathways in plant abiotic stress responses Biotechnol Adv, 2014,32:40-52. DOI:10.1016/j.biotechadv.2013.09.006URL [本文引用: 1]
BalestriniS, VartanianN . Rhizogenic activity during water stress-induced senescence in Brassica napus var oleifera. Physiol Veg, 1983,21:269-277. [本文引用: 1]
Jaradat MR, Feurtado JA, Huang DQ, Lu YQ, Cutler AJ . Multiple roles of the transcription factor AtMYBR1/AtMYB44 in ABA signaling, stress responses, and leaf senescence BMC Plant Biol, 2013,13:192-210. DOI:10.1186/1471-2229-13-192URL [本文引用: 1]
Di FF, Jian HJ, Wang TY, Chen XP, Ding YR, DuH, LuK, Li JN, LiuLZ . Genome-wide analysis of the PYL gene family and identification of PYL genes that respond to abiotic stress in Brassica napus Genes, 2018,9:156-173. [本文引用: 1]
Luo QX, PengM, Zhang XL, LeiP, Ji XM, WahsoonC, Meng FJ, Sun GY . Comparative mitochondrial proteomic, physiological, biochemical and ultrastructural profiling reveal factors underpinning salt tolerance in tetraploid black locust ( Robinia pseudoacacia L.) BMC Genomics, 2017,18:648-670. [本文引用: 1]
Fomeju BF, FalentinC, LassalleG , Manzanares-DauleuxM J, RégineD . Comparative genomic analysis of duplicated homoeologous regions involved in the resistance of Brassica napus to stem canker Front Plant Sci, 2015,6:772-785. [本文引用: 1]
RamseyJ . Polyploidy and ecological adaptation in wild yarrow Proc Natl Acad Sci USA, 2011,108:7096-7101. DOI:10.1073/pnas.1016631108URL [本文引用: 1]
AllarioT, BrumosJ , Colmenero-FloresJ M, IglesiasD J, PinaJ A, NavarroL, TalonM, OllitraultP, MorillonR . Tetraploid Rangpur lime rootstock increases drought tolerance via enhanced constitutive root abscisic acid production Plant Cell Environ, 2013,36:856-868. DOI:10.1111/pce.2013.36.issue-4URL [本文引用: 1]
Eliá?ováA, MünzbergováZ . Factors influencing distribution and local coexistence of diploids and tetraploids of Vicia cracca: inferences from a common garden experiment J Plant Res, 2017,130:677-687. [本文引用: 1]
Manzaneda AJ, Rey PJ, Bastida JM, Weiss-LehmanC, RaskinE, Mitchell-OldsT . Environmental aridity is associated with cytotype segregation and polyploidy occurrence in Brachypodium distachyon( Poaceae) New Phytol, 2012,193:797-805. [本文引用: 1]
Novikova PY, HohmannN , Vande Peer Y . Polyploid Arabidopsis species originated around recent glaciation maxima Curr Opin Plant Biol, 2018,42:8-15. [本文引用: 1]
XuY, XuH, WuX, FangX, WangJ . Genetic changes following hybridization and genome doubling in synthetic Brassica napus. Biochem Genet, 2012,50:616-624. [本文引用: 1]
Golicz AA, Bayer PE, Barker GC, Edger PP, KimH, Martinez PA , ChanC K K, Severn-EllisA, McCombieW R, ParkinI A P, PatersonA H, PiresJ C, SharpeA G, TangH B, TeakleG R, TownC D, BatleyJ, EdwardsD . The pangenome of an agronomically important crop plant Brassica oleracea. Nat Commun, 2016,7:13390-13397. [本文引用: 1]
李勤菲, 陈致富, 刘瑶, 梅家琴, 钱伟 . 六倍体( A nA nC nC nC oC o)与白菜型油菜杂交可交配性及后代菌核病抗性 中国农业科学, 2017,50:123-130. [本文引用: 1]
Li QF, Chen ZF, LiuY, Mei JQ, QianW . Crossability and Sclerotinia resistance among hybrids between hexaploid (A nA nC nC nC oC o) and Brassica rapa Sci Agric Sin, 2017,50:123-130 (in Chinese with English abstract). [本文引用: 1]
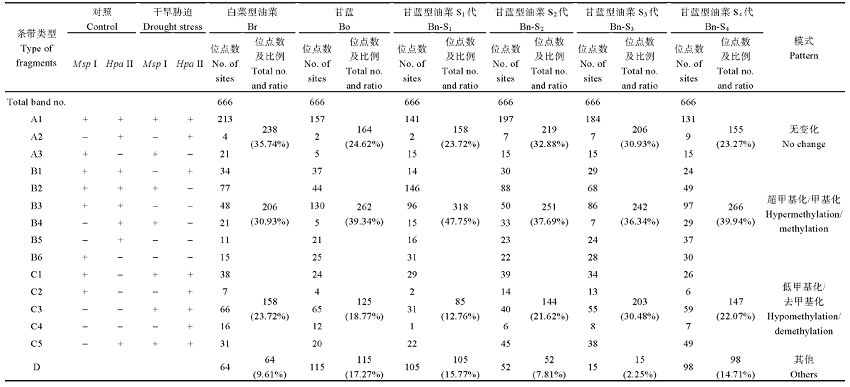
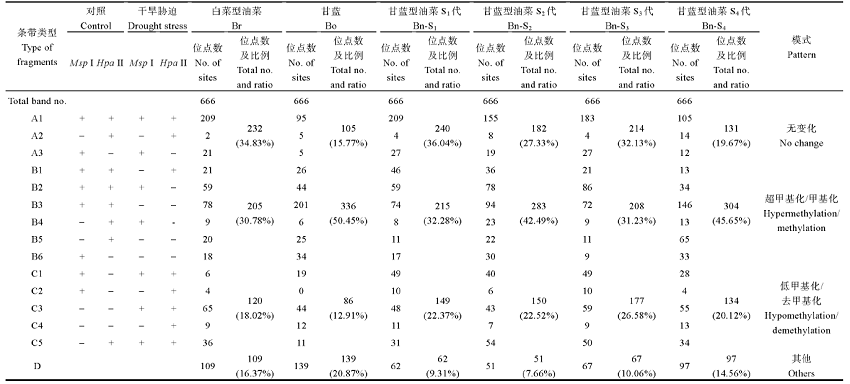
Ran LP, Fang TT, RongH, Jiang JJ, Fang YJ, Wang YP . Analysis of cytosine methylation in early generations of resynthesized Brassica napus . J Integr Agric, 2016,15:1228-123. [本文引用: 2]
Barrs HD, Weatherley PE . A re-examination of the relative turgidity technique for estimating water deficits in leaves Aust J Biol Sci, 1968,15:413-428. [本文引用: 1]
Hodges DM, Delong JM, Forney CF, Prangel RK . Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds Planta, 1999,207:604-611. DOI:10.1007/s004250050524URL [本文引用: 1]
Snyder JC, Desborough SL . Rapid estimation of potato tuber total protein content with coomassie brilliant blue G-250 Theor Appl Genet, 1978,52:135-139. [本文引用: 1]
Giannopolitis CN, Ries SK . Superoxide dismutases: II. Purification and quantitative relationship with water-soluble protein in seedlings (of corn, peas, and oats) Plant Physiol, 1977,59:315-318. DOI:10.1104/pp.59.2.315URL [本文引用: 1]
Xiong LZ, Xu CG , Saghai MaroofM A, ZhangQ F . Patterns of cytosine methylation in an elite rice hybrid and its parental lines, detected by a methylation-sensitive amplification polymorphism technique Mol Gen Genet, 1999,261:439-446. DOI:10.1007/s004380050986URL [本文引用: 1]
XieT, RongH, Jiang JJ, Kong YQ, Ran LP, WuJ, Wang YP . Analysis of DNA methylation patterns in resynthesized Brassica napus and diploid parents Acta Agron Sin, 2016,42:513-524 (in Chinese with English abstract). [本文引用: 2]
ValliyodanB, Nguyen HT . Understanding regulatory networks and engineering for enhanced drought tolerance in plants Curr Opin Plant Biol, 2006,9:189-195. DOI:10.1016/j.pbi.2006.01.019URL [本文引用: 1]
Qaderi MM, Kurepin LV, Reid DM . Growth and physiological responses of canola ( Brassica napus) to three components of global climate change: temperature, carbon dioxide and drought Physiol Plant, 2006,128:710-721. [本文引用: 1]
JoshiR, Wani SH, SinghB, BohraA, Dar ZA, Lone AA, PareekA , Singla-PareekSneh L . Transcription factors and plants response to drought stress: current understanding and future directions Front Plant Sci, 2016,7:1029-1043. [本文引用: 1]
Shao HB, Chu LY, Jaleel CA, ManivannanP, PanneerselvamR, Shao MA . Understanding water deficit stress-induced changes in the basic metabolism of higher plants-biotechnologically and sustainably improving agriculture and the ecoenvironment in arid regions of the globe Crit Rev Biotechnol, 2009,29:131-151. DOI:10.1080/07388550902869792URL [本文引用: 1]
LiuD, Pei ZF, Naeem MS, Ming DF, Liu HB, KhanF, Zhou WJ . 5-Aminolevulinic acid activates antioxidative defence system and seedling growth in Brassica napus L. under water-deficit stress J Agron Crop Sci, 2011,197:284-295. [本文引用: 1]
ShafiqS, Akram NA, AshrafM, ArshadA . Synergistic effects of drought and ascorbic acid on growth, mineral nutrients and oxidative defense system in canola ( Brassica napus L.) plants Acta Physiol Plant, 2014,36:1539-1553. [本文引用: 1]
AhmadJ, BashirH, BagheriR, BaigA, Al-HuqailA, Ibrahim MM, Qureshi MI . Drought and salinity induced changes in ecophysiology and proteomic profile of Parthenium hysterophorus PLoS One, 2017,12:e0185118. [本文引用: 1]
Akram NA, IqbalM, MuhammadA, AshrafM, Al-QurainyF, ShafiqS . Aminolevulinic acid and nitric oxide regulate oxidative defense and secondary metabolisms in canola ( Brassica napus L.) under drought stress Protoplasma, 2018,255:163-174. [本文引用: 1]
ShafiqS, Akram NA, AshrafM . Does exogenously-applied trehalose alter oxidative defense system in the edible part of radish ( Raphanus sativus L.) under water-deficit conditions? Sci Hortic, 2015,185:68-75. [本文引用: 1]
LatifM, Akram NA, AshrafM . Regulation of some biochemical attributes in drought-stressed cauliflower ( Brassica oleracea L.) by seed pretreatment with ascorbic acid Sci Hortic, 2016,91:129-137. [本文引用: 1]
CaverzanA, PassaiaG, Rosa SB, Ribeiro CW, LazzarottoF, Margis-PinheiroM . Plant responses to stresses: role of ascorbate peroxidase in the antioxidant protection Genet Mol Biol, 2012,35:1011-1019. DOI:10.1590/S1415-47572012000600016URL [本文引用: 1]
RenJ, Sun LN, Zhang QY, Song XS . Drought tolerance is correlated with the activity of antioxidant enzymes in Cerasus humilis seedlings Biomed Res Int, 2016,2016:2038-2047. [本文引用: 1]
Xia LJ, Yang LQ, Sun NL, LiJ, Fang YJ, Wang YP . Physiological and antioxidant enzyme gene expression analysis reveals the improved tolerance to drought stress of the somatic hybrid offspring of Brassica napus and Sinapis alba, at vegetative stage Acta Physiol Planta, 2016,38:1-10. [本文引用: 1]
WangT, Huang DY, Chen BY, Mao NN, Qiao YS, Ji MX . Differential expression of photosynthesis-related genes in pentaploid interspecific hybrid and its decaploid of Fragaria spp Genes Genomics, 2018,40:321-331. [本文引用: 1]
JiaoJ, WuJ, LyuZ, SunC, GaoL, YanX, CuiL, TangZ, YanB, JiaY . Methylation-sensitive amplified polymorphism-based genome-wide analysis of cytosine methylation profiles in Nicotiana tabacum cultivars Genet Mol Res, 2015,14:15177-15187. [本文引用: 1]
FangL, GongH, HuY, Liu CX, Zhou BL, HuangT, Wang YK, Chen SQ, Fang DD, Du XM, ChenH, Chen JD, WangS, WangQ, WanQ, Liu BL, Pan MQ, Chang LJ, Wu HT, Mei GF, XiangD, Li XH, Cai CP, Zhu XF, Chen ZJ, HanB, Chen XY, Guo WZ, Zhang TZ, Huang XH . Genomic insights into divergence and dual domestication of cultivated allotetraploid cottons Genome Biol, 2017,18:33-45. DOI:10.1186/s13059-017-1167-5URL [本文引用: 1]
Song QX, Zhang TZ, Stelly DM, Chen ZJ . Epigenomic and functional analyses reveal roles of epialleles in the loss of photoperiod sensitivity during domestication of allotetraploid cottons Genome Biol, 2017,18:99-112. DOI:10.1186/s13059-017-1229-8URL [本文引用: 1]
 ,*, 王幼平*扬州大学生物科学与技术学院, 江苏扬州 225009
,*, 王幼平*扬州大学生物科学与技术学院, 江苏扬州 225009
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT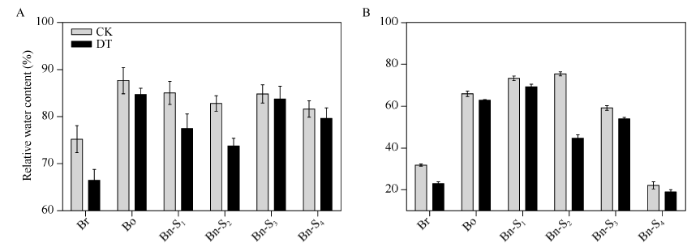 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT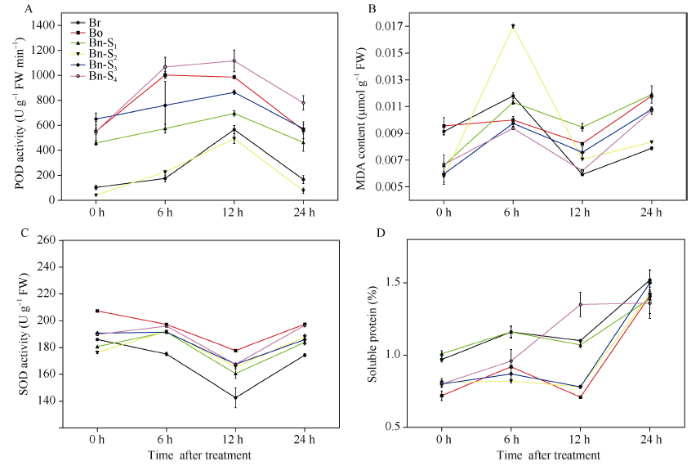 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT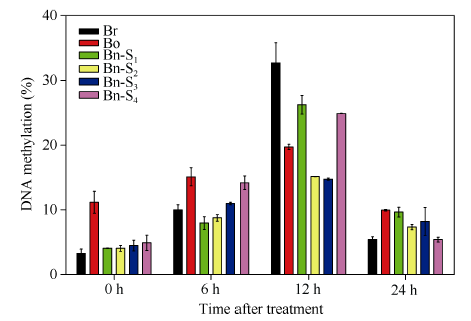 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT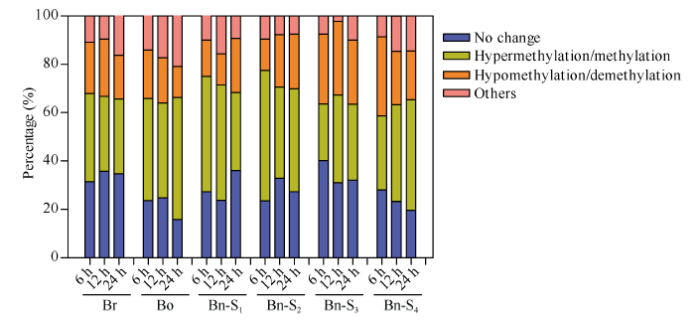 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT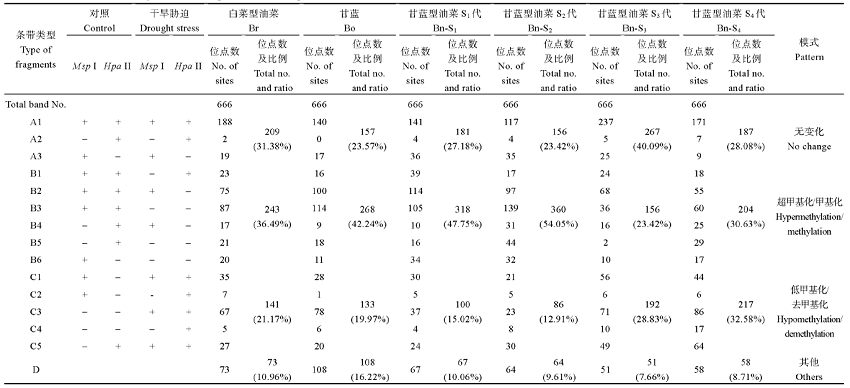

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}