 ,1,2,**, 钟鹏2,**, 刘杰,2,*, 唐中华3, 高亚冰2, 于洪久2, 郭炜21
,1,2,**, 钟鹏2,**, 刘杰,2,*, 唐中华3, 高亚冰2, 于洪久2, 郭炜21 2
3
Effect of low-temperature stress and gibberellin on seed germination and seedling physiological responses in peanut
CHANG Bo-Wen,1,2,**, ZHONG Peng2,**, LIU Jie,2,*, TANG Zhong-Hua3, GAO Ya-Bing2, YU Hong-Jiu2, GUO Wei21 2
3
通讯作者:
第一联系人:
收稿日期:2018-03-22接受日期:2018-08-20网络出版日期:2018-09-28
| 基金资助: |
Received:2018-03-22Accepted:2018-08-20Online:2018-09-28
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (404KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
常博文, 钟鹏, 刘杰, 唐中华, 高亚冰, 于洪久, 郭炜. 低温胁迫和赤霉素对花生种子萌发和幼苗生理响应的影响[J]. 作物学报, 2019, 45(1): 118-130. doi:10.3724/SP.J.1006.2019.84043
CHANG Bo-Wen, ZHONG Peng, LIU Jie, TANG Zhong-Hua, GAO Ya-Bing, YU Hong-Jiu, GUO Wei.
近年来, 黑龙江省倒春寒天气成为春季主要气象灾害之一。倒春寒天气产生的低温胁迫对作物种子出芽和幼苗生长有严重影响, 轻则致使出苗缓慢, 推迟作物的物候期, 重则导致种子“粉籽”、“烂种”或幼苗直接被冻死, 但至今对倒春寒天气引起的低温危害作物出苗和生长还未有系统研究[1]。花生(Arachis hypogaea L.)是我国主要的经济作物和油料作物, 也是我国食用蛋白和食用植物油源。黑龙江花生种植面积已达到13.3万公顷, 种植比例不断上升, 但倒春寒天气影响了黑龙江省春播花生出苗[2,3]。花生播种后常遭受低温威胁, 限制了春花生出芽、生长、发育, 甚至产量也受到影响。花生种子萌发最低温度为12℃[4], 封海胜[5]在2℃和6℃下筛选耐低温花生种质; 唐月异等[6]在种子吸胀期采用2℃冷浸的方法筛选耐低温花生种质; Chen等[7]在转录水平上筛选花生响应低温的基因, 但将倒春寒天气与花生种子萌发相结合的研究还未见报道。
赤霉素(gibberellic acid, GA)是一种植物激素, 作用于植物幼嫩组织, 适当浓度下能够促进种子萌发[8]。研究表明, 适量赤霉素可以有效地促进种子萌发。吕桂兰等[9]研究表明, 在10℃和15℃低温条件下GA3对大豆种子萌发有显著的促进作用。GA3可以部分或完全代替低温解除种子的休眠, 提高其发芽率[10]。许多研究表明, GA3参与植物生长发育的全过程, 如种子萌发、茎的伸长、花的诱导和发育、种子和果实的生长[11,12,13]。在种子萌发过程中, GA3促进淀粉酶和其他水解酶的合成, 并对受损的细胞膜起一定的修复作用[14]。此外, GA3在幼苗抗逆性形成方面也具有重要作用, 这可能和GA3诱导与抗逆性形成相关的基因表达和蛋白质的合成有关[15,16]。本研究将田间试验和温室试验相结合筛选耐低温花生种质, 以及探讨外源赤霉素处理对低温下花生种子萌发和幼苗生理响应的影响。
1 材料与方法
1.1 材料及处理
大田试验在黑龙江省农业科学院国家现代农业示范区(45°50′45.58″N, 126°51′3.81″E)进行。实验材料为2016年引进全国不同生态区的30个花生品种种子。2017年5月3日播种, 每穴1粒, 随机区组设计, 3次重复, 小区5 m×10 m, 行株距50 cm×25 cm。目前黑龙江省还没有统一的倒春寒规定和等级标准, 因此参照文献[17]倒春寒等级气象指标, 制定黑龙江省倒春寒等级指标(表1)。Table 1
表1
表1黑龙江省倒春寒等级指标
Table 1
| 倒春寒等级 Late spring chilling grades | 最大降温幅度ΔT Maximum cooling rate (oC) | 气温距平(δT) Temperature departure (oC) | 持续时间 Consecutive time (d) |
|---|---|---|---|
| 轻微 Light | 3.0 < ΔT24 ≤ 5.0 or 5.0 < ΔT48 ≤ 8.0 | -2.0 ≤ δT < -1.0 | ≥ 3 |
| 中度偏轻 Moderate partial light | 5.0 < Δ.0erate or 8.0 < Δ.0erate | -3.0 ≤ δT < -2.0 | ≥ 3 |
| 中度 Moderate | 8.0 < ΔT24 ≤ 10.0 or 10.0 < ΔT48 ≤ 12.0 | -5.0 ≤ δT < -3.0 | ≥ 5 |
| 严重 Severe | 10.0 < ΔT24 or 12.0 < ΔT48 | δT < -5.0 | ≥ 5 |
新窗口打开|下载CSV
温室试验在东北林业大学森林生态教育部重点实验室进行, 实验材料来自黑龙江省农业科学院农村能源研究所, 供试花生品种为大田试验筛选的4个较耐低温品种以及4个不耐低温品种。花生种子萌发试验使用人工气候箱(ZPW-400, China), 光照6级(day)/光照0级(night), 光照时间为6:00-18:00, 湿度80%, 温度28℃(day)/25℃(night)。
将种子置直径为20 cm的培养皿中温水浸泡吸胀24 h, 将温度调至2℃、4℃、6℃、12℃低温冷浸培养3 d, 28℃复温后在无菌水或加GA3溶液(浓度为100 μmol L-1、200 μmol L-1、300 μmol L-1)中培养5 d, 进行检测。每个实验组每个花生品种50粒种子, 重复3次。
将花生种子温水浸泡吸胀24 h后播种于珍珠岩中, 浇灌1/2 Hoagland营养液培养萌发后, 当幼苗3对真叶完全展开后, 进行4℃低温和GA3处理。低温处理是浇灌1/2 Hoagland营养液将幼苗置4℃人工气候箱中20 min, 复温1 h。GA3处理是提前浇灌分别含有不同浓度GA3 (浓度为100 μmol L-1、200 μmol L-1、300 μmol L-1)的1/2 Hoagland营养液12 h后将幼苗置4℃人工气候箱中20 min, 复温1 h。营养钵内口径10 cm、深10 cm, 每个实验组的每个品种12株, 试验重复3次。
1.2 试验方法
1.2.1 大田试验出苗率统计 于2017年6月3日在30个花生品种小区分别随机挑选3条垄, 统计播种穴数和出苗穴数。出苗率=出苗数(出苗穴数)/播种粒数(播种穴数)。1.2.2 温室发芽试验 以正常温度为对照, 统计8个花生品种在不同处理后种子发芽数、发芽时间、芽长等数据。参考文献[18,19]计算以下参数。芽长(下胚轴+胚根)≥10 mm为发芽。发芽率 = 发芽种子数/种子总数; 发芽指数(GI) = ∑Gt/Dt, 其中, Gt指t时的发芽率, Dt指发芽天数; 种子活力指数(VI) = S×GI, 其中, S指芽长(总芽长/种子总数), GI指发芽指数。
1.2.3 幼苗生理生化特性指标测定 参考文献[20]并改进。采集完全展开且叶位相同的叶片, 使用钻孔取样器(直径为1 cm)打取圆片, 将每个处理组样品分成2份, 每份1.0 g, 向烧杯中加入20 mL去离子水。一组使用真空泵抽取20 min, 室温静置30 min; 另一组使用沸水浴加热20 min, 冷却。使用电导仪分别测定电导率R1、R2, 相对膜透性以电解质外渗百分率表示。相对膜透性(%) = R1/R2×100%。
参照文献[21]的方法并改进。称取0.5 g叶片样品, 加入5 mL磷酸缓冲液冰浴研磨成匀浆, 离心后上清液为提取液。取2 mL提取液, 加入0.67%的TBA(硫代巴比妥酸)溶液4 mL, 沸水浴15 min, 冷却后离心取上清液。测定660 nm、532 nm、450 nm波长的吸光度。MDA含量C (mmol g-1 FW)= [6.45× (A532-A600)-0.56×A450]×Vt/FW×Vs, MDA (μmol g-1) = C×V/W, Vt为提取液总体积(mL); Vs为提取液试验体积(mL); FW为样品重(g)。
取新鲜叶片0.3 g, 加入10 mL ddH2O沸水浴1 h, 将提取液定容至25 mL。吸取1 mL提取液, 加入1 mL ddH2O、0.5 mL蒽酮乙酸乙酯和5 mL浓硫酸, 充分振荡, 沸水浴1 min冷却至室温, 630 nm波长下测吸光度。以不同浓度纯蔗糖制作标准曲线, 计算可溶性糖含量。
称取0.5 g叶片冻干粉末, 加入磷酸缓冲液研磨成匀浆后, 离心10 min, 取上清液为提取液。吸取1.0 mL提取液, 加入5 mL考马斯亮蓝试剂, 摇匀, 放置2 min后在595 nm波长下测量吸光度。将不同浓度的BSA (牛血清蛋白, Sigma, USA)溶液显色并测定吸光度, 制作标准曲线, 以外标法通过标准曲线计算可溶性蛋白含量。
称取0.5 g叶片冻干粉末, 加入3% (w/v)磺基水杨酸溶液5 mL, 沸水浴中提取15 min。吸取2 mL提取液分别加入冰醋酸和2.5% (w/v)酸性茚三酮溶液各2 mL。摇匀后, 沸水浴中加热显色30 min, 取出冷却至室温。加入5 mL甲苯充分摇匀, 避光静置4 h, 完全分层后用吸管吸取甲苯层, 使用紫外可见光分光光度计在520 nm下检测吸光度。将不同浓度和脯氨酸标准液也以同样方法显色并测定吸光度, 制作标准曲线, 以外标法通过标准曲线计算游离脯氨酸含量。
1.2.4 统计分析 使用SPSS 17.0软件进行单因素方差分析(One-way-ANOVA)和相关性分析检验各指标之间的相关性, Microsoft Excel软件作图。用Duncan’s检验(α=0.05)进行多重比较样品之间差异显著性。
2 结果与分析
2.1 春季播种期气温变化趋势和花生出苗率
由图1可知, 5月3日至6月3日1个月内发生2次倒春寒。5月1日至3日平均气温分别为19.5℃、22.5℃、23.5℃, 适合花生萌发, 但5月4日温度剧烈下降, 平均温度为15℃, ΔT24 > 8℃; 5月5日平均温度为12.5℃, 10.0 < ΔT48≤12.0; 5月6日平均温度6.5℃, 10.0 < ΔT48≤12.0, δT < -5.0, 持续3 d, 符合倒春寒天气的标准, 属于中度偏轻。第二次倒春寒在5月19-22日, 温度下降3.0 < ΔT24 < 8.0或5.0 < ΔT48≤8.0, -2.0≤δT < -1.0, 持续4 d, 程度为轻度。图1
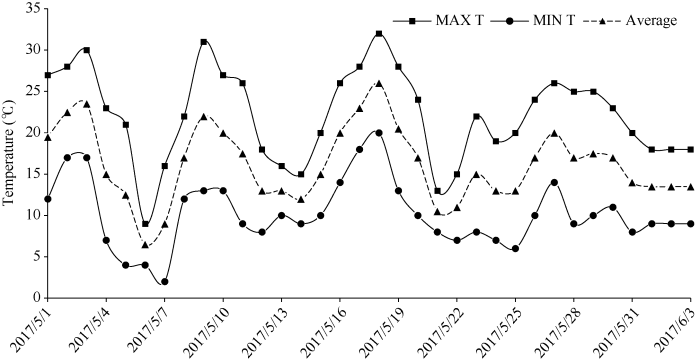 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图12017年5月至6月气温变化
MAX T: 日最高气温; MIN T: 日最低气温; Average: 日平均气温。
Fig. 1Changes of daily temperature from May 1 to June 3 in 2017
=MAX T: daily maximum of air temperature; MIN T: daily minimum of air temperature; Average: average value of air temperature.
统计花生播种1个月后的出苗率, 由表2可知, 出苗率较高的品种为阜花17 (FH17)、阜花12 (FH12)、冀花16 (JH16)、冀花18 (JH18), 出苗率分别为97.22%、95.36%、95.27%和94.56%; 而出苗率较低的品种分别为白玉花生(BHS)、正农黑花1号(HHS)、白沙1016 (BS1016)、七彩花生(QCHS)、鲁花11(LH11), 出苗率分别为45.51%、49.79%、50.28%、58.63%和59.63%, 选取FH17、FH12、JH16、JH18为耐低温实验组品种, BHS、HHS、BS1016和LH11为不耐低温实验组品种进行温室试验。
Table 2
表2
表2播种1个月后各花生品种出苗率
Table 2
| 品种 Cultivar | 出苗率 Seedling emergence rate (%) | 标准误差 Standard error | 差异显著性 Sig. 5% | |
|---|---|---|---|---|
| 白玉 Baiyu | 45.51 | 8.43 | a | |
| 正农黑花生1号 Zhengnongheihuasheng 1 | 49.79 | 10.08 | ab | |
| 白沙1016 Baisha 1016 | 50.28 | 8.52 | ab | |
| 七彩花生 Qicaihuasheng | 58.63 | 3.89 | bc | |
| 鲁花11 Luhua 11 | 59.63 | 9.76 | bcd | |
| 花育22 Huayu 22 | 67.33 | 10.25 | cde | |
| 荣丰8号 Rongfeng 8 | 68.42 | 9.58 | cde | |
| 丰花5号 Fenghua 5 | 69.29 | 5.32 | de | |
| 丰花3号 Fenghua 3 | 72.75 | 5.03 | ef | |
| 冀花11 Jihua 11 | 72.99 | 6.34 | ef | |
| 丰花1号 Fenghua 1 | 73.30 | 4.82 | efg | |
| 海花1号 Haihua 1 | 73.54 | 3.22 | efg | |
| 中农108 Zhongnong 108 | 74.00 | 8.92 | efg | |
| 山花10号 Shanhua 10 | 74.33 | 3.24 | efg | |
| 中花800 Zhonghua 800 | 75.00 | 4.77 | efg | |
| 拔二罐 Baerguan | 76.38 | 6.64 | efgh | |
| 花育23 Huayu 23 | 81.73 | 4.38 | fghi | |
| 品种 Cultivar | 出苗率 Seedling emergence rate (%) | 标准误差 Standard error | 差异显著性 Sig. 5% | |
| 冀花4号 Jihua 4 | 82.06 | 4.22 | fghi | |
| 冀花9号 Jihua 9 | 82.63 | 1.04 | fghij | |
| 冀花8号 Jihua 8 | 84.22 | 4.13 | ghijk | |
| 寿花820 Shouhua 820 | 86.38 | 5.66 | hijkl | |
| 花育36 Huayu 36 | 89.71 | 4.09 | ijkl | |
| 四粒红 Silihong | 90.19 | 5.24 | ijkl | |
| 冀花10号 Jihua 10 | 90.95 | 2.74 | ijkl | |
| 花育20 Huayu 20 | 91.43 | 2.74 | ijkl | |
| 冀花324 Jihua 324 | 93.40 | 1.89 | jkl | |
| 冀花18 Jihua 18 | 94.56 | 4.16 | kl | |
| 冀花16 Jihua 16 | 95.27 | 1.49 | l | |
| 阜花12 Fuhua 12 | 95.36 | 3.27 | l | |
| 阜花17 Fuhua 17 | 97.22 | 1.26 | l | |
新窗口打开|下载CSV
2.2 花生耐低温鉴定温度选择
选取2℃、4℃、6℃、12℃进行发芽对比试验, 旨在筛选合适的低温处理温度。由图2可知, 2℃处理时所有品种的发芽率显著下降, 耐低温品种发芽率都在60%以下; 4℃处理时不同耐低温性的品种之间差异明显, 相同耐低温性的品种发芽率差异较小; 12℃处理下所有花生品种发芽率差异不显著。综上, 以下试验都采用4℃作为试验处理温度。图2
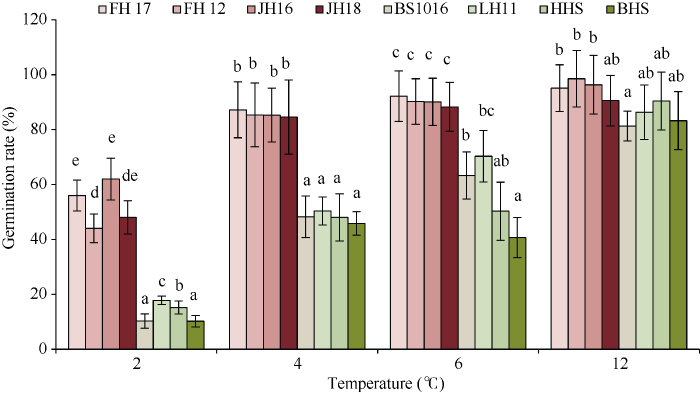 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同温度对花生种子发芽率的影响
FH17: 阜花17; FH12: 阜花12; JH16: 冀花16; JH18: 冀花18; BS1016: 白沙1016; LH11: 鲁花11; HHS: 正农黑花1号; BHS: 白玉花生。图柱上不同字母表示处理之间的0.05水平差异显著。
Fig. 2Effects of different temperatures on germination rate of peanut seeds
FH17: Fuhua 17; FH12: Fuhua 12; JH16: Jihua 16; JH18: Jihua 18; BS1016: Baisha 1016; LH11: Luhua 11; HHS: Zhengnongheihua 1; BHS: Baiyuhuasheng. Bars superscripted by different letters are significantly different at the 0.05 probability level among treatments.
2.3 低温胁迫和GA3对花生种子发芽率的影响
由表3可知, 常温条件下(对照组)耐低温和不耐低温的花生品种发芽率都达到95%以上, 品种间无明显差异。4℃处理后, 所有品种发芽率都有所下降, 耐低温品种发芽率比不耐低温品种下降幅度小, BHS发芽率下降幅度最大, 降低了53.29%, FH17发芽率下降了11.10%, 降低幅度最小。GA3处理后, 所有品种发芽率有显著提高, 并随GA3浓度升高而增加, 300 μmol L-1 GA3处理促进发芽率升高幅度最大, 不耐低温品种发芽率增加幅度较大, 都提高了25%以上。200~300 μmol L-1 GA3处理后, 耐低温品种发芽率达到了90%以上。Table 3
表3
表3低温胁迫和GA3对花生种子发芽率、发芽指数、芽长和种子活力的影响
Table 3
| 花生品种 Peanut cultivar | 对照组 Control group | 处理组 Treatment group | |||
|---|---|---|---|---|---|
| 4℃ | 4℃+100 μmol L-1 GA3 | 4℃+200 μmol L-1 GA3 | 4℃+300 μmolL-1 GA3 | ||
| 发芽率Seed germination rate (%) | |||||
| 阜花17 Fuhua 17 | 98.00±1.27 ab | 87.33±1.73 b | 89.33±1.04 b | 91.67±0.99 b | 95.33±1.46 b |
| 阜花12 Fuhua 12 | 97.33±1.44 ab | 85.33±1.02 b | 89.33±2.13 b | 92.33±0.57 b | 94.00±0.98 b |
| 冀花16 Jihua 16 | 98.67±0.67 ab | 85.00±1.67 b | 90.33±2.06 b | 92.33±1.33 b | 94.00±1.04 b |
| 冀花18 Jihua 18 | 99.00±1.09 b | 84.67±2.56 b | 90.00±2.01 b | 93.67±1.77 b | 95.00±2.76 b |
| 白沙1016 Baisha 1016 | 99.00±0.78 b | 48.33±4.37 a | 51.67±3.22 a | 57.00±4.06 a | 60.67±1.34 a |
| 鲁花11 Luhua 11 | 96.33±1.78 a | 50.67±5.34 a | 54.00±2.35 a | 60.67±3.33 a | 65.33±3.32 a |
| 正农黑花生1号 Zhengnongheihuasheng 1 | 99.00±0.33 b | 48.67±4.43 a | 54.00±4.78 a | 56.00±5.49 a | 60.67±3.97 a |
| 白玉Baiyu | 98.67±1.08 ab | 45.67±4.56 a | 50.00±1.98 a | 54.33±3.46 a | 61.67±3.22 a |
| 芽长Hypocotyl length (cm) | |||||
| 阜花17 Fuhua 17 | 5.36±0.46 a | 4.08±0.55 d | 4.23±0.33 c | 4.48±0.18 d | 4.94±0.25 c |
| 阜花12 Fuhua 12 | 4.98±0.45 a | 3.55±0.34 cd | 3.76±0.26 c | 4.01±0.18 c | 4.66±0.21 c |
| 冀花16 Jihua 16 | 5.23±0.22 a | 3.46±0.11 cd | 3.99±0.32 c | 4.31±0.15 cd | 4.79±0.54 c |
| 冀花18 Jihua 18 | 5.06±0.33 a | 3.33±0.21 c | 3.79±0.14 c | 4.29±0.21 cd | 4.48±0.28 c |
| 白沙1016 Baisha 1016 | 5.16±0.42 a | 1.88±0.09 a | 2.06±0.10 a | 2.14±0.13 a | 2.36±0.18 a |
| 鲁花11 Luhua 11 | 4.92±0.46 a | 2.19±0.08 b | 2.36±0.09 b | 2.65±0.12 b | 2.95±0.15 b |
| 正农黑花生1号 Zhengnongheihuasheng 1 | 4.86±0.50 a | 2.21±0.09 b | 2.37±0.21 b | 2.56±0.13 b | 2.83±0.14 b |
| 白玉 Baiyu | 5.26±0.45 a | 2.06±0.12 ab | 2.24±0.21 ab | 2.41±0.19 ab | 2.61±0.25 ab |
| 发芽指数 Germination index | |||||
| 阜花17 Fuhua 17 | 21.34±2.14 a | 16.24±1.24 b | 17.89±2.45 b | 18.56±1.11 b | 20.55±2.01 b |
| 阜花12 Fuhua 12 | 20.16±1.99 a | 15.06±1.45 b | 16.63±1.84 b | 17.94±1.86 b | 19.34±1.08 b |
| 冀花16 Jihua 16 | 21.33±2.01 a | 16.12±1.33 b | 17.64±1.04 b | 18.48±1.75 b | 20.26±1.88 b |
| 冀花18 Jihua 18 | 22.21±1.99 a | 14.90±1.23 b | 15.78±1.64 b | 17.30±1.13 b | 19.10±1.86 b |
| 白沙1016 Baisha 1016 | 20.55±2.01 a | 9.00±1.08 a | 10.35±1.44 a | 11.56±1.07 a | 12.62±2.33 a |
| 鲁花11 Luhua 11 | 19.17±2.88 a | 10.72±1.64 a | 11.69±1.37 a | 13.16±1.05 a | 14.97±1.75 a |
| 正农黑花生1号 Zhengnongheihuasheng 1 | 20.22±2.54 a | 9.90±1.85 a | 11.00±1.32 a | 12.02±1.56 a | 13.34±1.33 a |
| 白玉 Baiyu | 19.31±2.04 a | 9.38±1.15 a | 10.94±1.46 a | 12.26±1.35 a | 13.15±1.78 a |
| 种子活力指数 Seed vigor | |||||
| 阜花17 Fuhua 17 | 114.38±10.11 b | 66.28±6.91 c | 75.67±5.90 d | 83.17±7.45 c | 101.52±8.99 c |
| 阜花12 Fuhua 12 | 100.38±8.45 ab | 53.46±6.23 bc | 62.53±5.34 c | 71.94±7.89 c | 90.12±10.11 c |
| 冀花16 Jihua 16 | 111.58±10.58 ab | 55.78±6.03 bc | 70.38±5.84 cd | 79.65±7.77 c | 97.05±8.09 c |
| 冀花18 Jihua 18 | 112.39±8.23 b | 49.61±5.89 b | 59.81±7.82 c | 74.21±8.19 c | 85.56±8.74 c |
| 白沙1016 Baisha 1016 | 106.04±11.56 ab | 16.92±3.65 a | 21.33±2.98 a | 24.74±3.02 a | 29.78±2.00 a |
| 鲁花11 Luhua11 | 93.15±10.67 a | 23.68±3.98 a | 27.70±2.76 b | 33.69±3.14 b | 42.35±4.23 b |
| 正农黑花生1号 Zhengnongheihuasheng 1 | 99.46±11.49 ab | 21.67±2.89 a | 25.97±3.01 ab | 31.85±2.76 b | 39.35±3.45 b |
| 白玉 Baiyu | 101.59±10.97 ab | 19.32±2.05 a | 24.50±2.86 ab | 29.55±3.56 ab | 34.33±3.89 ab |
新窗口打开|下载CSV
2.4 低温胁迫和GA3对花生种子发芽指数、芽长和种子活力的影响
由表3可知, 低温处理后不耐低温品种的发芽指数、芽长、种子活力指数显著下降, 耐低温品种也有所下降, 但下降幅度比不耐低温品种幅度明显减小。施加GA3促进了各个品种发芽指数、芽长和种子活力指数, 随着GA3浓度升高, 促进作用也逐渐加强。300 μmol L-1 GA3处理对不耐低温品种芽长、发芽指数和种子活力指数促进作用稍微大于对耐低温品种。LH11芽长上升幅度最大, 增加了34.70%; BS1016和BHS发芽指数上升了40.21%和40.28%; 种子活力指数HHS上升了81.58%。JH16在300 μmol L-1 GA3处理后种子活力指数升高幅度最大, 上升了73.99%。2.5 低温胁迫和GA3对花生幼苗相对膜透性和丙二醛(MDA)含量的影响
4℃低温处理促进所有花生品种相对膜透性都升高, GA3处理后相对膜透性降低(表4)。4℃低温下, 耐低温品种相对膜透性上升幅度远远小于不耐低温品种, BHS和BS1016上升幅度为对照的0.93倍和0.90倍, FH17上升幅度最小, 上升了44.04%。相对膜透性随着GA3浓度的增大而减小, 说明GA3对低温诱导的相对膜透性升高有抑制作用, 300 μmol L-1 GA3的抑制作用最大, 其中FH12的相对膜透性比4℃处理时下降了25.06%。Table 4
表4
表4低温胁迫和GA3对花生幼苗相对膜透性、丙二醛、可溶性糖、可溶性蛋白和游离脯氨酸含量的影响
Table 4
| 生理指标 Physical indexes | 花生品种 Peanut cultivar | 对照组 Control group | 处理组Treatment group | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 4℃ | 4℃+100 μmol L-1 GA3 | 4℃+200 μmol L-1 GA3 | 4℃+300 μmol L-1 GA3 | |||||||
| 相对膜透性 Relative membrane permeability (%) | 阜花17 Fuhua 17 | 17.10±1.89 ab | 24.63±2.08 a | 21.63±1.65 a | 20.33±1.32 a | 18.71±1.16 a | ||||
| 阜花12 Fuhua 12 | 18.50±1.92 b | 26.98±1.36 a | 24.56±1.01 b | 22.68±1.06 ab | 20.22±1.45 ab | |||||
| 冀花16 Jihua16 | 15.92±1.65 a | 26.88±2.02 a | 25.69±1.54 b | 23.79±0.98 b | 21.25±0.99 b | |||||
| 冀花18 Jihua 18 | 15.32±1.54 a | 25.54±1.85 a | 24.46±1.99 ab | 23.03±1.33 b | 21.46±1.25 b | |||||
| 白沙1016 Baisha 1016 | 16.43±1.98 ab | 31.23±1.89 b | 30.36±1.45 c | 28.46±1.85 c | 26.18±1.28 c | |||||
| 鲁花11 Luhua 11 | 17.45±2.01 ab | 31.23±1.97 b | 30.56±1.67 c | 28.12±1.11 c | 26.54±1.98 c | |||||
| 正农黑花生1号 Zhengnongheihuasheng 1 | 17.95±1.85 ab | 30.91±1.16 b | 28.63±1.22 c | 27.56±1.00 c | 25.87±1.18 c | |||||
| 白玉Baiyu | 15.99±1.89 a | 30.88±1.96 b | 29.36±1.02 c | 27.98±1.45 c | 26.87±1.08 c | |||||
| 丙二醛含量 MDA content (μmol g-1 FW-1) | 阜花17 Fuhua 17 | 7.13±0.95 a | 9.56±1.04 a | 8.66±0.83 a | 7.84±0.66 a | 7.16±0.58 a | ||||
| 阜花12 Fuhua 12 | 7.56±0.96 a | 9.84±0.68 a | 8.92±0.51 a | 8.06±0.53 a | 7.98±0.73 a | |||||
| 冀花16 Jihua 16 | 6.35±0.83 a | 10.19±1.01 a | 9.13±0.77 a | 8.46±0.49 a | 7.39±0.50 a | |||||
| 冀花18 Jihua 18 | 6.84±0.77 a | 10.23±0.93 a | 9.04±1.00 a | 8.16±0.67 a | 7.28±0.63 a | |||||
| 白沙1016 Baisha 1016 | 6.45±0.99 a | 19.03±0.95 b | 17.85±0.73 b | 16.32±0.93 c | 14.88±1.06 c | |||||
| 鲁花11 Luhua 11 | 7.26±1.01 a | 18.32±0.99 b | 16.98±0.84 b | 15.36±0.56 bc | 13.86±0.99 bc | |||||
| 正农黑花生1号 Zhengnongheihuasheng 1 | 7.06±0.93 a | 19.26±0.58 b | 16.54±1.05 b | 14.46±0.50 b | 11.65±1.46 b | |||||
| 白玉 Baiyu | 6.98±0.95 a | 19.69±0.98 b | 17.05±1.51 b | 15.24±0.73 bc | 12.74±1.22 bc | |||||
| 可溶性糖 Soluble sugar content (mg g-1 FW-1) | 阜花17 Fuhua 17 | 7.46±1.95 b | 18.75±1.49 b | 21.36±0.83 b | 24.68±1.66 bc | 29.00±1.28 b | ||||
| 阜花12 Fuhua 12 | 7.96±1.36 b | 18.32±1.38 b | 20.69±0.51 b | 24.12±1.53 bc | 28.56±1.73 b | |||||
| 冀花16 Jihua 16 | 8.63±1.26 b | 17.11±1.51 b | 21.63±0.77 b | 25.55±1.49 c | 27.36±1.00 b | |||||
| 冀花18 Jihua 18 | 7.78±1.53 b | 18.56±1.43 b | 20.45±1.00 b | 22.69±1.67 b | 28.33±1.63 b | |||||
| 白沙1016 Baisha 1016 | 5.55±0.99 ab | 8.78±1.95 a | 10.96±0.73 a | 12.15±1.90 a | 14.15±1.56 a | |||||
| 鲁花11 Luhua 11 | 4.99±1.01 a | 8.82±1.49 a | 10.65±0.84 a | 12.21±1.56 a | 14.54±0.99 a | |||||
| 正农黑花生1号 Zhengnongheihuasheng 1 | 5.87±0.93 ab | 9.74±1.28 a | 11.96±1.05 a | 13.33±1.51 a | 15.78±1.46 a | |||||
| 白玉Baiyu | 6.45±1.34 ab | 8.64±1.48 a | 11.62±1.51 a | 13.05±1.73 a | 14.51±1.22 a | |||||
| 生理指标 Physical indexes | 花生品种 Peanut cultivar | 对照组 Control group | 处理组Treatment group | |||||||
| 4℃ | 4℃+100 μmol L-1 GA3 | 4℃+200 μmol L-1 GA3 | 4℃+300 μmol L-1 GA3 | |||||||
| 可溶性蛋白 Soluble protein content (μmol g-1 FW-1) | 阜花17 Fuhua 17 | 16.49±2.09 a | 25.45±0.98 b | 26.85±1.65 b | 27.96±1.66 b | 29.64±1.28 b | ||||
| 阜花12 Fuhua 12 | 17.68±1.22 a | 23.78±0.76 ab | 26.45±1.01 c | 28.65±1.53 b | 30.77±1.73 b | |||||
| 冀花16 Jihua 16 | 17.45±2.52 a | 24.36±1.22 ab | 25.85±1.54 bc | 26.56±1.49 ab | 29.86±1.00 b | |||||
| 冀花18 Jihua 18 | 16.12±2.06 a | 25.88±1.85b | 26.74±1.99 b | 27.65±1.67 b | 30.55±1.03 b | |||||
| 白沙1016 Baisha 1016 | 18.02±1.98 a | 23.71±1.89 ab | 24.56±1.45 ab | 25.74±1.23 ab | 26.16±1.56 a | |||||
| 鲁花11 Luhua 11 | 17.68±2.01 a | 23.52±1.67 ab | 23.65±1.67 ab | 24.31±1.56 a | 26.99±0.99 a | |||||
| 正农黑花生1号 Zhengnongheihuasheng 1 | 16.59±1.85 a | 22.34±1.56 a | 23.99±1.35 ab | 24.15±1.50 a | 26.59±1.46 a | |||||
| 白玉Baiyu | 17.66±2.68 a | 21.90±1.96 a | 23.06±1.02 a | 23.94±1.73 a | 25.35±1.22 a | |||||
| 游离脯氨酸 Free proline (μmol g-1 FW-1) | 阜花17 Fuhua 17 | 60.01±7.23 a | 83.95±7.24 a | 100.65±6.23 ab | 112.14±8.66 ab | 135.63±7.85 bc | ||||
| 阜花12 Fuhua 12 | 65.99±7.01 a | 85.45±8.36 a | 99.46±7.12 ab | 116.32±8.12 b | 140.63±7.78 c | |||||
| 冀花16 Jihua 16 | 61.23±8.96 a | 86.56±8.11 a | 101.25±7.52 ab | 110.36±7.78 ab | 136.44±8.33 bc | |||||
| 冀花18 Jihua 18 | 59.46±8.25 a | 80.11±8.46 a | 103.66±6.66 b | 120.56±8.36 b | 133.78±7.55 b | |||||
| 白沙1016 Baisha 1016 | 63.19±4.48 a | 78.25±7.12 a | 90.26±7.16 a | 108.36±7.45 ab | 122.32±6.06 ab | |||||
| 鲁花11 Luhua 11 | 61.85±6.85 a | 79.33±7.56 a | 90.29±8.56 a | 104.85±6.21 a | 118.62±7.60 a | |||||
| 正农黑花生1号 Zhengnongheihuasheng 1 | 58.36±6.99 a | 74.63±7.98 a | 91.29±7.31 a | 102.24±6.65 a | 121.55±8.32 ab | |||||
| 白玉 Baiyu | 62.45±7.11 a | 79.35±8.24 a | 95.11±8.56 ab | 104.15±7.12 a | 119.62±8.41 a | |||||
新窗口打开|下载CSV
丙二醛的变化趋势与相对膜透性相似, 4℃处理后所有花生品种丙二醛含量显著上升, GA3处理抑制丙二醛(表4)。4℃处理后, 不耐低温品种丙二醛含量上升幅度大于耐低温品种, 其中BS1016和BHS上升幅度较大, 分别是对照的1.95倍和1.82倍; FH12和FH17上升幅度较小, 增加了30.16%和34.08%。加入GA3抑制了4℃低温诱导的丙二醛含量升高, 随着GA3含量升高, 所有品种丙二醛含量都有所下降, 加入300 μmol L-1 GA3后丙二醛含量下降幅度最大, 特别是HHS降低了39.51%。
2.6 低温胁迫和GA3对花生幼苗可溶性糖、可溶性蛋白和游离脯氨酸含量的影响
由表4可知, 对照组中耐低温品种可溶性糖含量高于不耐低温品种。4℃处理后花生可溶性糖含量上升, 耐低温性强的品种可溶性糖含量显著上升, FH17和JH18上升较大, 增加了1.51倍和1.39倍; 不耐低温品种上升幅度较小, 其中BHS上升最小, 升高了33.95%。加入GA3后促进各品种可溶性糖含量升高, 并且随着GA3浓度的升高而上升。100~300 μmol L-1 GA3处理不耐低温品种LH11、BHS和HHS上升幅度超过耐低温品种, 特别是BHS和LH11在300 μmol L-1 GA3处理后上升幅度较大, 升高了67.94%和64.85%。4℃处理后各品种可溶性蛋白含量变化趋势与可溶性糖含量相似。常温条件下各品种可溶性蛋白含量差异不显著。4℃条件下各个品种可溶性蛋白显著升高, 耐低温品种可溶性蛋白升高幅度与不耐低温品种升高幅度差异不显著, JH18和FH17上升幅度达到了60.55%和54.34%, HHS上升幅度最小, 上升了6.83%。加入GA3后所有品种可溶性蛋白含量都有上升趋势。不耐低温品种在4℃处理后加入100 μmol L-1 GA3, 可溶性蛋白含量上升显著。加入300 μmol L-1 GA3所有品种可溶性蛋白含量上升幅度最大, 其中FH12可溶性蛋白含量最高, 上升了29.39% (表4)。
低温导致所有花生品种游离脯氨酸含量上升, 其中上升较多的是JH18和FH17, 增加了41.37%和39.89%。各花生品种在4℃处理后加入GA3, 随着处理浓度升高游离脯氨酸含量上升。GA3浓度为300 μmol L-1时游离脯氨酸含量最高, 与低温处理相比JH18上升幅度最大, 上升了67.00%。耐低温品种的游离脯氨酸含量显著高于不耐低温品种(表4)。
2.7 低温胁迫下种子萌发相关指标与幼苗生理生化指标相关性分析
由表5可知, 种子萌发相关指标发芽率、发芽指数、种子活力和芽长之间存在极显著正相关(P<0.01)。种子萌发相关指标与相对膜透性、丙二醛存在极显著负相关(P<0.01), 与可溶性糖呈现极显著正相关(P<0.01)。可溶性蛋白与发芽率和芽长为显著正相关(P<0.05), 但与发芽指数和种子活力相关性不显著。脯氨酸含量与发芽率和发芽指数存在极显著正相关(P<0.01), 与种子活力和芽长呈现显著正相关(P<0.05)。Table 5
表5
表5种子萌发相关指标与幼苗生理生化指标的相关系数
Table 5
| GR | GI | SV | HL | RMP | MDA | SS | SP | FP | |
|---|---|---|---|---|---|---|---|---|---|
| 发芽率GR | 1 | ||||||||
| 发芽指数GI | 0.935** | 1 | |||||||
| 种子活力指数SV | 0.923** | 0.982** | 1 | ||||||
| 芽长HL | 0.961** | 0.936** | 0.964** | 1 | |||||
| 相对膜透性RMP | -0.923** | -0.888** | -0.938** | -0.981** | 1 | ||||
| MDA含量 MDA | -0.997** | -0.934** | -0.934** | -0.972** | 0.943** | 1 | |||
| 可溶性糖含量SS | 0.973** | 0.872** | 0.897** | 0.970** | -0.960** | -0.983** | 1 | ||
| 可溶性蛋白含量SP | 0.742* | 0.556 | 0.583 | 0.737* | -0.718* | -0.742* | 0.829* | 1 | |
| 游离脯氨酸含量FP | 0.864** | 0.882** | 0.810* | 0.820* | -0.745* | -0.834* | 0.779* | 0.64 | 1 |
新窗口打开|下载CSV
本研究以发芽率为参考指标, 将不同品种(A)、温度(B)和赤霉素浓度(C)作为效应因子, 分析其效应相互影响的显著性对发芽率的影响。首先, 各水平结合下的数据正态分布检验, Shapiro-Wilk检验结果表明除了耐低温品种在常温下使用300 μmol L-1 GA3处理后发芽率(P=0.024)不符合, 其余各组数据均符合正态分布(P>0.05)(表6)。由方差检验分析结果(表7)可知A、B、C主效应都已达到显著, A与B的交互效应达到显著(P<0.01), 但A×C、A×B×C之间的交互效应不显著(P>0.01)。对效应因子B和C在A1和A2两种水平上进行简单效应检验(表8)表明, B在A1水平上简单效应不显著(P>0.05), B在A2水平上简单效应极显著(P<0.01), 效应因子C在A1和A2水平上都不显著(P>0.05)。
Table 6
表6
表6效应因子正态分布检验
Table 6
| A | B | C | Shapiro-Wilk检验 Shapiro-Wilk test | ||
|---|---|---|---|---|---|
| 统计量Statistic | 自由度df | 显著性Sig. | |||
| 耐低温品种 Low-temperature tolerant varieties (A1) | 对照Control | 0 | 0.966 | 4 | 0.814 |
| 300 μmol L-1 | 0.873 | 4 | 0.310 | ||
| 4℃ | 0 | 0.876 | 4 | 0.322 | |
| 300 μmol L-1 | 0.729 | 4 | 0.024 | ||
| 不耐低温品种 Low-temperature susceptible varieties (A2) | 对照Control | 0 | 0.827 | 4 | 0.161 |
| 300 μmol L-1 | 0.917 | 4 | 0.519 | ||
| 4℃ | 0 | 0.965 | 4 | 0.811 | |
| 300 μmol L-1 | 0.791 | 4 | 0.086 | ||
新窗口打开|下载CSV
Table 7
表7
表7不同效应因子交互效应方差分析结果
Table 7
| 效应因子 Source | 显著性 Sig. |
|---|---|
| A | 0.000 |
| B | 0.000 |
| C | 0.000 |
| A×B | 0.000 |
| A×C | 0.011 |
| B×C | 0.000 |
| A×B×C | 0.011 |
新窗口打开|下载CSV
Table 8
表8
表8效应因子交互作用简单效应检验
Table 8
| 效应因子 Source | F值 F-value | 显著性 Sig. |
|---|---|---|
| B WITHIN A1 | 3.23 | 0.084 |
| B WITHIN A2 | 77.70 | 0.000 |
| C WITHIN A1 | 1.19 | 0.286 |
| C WITHIN A2 | 2.52 | 0.124 |
新窗口打开|下载CSV
3 讨论
3.1 低温对花生种子萌发和生长的影响
近年来, 我国全国范围内倒春寒发生频率逐渐上升, 但各地区对倒春寒研究进度不尽相同, 以西南(滇桂黔)发表研究论文最多, 东北地区发表论文最少[1]。温度对花生种植的影响很大, 针对黑龙江地区倒春寒研究相对较少的问题, 开展了倒春寒对花生出苗影响的大田试验和低温胁迫对花生种子萌发影响的相关研究。研究表明, 春季温度在12℃以上种植不覆膜花生最为适宜。低温常导致花生发芽延缓、生长缓慢、干物质积累减少, 甚至种子丧失发芽能力等后果[22], 我国北方花生产区出现倒春寒天气时, 易导致大面积烂种。王晶珊等[23]研究结果表明, 花生在出苗阶段, 种子吸胀萌动和初始发育阶段对低温最为敏感。本研究大田试验时期花生播种后出现了2次倒春寒天气。其后, 各花生品种出苗率差异显著, 阜花12、阜花17、冀花16、冀花18、花育20、四粒红等品种出苗率可达到90%以上, 说明其耐低温能力较强, 低温对其种子萌发出苗影响不大, 但鲁花11、白沙1016、七彩花生、白花生、黑花生等品种发芽率在60%以下, 说明这些品种相对耐低温性较弱, 倒春寒对其出苗影响较大。温室试验中发芽率等萌发相关指数与大田试验结果一致。值得关注的是白花生、黑花生和七彩花生这类“彩色花生”品种耐低温性较差。丙二醛(MDA)是植物在遭受逆境胁迫时细胞膜脂过氧化作用生成的最终产物, 所以MDA浓度一定程度上反映了细胞膜受伤害的程度。温室试验结果表明, 较耐低温品种(阜花12、阜花17、冀花16、冀花18)在低温处理后MDA含量上升幅度低于不耐低温品种(鲁花11、白沙1016、白花生和黑花生), 说明耐低温性强的花生品种细胞膜受低温伤害较小。细胞膜相对透性的变化能反映植物低温伤害程度, 相对膜透性越大说明低温伤害越大[24]。正常生长条件下各品种相对膜透性无显著差异, 但低温处理后不耐低温品种叶片的电导率上升明显, 说明其相对膜透性增大。通过MDA和相对膜透性的结果表明不耐低温品种在低温处理后受伤害更严重。植物体内可溶性糖、可溶性蛋白有较强亲水性, 明显增强细胞持水力, 减少低温导致原生质结冰致死的可能, 从而提高植物耐低温能力[25]。本研究中耐低温品种在低温处理后可溶性糖和可溶性蛋白量上升显著, 正常条件下不耐低温品种的可溶性糖含量较低。植物细胞内游离脯氨酸也是一类重要的渗透调节物[26], 作用与可溶性糖等相似, 增加细胞内溶质量, 防止细胞过度失水造成渗透胁迫, 降低冰点, 减轻低温对细胞的损害[27]。以前研究中脯氨酸含量与抗渗透胁迫之间呈现显著正相关, 脯氨酸含量也作为植物抗逆性评价的重要指标之一[28,29,30]。本研究结果表明低温处理后较耐低温品种脯氨酸含量较高, 升高幅度最大, 与前人研究结果一致。
3.2 赤霉素对花生种子萌发和生长的影响
赤霉素(GA)是一类植物激素, 在植物生长的整个生命周期中都发挥重要作用, 在植物种子萌发、幼苗发育过程中茎和下胚轴伸长和叶子扩张[31]等方面具有必不可少的作用。许多研究表明, 赤霉素有促进种子萌发的作用, 并且对打破种子休眠、提高发芽率、缩短发芽时间有显著效果[32,33,34], 同时赤霉素还可以缓解逆境胁迫对种子萌发和幼苗生长的抑制作用[35]。本研究中GA3显著缓解低温对花生种子萌发的抑制, 特别是300 μmol L-1 GA3对花生种子萌发的促进作用最大。代勋等[36]报道, 赤霉素能够抑制低温和干旱导致的电解质渗透率增加和MDA积累。刘永庆等[14]证明赤霉素能破除种子休眠, 促进淀粉酶和水解酶合成, 修复受损的细胞膜。本研究中, GA3处理后各花生品种MDA含量、相对膜透性都显著下降, 说明GA3对低温造成的MDA含量升高和相对膜透性上升具有抑制作用, GA3缓解低温对花生幼苗伤害, 与前人研究结果一致。另外, GA3提高花生幼苗可溶性糖、可溶性蛋白和游离脯氨酸含量(表4), 也从侧面说明花生幼苗抗寒性有所增加。研究表明[37,38,39]植物幼苗时期施加赤霉素不仅可以促进幼苗叶片可溶性糖含量, 也能促进植物结实量和果实品质, 赤霉素对花生果实品质的影响需要进一步研究。3.3 低温和赤霉素对不同花生品种发芽率的简单效应分析
作物的耐低温性是作物在遭受低温胁迫时能够抵御胁迫, 维持生命延续的一种能力, 同一种作物不同品种之间也会存在较大差异。通过对品种、温度、赤霉素3个效应因子进行简单效应分析可知低温对不同品种影响不同, 低温对较耐低温品种影响不显著, 但对不耐低温品种影响非常显著; 另外简单效应检验表明, 赤霉素在品种间差异不显著, 这可能是由于赤霉素对种子萌发的促进作用具有普适性。本研究通过大田试验统计不同花生品种在倒春寒后出苗情况结合温室试验检测低温胁迫对花生发芽率、种子活力、发芽指数和幼苗低温下的生理生化指标综合分析, 筛选了耐低温花生品种, 今后可进一步验证在黑龙江地区种植的可行性并进行推广。另外, 在花生播种前使用一定浓度赤霉素处理种子, 能够促进花生发芽和提高幼苗耐低温能力。4 结论
筛选出耐低温的品种(阜花12、阜花17、冀花16、冀花18)和不耐低温的品种(鲁花11、白沙1016、黑花生、白花生)。前者在遭遇低温胁迫后种子发芽率、发芽指数、种子活力和芽长高于后者; MDA含量、相对膜透性低于后者, 可溶性糖、脯氨酸含量显著高于后者。赤霉素对低温导致的种子萌发降低和幼苗生长阻碍有一定缓解作用, 并且300 μmol L-1赤霉素的作用效果最大。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.11937/bfyy.201516046URL [本文引用: 2]

近年来,随着气候变化而产生的"倒春寒"天气迹象日趋明显,然而中国尚未完全掌握其对农业生态系统的危害原理,尤其有关树木生长方面的信息反馈十分有限。资料显示:中国对"倒春寒"的关注始于1975年,但主要研究区主要集中于中国西南地区,因此目前尚缺乏全国尺度上的把握;研究内容方面国内侧重"倒春寒"的大气成因及其对农作物冻害的评估,有关树木受害的关注度明显不足,相反国外研究更加注重探讨"倒春寒"对森林生态系统的影响方面并且已经取得了一定的进展;在"倒春寒"对植物的冻害机制方面,中国研究更倾向于"持续低温"和"气温骤降"的影响,而国外则认为是在全球变暖作用下植物物候提前所致。通过国外有关树木的研究结论对中国全境尺度春季温度数据进行分析后,发现近年来"倒春寒"很有可能已经给中国树木的春季生长造成了严重影响,希望引起同行们对此问题的重视。
DOI:10.11937/bfyy.201516046URL [本文引用: 2]
近年来,随着气候变化而产生的"倒春寒"天气迹象日趋明显,然而中国尚未完全掌握其对农业生态系统的危害原理,尤其有关树木生长方面的信息反馈十分有限。资料显示:中国对"倒春寒"的关注始于1975年,但主要研究区主要集中于中国西南地区,因此目前尚缺乏全国尺度上的把握;研究内容方面国内侧重"倒春寒"的大气成因及其对农作物冻害的评估,有关树木受害的关注度明显不足,相反国外研究更加注重探讨"倒春寒"对森林生态系统的影响方面并且已经取得了一定的进展;在"倒春寒"对植物的冻害机制方面,中国研究更倾向于"持续低温"和"气温骤降"的影响,而国外则认为是在全球变暖作用下植物物候提前所致。通过国外有关树木的研究结论对中国全境尺度春季温度数据进行分析后,发现近年来"倒春寒"很有可能已经给中国树木的春季生长造成了严重影响,希望引起同行们对此问题的重视。
DOI:10.1017/S0960258500001847URL [本文引用: 1]
Germinating seeds of many species, especially those of tropical or subtropical origin, suffer chilling injury when exposed to low but non-freezing temperatures, resulting in poor seedling establishment and reduction in yield. The severity of injury depends upon several factors such as (i) the species or the cultivars involved, (ii) the initial water content of the seed, (iii) the temperature to which seed is exposed, (iv) the duration of chilling exposure and (v) the period during the course of germination when the chilling exposure takes place. The period of imbibition by seeds is the period of sensitivity to the stresses but especially to chilling temperatures. Imbibitional chilling injury is defined as sensitivity to a combination of low seed-water content and imbibition at cold temperature. The mechanism of chilling injury in seeds is different from chilling injury of hydrated tissues and is discussed in relation to the involvement of membranes, respiration, rate of hydration and the concept of discreet moisture levels. Furthermore, the mechanistic and applied aspects of pre-sowing seed treatments for improved germination performance under chilling stress are discussed with an emphasis on hydration-dehydration, osmotic priming and growth regulator treatments.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
Tolerance to low temperaterature during germination for fifty-five accessions of peanut germplasm were evaluated when the seeds were imbibed at 2℃ at 96h followed by 25℃ for 72h. Percentage of the number of seeds with radicals breaking through testa(PSWRBT) and ratio of the length of hypocotyls and radicals to the length of seeds(RHRS) were calculated. Main quality traits including oleic acid, linoleic acid, palmitic acid, oil, protein and sucrose contents were determined by near-infrared spectroscopy. Relationship between low temperature tolerance of peanut seed during imbibition and individual quality attributes was analyzed. The results showed that under low temperature stress condition, 3,3,5 and 43 accessions had the PSERTBT of ≥80%, 70%~80%, 60%~70% and0.5, 0.4~0.5, 0.4~0.3 and
URL [本文引用: 1]
Tolerance to low temperaterature during germination for fifty-five accessions of peanut germplasm were evaluated when the seeds were imbibed at 2℃ at 96h followed by 25℃ for 72h. Percentage of the number of seeds with radicals breaking through testa(PSWRBT) and ratio of the length of hypocotyls and radicals to the length of seeds(RHRS) were calculated. Main quality traits including oleic acid, linoleic acid, palmitic acid, oil, protein and sucrose contents were determined by near-infrared spectroscopy. Relationship between low temperature tolerance of peanut seed during imbibition and individual quality attributes was analyzed. The results showed that under low temperature stress condition, 3,3,5 and 43 accessions had the PSERTBT of ≥80%, 70%~80%, 60%~70% and0.5, 0.4~0.5, 0.4~0.3 and
DOI:10.1016/j.scienta.2014.01.043URL [本文引用: 1]
The cultivated peanut (Arachis hypogaea L.) is important oil crop and cold stress seriously influences its development and yields. Tolerant varieties produced using transgenic techniques will effectively increase peanut plantation area and enhance yields. However, little is known about the network of gene expression regulation related to cold stress in peanut. Therefore, we screened genes regulated by cold stress in peanut. In total, 1478 up-regulated and 1510 down-regulated probe sets were successfully identified in leaves of peanut subjected to cold stress using the microarray technique. Data analysis indicated that the main biological processes involved in cold stress response included signal transduction, transcript regulations and accumulation of soluble metabolites. Regulation of cell division, translation, protein modification, transport and DNA/RNA-related processes were also involved in peanut cold acclimation. Our study also revealed that protein kinases, transcription factors, heat shock proteins, resistance proteins, stilbene synthase, and so on, may play important functions in cold stress regulation of peanut. The function of some probe sets in cold stress regulation had not been clarified (e.g. proteins functioning in glycerol ether metabolic process, microtubule-based movement or nutrient reservoir activity). Some genes we identified lack functional annotation and their roles in response to cold are yet to be elucidated. These results showed an overview of the transcription map of peanut under cold stress, which may yield some useful insights into cold-mediated signal transduction pathways in peanut and offer some candidate genes as potential markers of tolerance to cold stress.
[本文引用: 1]
URL [本文引用: 1]
春播大豆会因低温而延迟萌发和出苗,本研究在控制温度下用不同浓度赤霉素和激动素处理大豆种子,以促进大豆种子在低温下提早萌发,结果表明在10℃和15℃低温条件下赤霉素和激动素有显著地促进大豆种子萌发的作用,并且对大豆幼根发育也有明显的促进作用。
URL [本文引用: 1]
春播大豆会因低温而延迟萌发和出苗,本研究在控制温度下用不同浓度赤霉素和激动素处理大豆种子,以促进大豆种子在低温下提早萌发,结果表明在10℃和15℃低温条件下赤霉素和激动素有显著地促进大豆种子萌发的作用,并且对大豆幼根发育也有明显的促进作用。
DOI:10.3969/j.issn.1001-0009.2006.05.022URL [本文引用: 1]
越橘种子具有休眠特性,自然条件下萌发困难,发芽率低.本试验采 用低温和GA3处理研究解除越橘种子休眠、促进萌发、提高发芽率的方法.结果表明:GA3能代替低温层积,促进越橘种子发芽.GA3处理和低温层积相结 合,效果明显高于单独使用GA3.试验中最有效的处理是GA3200 mg/L浸渍12 h后,4℃层积处理20 d,其发芽势达到40%,发芽率达到50%,芽的生长情况较其它处理好.
DOI:10.3969/j.issn.1001-0009.2006.05.022URL [本文引用: 1]
越橘种子具有休眠特性,自然条件下萌发困难,发芽率低.本试验采 用低温和GA3处理研究解除越橘种子休眠、促进萌发、提高发芽率的方法.结果表明:GA3能代替低温层积,促进越橘种子发芽.GA3处理和低温层积相结 合,效果明显高于单独使用GA3.试验中最有效的处理是GA3200 mg/L浸渍12 h后,4℃层积处理20 d,其发芽势达到40%,发芽率达到50%,芽的生长情况较其它处理好.
DOI:10.1016/S0958-1669(00)00071-9URLPMID:10753762 [本文引用: 1]
Modification of plant hormone biosynthesis through the introduction of bacterial genes is a natural form of genetic engineering, which has been exploited in numerous studies on hormone function. Recently, biosynthetic pathways have been largely elucidated for most of the plant hormone classes, and genes encoding many of the enzymes have been cloned. These advances offer new opportunities to manipulate hormone content in order to study their mode of action and the regulation of their biosynthesis. Furthermore, this technology is providing the means to introduce agriculturally useful traits into crops.
DOI:10.1016/S0304-4238(02)00018-3URL [本文引用: 1]
Small tubers (1.5 cm diameter) of Zantedeschia ‘Black Magic’ were used to study the induction of flowering by gibberellic acid (GA 3) and gibberellins 4+7 (GA 4+7). When planted these tubers remained vegetative unless treated with gibberellin. To control gibberellin dose, tubers were dipped into treatment solution (0–1000 mg l 611) for defined periods (0–16 h) and then rinsed with water to remove gibberellin from the tuber surface. The tubers were then planted and grown in a greenhouse. Flower emergence was recorded and plants were then dissected to determine whether initiation had occurred on non-flowering plants and whether axillary buds were vegetative or floral. No differences were found between responses to GA 3 or GA 4+7 treatments. The proportion of shoots that initiated inflorescences increased with an increase in both gibberellin concentration and/or treatment duration. Increased gibberellin dose also increased in the proportion of initiated flowers that subsequently emerged and the proportion of axillary buds that initiated flowers. Plants treated with high gibberellin doses also flowered earlier and with a lower leaf number. Our results indicate that high gibberellin doses induce more rapid floral initiation than lower doses. In addition to their role in floral initiation, gibberellins play a role in floral development. Uptake of gibberellin is shown to be primarily through the buds on the upper surface of the tuber.
DOI:10.1016/j.plaphy.2004.03.011URLPMID:15331092 [本文引用: 1]
In plants, Lewis a type N-glycans may be involved in cell-to-cell communication and recognition. N-glycoproteins harboring Lewis a glycotopes are mainly found in plasma membranes and cell walls. Some can be also involved in cell wall synthesis or the loosening process, and subsequently in cell elongation. In order to determine the potential role(s) of the 伪4-fucosylation during vegetative development, transgenic tobacco plants overexpressing a human Lewis fucosyltransferase (hFUT3), which transfers a fucose residue in a (1,4)-linkage on complex glycans, have been developed. The heterologous enzyme hFUT3 was strongly expressed and fully functional in transgenic tobacco. Transgenic plants showed a delay in growth linked to a reduction of internode length. Furthermore, transgenic seedling roots were significantly shorter than wild-type roots and the length of their epidermis cells was reduced. Strikingly, root growth was completely and specifically restored following gibberellin treatment. Etiolated hypocotyls of hFUT3 overexpressors were also more sensitive to exogenous gibberellin. Furthermore, paclobutrazol, an inhibitor of gibberellin synthesis, induced a similar effect on control and transgenic dark-grown hypocotyls suggesting that gibberellin biosynthesis was probably not altered in seedlings overexpressing hFUT3. Thus, 4-fucosylation could act as a possible modulator of conformation and/or functioning of N-glycoproteins involved in the gibberellin-dependent elongation process.
DOI:10.1007/BF02007173URL [本文引用: 2]
利用细胞流检仪测番茄(LycopersiconesculentumMill.(GA-缺 陷型、ABA-缺陷型和相应的正常品种(野生型)成熟种子胚根尖细胞倍性水平时发现:GA-缺陷型和野生型种子绝大多数细胞DNA水平为2C,而ABA- 缺陷型种子则含有较多的4C细胞。在标准发芽条件下,ABA-缺陷型和野型种子浸种1d后胚根尖细胞DNA开始复制,随后胚根突破种皮而发芽。然而GA- 缺陷型子除非加入外源GA,
DOI:10.1007/BF02007173URL [本文引用: 2]
利用细胞流检仪测番茄(LycopersiconesculentumMill.(GA-缺 陷型、ABA-缺陷型和相应的正常品种(野生型)成熟种子胚根尖细胞倍性水平时发现:GA-缺陷型和野生型种子绝大多数细胞DNA水平为2C,而ABA- 缺陷型种子则含有较多的4C细胞。在标准发芽条件下,ABA-缺陷型和野型种子浸种1d后胚根尖细胞DNA开始复制,随后胚根突破种皮而发芽。然而GA- 缺陷型子除非加入外源GA,
URL [本文引用: 1]
以番茄幼苗为材料,研究了50 mg/g的赤霉素(GA3)和清水浸种后用不同浓度的聚乙二醇(0、40、80、120 mg/g)胁迫对番茄种子萌发及幼苗生长的影响。结果表明:40 mg/g的聚乙二醇胁迫对番茄种子的萌发率、发芽指数、幼苗株高、根长有促进作用,120 mg/g的聚乙二醇明显抑制种子的萌发率、发芽指数、幼苗株高、根长;不同浓度的聚乙二醇胁迫明显降低了幼苗的叶绿素含量。用50 mg/g GA3浸种后明显增加聚乙二醇胁迫下番茄种子的萌发率、发芽指数、幼苗株高、根长和叶绿素含量。用50 mg/g GA3浸种对番茄幼苗的生长有一定的促进作用。
URL [本文引用: 1]
以番茄幼苗为材料,研究了50 mg/g的赤霉素(GA3)和清水浸种后用不同浓度的聚乙二醇(0、40、80、120 mg/g)胁迫对番茄种子萌发及幼苗生长的影响。结果表明:40 mg/g的聚乙二醇胁迫对番茄种子的萌发率、发芽指数、幼苗株高、根长有促进作用,120 mg/g的聚乙二醇明显抑制种子的萌发率、发芽指数、幼苗株高、根长;不同浓度的聚乙二醇胁迫明显降低了幼苗的叶绿素含量。用50 mg/g GA3浸种后明显增加聚乙二醇胁迫下番茄种子的萌发率、发芽指数、幼苗株高、根长和叶绿素含量。用50 mg/g GA3浸种对番茄幼苗的生长有一定的促进作用。
DOI:10.3724/SP.J.1006.2009.00483URL [本文引用: 1]
用粳稻日本晴(Oryza sativa L.cv.Nipponbare),研究了盐胁迫对水稻种子萌发的抑制作用和赤霉酸(GA3)对盐胁迫的缓解作用;分别以H20(对照),5gL^-1NaCl(处理Ⅰ),5gL^-1NaCl+100μmolL^-1GA3(处理Ⅱ)培养水稻种苗48h,提取芽中的蛋白质,利用双向电泳(2-DE)和基质辅助激光解吸电离飞行时间质谱(MALDI—TOFMS)技术分析了水稻蛋白质组的变化。结果表明,在盐胁迫条件下,日本晴种子的萌发显著受到抑制,而GA3能显著缓解这种抑制作用;用Image Master软件分析2-DE凝胶,发现有4个蛋白质斑点表现出显著的变化,在盐胁迫下斑点S1、s2和s3表达下调而斑点S4消失,在GA3与盐共处理时,这4个蛋白质点的表达均有不同程度的恢复;经MALDI—TOFMS分析,其中2个蛋白质斑点(S1,S3)分别被鉴定为isoflavonereductase-like蛋白与葡萄糖磷酸变位酶,这些蛋白可能与GA3提高水稻耐盐性途径相关。
DOI:10.3724/SP.J.1006.2009.00483URL [本文引用: 1]
用粳稻日本晴(Oryza sativa L.cv.Nipponbare),研究了盐胁迫对水稻种子萌发的抑制作用和赤霉酸(GA3)对盐胁迫的缓解作用;分别以H20(对照),5gL^-1NaCl(处理Ⅰ),5gL^-1NaCl+100μmolL^-1GA3(处理Ⅱ)培养水稻种苗48h,提取芽中的蛋白质,利用双向电泳(2-DE)和基质辅助激光解吸电离飞行时间质谱(MALDI—TOFMS)技术分析了水稻蛋白质组的变化。结果表明,在盐胁迫条件下,日本晴种子的萌发显著受到抑制,而GA3能显著缓解这种抑制作用;用Image Master软件分析2-DE凝胶,发现有4个蛋白质斑点表现出显著的变化,在盐胁迫下斑点S1、s2和s3表达下调而斑点S4消失,在GA3与盐共处理时,这4个蛋白质点的表达均有不同程度的恢复;经MALDI—TOFMS分析,其中2个蛋白质斑点(S1,S3)分别被鉴定为isoflavonereductase-like蛋白与葡萄糖磷酸变位酶,这些蛋白可能与GA3提高水稻耐盐性途径相关。
DOI:10.3969/j.issn.2095-0756.2011.06.015URL [本文引用: 1]
<0.05),the same positive correlation with the moderate and serious disease area (<0.01),and no significant level was achieved with the mild disease area (>0.05). There was negative correlation between abnormal coldness in spring and the mild disease incidence rate (<0.01) and positive correlation with the moderate and severe disease incidence rate (<0.01). The year of severe abnormal coldness in spring matched with large acreage disease incidence of Valsa sordida and the more severe the coldness,the more obviously the acreage and incidence of moderate and serious disease increased. [Ch,4 tab. 10 ref.]
DOI:10.3969/j.issn.2095-0756.2011.06.015URL [本文引用: 1]
<0.05),the same positive correlation with the moderate and serious disease area (<0.01),and no significant level was achieved with the mild disease area (>0.05). There was negative correlation between abnormal coldness in spring and the mild disease incidence rate (<0.01) and positive correlation with the moderate and severe disease incidence rate (<0.01). The year of severe abnormal coldness in spring matched with large acreage disease incidence of Valsa sordida and the more severe the coldness,the more obviously the acreage and incidence of moderate and serious disease increased. [Ch,4 tab. 10 ref.]
DOI:10.3969/j.issn.1002-4093.2012.01.007URL [本文引用: 1]
分析研究了温度条件对不同花生品种种子活力的影响,以便掌握花生播种前最佳的预处理温度。试验通过生物化学、光谱分析和生理学综合进行。研究表明,在不同温度条件下,温度对不同花生品种的发芽率、发芽势、发芽指数影响极显著。在低于最适温度时,种子活力随预处理温度的升高而增强;在温度超过最适温度时,种子活力随温度升高逐渐降低直至丧失活力。六个品种分别通过响应曲面法拟合获得的最佳预处理温度为38.11℃、44.05℃、43.37℃、47.18℃、40.23℃和44.41℃。
DOI:10.3969/j.issn.1002-4093.2012.01.007URL [本文引用: 1]
分析研究了温度条件对不同花生品种种子活力的影响,以便掌握花生播种前最佳的预处理温度。试验通过生物化学、光谱分析和生理学综合进行。研究表明,在不同温度条件下,温度对不同花生品种的发芽率、发芽势、发芽指数影响极显著。在低于最适温度时,种子活力随预处理温度的升高而增强;在温度超过最适温度时,种子活力随温度升高逐渐降低直至丧失活力。六个品种分别通过响应曲面法拟合获得的最佳预处理温度为38.11℃、44.05℃、43.37℃、47.18℃、40.23℃和44.41℃。
DOI:10.3969/j.issn.1002-4093.2007.03.007URL [本文引用: 1]
以常温贮藏条件下的18个春花 生品种及其对应的秋花生品种为材料进行种子活力对比试验。结果表明,(1)经过常温干燥贮藏至第二年春播时,秋花生种子活力各项指标显著高于春花生,秋花 生种子的鲜重、发芽势、发芽率、发芽指数和活力指数分别是春花生的4.009倍、1.812倍、1.600倍、2.602倍和9.433倍;(2)无论春 花生或秋花生,活力指数各项指标品种间均有显著差异;(3)春花生与秋花生两者间活力指数呈显著正相关。
DOI:10.3969/j.issn.1002-4093.2007.03.007URL [本文引用: 1]
以常温贮藏条件下的18个春花 生品种及其对应的秋花生品种为材料进行种子活力对比试验。结果表明,(1)经过常温干燥贮藏至第二年春播时,秋花生种子活力各项指标显著高于春花生,秋花 生种子的鲜重、发芽势、发芽率、发芽指数和活力指数分别是春花生的4.009倍、1.812倍、1.600倍、2.602倍和9.433倍;(2)无论春 花生或秋花生,活力指数各项指标品种间均有显著差异;(3)春花生与秋花生两者间活力指数呈显著正相关。
URL [本文引用: 1]
叶片相对含水量随聚乙二醇(PEG)处理浓度的增加而依次降低,复水后它能恢复到对照水平;叶片质膜透性随PEG处理浓度的增加而依次增大,复水后它能不同程度地恢复;PEG处理时叶片膜脂的饱和脂肪酸含量增加,不饱和脂肪酸含量降低。复水后膜脂脂肪酸配比与复水前相似。干旱条件下叶片细胞的各类细胞器膜脂脂肪酸配比的变化与叶片总膜脂的变化相似;在轻度干旱条件下的叶绿体Mg~(++)-ATP酶活力低于对照,复水后能迅速恢复到对照水平,中度干旱条件下Mg~(++)-ATP酶活力明显高于对照,但复水后均有不周程度的恢复。
URL [本文引用: 1]
叶片相对含水量随聚乙二醇(PEG)处理浓度的增加而依次降低,复水后它能恢复到对照水平;叶片质膜透性随PEG处理浓度的增加而依次增大,复水后它能不同程度地恢复;PEG处理时叶片膜脂的饱和脂肪酸含量增加,不饱和脂肪酸含量降低。复水后膜脂脂肪酸配比与复水前相似。干旱条件下叶片细胞的各类细胞器膜脂脂肪酸配比的变化与叶片总膜脂的变化相似;在轻度干旱条件下的叶绿体Mg~(++)-ATP酶活力低于对照,复水后能迅速恢复到对照水平,中度干旱条件下Mg~(++)-ATP酶活力明显高于对照,但复水后均有不周程度的恢复。
[本文引用: 1]
DOI:10.3969/j.issn.1001-7283.2009.01.022URL [本文引用: 1]
在室内培养箱条件下,研究了不 同成熟度花生种子萌动期低温对花生出苗及幼苗生长发育的影响。结果表明:不同成熟度的种子受萌动期间低温的影响不同。种子成熟度好,活力强,抗低温能力 强,出苗率高,幼苗根系发达、生长迅速整齐,单株叶面积大,有利于形成壮苗;叶片NR活性强,叶绿素含量高,有利于干物质积累。种子成熟度差,萌动期遭遇 低温受影响较大,不利于壮苗,势必影响中后期植株的生长发育,降低产量。
DOI:10.3969/j.issn.1001-7283.2009.01.022URL [本文引用: 1]
在室内培养箱条件下,研究了不 同成熟度花生种子萌动期低温对花生出苗及幼苗生长发育的影响。结果表明:不同成熟度的种子受萌动期间低温的影响不同。种子成熟度好,活力强,抗低温能力 强,出苗率高,幼苗根系发达、生长迅速整齐,单株叶面积大,有利于形成壮苗;叶片NR活性强,叶绿素含量高,有利于干物质积累。种子成熟度差,萌动期遭遇 低温受影响较大,不利于壮苗,势必影响中后期植株的生长发育,降低产量。
URL [本文引用: 1]
正 低温干旱是我国北方大花生区播种期的主要自然灾害。播种稍早,遇寒流即发生大面积低温烂种;播种稍晚,则易发生春旱,落干缺苗。晚熟大花生由于积温不足,还易出现荚果不饱,造成大幅度减产。要改变这种状况,一是改变花生的生长环境,二是改进花生对不良
URL [本文引用: 1]
正 低温干旱是我国北方大花生区播种期的主要自然灾害。播种稍早,遇寒流即发生大面积低温烂种;播种稍晚,则易发生春旱,落干缺苗。晚熟大花生由于积温不足,还易出现荚果不饱,造成大幅度减产。要改变这种状况,一是改变花生的生长环境,二是改进花生对不良
DOI:10.3724/SP.J.1006.2012.01247URL [本文引用: 1]
为探索冬小麦抗寒性的分子机制,以168个花培3号×豫麦57的双单倍体株系为作图群体,利 用已构建含有324个SSR标记的遗传图谱,对电导法测定低温(-18℃)处理后的叶片膜透性进行QTL定位。利用完全区间作图法,在3种环境下共检测到 21个与叶片膜透性相关的加性QTLs,分布于1B、2A、3A、3B、5B、6A、6B、6D、7B和7D染色体上,其中4个位点(qCMP—1B- 1、qCMP-3B-2、qCMP-5B-1和qCMP-5B-4)遗传贡献率大于10%,属主效基因,其余QTL的遗传贡献率较小,属微效基因。3种环 境条件下在5B染色体的Xgwm213-Xswes861.2区间检测到共同位点,与Xswes861.2的遗传距离为0cM,其中qCMP-5B- 1(环境1)和qCMP-5B-4(环境3)的贡献率高达17.5%和14.O%。研究结果对于小麦抗寒标记选择和抗寒育种具有应用价值。
DOI:10.3724/SP.J.1006.2012.01247URL [本文引用: 1]
为探索冬小麦抗寒性的分子机制,以168个花培3号×豫麦57的双单倍体株系为作图群体,利 用已构建含有324个SSR标记的遗传图谱,对电导法测定低温(-18℃)处理后的叶片膜透性进行QTL定位。利用完全区间作图法,在3种环境下共检测到 21个与叶片膜透性相关的加性QTLs,分布于1B、2A、3A、3B、5B、6A、6B、6D、7B和7D染色体上,其中4个位点(qCMP—1B- 1、qCMP-3B-2、qCMP-5B-1和qCMP-5B-4)遗传贡献率大于10%,属主效基因,其余QTL的遗传贡献率较小,属微效基因。3种环 境条件下在5B染色体的Xgwm213-Xswes861.2区间检测到共同位点,与Xswes861.2的遗传距离为0cM,其中qCMP-5B- 1(环境1)和qCMP-5B-4(环境3)的贡献率高达17.5%和14.O%。研究结果对于小麦抗寒标记选择和抗寒育种具有应用价值。
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1006.2017.01677URL [本文引用: 1]
水杨酸(SA)、脯氨酸(Pro)和γ-氨基丁酸(GABA)在植物生长发育以及抵御逆境胁迫中起重要作用.为了研究三者复配对盐胁迫下水稻产量是否具有协同增效作用, 本文采用三元二次通用旋转组合设计, 建立了 SA、Pro 和GABA复配剂与盐胁迫下水稻产量之间的数学模型, 通过频数分析获得SA、Pro与GABA复配优化组合方案.结果表明, 二次模型拟合较好, 对盐胁迫下水稻产量的影响程度为SA>GABA>Pro, SA与GABA, Pro与GABA之间交互作用对盐胁迫下水稻产量有显著影响.模拟寻优获得提高盐胁迫下水稻产量10%以上的最佳复配组合为SA 0.44~0.51 mmol L-1、Pro 27.63~31.20 mmol L-1、GABA 3.55~4.28 mmol L-1.外源SA、Pro与GABA单剂和复配剂对盐敏感品种牡丹江30的调控作用大于耐盐品种龙稻5号.SA或Pro与GABA复配对盐胁迫下水稻产量的影响具有协同作用,而 SA 与 Pro 复配效果与品种耐盐性差异有关.SA、Pro 和 GABA 三者复配效果优于两两复配, 任意单剂与其他2种外源物质复配对水稻产量的影响均存在协同作用.
DOI:10.3724/SP.J.1006.2017.01677URL [本文引用: 1]
水杨酸(SA)、脯氨酸(Pro)和γ-氨基丁酸(GABA)在植物生长发育以及抵御逆境胁迫中起重要作用.为了研究三者复配对盐胁迫下水稻产量是否具有协同增效作用, 本文采用三元二次通用旋转组合设计, 建立了 SA、Pro 和GABA复配剂与盐胁迫下水稻产量之间的数学模型, 通过频数分析获得SA、Pro与GABA复配优化组合方案.结果表明, 二次模型拟合较好, 对盐胁迫下水稻产量的影响程度为SA>GABA>Pro, SA与GABA, Pro与GABA之间交互作用对盐胁迫下水稻产量有显著影响.模拟寻优获得提高盐胁迫下水稻产量10%以上的最佳复配组合为SA 0.44~0.51 mmol L-1、Pro 27.63~31.20 mmol L-1、GABA 3.55~4.28 mmol L-1.外源SA、Pro与GABA单剂和复配剂对盐敏感品种牡丹江30的调控作用大于耐盐品种龙稻5号.SA或Pro与GABA复配对盐胁迫下水稻产量的影响具有协同作用,而 SA 与 Pro 复配效果与品种耐盐性差异有关.SA、Pro 和 GABA 三者复配效果优于两两复配, 任意单剂与其他2种外源物质复配对水稻产量的影响均存在协同作用.
DOI:10.14088/j.cnki.issn0439-8114.2016.06.034URL [本文引用: 1]
以矮牵牛[Petunia hybrida(J.D.Hooker)Vilmorin]H株系为试验材料,利用人工气候箱进行低温胁迫试验,调查低温胁迫对矮牵牛叶片中花青素和渗透调节物质游离脯氨酸、可溶性糖含量的影响。结果表明,花青素在低温处理前期2 h和中期24 h时相对含量变化较对照无显著性差异,当胁迫增至120 h,其相对含量显著提高;游离脯氨酸含量在低温处理前期2 h时含量变化较对照无明显变化,而在中期24 h达到峰值,此后表现为下降趋势;可溶性糖含量在低温处理前期2 h时含量较对照有明显提高,此后含量略微上升,直至后期120 h一直保持在这一较高水平。研究表明,矮牵牛可能通过积累花青素、渗透调节物质来提高植株的抗寒能力,从而抵御低温胁迫。
DOI:10.14088/j.cnki.issn0439-8114.2016.06.034URL [本文引用: 1]
以矮牵牛[Petunia hybrida(J.D.Hooker)Vilmorin]H株系为试验材料,利用人工气候箱进行低温胁迫试验,调查低温胁迫对矮牵牛叶片中花青素和渗透调节物质游离脯氨酸、可溶性糖含量的影响。结果表明,花青素在低温处理前期2 h和中期24 h时相对含量变化较对照无显著性差异,当胁迫增至120 h,其相对含量显著提高;游离脯氨酸含量在低温处理前期2 h时含量变化较对照无明显变化,而在中期24 h达到峰值,此后表现为下降趋势;可溶性糖含量在低温处理前期2 h时含量较对照有明显提高,此后含量略微上升,直至后期120 h一直保持在这一较高水平。研究表明,矮牵牛可能通过积累花青素、渗透调节物质来提高植株的抗寒能力,从而抵御低温胁迫。
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.1005-3395.2000.3.009URL [本文引用: 1]
Drought, salt and chilling effects on proline accumulation were studied in 3-leaf-stage seedlings of three rice (Oryza sativa L.) varieties: IAC (sensitive to drought and salt, and tolerant to low temperature), Dahuanggu (salt-resistant but sensitive to low temperature) and Guixi (drought-tolerant). Treatments were carried out at -0.6 MPa PEG (simulation) for drought, 0.15 mol/L NaC1 for salinity, and 6 ℃ for chilling stresses. Proline content in aboveground part of rice seedlings were tested. The results showed that proline accumulation in rice seedlings of various varieties was seen under all stresses,and the proline content increased with increasing time of stress.Sensitive variety accumulated more proline content than tolerant one. It seems,however, that proline accumulation can not be used as a screening index of resistance to stres ses in rice seedlings.
DOI:10.3969/j.issn.1005-3395.2000.3.009URL [本文引用: 1]
Drought, salt and chilling effects on proline accumulation were studied in 3-leaf-stage seedlings of three rice (Oryza sativa L.) varieties: IAC (sensitive to drought and salt, and tolerant to low temperature), Dahuanggu (salt-resistant but sensitive to low temperature) and Guixi (drought-tolerant). Treatments were carried out at -0.6 MPa PEG (simulation) for drought, 0.15 mol/L NaC1 for salinity, and 6 ℃ for chilling stresses. Proline content in aboveground part of rice seedlings were tested. The results showed that proline accumulation in rice seedlings of various varieties was seen under all stresses,and the proline content increased with increasing time of stress.Sensitive variety accumulated more proline content than tolerant one. It seems,however, that proline accumulation can not be used as a screening index of resistance to stres ses in rice seedlings.
DOI:10.11686/cyxb20140223URLMagsci [本文引用: 1]
<p>以苦马豆为实验材料,在8种不同浓度NaCl (0, 80, 160, 240, 320, 400, 480和 560 mmol/L)胁迫下,分别向叶面喷施细胞分裂素类物质6-BA和脱落酸(ABA),通过2周的胁迫生长后,对其根冠比、脯氨酸以及脯氨酸代谢酶活性进行测定,进行外源植物激素对苦马豆耐盐性影响的研究,为进一步培育耐盐牧草提供一定的理论基础。结果表明,外源植物激素可以提高苦马豆的抗盐性,且根系中酶活性对植物激素的敏感程度大于叶片。植物激素6-BA比ABA的作用更为显著。外源植物激素使苦马豆幼苗中脯氨酸不再是抗逆性的主要检测指标。</p>
DOI:10.11686/cyxb20140223URLMagsci [本文引用: 1]
<p>以苦马豆为实验材料,在8种不同浓度NaCl (0, 80, 160, 240, 320, 400, 480和 560 mmol/L)胁迫下,分别向叶面喷施细胞分裂素类物质6-BA和脱落酸(ABA),通过2周的胁迫生长后,对其根冠比、脯氨酸以及脯氨酸代谢酶活性进行测定,进行外源植物激素对苦马豆耐盐性影响的研究,为进一步培育耐盐牧草提供一定的理论基础。结果表明,外源植物激素可以提高苦马豆的抗盐性,且根系中酶活性对植物激素的敏感程度大于叶片。植物激素6-BA比ABA的作用更为显著。外源植物激素使苦马豆幼苗中脯氨酸不再是抗逆性的主要检测指标。</p>
DOI:10.1105/tpc.105.036707URLPMID:16339853 [本文引用: 1]
Gibberellic acid (GA) promotes germination, stem/hypocotyl elongation, and leaf expansion during seedling development. Using activation-tagging mutagenesis, we identified a mutation, sob2-D (for suppressor of phytochromeB-4 [phyB-4]#2 dominant), which suppresses the long-hypocotyl phenotype of a phyB missense allele, phyB-4. This mutant phenotype is caused by the overexpression of an APETALA2 transcription factor, SOB2, also called DRN-like. SOB2/DRN-like transcript is not detectable in wild-type seedling or adult tissues via RT-PCR analysis, suggesting that SOB2/DRN-like may not be involved in seedling development under normal conditions. Adult sob2-D phyB-4 plants have curled leaves and club-like siliques, resembling plants that overexpress a closely related gene, LEAFY PETIOLE (LEP). Hypocotyls of a LEP-null allele, lep-1, are shorter in the light and dark, suggesting LEP involvement in seedling development. This aberrant hypocotyl phenotype is due at least in part to a delay in germination. In addition, lep-1 is less responsive to GA and more sensitive to the GA biosynthesis inhibitor paclobutrazol, indicating that LEP is a positive regulator of GA-induced germination. RT-PCR shows that LEP transcript accumulates in wild-type seeds during imbibition and germination, and the transcript levels of REPRESSOR OF ga1-3-LIKE2 (RGL2), a negative regulator of GA signaling during germination, is unaffected in lep-1. These results suggest LEP is a positive regulator of GA-induced germination acting independently of RGL2. An alternative model places LEP downstream of RGL2 in the GA-signaling cascade.
URL [本文引用: 1]
为了比较分析乌饭树和短尾越橘种子的发芽特性,就不同温度条件下不同浓度赤霉素GA3溶液浸种处理对乌饭树和短尾越橘种子萌发的影响情况进行了恒温培养试验。结果表明:GA3溶液对打破乌饭树和短尾越橘种子休眠、提高发芽率、缩短发芽时间有显著的效果,发芽率比未经处理的高出3~5倍;处理后的乌饭树和短尾越橘种子对温度有很高的敏感度,25℃是最佳发芽温度,温度升高发芽率则降低,25℃时的发芽率是27℃时的3~5倍,是30℃时的10至几十倍;GA3溶液的浓度变化对乌饭树种子发芽率的影响不大,在25℃恒温条件下,用浓度分别为200、600、1 000 mg/L的GA3溶液浸种后的种子其最高发芽率分别为60%、65%和64%;而短尾越橘种子的发芽率随GA3溶液浓度不同而有所变化,用浓度为1 000 mg/L的GA3溶液处理后的种子其发芽率始终最高,在25℃时,用浓度分别为200 mg/L、600 mg/L、1 000 mg/L的GA3溶液浸种后的最高发芽率分别为41%、25%和57%。(4)提高温度,对两者的发芽都有抑制作用,30℃时,乌饭树几乎不发芽,而短尾越橘最终发芽率为16%。说明短尾越橘种子休眠更深,这可能与其自身种皮密实、坚硬等物理特性有关。
URL [本文引用: 1]
为了比较分析乌饭树和短尾越橘种子的发芽特性,就不同温度条件下不同浓度赤霉素GA3溶液浸种处理对乌饭树和短尾越橘种子萌发的影响情况进行了恒温培养试验。结果表明:GA3溶液对打破乌饭树和短尾越橘种子休眠、提高发芽率、缩短发芽时间有显著的效果,发芽率比未经处理的高出3~5倍;处理后的乌饭树和短尾越橘种子对温度有很高的敏感度,25℃是最佳发芽温度,温度升高发芽率则降低,25℃时的发芽率是27℃时的3~5倍,是30℃时的10至几十倍;GA3溶液的浓度变化对乌饭树种子发芽率的影响不大,在25℃恒温条件下,用浓度分别为200、600、1 000 mg/L的GA3溶液浸种后的种子其最高发芽率分别为60%、65%和64%;而短尾越橘种子的发芽率随GA3溶液浓度不同而有所变化,用浓度为1 000 mg/L的GA3溶液处理后的种子其发芽率始终最高,在25℃时,用浓度分别为200 mg/L、600 mg/L、1 000 mg/L的GA3溶液浸种后的最高发芽率分别为41%、25%和57%。(4)提高温度,对两者的发芽都有抑制作用,30℃时,乌饭树几乎不发芽,而短尾越橘最终发芽率为16%。说明短尾越橘种子休眠更深,这可能与其自身种皮密实、坚硬等物理特性有关。
DOI:10.3969/j.issn.1001-4705.2012.11.030URL [本文引用: 1]
使用不同浓度的赤霉素(GA)溶液对来自4个不同产区的秋葵种子进行处理以探讨赤霉素对秋葵 种子发芽的影响。结果表明:赤霉素可提高秋葵种子的发芽集中度、发芽势和发芽率,还可缩短发芽时间;赤霉素浓度为60mg/L时,种子的发芽集中度最高, 发芽所需时间最短;当赤霉素浓度超过60mg/L时,种子发芽集中度开始下降,发芽所需时间开始延长。
DOI:10.3969/j.issn.1001-4705.2012.11.030URL [本文引用: 1]
使用不同浓度的赤霉素(GA)溶液对来自4个不同产区的秋葵种子进行处理以探讨赤霉素对秋葵 种子发芽的影响。结果表明:赤霉素可提高秋葵种子的发芽集中度、发芽势和发芽率,还可缩短发芽时间;赤霉素浓度为60mg/L时,种子的发芽集中度最高, 发芽所需时间最短;当赤霉素浓度超过60mg/L时,种子发芽集中度开始下降,发芽所需时间开始延长。
URL [本文引用: 1]
研究了赤霉素(GA3)和发芽温度对圆叶牵牛种子萌发的影响.结 果表明:28℃较高温度处理圆叶牵牛种子,发芽指数和发芽率高于23℃较低温度处理,但未达到显著水平.200 mg/L GA3浸种可显著提高圆叶牵牛种子的发芽率和发芽指数,发芽起始时间和终止时间也有所提前.
URL [本文引用: 1]
研究了赤霉素(GA3)和发芽温度对圆叶牵牛种子萌发的影响.结 果表明:28℃较高温度处理圆叶牵牛种子,发芽指数和发芽率高于23℃较低温度处理,但未达到显著水平.200 mg/L GA3浸种可显著提高圆叶牵牛种子的发芽率和发芽指数,发芽起始时间和终止时间也有所提前.
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1142.2012.20204URL [本文引用: 1]
Jatropha curcas L.is an energy plant of the Euphorbiaceae family.Using seeds and seedlings of J.curcas as materials,the effect of different concentrations of gibberellin or calcium treatment,or combination treatment of different concentrations of gibberellin,calcium, and betaine on seed germination and resistance of seedlings to low temperature or drought stress were investigated.The application of 10 mg/L gibberellin or 10 mmol/L calcium not only improved germination percentage of J.curcas seeds under normal conditions (26℃),low temperature (18℃),and drought stress (5% PEG6000),but also alleviated electrolyte leakage and accumulation of MDA in J.curcas seedlings under low temperature (2℃) and drought stress (25% PEG6000) compared with the control. In addition,combination treatments with 10 mg/L gibberellin,5 mmol/L calcium,and 15 mmol/L betaine also enhanced germination percentage of J.curcas seeds under normal conditions (26℃),low temperature (18℃),and drought stress (5% PEG6000),and alleviated electrolyte leakage and accumulation of MDA in J.curcas seedlings under low temperature (2℃) and drought stress (25% PEG6000),illustrating that treatment with 10 mg/L gibberellin or 10 mmol/L calcium,or combination treatments of 10 mg/L gibberellin,5 mmol/L calcium,and 15 mmol/L betaine increased germination percentage of J.curcas seeds under low temperature and drought stress,and resistance of seedlings to low temperature or drought stress.
DOI:10.3724/SP.J.1142.2012.20204URL [本文引用: 1]
Jatropha curcas L.is an energy plant of the Euphorbiaceae family.Using seeds and seedlings of J.curcas as materials,the effect of different concentrations of gibberellin or calcium treatment,or combination treatment of different concentrations of gibberellin,calcium, and betaine on seed germination and resistance of seedlings to low temperature or drought stress were investigated.The application of 10 mg/L gibberellin or 10 mmol/L calcium not only improved germination percentage of J.curcas seeds under normal conditions (26℃),low temperature (18℃),and drought stress (5% PEG6000),but also alleviated electrolyte leakage and accumulation of MDA in J.curcas seedlings under low temperature (2℃) and drought stress (25% PEG6000) compared with the control. In addition,combination treatments with 10 mg/L gibberellin,5 mmol/L calcium,and 15 mmol/L betaine also enhanced germination percentage of J.curcas seeds under normal conditions (26℃),low temperature (18℃),and drought stress (5% PEG6000),and alleviated electrolyte leakage and accumulation of MDA in J.curcas seedlings under low temperature (2℃) and drought stress (25% PEG6000),illustrating that treatment with 10 mg/L gibberellin or 10 mmol/L calcium,or combination treatments of 10 mg/L gibberellin,5 mmol/L calcium,and 15 mmol/L betaine increased germination percentage of J.curcas seeds under low temperature and drought stress,and resistance of seedlings to low temperature or drought stress.
DOI:10.3969/j.issn.1673-1409.2006.04.002URL [本文引用: 1]
以大棚栽培的3年生早紫葡萄(Vitis vinifera )为材料,研究了在花前10 d喷施不同浓度的赤霉素(GA3)溶液对果实品质、无核率及座果率的影响.结果表明:喷施赤霉素(GA3)溶液后都能达到无核化的效果,其中以50 mg/L的赤霉素(GA3)溶液无核处理的效果最好,无核率可达93.33%;果实的可溶性糖及可溶性固性物含量也均最高,而且其着色度、表面光洁度等外观品质也最好.
DOI:10.3969/j.issn.1673-1409.2006.04.002URL [本文引用: 1]
以大棚栽培的3年生早紫葡萄(Vitis vinifera )为材料,研究了在花前10 d喷施不同浓度的赤霉素(GA3)溶液对果实品质、无核率及座果率的影响.结果表明:喷施赤霉素(GA3)溶液后都能达到无核化的效果,其中以50 mg/L的赤霉素(GA3)溶液无核处理的效果最好,无核率可达93.33%;果实的可溶性糖及可溶性固性物含量也均最高,而且其着色度、表面光洁度等外观品质也最好.
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.1000-2561.2014.09.021URL [本文引用: 1]
以海南主栽杧果品种贵妃杧为试材,研究了赤霉素(GA3)和氯吡脲(CPPU或KT-30)对其产量和果实品质的影响。结果表明:(1)GA3处理明显增加产量,且不同程度地改善果实品质,以GA3150 mg/L和GA3250 mg/L效果较好,两者的产量、果形指数、单果重、可食率、糖酸比(显著降低可滴定酸含量)均显著高于对照;(2)CPPU处理促进果实膨大的效果比GA3处理明显,且可食率和果实硬度显著高于对照,但产量增加不明显,具有疏果的作用,明显增加畸形果数量,浓度过高会使糖酸比(显著增加可滴定酸含量)明显下降,以较低浓度的CPPU 5 mg/L和CPPU 10 mg/L为宜;(3)GA3150 mg/L+CPPU 5 mg/L的混合处理对果实膨大的效果明显优于单一处理,但增加产量不明显,与单用GA3150 mg/L处理相比,明显降低了产量、可溶性固形物、可溶性糖含量和糖酸比(显著增加可滴定酸含量);(4)CPPU处理和较低浓度的GA3处理(GA350 mg/L和GA3150 mg/L)延缓了果皮叶绿素的降解,较高浓度(GA3250 mg/L和GA3500 mg/L)则相反,GA3和CPPU处理均促进了果皮类胡萝卜素的积累,较高浓度的GA3处理促进了果皮花青素的合成,CPPU和混合处理抑制了果皮花青素的合成。该研究为GA3和CPPU在海南贵妃杧种植中的合理应用提供了科学依据。
DOI:10.3969/j.issn.1000-2561.2014.09.021URL [本文引用: 1]
以海南主栽杧果品种贵妃杧为试材,研究了赤霉素(GA3)和氯吡脲(CPPU或KT-30)对其产量和果实品质的影响。结果表明:(1)GA3处理明显增加产量,且不同程度地改善果实品质,以GA3150 mg/L和GA3250 mg/L效果较好,两者的产量、果形指数、单果重、可食率、糖酸比(显著降低可滴定酸含量)均显著高于对照;(2)CPPU处理促进果实膨大的效果比GA3处理明显,且可食率和果实硬度显著高于对照,但产量增加不明显,具有疏果的作用,明显增加畸形果数量,浓度过高会使糖酸比(显著增加可滴定酸含量)明显下降,以较低浓度的CPPU 5 mg/L和CPPU 10 mg/L为宜;(3)GA3150 mg/L+CPPU 5 mg/L的混合处理对果实膨大的效果明显优于单一处理,但增加产量不明显,与单用GA3150 mg/L处理相比,明显降低了产量、可溶性固形物、可溶性糖含量和糖酸比(显著增加可滴定酸含量);(4)CPPU处理和较低浓度的GA3处理(GA350 mg/L和GA3150 mg/L)延缓了果皮叶绿素的降解,较高浓度(GA3250 mg/L和GA3500 mg/L)则相反,GA3和CPPU处理均促进了果皮类胡萝卜素的积累,较高浓度的GA3处理促进了果皮花青素的合成,CPPU和混合处理抑制了果皮花青素的合成。该研究为GA3和CPPU在海南贵妃杧种植中的合理应用提供了科学依据。

{kind=link}
{kind=link}
{kind=link}
{kind=link}