 ,1,*, 张丽娜2,3, 潘映红,2,3,*
,1,*, 张丽娜2,3, 潘映红,2,3,*Proteomic Analysis of the Effect of 2,4-Epibrassinolide on Rice Seedlings Response to Cold Stress
WANG Dao-Ping1,2, XU Jiang2, MU Yong-Ying2,3, YAN Wen-Xiu2,3, ZHAO Meng-Jie3, MA Bo3, LI Qun,1,*, ZHANG Li-Na2,3, PAN Ying-Hong,2,3,*通讯作者:
第一联系人:
收稿日期:2017-08-29接受日期:2018-03-18网络出版日期:2018-06-12
| 基金资助: |
Received:2017-08-29Accepted:2018-03-18Online:2018-06-12
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (1748KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王道平, 徐江, 牟永莹, 闫文秀, 赵梦洁, 马博, 李群, 张丽娜, 潘映红. 表油菜素内酯影响水稻幼苗响应低温胁迫的蛋白质组学分析[J]. 作物学报, 2018, 44(6): 897-908. doi:10.3724/SP.J.1006.2018.00897
WANG Dao-Ping, XU Jiang, MU Yong-Ying, YAN Wen-Xiu, ZHAO Meng-Jie, MA Bo, LI Qun, ZHANG Li-Na, PAN Ying-Hong.
作为植物内源信号的乙烯、生长素、细胞分裂素、脱落酸、赤霉素、油菜素内酯(brassinosteroids, BRs)等激素在植物生长发育和逆境响应中发挥着重要的调节作用[1,2]。其中BRs是一类在体内分布广泛但不长距离运输的甾体激素, 这类激素全面参与植物的细胞扩增、分化、生长发育、衰老以及光形态建成等生物过程[3,4,5], 对植物响应高温、低温、干旱和盐胁迫均有调控作用[5,6]。有研究用BRs类植物生长调节剂表油菜素内酯(2,4-Epibrassinolide, EBR)处理拟南芥和欧洲油菜的幼苗, 增强了植物对干旱和低温胁迫的耐受性[7]; BRs生物合成基因AtDWF4在种子中过表达也可提高拟南芥幼苗的耐冷性[8]。在苦瓜中发现BRs可以通过增加植物鲜重、叶绿素含量、可溶性蛋白质含量以及增强超氧化物歧化酶(SOD)活性等来降低低温胁迫对苦瓜生长的影响[9]。对玉米的研究发现, EBR处理幼苗可增加株高, 提高叶绿素和可溶性蛋白质含量, 并显著增强植株的低温耐受性[10]。番茄叶面喷施EBR后, 可缓解低温对光合作用和氮代谢的影响, 提高低温弱光胁迫下番茄的鲜重和干重[11]。此外, BRs还可以通过增强抗氧化系统和维持光系统II来缓解辣椒中由低温诱导产生的氧化胁迫[12]。这些研究均表明BRs可以增强植物的低温抗性, 但有关调控机制的研究尚不多见。从现有的BRs影响植物低温胁迫蛋白质组学研究结果来看, 绿豆中的上调蛋白质主要参与甲硫氨酸的同化、ATP合成、细胞壁构建和应激反应[13]; 辣椒的差异蛋白主要参与催化及活性调节、蛋白质结构性分子活性、能量转运过程和蛋白质结合等生物过程及光合固碳途径、氧化磷酸化途径、谷胱甘肽代谢途径、苯丙素生物合成途径、苯丙氨酸代谢途径和核糖体途径等[12]。上述结果提示, BRs可能通过调节多种蛋白质的丰度来影响植物对低温胁迫的响应。
水稻是一种被广泛研究的重要作物, 与水稻相关的蛋白质组研究主要集中在响应盐胁迫、干旱胁迫等方面, 有关低温胁迫的研究相对较少。对1周龄水稻幼苗进行低温处理后发现, 膜结构受损, 代谢变化剧烈, 水稻防御系统在低温胁迫下被激活[14]。2周龄幼苗低温胁迫6 h后, 一些蛋白质包括应激抗性蛋白质和负调控蛋白质等出现上调[15]; 低温胁迫12 h后, 根部组织的烯醇化酶、甘油醛-3-磷酸脱氢酶、核苷二磷酸激酶、抗坏血酸过氧化物酶等的蛋白丰度可增加2倍以上[16]; 低温胁迫24 h后叶片的差异蛋白主要包括蛋白质生物合成因子、分子伴侣、细胞壁组分生物合成酶、抗氧化/解毒酶以及与能量代谢通路相关的蛋白质, 这些蛋白主要定位于叶绿体中[17]; 幼苗经48 h低温胁迫, 能量代谢和应激相关蛋白质明显上调, 而防御相关蛋白质下调[18]。进一步对水稻幼苗12~14°C处理48 h、72 h和96 h的研究发现, 差异蛋白质主要是物质转运、光合作用、前体代谢产物和能量代谢相关类蛋白, 组蛋白和维生素B生物合成相关蛋白的丰度明显受到冷胁迫的影响[19]。这些蛋白质组学研究结果说明水稻幼苗受不同程度低温胁迫处理后, 光合作用、能量代谢和应激相关蛋白会出现明显的变化, 但是目前仍然不清楚BRs如何在蛋白质水平上影响水稻幼苗对低温胁迫的响应。
本研究采用表油菜素内酯(EBR)处理萌发的水稻种子, 利用蛋白质组学技术筛选和分析可能与EBR影响水稻幼苗响应低温胁迫有关的蛋白, 旨在为进一步研究BRs影响植物响应低温胁迫机制提供新的线索。
1 材料与方法
1.1 实验材料
日本晴种子在16 h光照(26°C)/8 h黑暗(22°C)条件下萌发培养7 d后, 用0.1 mg L-1 EBR浸泡12 h, 对照为蒸馏水浸泡, 浸泡后的萌发种子分别在16 h光照/8 h黑暗的4°C和26°C培养箱中培养24 h后取芽, EBR处理4°C培养样品, 单独4°C培养样品, EBR处理26°C培养样品和单独26°C培养样品分别标记为4B、4、26B和26, -80°C保存备用。1.2 叶绿素测定方法
取每个样品新鲜芽0.2 g, -80°C冷冻1 h, 加入10 mL80%丙酮溶液, 50°C水浴3 h至叶片发白。以80%丙酮为空白, 分别测定叶绿素提取液在663 nm和645 nm波长的光密度, 按下列公式计算叶绿素a和b含量(D645和D663表示在645 nm和663 nm波长下的光密度, V为定容体积10 mL, W为称样量0.2 g)。叶绿素a (mg g-1) = (12.72D663 - 2.69D645) V/(W1000)
叶绿素b (mg g-1) = (22.88D645 - 4.68D663) V/(W1000)
1.3 蛋白提取及酶切
称取0.2 g样品用细胞破碎仪20 Hz s-1振荡破碎, 加入0.4 mL提取缓冲液(8 mol L-1 Urea, 2 mmol L-1 EDTA, 20 mmol L-1 CaCl2, 200 mmol L-1 NaCl, 100 mmol L-1 Tris-HCl, pH 8.1), 13 000×g离心30 min; 转移上清液到10 K超滤管内超滤离心(13 000×g, 30 min, 下同), 弃滤出液, 加入200 μL现配的积二硫苏糖醇(dithiothreitol, DTT)溶液(50 mmol L-1 DTT, 8 mol L-1 Urea, 100 mmol L-1 Tris-HCl, pH 8.1), 30°C孵育60 min; 超滤离心除去滤出液, 分3次加入200 μL尿素缓冲液(8 mol L-1 Urea, 100 mmol L-1 Tris-HCl, pH 8.1)洗涤超滤, 加入100 μL现配碘乙酰胺(iodoacetamide, IAA)溶液(50 mmol L-1 IAA, 8 mol L-1 Urea, 100 mmol L-1 Tris-HCl, pH 8.1)暗置30 min; 超滤离心除去滤出液, 用100 μL尿素缓冲液(8 mol L-1 Urea, H2O)洗涤超滤3次, 加入200 μL NH4HCO3溶液(50 mmol L-1)洗涤超滤2次; 加入50 μL蛋白酶解液(含0.5 μg Trypsin, 50 mmol L-1 NH4HCO3), 同时更换超滤管外管, 37°C酶切10 h, 超滤离心得到质谱分析样品。1.4 质谱分析
液相色谱分离在Easy nLC 1000纳升级液相色谱仪(Thermo Fisher公司产品)上完成。流动相A为0.1%甲酸水(Formic Acid, FA), 双蒸水; 流动相B: 0.1% FA, CAN。设置流动相的梯度为流速0.4 mL min-1, 3%~6% 3 min, 6%~22% 78 min, 22%~100% 1 min, 100% 8 min。质谱分析在依靠静电场轨道离子阱(Orbitrap)检测离子质量的Q Exactive Plus (QE Plus, Thermo Fisher公司产品)质谱仪上完成, 采集模式为数据依赖型(data dependent acquisition, DDA)。设一级质谱的分辨率为70 000, 扫描范围为300~1800 m/z, 自动增益控制值为3e6, 注入时间为50 ms。二级质谱的分辨率为17 500, 自动增益控制值1e5, 注入时间45 ms, 碰撞能量27 NCE。二级采集的标准以前20个最强离子为母离子, 再进行Orbitrap检测。每个样品进行3次质谱技术重复。1.5 蛋白质鉴定及数据库搜索
利用Proteome Discoverer 2.0定性分析软件(Thermo Fisher公司产品), 对质谱的原始数据RAW文件进行定性分析。检索引擎设定为SEQUEST HT, 检索数据库为植物基因组中水稻数据库(https:// phytozome.jgi.doe.gov/, 版本为Osativa_323_v7.0. protein)。设定如下定性搜库参数, 即肽段置信度为高, 最多蛋白漏切位点数2个, 肽段的长度范围6~144个氨基酸, 母离子质量偏差为±10 μmol L-1, 碎片离子质量偏差0.02 Da, 固定修饰选择半胱氨酸(Cys)碘乙酰胺化(carbamidomethy/+57.021 Da), 可变修饰选择甲硫氨酸(Met)氧化(oxidation/+15.995 Da)和N-乙酰化(Acetyl/+42.011 Da), 肽段搜库假阳性率FDR为1%。将每个样品3次重复的数据一起检索, 得到每个样品的定性结果, 分别以Excel表格的形式输出。采用Maxquant软件(http://www.coxdocs.org/, 版本为1.3.0.5) 定量分析, 检索数据库同上。用全部样品(各自重复3次)的质谱数据一起进行非标定量(label-free)分析, 设定如下参数, 即胰蛋白酶为默认的蛋白酶, 2个漏切位点, 固定修饰选择半胱氨酸碘乙酰胺化, 可变修饰选择甲硫氨酸氧化和N-乙酰化, 先驱离子质量偏差为±20 μmol L-1, 碎片离子质量偏差0.02 Da, 能够检测到的肽段长度的最小值是7个氨基酸, 肽段搜库假阳性率FDR为1%。4个样品的相对定量结果以Excel表格的形式输出, 供后续比较分析。
1.6 数据相关性分析
使用SPSS 22.0数据处理软件(https://www.ibm. com/analytics)对各样品定量数据进行皮尔逊(Pearson)相关性分析。通过绝对值的大小表示样品间线性相关强弱的程度, 绝对值越大表示相关性越高, 如果样品间相关系数的绝对值为0则表示无关, 在0.9以上为高度相关, 1表示变量完全正相关。1.7 生物信息学分析
用于生物信息分析的定量蛋白质假阳性率q-value值<0.01, 4个样品的总信号强度Intensity>0, 蛋白质信号强度的相对比值大于或等于2倍即视为差异蛋白质。从植物基因组水稻数据库中分别批量提取上调和下调差异蛋白质的FASTA格式序列导入Omicsbean软件(http://www.omicsbean.cn/)进行本体(Gene Ontology, GO)和代谢通路(Kyoto Encyclopedia of Genes and Genomes, KEGG)富集分析。1.8 PRM靶向定量
4个样品混合样的上样量为1 μL, DDA分析和定性搜库参数设置同上。将混合样中待验证蛋白的定性结果导入Skyline软件(https://skyline.ms/), 选择待验证目标蛋白肽段并编辑PRM检测方法, 使用QE-Plus分别采集4个样品中待验证蛋白的PRM定量数据, 离子隔离窗口(isolation window)为1.6 m z-1, 一级全扫描1次结合30次PRM扫描, 一级全扫描的分辨率为70 000, PRM扫描分辨率设为17 500, 用Skyline完成待验证蛋白的定量分析。2 结果与分析
2.1 水稻低温胁迫表型观察及叶绿素含量测定
EBR处理26°C常温培养的样品与单独26°C培养样品在表型上无差异, 低温培养的两组样品中, 单独4°C培养样品明显偏黄, 芽也相对细弱(图1-A)。常温培养样品的叶绿素a和叶绿素b的含量均没有显著差异, 但明显高于低温培养的样品; EBR处理4°C培养样品与单独4°C培养样品相比, 叶绿素a含量差异显著, 叶绿素b含量差异不显著(图1-B); 全株称重3次, 每次10粒发芽种子, 结果显示4个样品间的全株鲜重无显著差异(图1-C)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1水稻低温胁迫表型及叶绿素含量
A: 4组样品表型图; B: 4组样品叶绿素a和叶绿素b含量比较; C: 4组样品全株重比较。随机选取10株幼苗称重, 重复3次, 算其平均值作为该组样品的全株鲜重。4B: EBR处理4°C培养样品, 4: 单独4°C培养样品, 26B: EBR处理26°C培养样品, 26: 单独26°C培养样品。B和C中标以不同小字母的同组数值在0.05水平差异显著。
Fig. 1Phenotypes under cold stress and of chlorophyll content in rice
A: four groups of sample phenotype; B: comparison of chlorophyll a and chlorophyll b contents in four groups of samples; C: comparison of whole plant weight in four groups of samples, randomly selected 10 seedlings and weighed which was repeated three times and calculate the average value as the whole plant fresh weight of a group. 4B: samples treated with EBR then cultured at 4°C, 4: samples cultured at 4°C, 26B: samples treated with EBR then cultured at 26°C, 26: samples cultured at 26°C. Values within a group followed by a different small letter are significantly different at the 0.05 probability level in B and C.
2.2 质谱分析
从4个样品定性鉴定到5778个蛋白质, 平均鉴定到每个样品4331个蛋白质, 低温胁迫样品与正常培养样品(4 vs 26, 4B vs 26B)相比鉴定出的蛋白质数量较少, 但同温度培养的EBR处理样品蛋白鉴定数有所增加。2个样品间共有的蛋白质数在3508到3631之间, 其中正常培养的2个样品的共有蛋白质数量最多, 占2个样品总蛋白数的72.4%; 低温胁迫处理的2个样品间共有蛋白质数量最少, 仅占2个样品总蛋白数的69.3%; 4个样品共有蛋白数为3112, 占4个样品总蛋白数的53.9% (图2-A)。对质谱定量数据的Pearson相关分析表明, 样品4B与其他几组样品的相似度普遍较高, 样品4B和26的定量结果相似度最高, 样品4B和26B以及不加EBR处理的两组样品的相似度居中, 样品4和26B的相似度低于其他的组合(图2-B)。4组样品定量鉴定结果还显示, EBR处理后正常培养及低温胁迫的样品中检测到有定量信息的蛋白质均增加, 且蛋白质信号强度多为107~108 (图2-C)。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2质谱定性和定量分析结果
A: 定性鉴定蛋白数及样品间共有蛋白数; B: 样品间的皮尔逊相关性分析图; C: 定量鉴定蛋白质信号强度分布表。
Fig. 2Qualitative and quantitative analysis of samples by mass spectrometry
A: qualitatively identified proteins and common proteins between samples; B: Pearson correlation analysis between samples; C: signal intensity distribution of quantitative identified proteins.
2.3 定量蛋白差异特性分析
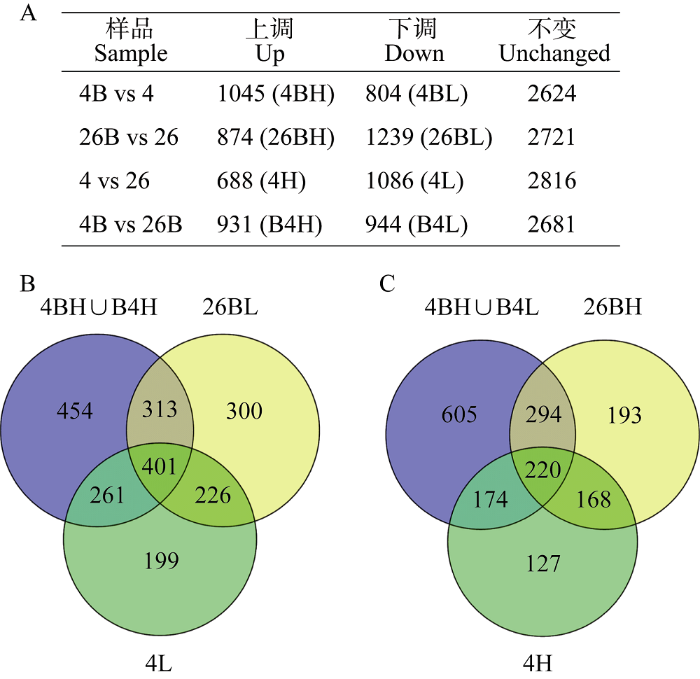
利用DDA质谱数据, 采用非标定量方法共鉴定到4834个有可靠定量信息的蛋白质, 其中样品4和26 (4 vs 26), 样品4B和26B (4B vs 26B), 样品4B和4 (4B vs 4), 以及样品26B和26 (26B vs 26)两两相比丰度差异大于2倍的上、下调蛋白数见图3-A。进一步分析后发现, 在“4B vs 26B”和“4B vs 4”上调蛋白的并集(4BH∪B4H)中, 即EBR处理4°C培养样品分别与EBR处理26°C培养及无EBR处理4°C培养样品相比上调的1429个蛋白中, 与“26B vs 26”和“4 vs 26”下调蛋白有交集的有401个, 记为与EBR影响水稻幼苗响应低温胁迫有关的上调蛋白(图3-B); “4B vs 26B”和“4B vs 4”下调蛋白的并集(4BL∪B4L)有1293个, 其中与“26B vs 26”和“4 vs 26”上调蛋白共有的有220个, 记为与EBR影响水稻幼苗响应低温胁迫有关的下调蛋白(图3-C)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3定量蛋白的差异性比较
A: 样品间的差异蛋白数; B: 与EBR影响水稻幼苗响应低温胁迫有关上调蛋白维恩图; C: 与EBR影响水稻幼苗响应低温胁迫有关下调蛋白维恩图。H表示上调, L表示下调。
Fig. 3Comparison of quantitative protein
A: proteins with intensity difference between every two samples; B: Venn diagram shows the 401 up-regulated proteins related to the effect of EBR on rice seedings response to cold stress; C: Venn diagram shows the 220 down-regulated proteins related to the effect of EBR on rice seeding response to cold stress. H means up regulated and L means down regulated.
2.4 GO分析
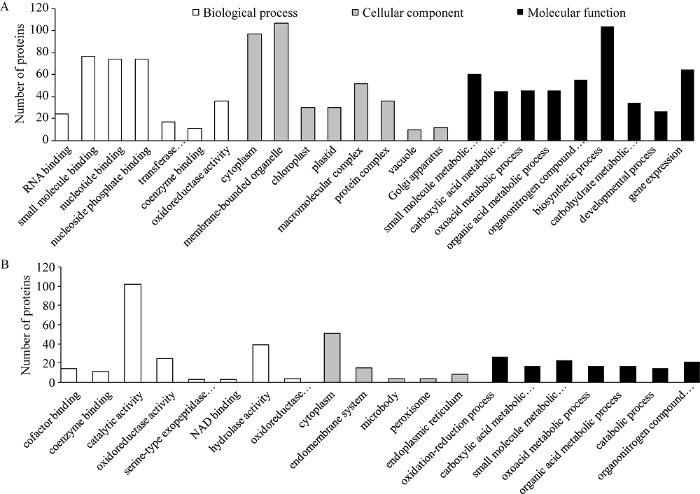
在与EBR影响水稻幼苗响应低温胁迫有关差异蛋白中, 上调蛋白质的分子功能主要涉及RNA结合、小分子结合、转移酶活性、氧化还原酶活性和辅酶结合等, 多数蛋白位于细胞质、叶绿体和有膜细胞器等, 参与了小分子代谢、有机氮化合物代谢、羧酸代谢、含氧酸代谢、有机酸代谢、碳水化合物生物合成、生物合成及基因表达等生物学过程(图4-A); 下调蛋白质其分子功能主要涉及辅因子结合、催化活性、辅酶结合、水解酶活性等, 多数位于细胞质、内质网和内膜系统, 生物过程富集分析显示这些蛋白主要参与了氧化还原、分解、含氧酸代谢、有机氮化合物代谢和小分子代谢等过程(图4-B)。图4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4影响水稻幼苗响应低温胁迫相关蛋白的本体分析
A: 401个上调蛋白质的本体分析; B: 220个下调蛋白质的本体分析。
Fig. 4Gene ontology analysis of proteins related to the effect of EBR on rice seeding response to cold stress
A: gene ontology analysis of 401 up-regulated proteins; B: gene ontology analysis of 220 down-regulated proteins.
2.5 代谢途径分析
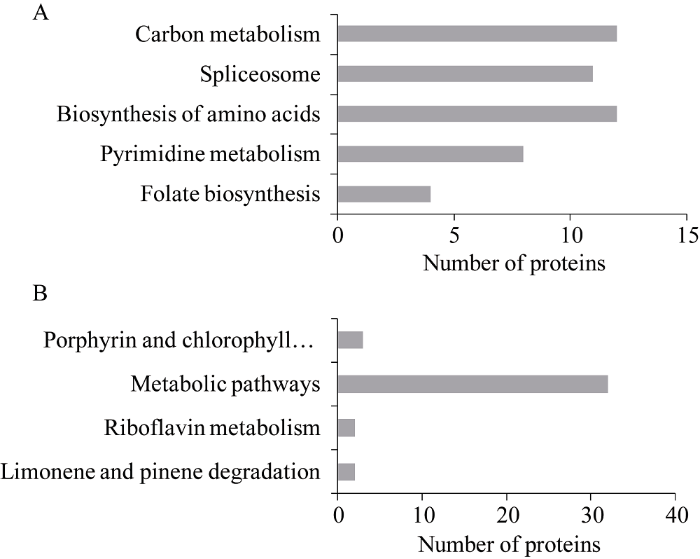
对差异蛋白进行代谢通路富集分析, 发现在P<0.05的范围内, 上调蛋白与叶酸生物合成、嘧啶代谢、氨基酸的生物合成、剪接体、碳代谢等代谢途径相关(图5-A)。下调蛋白与卟啉和叶绿素代谢、核黄素代谢、柠檬烯和蒎烯降解、其他代谢途径等相关(图5-B)。图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5代谢通路分析
A: 401个上调差异蛋白质代谢通路分析; B: 220个下调差异蛋白质的代谢通路分析。
Fig. 5Analysis of KEGG pathways
A: KEGG pathways analysis of 401 up-regulated proteins; B: KEGG pathways analysis of 220 down-regulated proteins.
在KEGG富集分析中, P值低于0.16且富集蛋白数多于4的重要代谢通路及其富集到的蛋白质种类见表1。
Table 1
表1
表1与EBR影响水稻幼苗响应低温胁迫相关的重要通路及富集的蛋白质
Table 1
| 通路和蛋白 Pathway name and protein IDs | 注解 Annotation | 信号强度Intensity | |||
|---|---|---|---|---|---|
| 26 | 26B | 4 | 4B | ||
| 碳代谢 Carbon metabolism | |||||
| LOC_Os01g54030.1 | Nadp-dependent malic enzyme | 0.00E+00 | 0.00E+00 | 0.00E+00 | 1.78E+07 |
| LOC_Os02g38200.1 | Dehydrogenase | 3.97E+08 | 1.78E+08 | 9.81E+07 | 2.64E+08 |
| LOC_Os03g15050.2 | Phosphoenolpyruvate carboxykinase | 1.22E+08 | 5.70E+07 | 4.21E+07 | 1.26E+08 |
| LOC_Os04g24140.1 | Ribose-5-phosphate isomerase a | 1.10E+08 | 8.97E+06 | 1.79E+07 | 7.07E+07 |
| LOC_Os06g04510.1 | Enolase | 5.68E+07 | 1.08E+07 | 0.00E+00 | 2.00E+07 |
| LOC_Os06g05700.1 | Cysteine synthase | 0.00E+00 | 0.00E+00 | 0.00E+00 | 8.73E+06 |
| LOC_Os06g45590.1 | Glyceraldehyde-3-phosphate dehydrogenase | 0.00E+00 | 0.00E+00 | 0.00E+00 | 3.28E+07 |
| LOC_Os07g09890.1 | Hexokinase | 2.82E+07 | 0.00E+00 | 0.00E+00 | 2.09E+07 |
| LOC_Os08g02700.1 | Fructose-bisphospate aldolase isozyme | 3.73E+07 | 0.00E+00 | 0.00E+00 | 8.83E+06 |
| LOC_Os09g24910.2 | Phosphofructokinase | 0.00E+00 | 0.00E+00 | 0.00E+00 | 2.38E+07 |
| LOC_Os11g10980.1 | Pyruvate kinase | 0.00E+00 | 0.00E+00 | 0.00E+00 | 1.06E+07 |
| LOC_Os11g41160.3 | Phosphoserine phosphatase | 2.13E+07 | 1.00E+07 | 0.00E+00 | 2.83E+07 |
| LOC_Os12g05110.1 | Pyruvate kinase | 2.37E+08 | 1.29E+08 | 5.51E+07 | 2.95E+08 |
| LOC_Os06g35540.1 | Aminotransferase | 5.93E+07 | 1.63E+08 | 1.61E+08 | 7.89E+07 |
| LOC_Os06g44460.1 | D-3-phosphoglycerate dehydrogenase | 9.76E+06 | 7.49E+07 | 2.85E+07 | 3.37E+07 |
| LOC_Os05g49760.1 | Dehydrogenase | 0.00E+00 | 1.20E+08 | 1.00E+08 | 3.93E+07 |
| 苯丙素生物合成 Phenylpropanoid biosynthesis | |||||
| LOC_Os10g17650.1 | Os10bglu34—beta-glucosidase homologue | 1.02E+08 | 0.00E+00 | 7.90E+07 | 4.76E+08 |
| LOC_Os01g32364.1 | Os1bglu1—beta-mannosidase/glucosidase homologue | 0.00E+00 | 0.00E+00 | 0.00E+00 | 8.51E+06 |
| LOC_Os01g73200.1 | Peroxidase | 0.00E+00 | 0.00E+00 | 0.00E+00 | 2.12E+07 |
| LOC_Os02g41680.1 | Phenylalanine ammonia-lyase | 7.94E+06 | 0.00E+00 | 0.00E+00 | 6.33E+06 |
| LOC_Os04g56180.1 | Peroxidase | 1.25E+08 | 2.86E+07 | 5.14E+07 | 7.05E+07 |
| LOC_Os03g11420.1 | Os3bglu6—beta-glucosidase/beta-fucosidase/beta-galactosidase | 0.00E+00 | 2.25E+07 | 3.03E+07 | 6.87E+06 |
| LOC_Os01g22249.1 | Peroxidase | 1.24E+07 | 5.91E+07 | 2.05E+08 | 0.00E+00 |
| LOC_Os05g04500.1 | Peroxidase | 8.07E+06 | 1.92E+08 | 5.61E+07 | 0.00E+00 |
| LOC_Os07g01410.1 | Peroxidase | 2.01E+07 | 5.70E+06 | 8.45E+07 | 0.00E+00 |
| LOC_Os08g34280.1 | Cinnamoyl-coa reductase | 0.00E+00 | 6.70E+06 | 6.25E+07 | 0.00E+00 |
| LOC_Os09g33680.1 | Os9bglu31—beta-glucosidase, dhurrinase | 0.00E+00 | 4.08E+07 | 1.92E+07 | 1.48E+07 |
| 卟啉和叶绿素代谢 Porphyrin and chlorophyll metabolism | |||||
| LOC_Os03g22780.1 | DVR | 8.72E+07 | 0.00E+00 | 4.27E+07 | 9.48E+07 |
| LOC_Os01g16520.1 | Glutamyl-tRNA synthetase | 0.00E+00 | 1.59E+07 | 1.40E+07 | 0.00E+00 |
| LOC_Os01g57460.1 | Frataxin, putative, expressed | 4.04E+07 | 9.91E+07 | 1.15E+08 | 0.00E+00 |
| LOC_Os10g37210.1 | FAD dependent oxidoreductase domain containing Protein | 2.01E+07 | 3.51E+06 | 5.45E+07 | 0.00E+00 |
| LOC_Os04g41260.1 | Amine oxidase | 0.00E+00 | 2.87E+06 | 1.70E+07 | 0.00E+00 |
| 叶酸生物合成 Folate biosynthesis | |||||
| LOC_Os11g29390.1 | Bifunctional dihydrofolate reductase-thymidylate synthase | 4.47E+06 | 1.31E+07 | 3.66E+06 | 0.00E+00 |
| LOC_Os09g38759.1 | Dihydroneopterin aldolase | 2.42E+07 | 0.00E+00 | 5.66E+06 | 2.25E+07 |
| LOC_Os04g38950.1 | Class I glutamine amidotransferase | 6.99E+07 | 0.00E+00 | 2.89E+07 | 2.37E+07 |
| LOC_Os03g02030.2 | Folylpolyglutamate synthase | 0.00E+00 | 0.00E+00 | 0.00E+00 | 2.04E+07 |
| LOC_Os02g35200.1 | Vp15 | 0.00E+00 | 0.00E+00 | 0.00E+00 | 7.22E+06 |
| 通路和蛋白 Pathway name and protein IDs | 注解 Annotation | 信号强度Intensity | |||
| 26 | 26B | 4 | 4B | ||
| 不饱和脂肪酸生物合成 Biosynthesis of unsaturated fatty acids | |||||
| LOC_Os01g65830.1 | Acyl-desaturase | 6.12E+07 | 0.00E+00 | 0.00E+00 | 8.06E+06 |
| LOC_Os02g48560.6 | Fatty acid desaturase | 4.69E+08 | 7.57E+07 | 8.14E+07 | 3.83E+08 |
| LOC_Os08g10010.1 | Acyl-desaturase | 1.38E+08 | 2.72E+07 | 4.92E+07 | 6.52E+07 |
| LOC_Os11g39220.2 | Acyl-coenzyme A oxidase | 0.00E+00 | 4.07E+07 | 3.18E+07 | 0.00E+00 |
| 脂肪酸生物合成 Fatty acid biosynthesis | |||||
| LOC_Os01g65830.1 | Acyl-desaturase | 6.12E+07 | 0.00E+00 | 0.00E+00 | 8.06E+06 |
| LOC_Os08g10010.1 | Acyl-desaturase | 1.38E+08 | 2.72E+07 | 4.92E+07 | 6.52E+07 |
| LOC_Os03g28420.1 | 3-oxoacyl-synthase | 8.02E+07 | 3.53E+07 | 0.00E+00 | 1.01E+08 |
| LOC_Os01g48910.2 | Long-chain acyl-coa synthetase | 3.27E+07 | 2.61E+07 | 4.38E+06 | 5.34E+07 |
| LOC_Os12g04990.3 | Long-chain acyl-coa synthetase | 1.80E+07 | 2.67E+07 | 6.15E+07 | 9.72E+06 |
新窗口打开|下载CSV
如表1所示, 富集在碳代谢途径中的NADP依赖性苹果酸酶、甘油醛-3-磷酸脱氢酶、磷酸果糖激酶和丙酮酸激酶(LOC_Os11g10980.1), 苯丙素的生物合成途径中的过氧化物酶(LOC_Os01g73200.1)等仅在样品4B中被检测到。碳代谢途径中的烯醇化酶、卟啉和叶绿素代谢途径中的联乙烯还原酶(divinyl reductase, DVR)在样品4B中的蛋白丰度是样品4和26B中的2倍以上。
2.6 PRM验证
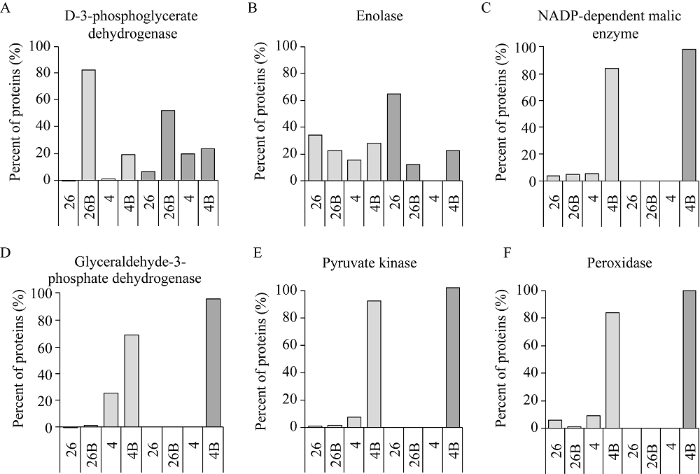
PRM技术可对样品中的目标肽段进行准确的特异性分析, 因此可用于验证非标记定量分析得到的差异蛋白。在对所有差异蛋白的生物学功能分析的基础上, 选取NADP-苹果酸酶(LOC_Os01g540 30.1)、过氧化物酶(LOC_Os01g73200.1)、3-磷酸甘油酸脱氢酶(LOC_Os06g44460.1)、烯醇化酶(LOC_Os06g04510.1)、甘油醛-3-磷酸脱氢酶(LOC_Os06g 45590.1)和丙酮酸激酶(LOC_Os11g10980.1)共6个蛋白进行PRM分析(图6)。表明EBR影响水稻幼苗响应低温胁迫相关的1个下调和5个上调蛋白在4个样品中的变化趋势与DDA数据(表1)基本一致。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6部分差异丰度蛋白PRM验证
A: 3-磷酸甘油酸脱氢酶; B: 烯醇化酶; C: NADP-苹果酸酶; D: 甘油醛-3-磷酸脱氢酶; E: 丙酮酸激酶; F: 过氧化物酶。
Fig. 6Targeted PRM quantification of some proteins with intensity difference
A: 3-phosphoglycerate dehydrogenase; B: enolase; C: NADP-dependent malic enzyme; D: glyceraldehyde-3-phosphate dehydrogenase; E: pyruvate kinase; F: peroxidase.
3 讨论
以往的研究证实, 低温胁迫可影响光合作用、细胞膜、活性氧累积等相关生物学过程[20], 而激素在植物对低温胁迫的响应中发挥着重要作用[21]。本研究利用蛋白质组学技术分析了EBR影响水稻幼苗响应低温胁迫的相关蛋白和代谢通路, 为进一步的相关机制研究提供了新的线索。3.1 表型和叶绿素含量分析
在本研究中, 未经EBR处理的低温胁迫样品明显比EBR处理的样品细弱, 且芽的颜色偏黄, 而在正常温度下生长的两组样品从表型上未看出明显的差异(图1-A), 提示EBR增强了低温胁迫水稻幼苗的光合作用。EBR处理后低温胁迫样品与未EBR处理的低温胁迫样品比, 叶绿素a含量显著增加(图1-B), 进一步提示EBR对水稻幼苗低温胁迫响应的影响与叶绿素a的合成或降解相关。低温胁迫后单株重显著降低, 但在相同培养条件下未见EBR对单株鲜重有明显影响, 可能与植物材料培养时间较短有关(图1-C)。3.2 质谱定性和定量数据分析
本文共鉴定到5778个水稻幼苗蛋白质, 说明所用的蛋白样品制备和质谱分析方法较适宜。定性鉴定结果(图2-A)显示, 两组EBR处理样品与未经EBR处理样品相比, 鉴定的蛋白质种类增加, 证明EBR处理可提高一些蛋白质的表达水平。由图2-C可知, 蛋白质的信号强度呈正态分布, 样品信号强度均较高, 说明鉴定结果比较稳定。Pearson相关分析(图2-B)显示, EBR处理后低温培养的样品(样品4B)与未经EBR处理正常培养的样品(样品26)相似度最高, 表明EBR可能使水稻幼苗在低温胁迫下趋向于恢复到正常的生理状态。定性鉴定显示低温培养的EBR处理和未经EBR处理样品的相似度最低(图2-A), 结果与Pearson分析相吻合。在EBR处理后低温胁迫条件下上调, 但EBR处理后正常培养下调和无EBR低温胁迫下调的401个蛋白质(图3-B), 以及220个在EBR处理后低温胁迫下调, 在EBR处理后正常培养和无EBR低温胁迫后上调的蛋白质(图3-C), 可能同时与EBR处理和温度胁迫相关, 是值得重点关注的丰度差异蛋白。3.3 功能和通路富集分析
功能和通路富集分析发现, 上调的蛋白主要与结合活性相关, 多位于细胞质和叶绿体 (图4-A), 涉及碳代谢、叶酸生物合成、氨基酸的生物合成等通路(图5-A), 下调的蛋白主要涉及到催化活性和氧化还原酶活性, 多位于细胞质、内膜系统和内质网(图4-B), 与卟啉和叶绿素代谢、核黄素代谢等相关(图5-B), 表明水稻幼苗经EBR处理后低温胁迫时的能量代谢和光合作用有所增强, 蛋白降解过程减弱。碳代谢途径中富集到16个与EBR影响水稻幼苗响应低温胁迫相关的蛋白(表1)。这16个蛋白质主要参与卡尔文循环、三羧酸循环和糖酵解, 其中参与卡尔文循环的果糖-双磷酸醛缩酶同功酶和核糖-5-磷酸异构酶A在未经EBR处理的低温胁迫样品(样品4)中, 与正常培养样品(样品26)相比丰度下调, 然而在EBR处理低温胁迫样品中上调; 参与糖酵解的烯醇化酶、甘油醛-3-磷酸脱氢酶、果糖-双磷酸醛缩酶同功酶和2个丙酮酸激酶均上调。有研究显示, 番茄植株的糖酵解过程在寒冷条件下显著增强, 有利于维持更快的叶呼吸和体内平衡[22], 苹果的糖酵解反应在4°C冷藏时也明显增强[23], 提示碳代谢途径中糖酵解等相关蛋白的变化与EBR影响水稻幼苗响应低温胁迫紧密相关。苯丙素生物合成途径富集到11个蛋白, 其中Os10bglu34、Os1bglu1、苯丙氨酸解氨酶和2个过氧化物酶(分别由LOC_ Os01g73200.1和LOC_Os04g56180.1编码)上调, Os3bglu6、肉桂酰辅酶A还原酶等6个蛋白下调。苯丙素有助于植物响应各种生物和非生物胁迫且与木质素、类黄酮等活性成分的生物合成相关, 苯丙氨酸解氨酶参与催化苯丙氨酸的非氧化脱氨反应并将来自于莽草酸途径的碳流引向苯丙素代谢的各个分支[24], 肉桂酰辅酶A还原酶参与水稻的防御反应[25], 说明EBR也可能通过调控苯丙素生物合成途径产生影响。富集到5个叶绿素代谢相关蛋白, 其中DVR是水稻等植物中合成叶绿素a的关键酶[26,27]。文献报道EBR能增加低温胁迫下苦瓜和玉米的叶绿素含量[9,10], 本研究也发现低温胁迫时EBR处理样品与无EBR处理样品相比, 叶绿素a含量明显增加(图1-B), 表明EBR可能通过DVR调节低温胁迫幼苗叶绿素a的合成。但另一方面, 叶绿素代谢通路中的谷氨酰tRNA合成酶、共济蛋白、FAD依赖的氧化还原酶结构域蛋白及胺氧化酶蛋白下调, 提示EBR对水稻幼苗响应低温胁迫的影响与叶绿素相关代谢途径存在复杂的关系。本研究也富集到一些叶酸合成相关蛋白。叶酸参与多种生物学过程, 不仅是C1转移反应的重要辅助因子[28,29], 也在光呼吸、叶绿素合成、抗氧化胁迫等方面起着重要作用[30,31,32], 该途径中的二氢蝶呤醛缩酶、I类谷氨酰胺转氨酶、VP15和叶酰聚谷氨酸合酶上调, 而二氢叶酸还原酶- 胸苷酸合酶下调, 提示叶酸生物合成途径与EBR影响水稻幼苗响应低温胁迫的关系还有待进一步研究。另外, 有研究表明β-酮酰基酰基载体蛋白合成酶(3-氧代酰基合酶)与脂肪酸温度依赖性组分的调控相关[33], 低温条件下水稻类囊体PSII膜脂质中不饱和脂肪酸呈现先降低后升高的趋势[34]。本研究发现, 不饱和脂肪酸代谢和脂肪酸代谢途径中的脂肪酸去饱和酶、酰基辅酶A氧化酶、3-氧代酰基合酶、酰脱氢酶和酰基辅酶A合成酶在EBR处理后低温胁迫条件下丰度有较大变化, 虽未见上述蛋白与EBR相关性的报道, 但可能在EBR影响水稻幼苗响应低温胁迫中发挥了重要的作用。
3.4 PRM验证蛋白功能分析
PRM验证显示NADP-苹果酸酶、过氧化物酶、3-磷酸甘油酸脱氢酶、烯醇化酶、甘油醛-3-磷酸脱氢酶和丙酮酸激酶在4组样品中的丰度变化趋势与非标记定量结果近似(图6), 进一步证实了蛋白定量研究结果的可靠性。3-磷酸甘油酸脱氢酶(PGDH)是参与丝氨酸生物合成的磷酸化途径的第一种酶, 在向非光合组织供应丝氨酸中起重要作用[35]。本研究发现经EBR处理后PGDH上调, 但未低温胁迫的样品中上调更明显(图6-A), 说明低温胁迫会抑制非光合组织中的丝氨酸供应, 但EBR可能影响了这种抑制作用。烯醇化酶与植物冷胁迫的关系已有大量报道。作为CBF调节因子的活化剂, 烯醇化酶不仅在碳代谢中起重要的作用, 而且在冷诱导基因表达的调节中也起作用[36,37]。在三叶橙(Poncirus trifoliata L.)中分离到一个编码烯醇酶的基因PtrLOS2, 低温处理后该基因在叶中表达上调[38]。茶树(Camellia sinensis L.)烯醇酶基因CsENO也在低温下诱导表达, 通过糖酵解途径产生的能量逐渐积累并修复光合系统[39]。还有研究显示在小麦叶片中的烯醇酶在低温下的表达先上调后下调[40]。我们的研究显示烯醇化酶在低温胁迫下下调, 经EBR处理后上调(图6-B), 表明该蛋白与EBR调控植物响应低温胁迫密切相关。NADP-苹果酸酶参与了植物细胞质体和细胞溶质的各种代谢途径, 催化(L)-苹果酸和NADP的氧化脱羧以产生丙酮酸和二氧化碳[41,42]。有研究表明, 外源激素ABA和SA以及多种非生物胁迫可调节小麦中TaNADP- ME1和TaNADP-ME2基因的表达[43]。NADP-苹果酸酶在EBR处理低温胁迫样品中明显上调(图6-C), 表明该蛋白与EBR影响水稻幼苗响应低温胁迫有密切关系。甘油醛-3-磷酸脱氢酶(GAPDH)在能量代谢中有重要作用, 是糖酵解的中枢酶, 克隆的小麦GAPDH基因经ABA和低温处理后可显著表达[44]。蛋白质组分析显示GAPDH在EBR处理后低温胁迫的水稻幼苗中明显上调, PRM验证发现低温胁迫时该蛋白也有一定程度上调(图6-D), 表明该蛋白在EBR信号通路和低温胁迫相互关系作用还需要进一步研究。丙酮酸激酶是糖酵解途径中的一种转移酶, 在EBR处理后低温胁迫样品中明显上调(图6-E)。有研究显示低温等条件下糖酵解反应增强[22,23], 但该蛋白在植物中的相关功能未见报道, 在EBR影响水稻幼苗响应低温胁迫中的具体作用有待研究。在苯丙素生物合成途径中过氧化物酶的增加会使木质素含量增加, 从而提高低温抗性。有研究证明过氧化物酶基因的过表达增加了油菜素类固醇敏感突变体bri1-9的耐寒性[45], 也有研究表明过氧化物酶2 (OsAPX2)在干旱、盐和冷胁迫下对水稻生长和繁殖起着关键作用[46]。过氧化物酶(LOC_Os01g73200.1)在加EBR低温胁迫中明显增加(图6-F), 表明该蛋白可能与EBR影响水稻幼苗响应低温胁迫有密切关系。
4 结论
利用LC-MS-MS从水稻幼苗中鉴定到5778个蛋白质, 其中621个差异丰度蛋白质与EBR影响水稻幼苗响应低温胁迫有关。差异蛋白主要集中在碳代谢、苯丙素代谢、叶绿素合成、叶酸合成以及脂肪酸代谢等通路中。分布在碳代谢和苯丙素代谢通路中的NADP-苹果酸酶、过氧化物酶、3-磷酸甘油酸脱氢酶、烯醇化酶、甘油醛-3-磷酸脱氢酶和丙酮酸激酶可能在EBR对低温胁迫水稻幼苗的调控中发挥重要作用。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/j.febslet.2009.01.020URLPMID:19167382 [本文引用: 1]

Abstract Twenty-two conserved miRNAs were chosen to investigate the expression pattern in response to phytohormone treatments, in which the effects of five classic plant hormone stresses were surveyed in Oryza sativa. The results showed that 11 miRNAs were found to be dysregulated by one or more phytohormone treatments. The target genes of these miRNAs were validated in vivo and their expression profiling were revealed. We also analyzed the promoter regions of the 22 conserved miRNAs for phytohormone-responsive elements and the existence of the elements provided further evidences supporting our results. These findings enable us to further investigate the role of miRNAs in phytohormone signaling.
DOI:10.1016/j.pbi.2011.02.001URL [本文引用: 1]
DOI:10.1016/j.molp.2016.06.010URLPMID:27375203 [本文引用: 1]
Brassinosteroids (BR ) 是为植物生长和开发是必要的类固醇荷尔蒙的一个班。在 dicot 模型植物 Arabidopsis 的 BR 信号 transduction 小径很好被建立,但是连接在米饭表明步的 BR 的部件一直不是充分探索了。例如,特别在 BR 受体水平,发信号的 BR 怎么在米饭被调整,大部分是未知的。这里,我们显示出那 OsPRA2,米饭小 G 蛋白质,在发信号的 BR 起一个压抑的作用小径。薄板倾斜, coleoptile 延伸,和根抑制试金显示有 OsPRA2 的压制的表示的米饭植物比野类型的植物对外长地应用的 brassinolide 更敏感。相反地,米饭 overexpressing OsPRA2 对外长的 brassinolide 不太敏感。进一步的学习揭开了那 OsPRA2 禁止了 dephosphorylation,并且因此使抄写因素失去活性 BRASSINAZOLE 抵抗 1 (OsBZR1 ) 。更重要地, OsPRA2 被发现到共同本地化与并且直接绑在米饭 BR 受体 BRASSINOSTEROID 感觉迟钝 1 (OsBRI1 ) 在血浆膜。另外,在里面 vitro 试金证明 OsPRA2 禁止它的 autophosphorylation。这个 OsPRA2-OsBRI1 相互作用从它的合作受体 OsBAK1 导致了 OsBRI1 的分离,并且废除了 OsBAK1 的调停 OsBRI1 的 phosphorylation。一起,这些结果在 OsBRI1 和 OsBZR1 上作为一个新奇否定管理者揭示 OsPRA2 的可能的工作机制并且关于米饭 BR 发信号的规章的机制扩大知识。
DOI:10.1093/mp/ssu050URL [本文引用: 1]
DOI:10.1007/s00299-014-1578-7URL [本文引用: 2]
DOI:10.1007/978-1-4939-6813-8URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.2095-1191.2011.05.007URL [本文引用: 2]
[目的]筛选出油菜素内酯(BR)提高苦瓜抗冷性的最佳浓度,为 解决苦瓜低温胁迫问题提供理论依据.[方法]设CK1和CK2为清水对照(CK1为常温处理,CK2为低温胁迫处理),通过对不同浓度(10-4、 10-3、10-2、10-1、10 mg/L)BR处理后的苦瓜幼苗进行(8.0±0.5)℃ 72 h低温胁迫,并测定植株生长指标及功能叶片中的丙二醛(MDA)含量、可溶性蛋白含量、细胞质相对电导率及超氧化物岐化酶(SOD)、过氧化物酶 (POD)活性等.[结果]与CK2相比,不同浓度BR处理可以促进苦瓜幼苗在低温下生长,其鲜物重和叶片叶绿素含量增加,冷害指数降低,POD、SOD 活性增加,可溶性蛋白和Pro含量升高,并在一定程度上减缓叶片中MDA的积累,同时可以明显降低细胞质相对电导率.[结论]喷施不同浓度BR可有效提高 苦瓜幼苗的抗冷性,其中以10-2 mg/L浓度处理最佳.
DOI:10.3969/j.issn.2095-1191.2011.05.007URL [本文引用: 2]
[目的]筛选出油菜素内酯(BR)提高苦瓜抗冷性的最佳浓度,为 解决苦瓜低温胁迫问题提供理论依据.[方法]设CK1和CK2为清水对照(CK1为常温处理,CK2为低温胁迫处理),通过对不同浓度(10-4、 10-3、10-2、10-1、10 mg/L)BR处理后的苦瓜幼苗进行(8.0±0.5)℃ 72 h低温胁迫,并测定植株生长指标及功能叶片中的丙二醛(MDA)含量、可溶性蛋白含量、细胞质相对电导率及超氧化物岐化酶(SOD)、过氧化物酶 (POD)活性等.[结果]与CK2相比,不同浓度BR处理可以促进苦瓜幼苗在低温下生长,其鲜物重和叶片叶绿素含量增加,冷害指数降低,POD、SOD 活性增加,可溶性蛋白和Pro含量升高,并在一定程度上减缓叶片中MDA的积累,同时可以明显降低细胞质相对电导率.[结论]喷施不同浓度BR可有效提高 苦瓜幼苗的抗冷性,其中以10-2 mg/L浓度处理最佳.
DOI:10.1007/s12298-012-0122-xURLPMID:3550514 [本文引用: 2]
Abstract Germination and early seedling growth are important for establishment of maize because maize is chilling sensitive crop and low temperature during early period of growth can be detrimental to subsequent crop growth and productivity. Therefore, it is important to protect maize seedling from cold stress. A study was conducted on induced cold tolerance by 24-epibrassinoslide (EBR) at the Indian Agricultural Research Institute, New Delhi, India. Maize seedlings were raised in green house condition (25/18 °C02day-night temperatures). Ten days old seedlings were treated with EBR (0.0, 0.01, 0.1, 1.0 and 1002μM) and then divided into two sets, one set was kept in greenhouse (25/18 °C02day-night temperatures) and another was transferred to net house (cold stress). Data on various morpho-physiological traits was recorded after 7, 14 and 2102days of treatment. Exogenous application of 1.002μM EBR had significant effect on growth and morpho-physiological traits under both conditions. The maize seedlings treated with EBR were more tolerant to cold stress than the untreated one. Significant increase in plant height, dry matter accumulation, chlorophyll content, total soluble proteins and starch contents was observed under both conditions, however, the results were more pronounced under cold stress. 1.002μ65M concentration being the most effective under both conditions. Maintenance of high tissue water content, reduced membrane injury index, increased total chlorophyll, soluble sugar and protein content were taken as the possible indicators of EBR induced chilling tolerance.
DOI:10.1016/j.plaphy.2016.06.021URLPMID:27362298 [本文引用: 1]
61Low temperature and weak light stress cause the inhibition of photochemical efficiency and nitrogen metabolism in tomato seedlings.61Chlorophyll fluorescence imaging technique was used to analyze the photochemical efficiency of photosystem II.61EBR could alleviate the photoinhibition and improve the photochemical efficiency of stressed-tomato seedlings.61EBR could enhance the assimilation of nitrate and ammonium in the primary nitrogen metabolism pathway.61EBR could confer resistance to tomato plants against low temperature and weak light stress.
URL [本文引用: 2]
近年来,随着人类生产活动和温室效应的作用,全球极端天气频发。冬春季节,我国北方设施蔬菜生产面临冷害和冻害的严重威胁。因此,在改善农业基础设施和提高设施环境调控能力的同时,研究蔬菜作物对逆境胁迫的应答机制,运用生长调节剂等提高蔬菜作物对不良环境条件的耐性,对提高蔬菜产量、品质和经济效益具有十分重要的科学意义。油菜素内酯(BRs)作为一种新型植物生长调节剂,广泛参与植物多种生理过程,尤其在植物生长发育及其对逆境的响应等方面具有重要的调节作用。然而,目前人们对BRs介导的耐低温机制仍不清楚。本试验以低温敏感型辣椒品种‘湘研16号’为试材,通过叶面喷施24-表油菜素内酯(24-epibrassinolide,EBR)研究了外源EBR对低温胁迫下(15/5℃,昼/夜)辣椒幼苗生长、激发能分配、渗透调节物质积累和抗氧化防御的影响;利用高通量测序RNA-seq和i TRAQ技术,分析了低温胁迫下EBR对辣椒幼苗转录组和蛋白质组的影响,并利用实时荧光定量PCR技术验证了高通量测序结果的可靠性。从生理生化、基因和蛋白质3个方面探讨BRs调控辣椒幼苗耐低温胁迫的生理和分子调控机制。主要试验结果如下:(1)低温胁迫下,辣椒幼苗生长受到抑制,根系形态指标显著降低,膜脂过氧化产物MDA含量显著提高,抗氧化酶活性(SOD、POD、CAT)显著增强。低温胁迫下,外源EBR处理可显著提高辣椒幼苗地上部干鲜重、根系活力、根系总长、根表面积和分根数及根系抗氧化酶SOD、POD和CAT活性,降低MDA含量,缓解低温对根系的伤害。EBR的适宜浓度为0.1μM。(2)EBR亦可显著提高低温胁迫下辣椒叶片光光系统II最大量子效率、光系统II实际光化学效率和光合系统II反应中心光能捕获效率,减少天线热耗散和过剩光能,缓解光抑制。低温胁迫增加了叶片MDA和活性氧H2O2、O2-和OH-的含量,产生了氧化应激反应,而通过喷施EBR可显著提高叶片SOD、POD、CAT、APX抗氧化酶活性(酶系统)和抗氧化剂As A和GSH的含量(非酶系统),降低ROS积累,缓解过氧化伤害。此外,低温下喷施EBR可以显著提高渗透调节物质脯氨酸、可溶性糖、可溶性蛋白质的含量。(3)采用RNA-Seq高通量测序技术共获得39,829个转录本,经过差异基因筛选,共得到656条差异表达基因,包括335条上调和321条下调差异表达基因。利用RT-PCR方法对20个差异表达基因进行了表达量验证,定量检测结果与测序结果一致,证明建立的转录组数据库可靠。通过GO功能注释和KEGG富集分析,EBR可诱导光合作用相关基因显著上调,诱导纤维素合成蛋白和UDP糖基转移酶的合成,引起细胞质中钙信号的转导途径,诱导细胞内氧化还原相关基因GSTX1,PER72、CAT2的上调表达,也诱导激素代谢、氧化还原过程、信号转导和防御反应相关基因的表达。(4)采用i TRAQ技术共鉴定得到4661个蛋白质。通过差异蛋白质筛选,筛出上调蛋白217个,下调蛋白129个。GO注释分析表明,细胞学组件(Cellular Component)中,鉴定的差异蛋白主要是细胞功能和细胞组成相关蛋白;生物学过程(Biological Process)中,主要行使代谢过程(metabolic process,20.83%)、细胞过程(cellular processes,18.28%)和应激反应(response to stimulus,9.85%);主要参与催化活性(catalytic activity,47.73%)、结合功能(binding,36.62%)、结构性分子活性(structural molecular activity,5.30%)和抗氧化活性(antioxidant activity,3.54%)等生物过程。Pathway富集分析表明,差异蛋白被显著富集在82个代谢通路中,主要是代谢途径、次生代谢生物合成、核蛋白、光合作用天线蛋白等。主要参与电子传递以感应和传导逆境信号,调节光合作用和能量代谢以维持细胞内正常的能量供给,降低细胞壁的延展性以维系低温胁迫下细胞完整性,生成抗逆活性物质以维持细胞内环境稳定。主要参与光合固碳途径、氧化磷酸化途径、谷胱甘肽代谢途径、苯丙素生物合成途径、苯丙氨酸代谢途径和核糖体途径等代谢途径及催化及活性调节、蛋白质结构性分子活性、能量转运过程和蛋白质结合等过程。
URL [本文引用: 2]
近年来,随着人类生产活动和温室效应的作用,全球极端天气频发。冬春季节,我国北方设施蔬菜生产面临冷害和冻害的严重威胁。因此,在改善农业基础设施和提高设施环境调控能力的同时,研究蔬菜作物对逆境胁迫的应答机制,运用生长调节剂等提高蔬菜作物对不良环境条件的耐性,对提高蔬菜产量、品质和经济效益具有十分重要的科学意义。油菜素内酯(BRs)作为一种新型植物生长调节剂,广泛参与植物多种生理过程,尤其在植物生长发育及其对逆境的响应等方面具有重要的调节作用。然而,目前人们对BRs介导的耐低温机制仍不清楚。本试验以低温敏感型辣椒品种‘湘研16号’为试材,通过叶面喷施24-表油菜素内酯(24-epibrassinolide,EBR)研究了外源EBR对低温胁迫下(15/5℃,昼/夜)辣椒幼苗生长、激发能分配、渗透调节物质积累和抗氧化防御的影响;利用高通量测序RNA-seq和i TRAQ技术,分析了低温胁迫下EBR对辣椒幼苗转录组和蛋白质组的影响,并利用实时荧光定量PCR技术验证了高通量测序结果的可靠性。从生理生化、基因和蛋白质3个方面探讨BRs调控辣椒幼苗耐低温胁迫的生理和分子调控机制。主要试验结果如下:(1)低温胁迫下,辣椒幼苗生长受到抑制,根系形态指标显著降低,膜脂过氧化产物MDA含量显著提高,抗氧化酶活性(SOD、POD、CAT)显著增强。低温胁迫下,外源EBR处理可显著提高辣椒幼苗地上部干鲜重、根系活力、根系总长、根表面积和分根数及根系抗氧化酶SOD、POD和CAT活性,降低MDA含量,缓解低温对根系的伤害。EBR的适宜浓度为0.1μM。(2)EBR亦可显著提高低温胁迫下辣椒叶片光光系统II最大量子效率、光系统II实际光化学效率和光合系统II反应中心光能捕获效率,减少天线热耗散和过剩光能,缓解光抑制。低温胁迫增加了叶片MDA和活性氧H2O2、O2-和OH-的含量,产生了氧化应激反应,而通过喷施EBR可显著提高叶片SOD、POD、CAT、APX抗氧化酶活性(酶系统)和抗氧化剂As A和GSH的含量(非酶系统),降低ROS积累,缓解过氧化伤害。此外,低温下喷施EBR可以显著提高渗透调节物质脯氨酸、可溶性糖、可溶性蛋白质的含量。(3)采用RNA-Seq高通量测序技术共获得39,829个转录本,经过差异基因筛选,共得到656条差异表达基因,包括335条上调和321条下调差异表达基因。利用RT-PCR方法对20个差异表达基因进行了表达量验证,定量检测结果与测序结果一致,证明建立的转录组数据库可靠。通过GO功能注释和KEGG富集分析,EBR可诱导光合作用相关基因显著上调,诱导纤维素合成蛋白和UDP糖基转移酶的合成,引起细胞质中钙信号的转导途径,诱导细胞内氧化还原相关基因GSTX1,PER72、CAT2的上调表达,也诱导激素代谢、氧化还原过程、信号转导和防御反应相关基因的表达。(4)采用i TRAQ技术共鉴定得到4661个蛋白质。通过差异蛋白质筛选,筛出上调蛋白217个,下调蛋白129个。GO注释分析表明,细胞学组件(Cellular Component)中,鉴定的差异蛋白主要是细胞功能和细胞组成相关蛋白;生物学过程(Biological Process)中,主要行使代谢过程(metabolic process,20.83%)、细胞过程(cellular processes,18.28%)和应激反应(response to stimulus,9.85%);主要参与催化活性(catalytic activity,47.73%)、结合功能(binding,36.62%)、结构性分子活性(structural molecular activity,5.30%)和抗氧化活性(antioxidant activity,3.54%)等生物过程。Pathway富集分析表明,差异蛋白被显著富集在82个代谢通路中,主要是代谢途径、次生代谢生物合成、核蛋白、光合作用天线蛋白等。主要参与电子传递以感应和传导逆境信号,调节光合作用和能量代谢以维持细胞内正常的能量供给,降低细胞壁的延展性以维系低温胁迫下细胞完整性,生成抗逆活性物质以维持细胞内环境稳定。主要参与光合固碳途径、氧化磷酸化途径、谷胱甘肽代谢途径、苯丙素生物合成途径、苯丙氨酸代谢途径和核糖体途径等代谢途径及催化及活性调节、蛋白质结构性分子活性、能量转运过程和蛋白质结合等过程。
[本文引用: 1]
DOI:10.1007/s10930-017-9721-2URL [本文引用: 1]
DOI:10.1111/jipb.2006.48.issue-8URL [本文引用: 1]
[本文引用: 1]
DOI:10.1002/(ISSN)1615-9861URL [本文引用: 1]
DOI:10.1002/(ISSN)1615-9861URL [本文引用: 1]
DOI:10.1002/pmic.201000727URL [本文引用: 1]
DOI:10.1016/S0065-2296(08)00602-2URL [本文引用: 1]
DOI:10.1007/s00018-015-2089-6URL [本文引用: 1]
DOI:10.1007/s00425-012-1799-3URLPMID:23229059 [本文引用: 2]
Abstract To find if cytosolic glycolysis dynamical metabolism plays a role in mediating respiration homeostasis and its relationship with mitochondrial electron transport chain (miETC) flexibility, we selected two tomato genotypes that differ in chilling tolerance and compared the responses of miETC, cytosolic glycolysis and respiratory homeostasis at 7000200°C. Our results showed that the transcripts of both classical and bypass component genes for miETC and glycolysis were comparable for both genotypes when grown at 25000200°C. However, there was a rapid global increase in the expression of most respiratory genes in response to chilling at 7000200°C for both genotypes. When normally grown plant was set as the control for each genotype, the transcripts of most COX family members, ATP synthase, AOX1b, and UCP are highly up-regulated in chilling-tolerant Zhefen No. 208 plants in contrast to the sensitive Zhefen No. 212 plants. Both genotypes mobilized the energy-saving sucrose synthase pathway for sucrose degradation by cytosolic glycolysis, but this mechanism is evidently more effective in tolerant Zhefen No. 208 plants. Furthermore, only Zhefen No. 208 plants were able to partially switch from low-energy efficiency pathways to ATP conserving pathways to carry out fructose-6-phosphate conversion and pyruvate production. This metabolic flexibility in miETC and cytosolic glycolysis were coupled to higher ATP synthesis and lower ROS accumulation, which may be essential for sustaining the higher leaf respiration and homeostasis of chilling-tolerant plants.
DOI:10.1034/j.1399-3054.1999.100103.xURL [本文引用: 2]
Abstract This study provides the first report on the occurrence of the respiratory climacteric during cold storage of apple fruit ( Malus domestica Borkh. cv. Reinette du Canada). The respiratory pattern at 4 C was very similar to that observed during postharvest ripening at room temperature, except that shelf life was considerably extended and the onset of the climacteric delayed. Increasing the calcium content of the apple fruit significantly reduced loss of firmness during cold storage, but showed no effect on respiration or on the other parameters determined. A gradual accumulation of soluble sugars occurred during the first 60 days after harvest and was effectively completed before the climacteric peak was reached. This increase in sugars correlated with an increase in the activity of sucrose-phosphate synthase (EC 2.4.1.14), and a marked change in the kinetic properties of the enzyme was observed after sucrose accumulation ceased. Changes in the hexose-phosphate pool and in glycolytic and gluconeogenic activities indicated an initial increase in the gluconeogenic flow at early stages of the climacteric, followed by activation of glycolysis, with the carbon flow being most likely regulated at the reversible phosphorylation of fructose-6-phosphate to fructose-1,6-bisphosphate (mostly via pyrophosphate:fructose-6-phosphate phosphotransferase, EC 2.7.1.90) and at the pyruvate kinase (EC 2.7.1.40) steps. The results presented indicate that the respiratory climacteric does not occur to accommodate extra ATP requirements during sucrose synthesis nor can it be a consequence of an increased supply of respiratory substrate.
DOI:10.1093/mp/ssp106URL [本文引用: 1]
DOI:10.1073/pnas.0509875103URLPMID:16380417 [本文引用: 1]
OsRac1, one of the Rac/Rop family of small GTPases, plays important roles in defense responses, including a role in the production of reactive oxygen species mediated by NADPH oxidase. We have identified an effector of OsRac1, namely rice (Oryza sativa) cinnamoyl-CoA reductase 1 (OsCCR1), an enzyme involved in lignin biosynthesis. Lignin, which is polymerized through peroxidase activity by using H(2)O(2) in the cell wall, is an important factor in plant defense responses, because it presents an undegradable mechanical barrier to most pathogens. Expression of OsCCR1 was induced by a sphingolipid elicitor, suggesting that OsCCR1 participates in defense signaling. In in vitro interaction and two-hybrid experiments, OsRac1 was shown to bind OsCCR1 in a GTP-dependent manner. Moreover, the interaction of OsCCR1 with OsRac1 led to the enzymatic activation of OsCCR1 in vitro. Transgenic cell cultures expressing the constitutively active OsRac1 accumulated lignin through enhanced CCR activity and increased reactive oxygen species production. Thus, it is likely that OsRac1 controls lignin synthesis through regulation of both NADPH oxidase and OsCCR1 activities during defense responses in rice.
[本文引用: 1]
[本文引用: 1]
DOI:10.1104/pp.110.158477URLPMID:20484022 [本文引用: 1]
3,8-Divinyl (proto)chlorophyll(ide) a 8-vinyl reductase (DVR) catalyzes the reduction of 8-vinyl group on the tetrapyrrole to an ethyl group, which is indispensable for monovinyl chlorophyll (Chl) synthesis. So far, three 8-vinyl reductase genes (DVR, bciA, and slr1923) have been characterized from Arabidopsis (Arabidopsis thaliana), Chlorobium tepidum, and Synechocystis sp. PCC6803. However, no 8-vinyl reductase gene has yet been identified in monocotyledonous plants. In this study, we isolated a spontaneous mutant, 824ys, in rice (Oryza sativa). The mutant exhibited a yellow-green leaf phenotype, reduced Chl level, arrested chloroplast development, and retarded growth rate. The phenotype of the 824ys mutant was caused by a recessive mutation in a nuclear gene on the short arm of rice chromosome 3. Map-based cloning of this mutant resulted in the identification of a gene (Os03g22780) showing sequence similarity with the Arabidopsis DVR gene (AT5G18660). In the 824ys mutant, nine nucleotides were deleted at residues 952 to 960 in the open reading frame, resulting in a deletion of three amino acid residues in the encoded product. High-performance liquid chromatography analysis of Chls indicated the mutant accumulates only divinyl Chl a and b. A recombinant protein encoded by Os03g22780 was expressed in Escherichia coli and found to catalyze the conversion of divinyl chlorophyll(ide) a to monovinyl chlorophyll(ide) a. Therefore, it has been confirmed that Os03g22780, renamed as OsDVR, encodes a functional DVR in rice. Based upon these results, we succeeded to identify an 8-vinyl reductase gene in monocotyledonous plants and, more importantly, confirmed the DVR activity to convert divinyl Chl a to monovinyl Chl a.
[本文引用: 1]
DOI:10.3724/SP.J.1259.2012.00525URL [本文引用: 1]
DOI:10.1002/jsfa.v81:9URL [本文引用: 1]
DOI:10.1111/j.1469-8137.2009.02790.xURL [本文引用: 1]
DOI:10.1038/nbt1351URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1744-7909.2007.00438.xURL [本文引用: 1]
Temperature is one of the abiotic factors limiting growth and productivity of plants. In the present work, the effect of low non-freezing temperature, as an inducer of "chilling resistance", was studied in three cultivars of rice ( Oryza sativa L.), japonica cv. 9516 (j-9516), the two parental lines of superhigh-yield hybrid rice between subspecies, Peiai/E32 (ji-PE), and the traditional indica hybrid rice Shanyou 63 (i-SY63). Leaves of chill-treated rice showed chilling-induced resistance, as an increase of their low-temperature tolerance was measured using chlorophyll fluorescence measurements, revealing a change in photosystem II (PSII) efficiency. After 5 d of exposure to 11 C under low light (100 mol m -2 s -1 ), levels of unsaturated fatty acids in PSII thylakoid membrane lipids decreased during the initial 1-2 d, then increased slowly and reached 99.2%, 95.3% and 90.1% of the initial value (0 d) in j-9516, ji-PE and i-SY63, respectively, on the fifth day. However, under medium light (600 mol m -2 s -1 ), all cultivars experienced similar substantial photoinhibition, which approached steady state levels after a decline in levels of unsaturated fatty acids in PSII thylakoid membrane lipids to about 57.1%, 53.8% and 44.5% of the initial values (0 d) in j-9516, ji-PE and I-SY63 on the fifth day. Under either chilling-induced resistance (the former) or low temperature photoinhibition (the latter) conditions, the changes of other physiological parameters such as D1 protein contents, electron transport activities of PSII (ETA), F v / F m , xanthophyl cycle activities expressed by DES (deepoxide state) were consistent with that of levels of unsaturated fatty acids in PSII thylakoid membrane lipids. So there were negative correlations between saturated levels of fatty acids (16:1(3t), 16:0, 18:0), especially the 16:1(3t) fatty acid on thylakoid membrane and other physiological parameters, such as D1 protein contents, ETA and (A+Z)/(A+V+Z). A specific role of desaturation of fatty acids and the photoprotective pigments of the xanthophyl cycle, leading to an acclimation response in thylakoid membrane lipids may be involved. We conclude that chilling-induced resistance is accelerated by the unsaturation of thylakoid membranes, and the ability of rice plants to cold-harden can be enhanced by genetic engineering.
[本文引用: 1]
DOI:10.1016/j.jprot.2011.02.006URLPMID:21329772 [本文引用: 1]
Abstract Plant acclimation to stress is associated with profound changes in proteome composition. Since proteins are directly involved in plant stress response, proteomics studies can significantly contribute to unravel the possible relationships between protein abundance and plant stress acclimation. In this review, proteomics studies dealing with plant response to a broad range of abiotic stress factors--cold, heat, drought, waterlogging, salinity, ozone treatment, hypoxia and anoxia, herbicide treatments, inadequate or excessive light conditions, disbalances in mineral nutrition, enhanced concentrations of heavy metals, radioactivity and mechanical wounding are discussed. Most studies have been carried out on model plants Arabidopsis thaliana and rice due to large protein sequence databases available; however, the variety of plant species used for proteomics analyses is rapidly increasing. Protein response pathways shared by different plant species under various stress conditions (glycolytic pathway, enzymes of ascorbate-glutathione cycle, accumulation of LEA proteins) as well as pathways unique to a given stress are discussed. Results from proteomics studies are interpreted with respect to physiological factors determining plant stress response. In conclusion, examples of application of proteomics studies in search for protein markers underlying phenotypic variation in physiological parameters associated with plant stress tolerance are given. Copyright 漏 2011 Elsevier B.V. All rights reserved.
DOI:10.1093/emboj/21.11.2692URL [本文引用: 1]
The Arabidopsis mutation, los2, impairs cold-responsive gene transcription, acquired freezing tolerance and plant resistance to chilling under certain conditions. LOS2 was isolated through positional cloning and shown to encode an enolase in the glycolytic pathway. In animal cells, enolase has also been known to function as a transcription factor that represses the expression of c-myc by binding to the c-myc gene promoter. LOS2 fused to green fluorescent protein is targeted to the nucleus as well as to the cytoplasm. LOS2/enolase protein can bind to the cis-element of the human c-myc gene promoter and to the gene promoter of STZ/ZAT10, a zinc finger transcriptional repressor from Arabidopsis. STZ/ZAT10 expression is induced rapidly and transiently by cold in the wild type, and this induction is stronger and more sustained in the los2 mutant. Furthermore, the expression of a RD29A-LUC reporter gene is repressed significantly by STZ/ZAT10 in transient expression assays in Arabidopsis leaves. Our results demonstrate that cold-responsive gene transcription in plants is controlled by a bi-functional enolase.
DOI:10.1007/s10535-011-0005-yURL [本文引用: 1]
Low expression of osmotically responsive genes 2 ( LOS2 ) encodes an enolase (2-phospho-D-glycerate hydrolase, EC 4.2.1.11) that converts 2-phospho-D-glycerate (PGA) to phosphoenolpyruvate (PEP) in the glycolytic pathway in Arabidopsis . Meanwhile, it is a transcriptional activator of cold-responsive gene, negatively controlling the expression of STZ/ZAT10 , a zinc finger transcriptional repressor of cold-responsive gene from Arabidopsis . A novel LOS2 gene, designated PtrLOS2 (GenBank accession number GQ144341), was isolated from trifoliate orange [ Poncirus trifoliata (L.) Raf.]. The PtrLOS2 cDNA is 1 662 bp in length with a 1 338 bp open reading frame (ORF), encoding a deduced 445 amino acid residue protein with a predicted molecular mass of 47.79 kDa and an isoelectric point of 5.54. The deduced protein of the PtrLOS2 gene shares high identity (over 86 %) with other plant species enolase, which suggests that the PtrLOS2 probably encodes an enolase. Sequence alignment showed that PtrLOS2 protein has a conserved DNA-binding and a repression domain. Moreover, a conserved start site of alternative translation for the c-myc promoter binding protein (MBP-1) was also found in PtrLOS2 protein. PtrLOS2 was constitutively expressed in leaves, stems and roots. PtrLOS2 expression in roots and stems was much higher than that in leaves under normal conditions, however, the expression of PtrLOS2 was up-regulated in leaves, but down-regulated in roots after cold treatments. The PtrLOS2 expression in stems was firstly up-regulated and then down-regulated after cold treatments. Meanwhile, after ABA treatment, the expression of PtrLOS2 was up-regulated in leaves but in stems and roots firstly down-regulated followed with up-regulation.
DOI:10.16420/j.issn.0513-353x.2016-0731URL [本文引用: 1]
从茶树‘黄旦’和‘金观音’抑制差减文库中筛选出1条烯醇酶(Enolase,ENO)cDNA片段序列,以‘铁观音’芽叶为材料克隆并验证该序列的全长编码c DNA序列。该序列全长1 753 bp,含有1个1 332 bp完整的开放阅读框,命名为CsENO,登录号KX962311。序列基因编码444个氨基酸,氨基酸序列分析发现,该c DNA与其他植物ENO高度保守,包含ENO特异结构域,与苹果(Malus×domestica)、白梨(Pyrus bretschneideri)的亲缘关系较近,相似性达84%。生物信息学预测结果显示,CsENO属于稳定的亲水蛋白,不存在跨膜结构,无信号肽,可能定位在过氧化体、溶酶体或核糖体等细胞质中,且具有多个磷酸化位点;二级结构主要由α–螺旋构成。实时荧光定量PCR结果表明,CsENO在低温、ABA、高盐、干旱逆境胁迫处理下均有表达,且表达受到不同程度的诱导。
DOI:10.16420/j.issn.0513-353x.2016-0731URL [本文引用: 1]
从茶树‘黄旦’和‘金观音’抑制差减文库中筛选出1条烯醇酶(Enolase,ENO)cDNA片段序列,以‘铁观音’芽叶为材料克隆并验证该序列的全长编码c DNA序列。该序列全长1 753 bp,含有1个1 332 bp完整的开放阅读框,命名为CsENO,登录号KX962311。序列基因编码444个氨基酸,氨基酸序列分析发现,该c DNA与其他植物ENO高度保守,包含ENO特异结构域,与苹果(Malus×domestica)、白梨(Pyrus bretschneideri)的亲缘关系较近,相似性达84%。生物信息学预测结果显示,CsENO属于稳定的亲水蛋白,不存在跨膜结构,无信号肽,可能定位在过氧化体、溶酶体或核糖体等细胞质中,且具有多个磷酸化位点;二级结构主要由α–螺旋构成。实时荧光定量PCR结果表明,CsENO在低温、ABA、高盐、干旱逆境胁迫处理下均有表达,且表达受到不同程度的诱导。
DOI:10.1111/aab.12019URL [本文引用: 1]
Seasonal low temperature (LT) adversely affects growth of plants. The onset of LT in temperate zones also entails the process of cold acclimation, preparing the plants to withstand freezing temperatures. During this process of cold acclimation a number of physiological, biochemical and molecular changes occur. A differentially expressed enolase gene in wheat plants exposed to LT was previously identified by cDNA-amplified fragment length polymorphism. In this study, two wheat enolase cDNA, TaENO-a and TaENO-b amplified by 5′,3′ rapid amplification of cDNA end (RACE)-PCR (polymerase chain reaction), were isolated and characterised. Quantitative real-time PCR (QPCR) was done to assess their expression patterns in leaf and crown tissues of wheat plants exposed to LT. BLAST searches and bioinformatic analyses were done to determine the structure, domains and phylogeny of the cloned sequences. The two cDNA sequences differed mostly in the 5′ and 3′ untranslated regions. Deduced amino acid sequence showed high identity to bacteria, yeast, fungi, human and plant enolases with conserved putative DNA-binding and repressor domains. A genomic clone containing 17 exons distributed over 4.5 kb structurally shared a high degree of similarity to rice enolase. QPCR revealed combined effects of LT and ageing on expression of TaENO-a and TaENO-b. Down-regulation of TaENO-a was observed with age in the crown tissues upon exposure to LT, but in leaf initial up-regulation was followed by down-regulation. Expression of TaENO-b was similar to expression patterns previously reported for cold-regulated (COR) genes in wheat, wherein the recessive vrnA-1 allele influenced its expression in the leaf and genetic background determines its expression in the crown.
DOI:10.1104/pp.90.2.367URL [本文引用: 1]
DOI:10.1016/0031-9422(92)80322-6URLPMID:1368216 [本文引用: 1]
Abstract NADP-malic enzyme functions in plant metabolism as a decarboxylase of malate in the chloroplast or cytosol. It serves as a source of CO2 for photosynthesis in the bundle sheath chloroplasts of C4 plants and in the cytosol of Crassulacean acid metabolism plants, and as a source of NADPH and pyruvate in the cytosol of various tissues. Mg2+ or Mn2+ is required as a cofactor. The enzyme has a high specificity and low Km for NADP+. It exists as a tetramer which may undergo changes in oligomerization and exhibit hysteresis. Its kinetic properties vary depending on the compartmentation and function of the enzyme. The chloroplast form in C4 plants has a high pH optimum (pH 8) under high malate, which favours the tetramer, whereas lower pH (pH 7) favours the dimer form. Generally, other forms of the enzyme, which are thought to be cytosolic, have lower pH optima than the chloroplast enzyme. In a number of cases these forms have been shown to have allosteric properties with malate as a substrate. Chemical modifications of the plant enzyme suggest sulphydryl, histidine and arginine residues are required for catalysis. Primary sequence studies on the chloroplastic enzyme from C4 plants show significant similarities to cytosolic NADP-ME in plants and animals, including a sequence motif which is indicative of the NADP+ binding site. The possible origin of the chloroplast form of the enzyme is discussed.
DOI:10.1016/j.crvi.2009.03.002URLPMID:19523599 [本文引用: 1]
The NADP-dependent malic enzyme (NADP-ME; EC1.1.1.40) found in many metabolic pathways catalyzes the oxidative decarboxylation of L-malate, producing pyruvate, CO 2 and NADPH. The NADP-MEs have been well studied in C4 plants but not well in C3 plants. In this study, we identified the NADP-ME isoforms from hexaploid wheat ( Triticum aestivum L). Two different NADP-ME transcripts were first identified in this C3 plant. The first is named TaNADP-ME1 [NCBI: EU170134] and encodes a putative plastidic isoform, while the second is named TaNADP-ME2 [NCBI: EU082065] and encodes a cytosolic counterpart. Sequence alignment shows that the two NADP-ME isoforms share an identity of 73.26% in whole amino acids and 64.08% in nucleotide sequences. The phylogenetic analysis deciphers the two NADP-MEs as belonging to the monocots (Group II), which closely resemble OschlME6 and OscytME2, respectively. Tissue-specific analyses indicate that the two NADP-ME genes are both expressed in root, stem and leaf, and that TaNADP-ME1 is a leaf-abundant isoform. Semi-quantitative RT-PCR analysis show that the two NADP-ME transcripts in wheat leaves respond differently to low temperature, salt, dark and drought stresses stimuli and to exogenous abscisic acid (ABA) and salicylic acid (SA). Our results demonstrate that exogenous hormones (ABA and SA), as well as salt, low temperature, dark and drought stresses can regulate the expressions of TaNADP-ME1 and TaNADP-ME2 in wheat. This indicates that the two NADP-ME genes may play an important role in the response of wheat to ABA, SA, low temperature, salt, dark and drought stress. To cite this article: Z.-Y. Fu et al., C. R. Biologies 332 (2009).
DOI:10.1186/s12864-016-2527-3URLPMID:4793594 [本文引用: 1]
Background Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) is a central enzyme in glycolysi, we performed genome-wide identification of GAPDHgenes in wheat and analyzed their structural...
DOI:10.1007/s10059-012-0230-zURLPMID:3887832 [本文引用: 1]
We previously reported that one of the brassinosteroidinsensitive mutants, bri1-9 , showed increased cold tolerance compared with both wild type and BRI1 -overexpressing transgenic plants, despite its severe growth retardation. This increased tolerance in bri1-9 resulted from the constitutively high expression of stress-inducible genes under normal conditions. In this report, we focused on the genes encoding class III plant peroxidases ( AtPrxs ) because we found that, compared with wild type, bri1-9 plants contain higher levels of reactive oxygen species (ROS) that are not involved with the activation of NADPH oxidase and show an increased level of expression of a subset of genes encoding class III plant peroxidases. Treatment with a peroxidase inhibitor, salicylhydroxamic acid (SHAM), led to the reduction of cold resistance in bri1-9 . Among 73 genes that encode AtPrxs in Arabidopsis, we selected four ( AtPrx1, AtPrx22, AtPrx39 , and AtPrx69 ) for further functional analyses in response to cold temperatures. T-DNA insertional knockout mutants showed increased sensitivity to cold stress as measured by leaf damage and ion leakage. In contrast, the overexpression of AtPrx22, AtPrx39 , and AtPrx69 increased cold tolerance in the BRI1-GFP plants. Taken together, these results indicate that the appropriate expression of a particular subset of AtPrx genes and the resulting higher levels of ROS production are required for the cold tolerance.
DOI:10.1371/journal.pone.0057472URLPMID:23468992 [本文引用: 1]
Plant ascorbate peroxidases (APXs), enzymes catalyzing the dismutation of H2O2 into H2O and O2, play an important role in reactive oxygen species homeostasis in plants. The rice genome has eight OsAPXs, but their physiological functions remain to be determined. In this report, we studied the function of OsAPX2 gene using a T-DNA knockout mutant under the treatment of drought, salt and cold stresses. The Osapx2 knockout mutant was isolated by a genetic screening of a rice T-DNA insertion library under 20% PEG-2000 treatment. Loss of function in OsAPX2 affected the growth and development of rice seedlings, resulting in semi-dwarf seedlings, yellow-green leaves, leaf lesion mimic and seed sterility. OsAPX2 expression was developmental- and spatial-regulated, and was induced by drought, salt, and cold stresses. Osapx2 mutants had lower APX activity and were sensitive to abiotic stresses; overexpression of OsAPX2 increased APX activity and enhanced stress tolerance. H2O2 and MDA levels were high in Osapx2 mutants but low in OsAPX2-OX transgenic lines relative to wild-type plants after stress treatments. Taken together, the cytosolic ascorbate peroxidase OsAPX2 plays an important role in rice growth and development by protecting the seedlings from abiotic stresses through scavenging reactive oxygen species.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}