关键词:水稻; MAGIC群体; 抽穗期; 株高; 关联分析; QTL Mapping of QTLs for Heading Date and Plant Height Using MAGIC Populations of Rice SHEN Cong-Cong1,2, ZHU Ya-Jun1,2, CHEN Kai1,2, CHEN Hui-Zhen3, WU Zhi-Chao1,2, MENG Li-Jun2, XU Jian-Long1,2,4,* 1 Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
2 Agricultural Genomics Institute, Chinese Academy of Agricultural Sciences, Shenzhen 518210, China
3 Pingxiang Institute of Agricultural Science, Pingxiang 337000, China
4 Shenzhen Institute of Breeding & Innovation, Chinese Academy of Agricultural Sciences, Shenzhen 518120, China
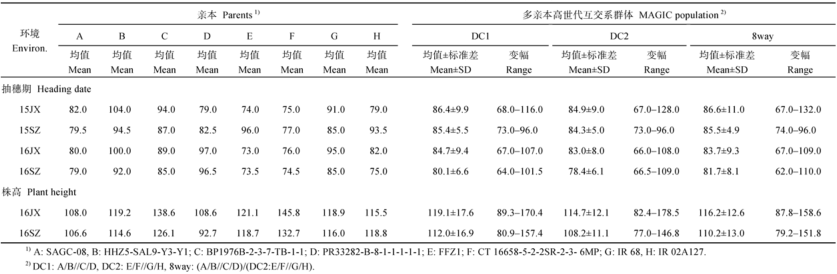
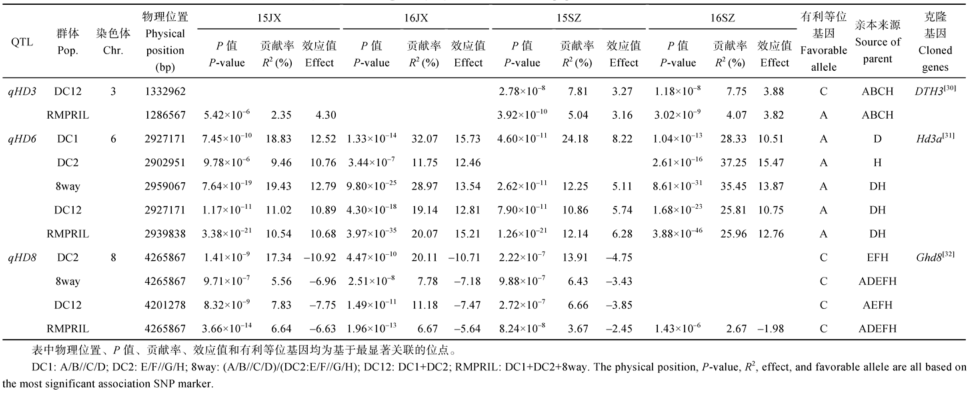
Fund:This study was supported by the National High Technology Research and Development Program of China (863 Program) (2014AA10A601), The Program of Introducing International Super Agricultural Science and Technology (948 Program) (2016-X16), Shenzhen Peacock Plan (20130415095710361), and the Scientific and Technological Innovation Project of Chinese Academy of Agricultural Sciences AbstractAssociation analysis was performed to map QTLs for heading date (HD) and plant height (PH) using three multi-parent advanced generation inter-cross (MAGIC) populations including two four-parent- and one eight-parent-derived populations (DC1, DC2, and 8way), and their composite populations (DC12=DC1+DC2, RMPRIL=DC12+8way) derived from eight elite indica lines. Using the phenotypic data of HD collected in the two sites (Jiangxi and Shenzhen) across two years (2015 and 2016), PH collected in the two sites (Jiangxi and Shenzhen) in 2016, and SNP genotypic data of rice 55K SNP chip, three HD QTLs, qHD3, qHD6and qHD8 were identified in the same regions of DTH3, Hd3a, and Ghd8, respectively. And five PH QTLs ( qPH1.1, qPH1.2, qPH1.3, qPH4, and qPH6) were also identified, of which qPH1.1 and qPH1.2 were located in the adjacent regions of Psd1 and sd1. The other three PH QTLs were new loci which were detected only in one population under one environment showing larger effects of genetic background and environment. Different MAGIC populations had different mapping effects for HD and PH. Using eight-parent-derived populations such as 8way, DC12 and RMPRIL not only detected more HD and PH QTLs (5, 5, and 6) but also had higher mapping resolution than four parents-derived populations such as DC1 and DC2 (2 and 4), showing an obvious advantage in two aspects of QTL mapping:highly significant level and closed to known genes. Especially, joint analyses of different populations such as DC12 and RMPRIL had significant advantage in QTL mapping.
Keyword:Rice; MAGIC population; Heading date; Plant height; Association analysis; Quantitative trait locus/loci (QTL) Show Figures Show Figures
Chang TT, Li CC, Vergara BS. Component analysis of duration from seeding to heading in rice by the basic vegetative phase and the photoperiod-sensitive phase. Euphytica, 1969, 18:79-91[本文引用:1]
[2]
岳兵, 邢永忠. 水稻抽穗期分子遗传研究进展. 分子植物育种, 2005, 3:222-228YueB, Xing YZ. Progress on molecular and genetic studies of heading date in rice. Mol Plant Breed, 2005, 3:222-228 (in Chinese with English abstract)[本文引用:1]
[3]
孙旭初. 水稻茎杆抗倒性的研究. 中国农业科学, 1987, 20(4):32-37Sun XC. Studies on the resistance of the culm of rice to lodging. Sci Agric Sin, 1987, 20(4):32-37 (in Chinese with English abstract)[本文引用:1]
[4]
李荣田, 姜廷波, 秋太权, 崔成焕, 龚振平. 水稻倒伏对产量影响及倒伏和株高关系的研究. 黑龙江农业科学, 1996, (1):13-17Li RT, Jiang TB, Qiu TQ, Cui CH, Gong ZP. Study on effect of loading to yield and relationship between lodging and plant height in rice. Agric Sci Heilongjiang, 1996, (1):13-17 (in Chinese with English abstract)[本文引用:1]
[5]
林鸿宣, 钱惠荣, 熊振民, 闵绍楷, 郑康乐. 几个水稻品种抽穗期主效基因与微效基因的定位研究. 遗传学报, 1996, 23:205-213Lin HX, Qian HR, Xiong ZM, Min SK, Zheng KL. Mapping of major genes and minor genes for heading date in several rice varieties. Acta Genet Sin, 1996, 23:205-213 (in Chinese with English abstract)[本文引用:1]
[6]
邱磊, 蒋海潮, 冯玉涛, 高冠军, 张庆路, 何予卿. 控制水稻抽穗期和株高的 QTL 的定位及遗传分析. 基因组学与应用生物学, 2014, 33:828-835QiuL, Jiang HC, Feng YT, Gao GJ, Zhang QL, He YQ. Mapping and genetic analysis of QTL for heading date and plant height in rice. Genom Appl Biol, 2014, 33:828-835 (in Chinese with English abstract)[本文引用:1]
[7]
李秀兰, 徐承水. 水稻株高基因及其在育种上的应用. 山东农业科学, 2009, (10):24-28Li XL, Xu CS. Plant height genes in rice and their application to breeding. Agric Sci Shand ong, 2009, (10):24-28 (in Chinese with English abstract)[本文引用:1]
[8]
杜雪树, 戚华雄, 廖世勇, 方非. 水稻抽穗期分子生物学研究进展. 湖北农业科学, 2013, 52:5958-5962Du XS, Qi HX, Liao SY, FangF. Advances on the molecular biology of rice heading date. Hubei Agric Sci, 2013, 52:5958-5962 (in Chinese with English abstract)[本文引用:1]
[9]
胡时开, 苏岩, 叶卫军, 郭龙彪. 水稻抽穗期遗传与分子调控机理研究进展. 中国水稻科学, 2012, 26:373-382Hu SK, SuY, Ye WJ, Guo LB. Advances in genetic analysis and molecular regulation mechanism of heading date in rice (Oryza sativa L. ). Chin J Rice Sci, 2012, 26:373-382 (in Chinese with English abstract)[本文引用:1]
[10]
张云辉, 张所兵, 林静, 汪迎节, 方先文. 水稻株高基因克隆及功能分析的研究进展. 中国农学通报, 2014, 30(12):1-7Zhang YH, Zhang SB, LinJ, Wang YJ, Fang XW. Research progress on cloning and functional analysis of plant height genes in rice (Oryza sativa L. ). Chin Agric Sci Bull, 2014, 30(12):1-7 (in Chinese with English abstract)[本文引用:1]
[11]
HoriK, MatsubaraK, YanoM. Genetic control of flowering time in rice:integration of Mendelian genetics and genomics. Int J Polym Anal Charact, 2010, 67:717-725[本文引用:1]
[12]
Guo LB, YeG. Use of major quantitative trait loci to improve grain yield of rice. Rice Sci, 2014, 21(2):65-82[本文引用:1]
[13]
Flint-Garcia SA, Thornsberry JM, Buckler IV ES. Structure of linkage disequilibrium in plants. Annu Rev Plant Biol, 2003, 54:357-374[本文引用:1]
[14]
YuJ, Buckler ES. Genetic association mapping and genome organization of maize. Curr Opin Biotechnol, 2006, 17:155-160[本文引用:2]
[15]
SalviS, TuberosaR. To clone or not to clone plant QTLs:present and future challenges. Trends Plant Sci, 2005, 10:297-304[本文引用:1]
[16]
ZhaoK, Tung CW, Eizenga GC, ZhaoK, Tung CW, Eizenga GC, Wright MH, Ali ML, Price AH, Norton GJ, Islam MR, ReynoldsA, MezeyJ, McClung A M, Bustamante C D, McCouch S R. Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat Commun, 2011, 2:467[本文引用:1]
[17]
Lipka AE, Kand ianis CB, Hudson ME, YuJ, DrnevichJ, Bradbury PJ, Gore MA. From association to prediction:statistical methods for the dissection and selection of complex traits in plants. Curr Opin Plant Biol, 2015, 24:110-118[本文引用:1]
[18]
Kover PX, ValdarW, TrakaloJ, ScarcelliN, Ehrenreich IM, Purugganan MD, DurrantC, MottR. A multiparent advanced generation inter-cross to fine-map quantitative traits in Arabidopsis thaliana. PLoS Genet, 2009, 5(7):e1000551[本文引用:2]
[19]
Higgins RH, Thurber CS, AssaranurakI, Brown PJ. Multiparental mapping of plant height and flowering time QTL in partially isogenic sorghum families. G3:Genes Genom Genet, 2014, 4:1593-1602[本文引用:1]
[20]
Mackay IJ, Bansept-BaslerP, BarberT, Bentley AR, CockramJ, GosmanN, Greenland AJ, HorsnellR, HowellsR, O’Sullivan D M, Rose G A, Howell P J. An eight-parent multiparent advanced generation inter-cross population for winter-sown wheat:creation, properties, and validation. G3:Genes Genom Genet, 2014, 4:1603-1610[本文引用:1]
[21]
RakshitS, RakshitA, Patil JV. Multiparent intercross populations in analysis of quantitative traits. J Genet, 2012, 91:111-117[本文引用:2]
[22]
LiZ, YeG, Yang ME, Liu ZX, Lu DB, Mao XX, Wu QH, Li XF. Genetic characterization of a multiparent recombinant inbred line rice population. Res Crops, 2014, 15:28-37[本文引用:1]
[23]
Rebetzke GJ, Verbyla AP, Verbyla KL, Morell MK, Cavanagh CR. Use of a large multiparent wheat mapping population in genomic dissection of coleoptile and seedling growth. Plant Biotechnol J, 2014, 12:219-230[本文引用:1]
[24]
ValdarW, FlintJ, MottR. Simulating the collaborative cross:power of quantitative trait loci detection and mapping resolution in large sets of recombinant inbred strains of mice. Genetics, 2006, 172:1783-1797[本文引用:1]
[25]
Broman KW. Genotype probabilities at intermediate generations in the construction of recombinant inbred lines. Genetics, 2012, 190:403-412[本文引用:1]
[26]
YamamotoE, IwataH, TanabataT, MizobuchiR, YonemaruJ, YamamotoT, YanoM. Effect of advanced intercrossing on genome structure and on the power to detect linked quantitative trait loci in a multi-parent population:a simulation study in rice. BMC Genet, 2014, 15(1):50[本文引用:1]
[27]
MengL, GuoL, PonceK, ZhaoX, YeG. Characterization of three rice multiparent advanced generation intercross (MAGIC) action. Plant Genome, 2016, 9(2):1-14[本文引用:6]
[28]
MengL, ZhaoX, PonceK, YeG, LeungH. QTL mapping for agronomic traits using multi-parent advanced generation inter- cross (MAGIC) populations derived from diverse elite indica rice lines. Field Crops Res, 2016, 189:19-42[本文引用:5]
[29]
陈天晓, 朱亚军, 密雪飞, 陈凯, 孟丽君, 左示敏, 徐建龙. 利用水稻MAGIC群体关联定位白叶枯病抗性QTL和创制抗病新种质. 作物学报, 2016, 42:1437-1447Chen TX, Zhu YJ, Mi XF, ChenK, Meng LJ, Zuo SM, Xu JL. Mapping of QTLs for bacterial blight resistance and screening of resistant materials using MAGIC populations of rice. Acta Agron Sin, 2016, 42:1437-1447 (in Chinese with English abstract)[本文引用:2]
[30]
Bian XF, LiuX, Zhao ZG, JiangL, GaoH, Zhang YH, ZhengM, Chen LM, Liu SJ, Zhai HQ, Wan JM. Heading date gene, dth3 controlled late flowering in O. glaberrima Steud. by down-regulating Ehd1. Plant Cell Rep, 2011, 30:2243-2254[本文引用:1]
[31]
TaokaK, OhkiI, TsujiH, FuruitaK, HayashiK, YanaseT, YamaguchiM, NakashimaC, Purwestri YA, TamakiS, OgakiY, ShimadaC, NakagawaA, KojimaC, ShimamotoK. 14-3-3 proteins act as intracellular receptors for rice Hd3a florigen. Nature, 2011, 476:332-335[本文引用:1]
[32]
Yan WH, WangP, Chen HX, Zhou HJ, Li QP, Wang CR, Ding ZH, Zhang YS, Yu SB, Xing YZ, Zhang QF. A major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice. Mol Plant, 2011, 4:319-330[本文引用:1]
[33]
LiR, XiaJ, XuY, ZhaoX, Liu YG, ChenY. Characterization and genetic mapping of a photoperiod-sensitive dwarf 1 locus in rice (Oryza sativa L. ). Theor Appl Genet, 2014, 127:241-250[本文引用:1]
[34]
MonnaL, KitazawaN, YoshinoR, SuzukiJ, MasudaH, MaeharaY, TanjiM, SatoM, NasuS, MinobeY. Positional cloning of rice semidwarfing gene, sd-1:rice “green revolution gene” encodes a mutant enzyme involved in gibberellin synthesis. DNA Res, 2002, 9:11-17[本文引用:1]
[35]
ZhaoM, LiuB, WuK, YeY, HuangS, WangS, WangY, HanR, LiuQ, FuX, WuY. Regulation of OsmiR156h through alternative polyadenylation improves grain yield in rice. PLoS One, 2015, 10(5):e0126154[本文引用:1]
[36]
PascualL, DesplatN, Huang BE, DesgrouxA, BruguierL, Bouchet JP, Le QH, ChauchardB, VerschaveP, CausseM. Potential of a tomato MAGIC population to decipher the genetic control of quantitative traits and detect causal variants in the resequencing era. Plant Biotechnol J, 2015, 13:565-577[本文引用:1]
[37]
StichB, MöhringJ, Piepho HP, HeckenbergerM, Buckler ES, Melchinger AE. Comparison of mixed-model approaches for association mapping. Genetics, 2008, 178:1745-1754[本文引用:1]
[38]
王韵, 程立锐, 孙勇, 周政, 朱苓华, 徐正进, 徐建龙, 黎志康. 利用双向导入系解析水稻抽穗期和株高QTL及其与环境互作表达的遗传背景效应. 作物学报, 2009, 35:1386-1394WangY, Cheng LR, SunY, ZhouZ, Zhu LH, Xu ZJ, Xu JL, L Z K. Genetic background effect on QTL expression of heading date and plant height and their interaction with environment in reciprocal introgression lines of rice. Acta Agron Sin, 2009, 35:1386-1394 (in Chinese with English abstract)[本文引用:1]
[39]
Lee YS, AnG. Regulation of flowering time in rice. J Plant Biol, 2015, 58:353-360[本文引用:1]
 , 朱亚军
, 朱亚军