关键词:玉米; 弱势粒; 库活性; 激素; 多胺 Relationship of Sink Activity with Endogenous Hormones and Polyamine Contents in Inferior Kernels of Maize WANG Zhi-Gang1,**, LIANG Hong-Wei1,2,**, GAO Ju-Lin1,*, YU Xiao-Fang1, SUN Ji-Ying1, SU Zhi-Jun1, HU Shu-Ping1, YU Shao-Bo1, LI Ya-Jian1, WEI Shu-Li1, YANG Zhe1 1 College of Agronomy, Inner Mongolia Agricultural University, Hohhot 010019, China
2 Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences, Hohhot 010031, China
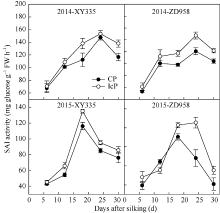
Fund:This study was supported by the National Natural Science Foundation of China (31301265), the State Program of China (2016YFD0300103), the Program for Young Talents of Science and Technology in Universities of Inner Mongolia Autonomous Region (NJYT-14-B04), and the National Key Technology Support Program of China (2013BAD07B04) AbstractSink activity at blister stage of maize is the primary limiting factor of the abortion and/or filling stagnation of inferior kernels. Clarifying the effect of endogenous hormone and polyamine contents on sink activity of inferior kernels in maize is of great importance for regulating inferior kernels in crowding maize colony. In this present study, two typical commercial maize hybrids Zhengdan 958 and Xianyu 335 were planted in 2014 and 2015. At silking stage, two pollination treatments, incomplete pollination (IcP) and complete pollination (CP), were imposed to each cultivar. The objective of incomplete pollination, in which the pollinated filaments within the basal region of ear was decreased by hand-cutting, was to stimulate the inferior kernel setting in the apical region of ear, where the inferior kernels should be aborted or filling-stagnated in complete pollination. The endogenous hormone and polyamine contents were compared between IcP and CP treatments during blister stage, and the relationship of sink activity with endogenous hormone and/or polyamine contents was analyzed. There was no remarkable effect of hybrids and years on sink activity, endogenous hormone and polyamine of kernels. The soluble acid invertase activity of IcP was significantly higher than that of CP, by 13.5% on an average and 21.8% at the maximum. There was no significant difference in contents of Z+ZR, IAA, GA3, and ABA of the inferior kernels between IcP and CP treatments during blister stage. The polyamine content of IcP was significantly higher than that of CP, but the ethylene producing rate was just the opposite. SAI activity correlated with polyamine content positively, but with ethylene release rate negatively, moreover, ethylene release rate showed negative correlation with polyamine content. The results suggested that, during the blister stage of maize, the contents of Z+ZR, IAA, GA3, and ABA of inferior kernels have nothing to do with SAI activity. SAI activity of inferior kernels is mainly affected by polyamine and ethylene, which is promoted by polyamine content and inhibited by ethylene release rate. The balance between polyamine and ethylene determines the result of inferior kernel development, which is radically regulated by assimilate supply of maize plant.
表1 不同授粉处理下玉米弱势粒的Z+ZR、IAA、GA3和ABA含量 Table 1 Z+ZR, IAA, GA3, and ABA content of inferior kernels of maize under different pollination treatments
激素 Hormone
年度 Year
品种 Hybrid
处理 Treatment
含量 Content (ng g-1 FW)
6 d
12 d
18 d
24 d
30 d
玉米素+玉米素核苷 Z+ZR
2014
先玉335
CP
7.59 b
13.43 a
14.41 a
16.09 a
15.92 a
Xianyu 335
IcP
9.56 a
14.47 a
15.64 a
16.43 a
14.33 a
郑单958
CP
6.54 b
13.44 a
15.00 a
15.31 a
11.71 a
Zhengdan 958
IcP
9.81 a
12.84 a
16.13 a
14.90 a
11.86 a
2015
先玉335
CP
4.16 a
9.32 a
15.62 a
15.25 a
9.17 a
Xianyu 335
IcP
3.99 a
11.18 a
16.05 a
12.28 a
9.33 a
郑单958
CP
4.28 a
7.92 a
15.35 a
14.90 a
11.18 a
Zhengdan 958
IcP
4.45 a
8.56 a
13.77 a
12.41 a
12.07 a
生长素 IAA
2014
先玉335
CP
59.16 a
70.89 a
94.64 a
58.27 a
52.19 a
Xianyu 335
IcP
56.42 a
68.34 a
85.97 b
61.26 a
58.74 a
郑单958
CP
44.96 a
78.85 a
108.14 a
87.65 a
74.79 a
Zhengdan 958
IcP
48.40 a
73.45 b
101.64 a
82.32 a
73.45 a
2015
先玉335
CP
16.88 a
55.03 a
86.00 b
80.57 a
72.26 a
Xianyu 335
IcP
15.40 a
36.55 b
102.58 a
85.13 a
73.91 a
郑单958
CP
14.50 a
34.78 b
88.24 a
101.99 a
81.81 a
Zhengdan 958
IcP
17.69 a
48.86 a
83.13 a
81.93 b
77.35 a
脱落酸 ABA
2014
先玉335
CP
103.05 a
78.89 a
48.45 a
58.55 a
68.00 a
Xianyu 335
IcP
93.47 a
68.23 a
57.03 a
80.03 a
57.16 a
郑单958
CP
77.81 a
47.08 a
62.43 a
78.88 a
60.46 a
Zhengdan 958
IcP
87.51 a
58.03 a
57.09 a
78.06 a
63.04 a
2015
先玉335
CP
87.77 a
81.69 a
97.67 a
90.08 a
121.46 a
Xianyu 335
IcP
108.08 a
86.00 a
88.62 a
82.90 a
124.56 a
郑单958
CP
102.62 a
98.76 a
88.87 a
92.21 a
133.72 a
Zhengdan 958
IcP
116.18 a
94.08 a
87.71 a
90.01 a
133.55 a
赤霉素 GA3
2014
先玉335
CP
7.49 a
6.41 a
5.97 a
5.01 a
4.11 a
Xianyu 335
IcP
7.36 a
5.91 a
5.13 a
4.33 a
3.88 a
郑单958
CP
8.14 a
6.42 a
5.94 a
5.85 a
4.30 a
Zhengdan 958
IcP
7.73 b
6.44 a
5.99 a
5.24 a
4.06 a
2015
先玉335
CP
7.22 a
6.74 a
6.25 a
5.52 a
4.70 a
Xianyu 335
IcP
7.30 a
6.33 a
6.06 a
5.61 a
4.60 a
郑单958
CP
7.98 a
6.89 a
5.72 a
5.59 a
4.54 a
Zhengdan 958
IcP
7.92 a
7.68 a
6.18 a
5.70 a
4.47 a
CP: complete pollination; IcP: incomplete pollination. Values within a column followed by different letters are significantly different at the 0.05 probability level among different treatments. CP: 完全授粉; IcP: 不完全授粉。同一列数字后不同小写字母表示不同处理间差异达0.05显著水平。
表1 不同授粉处理下玉米弱势粒的Z+ZR、IAA、GA3和ABA含量 Table 1 Z+ZR, IAA, GA3, and ABA content of inferior kernels of maize under different pollination treatments
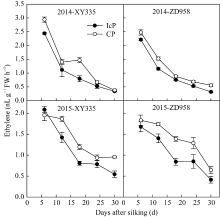
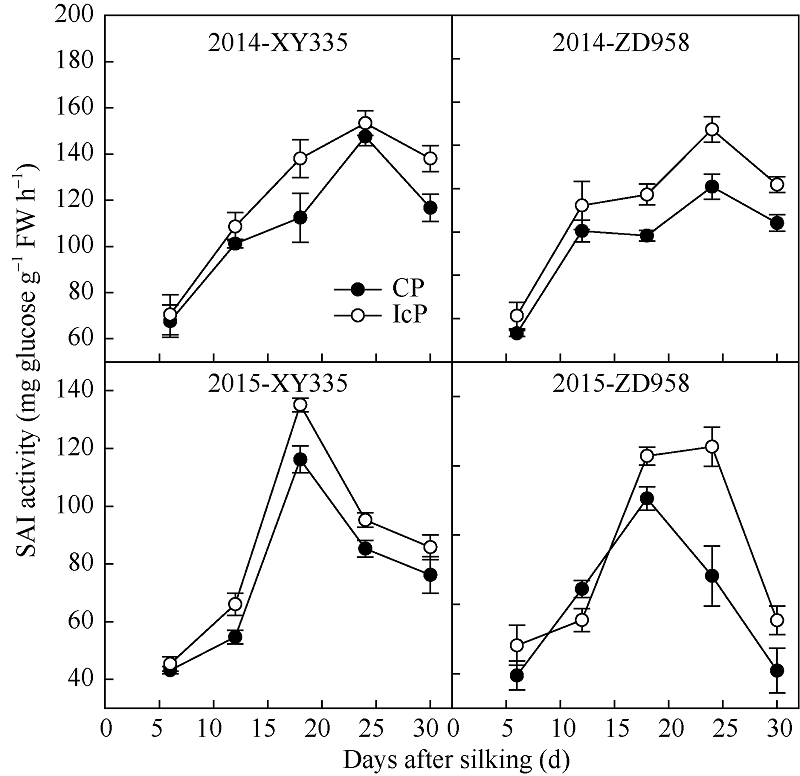
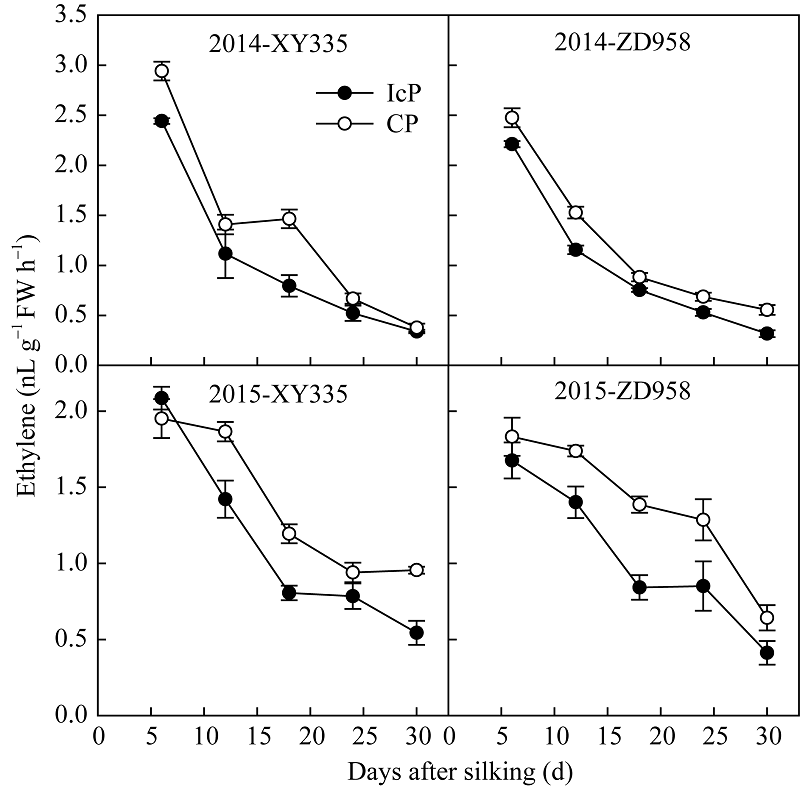
图3 不同控制授粉处理下玉米弱势粒乙烯释放速率变化 缩写同图2。Fig. 3 Dynamics of ethylene producing rate of inferior kernels of maize under different pollination treatments Abbreviations are the same as those given in Fig. 2.
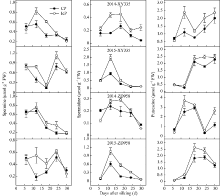
图4 不同控制授粉处理下玉米弱势粒多胺含量变化 缩写同图2。Fig. 4 Change of polyamine content of inferior kernels of maize under different pollination treatments Abbreviations are the same as those given in Fig. 2.
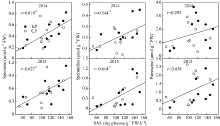
图5 玉米籽粒形成期弱势粒乙烯释放率与SAI活性的关系 缩写同图2。Fig. 5 Relationship between ethylene release rate and soluble acid invertase (SAI) activity of inferior kernels during the blister stage of maize Abbreviations are the same as those given in Fig. 2.
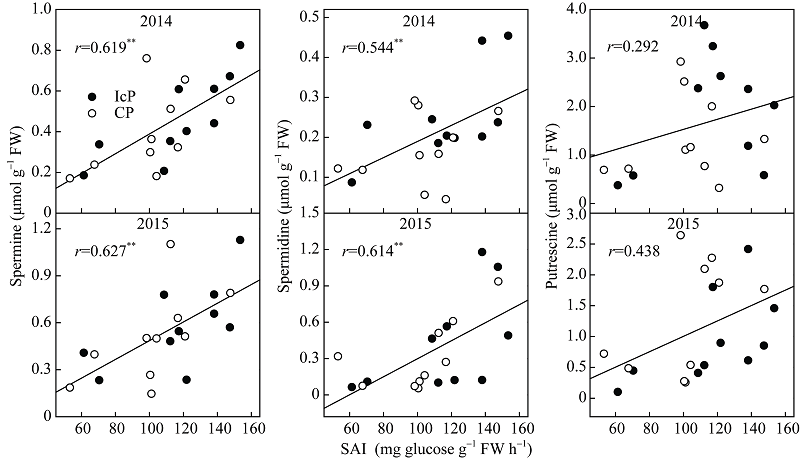
图6 玉米籽粒形成期弱势粒多胺含量与SAI活性的关系 缩写同图2。Fig. 6 Relationship between polyamine content and soluble acid invertase (SAI) of inferior kernels during the blister stage of maize Abbreviations are the same as those given in Fig. 2.
图7 玉米籽粒形成期弱势粒多胺含量与乙烯释放率的关系 缩写同图2。Fig. 7 Relationship between polyamine content and ethylene release rate of inferior kernels during the blister stage of maize Abbreviations are the same as those given in Fig. 2.
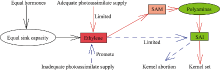
图8 乙烯、多胺平衡影响弱势粒库活性并主导弱势粒发育的模式图Fig. 8 Theoretical framework of effects of polyamines and ethylene contents and their balance on sink activity and inferior kernels setting
4 结论玉米籽粒形成期的弱势粒库活性即SAI活性受其内源多胺和乙烯含量及其平衡关系调控, 多胺促进SAI活性, 而乙烯抑制其活性。多胺和乙烯平衡受物质供应状况影响, 当同化物质供应充足时, 多胺大量合成抑制乙烯释放, 从而促进SAI活性使糖分向籽粒中卸载, 使弱势粒成功建成; 当同化物质供应不足时, 多胺合成受限而乙烯大量合成, 抑制SAI活性, 还原糖卸载受阻, 籽粒败育或灌浆受限。 The authors have declared that no competing interests exist.
Wang ZG, Gao JL, Ma BL. Concurrent improvement in maize yield and nitrogen use efficiency with integrated agronomic management strategies. Agron J, 2014, 106: 1243-1250[本文引用:1]
[2]
孙国伟, 张风路, 郭江, 赵久然, 郭景伦. 对玉米籽粒败育原因的再认识. 玉米科学, 2004, 12(增刊2): 35-37Sun GW, Zhang FL, GuoJ, Zhao JR, Guo JL. Discuss on the cause of maize kernel abortion. J Maize Sci, 2004, 12(suppl-2): 35-37 (in Chinese with English abstract)[本文引用:2]
[3]
申丽霞, 王璞, 张红芳, 易镇邪. 施氮对夏玉米不同部位籽粒灌浆的影响. 作物学报, 2005, 31: 532-534Shen LX, WangP, Zhang HF, Yi ZX. Effect of nitrogen supply on grain filling at different ear position in summer maize. Acta Agron Sin, 2005, 31: 532-534 (in Chinese with English abstract)[本文引用:1]
[4]
李玉玲, 台国琴. 玉米籽粒灌浆及种子萌发特性的粒位效应研究. 玉米科学, 2005, 13(1): 60-63Li YL, Tai GQ. Study on the kernel position effects of grain filling and seed germination characteristics in maize. J Maize Sci, 2005, 13(1): 60-63 (in Chinese with English abstract)[本文引用:1]
[5]
Yang JC, Zhang JH, Wang ZQ, Zhu QS. Hormones in the grains in relation to sink strength and postanthesis development of spikelets in rice. Plant Growth Regul, 2003, 41: 185-195[本文引用:3]
[6]
VenkateswarluB, Visperas RM. Source-sink relationships in crop plants. Int Rice Res Paper Ser, 1987, 125: 1-19[本文引用:2]
[7]
Warren WJ. Control of crop processes. In: Rees A R, Cockshull K E, Hand D W, Hurd R G, eds. Crop Processes in Controlled Environments. London/New York: Academic Press, 1972. pp 7-30[本文引用:2]
[8]
Ho LC. Metabolism and compartmentation of imported sugars in sink organs in relation to sink strength. Annu Rev Plant Physiol Plant Mol Biol, 1988, 39: 355-378[本文引用:1]
[9]
王纪华, 王树安, 赵冬梅, 倪军昌. 玉米籽粒发育的调控研究: III. 离体条件下的化学调控机理探讨. 作物学报, 1996, 22: 208-213Wang JH, Wang SA, Zhao DM, Ni JC. Study on regulation of development of maize kernels: III. The approach to chemical regulation under conditions of tissue culture. Acta Agron Sin, 1996, 22: 208-213 (in Chinese with English abstract)[本文引用:3]
[10]
唐祈林, 荣廷昭. 玉米秃尖与内源激素的关系. 核农学报, 2007, 21: 366-368Tang QL, Rong TZ. The relationship between endogenous hormone and barren ear tip of maize. J Nucl Agric Sci, 2007, 21: 366-368 (in Chinese with English abstract)[本文引用:2]
[11]
张凤路, 赵久然, 王纪华. 离体条件下玉米籽粒发育调控研究. 杂粮作物, 2001, 21(4): 34-35Zhang FL, Zhao JR, Wang JH. Regulating study on maize kernel development in vitro. Rain Fed Crops, 2001, 21(4): 34-35 (in Chinese with English abstract)[本文引用:3]
[12]
Pan YQ, Lou HL, Li YR. Soluble acid invertase and sucrose phosphate synthase: key enzymes in regulating sucrose accumulation in sugarcane stalk. J Sugar Tech, 2009, 11(1): 28-33[本文引用:2]
[13]
梁红伟, 高聚林, 王志刚, 于晓芳, 孙继颖, 胡树平, 王振, 余少波, 李雅剑. 基于控制授粉技术的玉米弱势粒发育与库特征的关系. 作物学报, 2016, 42: 927-933Liang HW, Gao JL, Wang ZG, Yu XF, Sun JY, Hu SP, WangZ, Yu SB, Li YJ. Relationship of inferior kernel setting with sink strength under controlling pollination. Acta Agron Sin, 2016, 42: 927-933 (in Chinese with English abstract)[本文引用:5]
[14]
徐云姬, 顾道健, 张博博, 张耗, 王志琴, 杨建昌. 玉米果穗不同部位籽粒激素含量及其与胚乳发育和籽粒灌浆的关系. 作物学报, 2013, 39: 1452-1461Xu YJ, Gu DJ, Zhang BB, ZhangH, Wang ZQ, Yang JC. Hormone contents in kernels at different positions on an ear and their relationship with endosperm development and kernel filling in maize. Acta Agron Sin, 2013, 39: 1452-1461 (in Chinese with English abstract)[本文引用:4]
[15]
徐云姬, 顾道建, 秦昊, 张耗, 王志琴, 杨建昌. 玉米灌浆期果穗不同部位籽粒碳水化合物积累与淀粉合成相关酶活性变化. 作物学报, 2015, 41: 297-307Xu YJ, Gu DJ, QingH, ZhangH, Wang ZQ, Yang JC. Changes in carbohydrate accumulation and activities of enzymes involved in starch synthesis in maize kernels at different positions on an ear during grain filling. Acta Agron Sin, 2015, 41: 297-307 (in Chinese with English abstract)[本文引用:3]
[16]
BangerthF. Dominance among fruits/sinks and the search for a correlative signal. Physiol Plant, 1989, 76: 608-614[本文引用:1]
[17]
Morris RD, Blevins DG, Dietrich JT, Durley RC, Gelvin SB, GrayJ, Hommes NG, KaminekM, Mathews LJ, MeilanR, Reinbott TM, Sayavedra-SotoL. Cytokinins in plant pathogenic bacteria and developing cereal grains. Aust J Plant Physiol, 1993, 20: 621-637[本文引用:2]
[18]
SinghG, Gerung SB. Hormonal role in the problem of sterility in Oryza sativa. Plant Physiol Biochem, 1982, 9: 22-23[本文引用:1]
[19]
孙庆泉, 吴元奇, 胡昌浩, 董树亭, 荣廷昭, 张颖. 不同产量潜力玉米子粒胚乳细胞增殖与子粒充实期的生理活性. 作物学报, 2005, 31: 612-618Sun QQ, Wu YQ, Hu CH, Dong ST, Rong TZ, ZhangY. Physiological activities and multiplication of endosperm cell at filling stage of kernels with different yield potential in maize. Acta Agron Sin, 2005, 31: 612-618 (in Chinese with English abstract)[本文引用:2]
[20]
冯汉宇, 王志敏, 孔凡娜, 张敏洁, 周顺利. 基于控制授粉技术的玉米籽粒生育特性与建成机制研究. 作物学报, 2011, 37: 1605-1615Feng HY, Wang ZM, Kong FN, Zhang MJ, Zhou SL. Mechanism of maize kernel setting and characteristics of kernel growth and development under controlled pollination. Acta Agron Sin, 2011, 37: 1605-1615 (in Chinese with English abstract)[本文引用:2]
[21]
Feng HY, Wang ZM, Kong FN, Zhang MJ, Zhou SL. Roles of carbohydrate supply and ethylene, polyamines in maize kernel set. J Integr Plant Biol, 2011, 53: 388-398[本文引用:3]
刘华英, 萧浪涛, 鲁旭东, 胡家金, 吴顺, 何长征, 邓秀新. 伏令夏橙愈伤组织体细胞胚发生中多胺水平的变化. 植物生理与分子生物学学报, 2005, 31: 275-280Liu HY, Xiao LT, Lu XD, Hu JJ, WuS, He CZ, Deng XX. Changes in polyamine levels in Citrus sinensis Osb. cv. valencia callus during somatic embryogenesis. J Plant Physiol Mol Biol, 2005, 31: 275-280 (in Chinese with English abstract)[本文引用:1]
[24]
王艳芳, 李红, 张立军, 马兴林, 关义新. 春玉米籽粒灌浆期IAA对淀粉积累的作用及其机制的研究. 化学与生物工程, 2006, 23(3): 35-37Wang YF, LiH, Zhang LJ, Ma XL, Guan YX. Effect and mechanism of IAA to starch accumulation in grain during grain-filling stage in spring maize. Chem Bioeng, 2006, 23(3): 35-37 (in Chinese with English abstract)[本文引用:1]
[25]
李连禄, 赵明, 丁在松. 玉米穗库发育的不同步性与内源激素作用的研究. 中国农业大学学报, 2001, 6(1): 30-36Li LL, ZhaoM, Ding ZS. Unsynchronism of the development of ear sinks and effects of endogenous hormones in maize. J China Agric Univ, 2001, 6(1): 30-36 (in Chinese with English abstract)[本文引用:1]
[26]
覃章铮, 唐锡华. 高等植物胚胎的发育生物学研究: VI. 粳稻胚分化发育期间一些大分子物质的动态. 植物生理学报, 1982, 8: 295-305Qin ZZ, Tang XH. Studies on the developmental biology of embryogenesis in higher plants: VI. Dynamic changes in some of the macromolecules during embryogenesis of Oryza sativa L. subsp. japonica. J Plant Physiol, 1982, 8: 295-305 (in Chinese with English abstract)[本文引用:1]
[27]
Reed AJ, Singletary GW. Roles of carbohydrate supply and phytohormones in maize kernel abortion. Plant Physiol, 1989, 91: 986-992[本文引用:1]
[28]
杨建昌, 彭少兵, 顾世梁, Visperas RM, 朱庆森. 水稻灌浆期籽粒中3个与淀粉合成有关的酶活性变化. 作物学报, 2001, 27: 157-164Yang JC, Peng SB, Gu SL, Visperas RM, Zhu QS. Changes in activities of three enzymes associated with starch synthesis in rice grains during grain filling. Acta Agron Sin, 2001, 27: 157-164 (in Chinese with English abstract)[本文引用:1]
[29]
Yang JC, Cao YY, ZhangH. Involvement of polyamines in the post-anthesis development of inferior and superior spikelets in rice. Planta, 2008, 228: 137-149[本文引用:2]
 , 梁红伟
, 梁红伟








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}