关键词:甘蓝型油菜; 半必需氨基酸; QTL定位; 胚; 母体植株; 遗传主效应; 环境互作效应 QTL Mapping and Analysis Based on Embryo and Maternal Genetic Systems for Semi-Essential Amino Acid Contents in Rapeseed ( Brassica napus L.) WEN Juan1, XU Jian-Feng1, LONG Yan2, XU Hai-Ming1, MENG Jin-Ling2, WU Jian-Guo3,*, SHI Chun-Hai1,* 1 College of Agriculture & Biotechnology, Zhejiang University, Hangzhou 310058, China
2 College of Plant Science & Technology, Huazhong Agricultural University, Wuhan 430070, China
3 College of Agriculture and Food Science, Zhejiang A&F University, Lin’an 311300, China
AbstractRapeseed meal is an important feed protein source, and the amino acid composition has a close relationship with the processing quality of feed, of which serine, cystine and tyrosine are the semi-essential amino acids for most animals. By using newly developed two-genetic-system QTL mapping software and method for analyzing seed quality traits of dicotyledonous plants, two backcross populations from a set of doubled haploid (DH) lines derived from an elite hybrid cross between Tapidor and Ningyou 7 were used to detect the QTLs simultaneously located in the amphidiploid embryo and maternal plant nuclear genomes for the semi-essential amino acid contents of rapeseed across environments. The results showed that five QTLs for serine content, two QTLs for cystine content and five QTLs for tyrosine content were identified, which were subsequently mapped on chromosomes A1, A4, A7, A8, A9, C2, C3, or C9 and could respectively explain 59.34%, 29.66%, and 59.26% of phenotypic variation in total. Five QTLs were major effect QTLs which could explain more than 10% of phenotypic variation for each. All of these QTLs had both notable embryo and maternal additive main effects, among which three QTLs were also found to have significant QE interaction effects. One QTL cluster on chromosome A4 was discovered to contain three QTLs related to serine, cystine and tyrosine contents. Some important QTLs and the tightly linked markers will have an important application value in the later map-based cloning and marker-assisted selection.
Keyword:Brassica napus L.; Semi-essential acid; QTL mapping; Embryo; Maternal plant; Genetic main effect; QTL#cod#x000D7;environment (QE) interaction effect Show Figures Show Figures
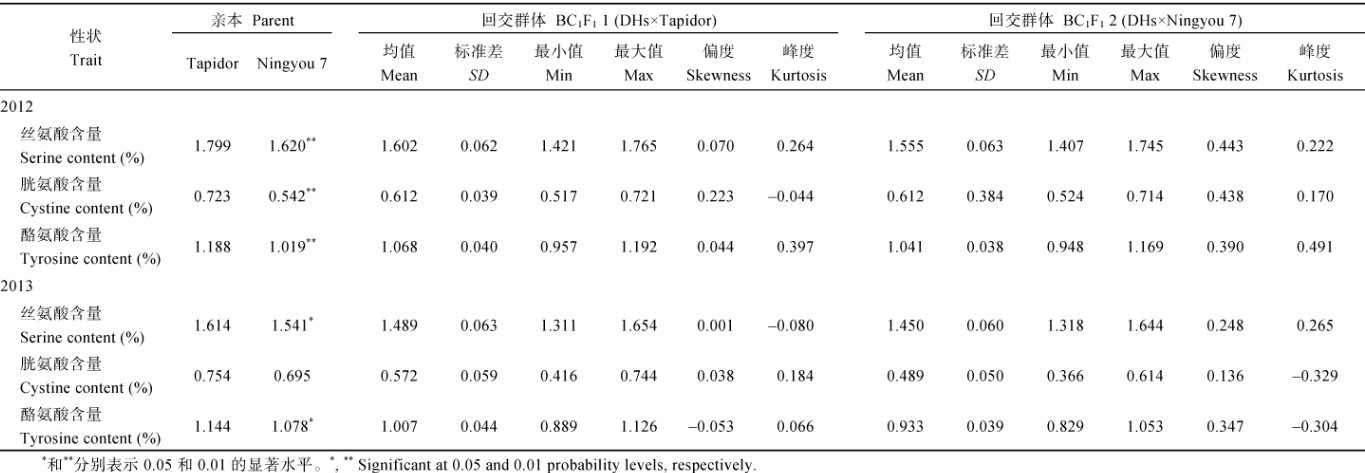
表 1 2012年和2013年亲本和回交群体油菜籽的半必需氨基酸含量 Table 1 Semi-essential amino acid contents of parents and the backeross populations of rapeseed DHs in 2012 and 2013
表 1 2012年和2013年亲本和回交群体油菜籽的半必需氨基酸含量 Table 1 Semi-essential amino acid contents of parents and the backeross populations of rapeseed DHs in 2012 and 2013
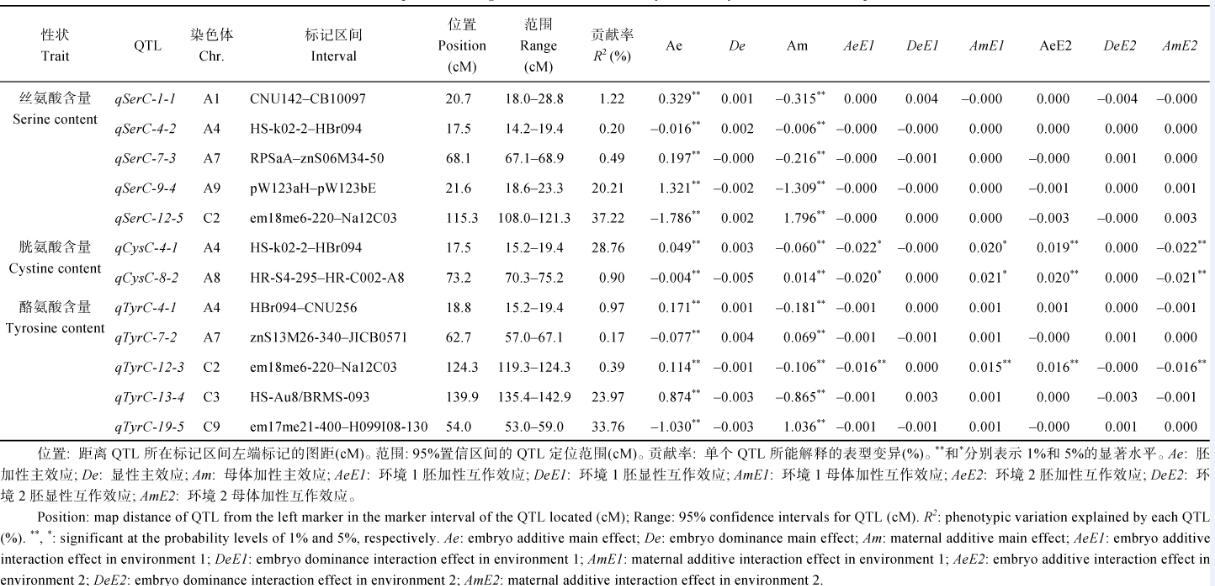
图1 油菜籽丝氨酸、胱氨酸和酪氨酸含量的QTL在染色体上的位置及其遗传效应Fig. 1 Positions (cM) and the genetic effects of QTLs for serine, cystine, and tyrosine contents in the chromosomes of rapeseed
王绍中, 李春喜, 罗艳蕊, 姜丽娜. 基因型和地域分布对小麦籽粒氨基酸含量影响的研究. 西北植物学报, 2001, 21: 437-445Wang SZ, Li CX, Luo YR, Jiang LN. Investigation on effects for genotypes and region distribution to the grain amino acid contents of winter wheat. Acta Bot Boreal-Occident Sin, 2001, 21: 437-445 (in Chinese with English abstract)[本文引用:1]
[2]
MunckaL, Pram NJ, MollerB, JacobsenS, SondergaardI, Engelsen SB, NorgaardL, BroaR. Exploring the phenotypic expression of a regulatory proteome-altering gene by spectroscopy and chemometrics. Anal Chim Acta, 2001, 446: 171-186[本文引用:1][JCR: 4.387]
[3]
Bell JM, RakowG, Downey RK. Comparisons of amino acid and protein levels in oil-extraeted seeds of Brassica and Sinapis species, with observations on environmental effects. Can J Anim Sci, 2000, 80: 169-174[本文引用:1][JCR: 0.956]
[4]
任玉玲, 石春海, 吴建国, 张海珍. 油菜籽三种氨基酸含量的胚、细胞质和母体遗传效应分析. 浙江大学学报(农业与生命科学版), 2005, 31: 41-46Ren YL, Shi CH, Wu JG, Zhang HZ. Genetic analysis of embryo, cytoplasmic and maternal effects on three amino acid traits in rapeseed. J Zhejiang Univ (Agric Life Sci), 2005, 31: 41-46 (in Chinese with English abstract)[本文引用:2][CJCR: 0.717]
[5]
Zeng ZB. Theoretical basis for separation of multiple linked gene effects in mapping quantitative trait loci. Proc Natl Acad Sci USA, 1993, 90: 10972-10976[本文引用:1][JCR: 9.737]
[6]
Jansen RC, StamP. High resolution of quantitative traits into multiple loci via interval mapping. Genetics, 1994, 136: 1447-1455[本文引用:1][JCR: 4.389]
[7]
ZhuJ, Weir BS, Chen LS. Mixed Model Approaches for Genetic Analysis of Quantitative Traits. In: Proceedings of the International Conference on Mathematical Biology. Singapore: World Scientific Publishing Co. , 1998. pp 321-330[本文引用:1]
[8]
Shi CH, ShiY, Lou XY, Xu HM, ZhengX, Wu JG. Identification of endosperm and maternal plant QTLs for protein and lysine contents of rice across different environments. Crop Pasture Sci, 2009, 60: 295-301[本文引用:2][JCR: 1.133]
[9]
Panthee DR, Pantalone VR, Sams CE, Saxton AM, West DR, Orf JH, Killam AS. Quantitative trait loci controlling sulfur containing amino acids, methionine and cysteine, in soybean seeds. Theor Appl Genet, 2006, 112: 546-553[本文引用:1][JCR: 3.658]
[10]
Jiang XL, Deng ZY, Ru ZG, WuP, Tian JC. Quantitative trait loci controlling amino acid contents in wheat (Triticum aestivum L. ). Aust J Crop Sci, 2013, 7: 820-829[本文引用:1][JCR: 1.632]
[11]
Liu HY, QuampahA, Chen JH, Li JR, Huang ZR, He QL, Zhu SJ, Shi CH. QTL Mapping based on different genetic systems for essential amino acid contents in cottonseeds in different environments. PLoS One, 2013, 8: e57531[本文引用:2][JCR: 3.73]
[12]
Xu JF, LongY, Wu JG, Xu HM, WenJ, Meng JL, Shi CH. QTL mapping and analysis of the embryo and maternal plant for three limiting amino acids in rapeseed meal. Eur Food Res Technol, 2014[本文引用:4][JCR: 1.436]
[13]
Zhang HZ, Shi CH, Wu JG, Ren YL, Li CT, Zhang DQ, Zhang YF. Analysis of genetic and genotype × environment interaction effects from embryo, cytoplasm and maternal plant for oleic acid content of Brassica napus L. Plant Sci, 2004, 167: 43-48[本文引用:2][JCR: 2.922]
[14]
Wu JG, Shi CH, Zhang HZ. Genetic analysis of embryo, cytoplasmic and maternal effects and their environment interactions for protein content in Brassica napus L. Aust J Agric Res, 2005, 56: 69-73[本文引用:2][JCR: 1.328]
[15]
Variath MT, WuJ, ZhangL, Shi CH. Analysis of developmental genetic effects from embryo, cytoplasm and maternal plant for oleic and linoleic acid contents of rapeseed. J Agric Sci, 2010, 148: 375-391[本文引用:2][JCR: 2.041]
[16]
YangJ, ZhuJ, Williams RW. Mapping the genetic architecture of complex traits in experimental populations. Bioinformatics, 2007, 23: 1527-1536[本文引用:3][JCR: 5.323]
[17]
郑希, 吴建国, 楼向阳, 徐海明, 石春海. 不同环境条件下稻米组氨酸和精氨酸的胚乳和母体植株QTL分析. 作物学报, 2008, 34: 369-375ZhengX, Wu JG, Lou XY, Xu HM, Shi CH. Mapping and analysis of QTLs on maternal and endosperm genomes for histidine and arginine in rice (Oryza sativa L. ) across environments. Acta Agron Sin, 2008, 34: 369-375 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[18]
Liu HY, QuampahA, Chen JH, Li JR, Huang ZR, He QL, Shi CH, Zhu SJ. QTL analysis for gossypol and protein contents in upland cottonseeds with two different genetic systems across environments. Euphytica, 2012, 188: 453-463[本文引用:1][JCR: 1.643]
[19]
QiuD, MorganC, ShiJ, LongY, LiuJ, LiR, ZhuangX, WangY, TanX, DietrichE, WeihmannT, EverettC, VanstraelenS, BeckettP, FraserF, TrickM, BarnesS, WilmerJ, SchmidtR, LiJ, LiD, MengJ, BancroftI. A comparative linkage map of oilseed rape and its use for QTL analysis of seed oil and erucic acid content. Theor Appl Genet, 2006, 114: 67-80[本文引用:1][JCR: 3.658]
[20]
Chen GL, ZhangB, Wu JG, Shi CH. Nondestructive assessment of amino acid composition in rapeseed meal based on intact seeds by near-infrared reflectance spectroscopy. Anim Feed Sci Tech, 2011, 165: 111-119[本文引用:1][JCR: 1.608]
[21]
Shi JQ, Li RL, QiuD, Jiang CC, LongY, MorganC, BancroftI, Zhao JY, Meng JL. Unraveling the complex trait of crop yield with quantitative trait loci mapping in Brassica napus. Genet, 2009, 182: 851-861[本文引用:1][JCR: 4.389]
[22]
McCouch SR, Cho YG, Yano PE, BlinstrubM, MorishimaH, KinoshitaT. Report on QTL nomenclature. Rice Genet Newslett, 1997, 14: 11-13[本文引用:1]
[23]
Falconer DS, MackayT F C. Introduction to Quantitative Genetics. 4th Edn. UK: Longmans Green, Harlow, Essex, 1996[本文引用:1]
[24]
Paterson AH, Land er ES, Hewitt JD, PetersonS, Lincoln SE, Tanksley SD. Resolution of quantitative traits into Mendelian factors using a complete linkage map of restriction fragment length polymorphisms. Nature, 1988, 335: 721-726[本文引用:1][JCR: 38.597]
SubhadraS, MohapatraT, RakeshS, HussainZ. Mapping of QTLs for oil content and fatty acid composition in Indian mustard [Brassica juncea (L. ) Czern. and Coss. ]. J Plant Biochem Biot, 2013, 22: 80-89[本文引用:2][JCR: 0.414]
[27]
MahmoodT, Rahman MH, Stringam GR, YehF, Good AG. Identification of quantitative trait loci (QTL) for oil and protein contents and their relationships with other seed quality traits in Brassica juncea. Theor Appl Genet, 2006, 113: 1211-1220[本文引用:1][JCR: 3.658]
[28]
梅德圣, 张垚, 李云昌, 胡琼, 李英德, 徐育松. 油菜油分、蛋白质和硫苷含量相关性分析及 QTL定位. 植物学报, 2009, 44: 536-545Mei DS, ZhangY, Li YC, HuQ, Li YD, Xu YS. Identification of quantitative trait loci for oil, protein and glucosinolate content in Brassica napus. Chin Bull Bot, 2009, 44: 536-545 (in Chinese with English abstract)[本文引用:1]
[29]
Zhang JF, Qi CK, Pu HM, ChenS, ChenF, Gao JQ, Chen XJ, GuH, Fu SZ. QTL identification for fatty acid content in rapeseed (Brassica napus L. ). Acta Agron Sin, 2008, 34: 54-60[本文引用:2][CJCR: 1.667]
[30]
Yan XY, Li JN, WangR, Jin MY, ChenL, QianW, Wang XN, Liu LZ, Mapping of QTLs controlling content of fatty acid composition in rapeseed (Brassica napus). Genes Genom, 2011, 33: 365-371[本文引用:1][JCR: 0.497]
 , 许剑锋
, 许剑锋


{kind=link}