关键词:BnMAPK1; 甘蓝型油菜; 菌核病; 抗性; 病程相关基因 Overexpression ofBnMAPK1 Enhances Resistance toSclerotinia sclerotiorum inBrassica napus WANG Shu-Wen, LU Jun-Xing, WAN Hua-Fang, WENG Chang-Mei, WANG Zhen, LI Jia-Na, LU Kun, LIANG Ying* College of Agronomy and Biotechnology, Southwest University / Chongqing Rapeseed Engineering & Technology Research Center / Engineering Research Center of South Upland Agriculture, Ministry of Education, Chongqing 400715, China Fund: AbstractPlant MAPKs (mitogen-activated protein kinases) play an important role in defense to biotic and abiotic stresses. In our previous studies,BnMAPK1 was cloned inBrassica napus and transgenic oilseed rape plants overexpressingBnMAPK1were obtained. In this study,B. napus ZY821DH line was used as control and the corresponding transgenic plants overexpressingBnMAPK1 were used as experimental materials. The resistance toS. sclerotiorum and oxalic acid content were tested by using detached leaves inoculated with the pathogen. The dynamic changes of the relative expression of four pathogenesis-related genes,OXO,Cu/Zn-SOD,PR2, andPR3 were detected in the leaves inoculated with the pathogen. The results showed that the overexpression ofBnMAPK1 significantly inhibited the invasion of the pathogen, controlled the accumulation of oxalic acid in the inoculated leaves, and maybe finally relieved the negative regulation ofOXO expression caused by the pathogen and up-regulated the expression of the other three genes,Cu/Zn-SOD,PR2, andPR3. The results indicated that overexpression ofBnMAPK1 can effectively improve the resistance toS. sclerotiorum in oilseed rape.
Keyword:BnMAPK1; Brassica napus; Sclerotinia rot; Resistance; Pathogenesis-related genes Show Figures Show Figures
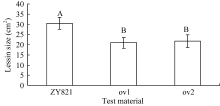
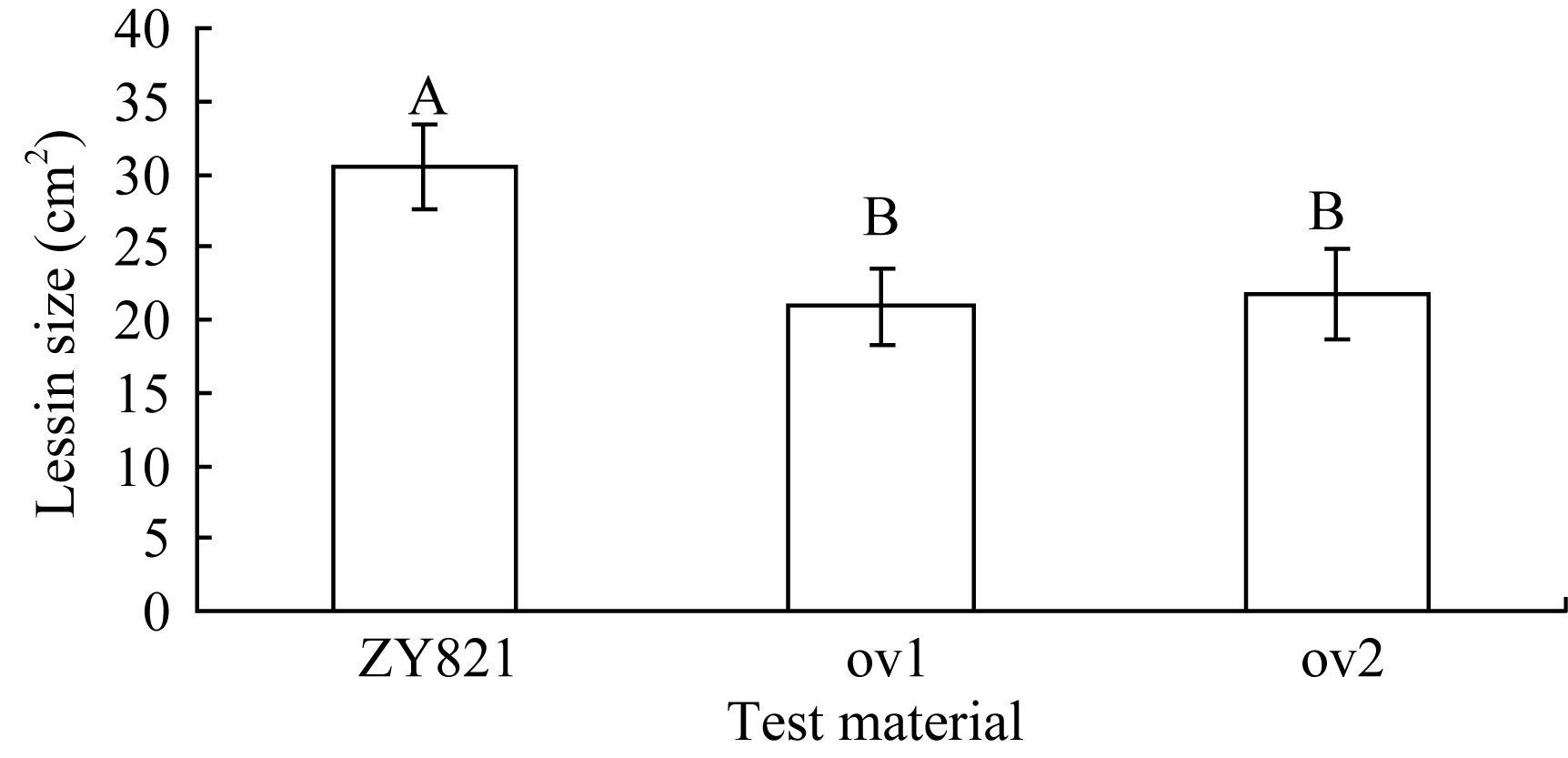
图2 离体叶片接种核盘菌36 h后病斑的差异比较标有不同字母的材料间差异极显著( P< 0.01)。Fig. 2 Comparison of lesion on the detached leaves after inoculation with S. sclerotiorum for 36 hoursBars superscripted by different letters are significantly different among the lines at P< 0.01.
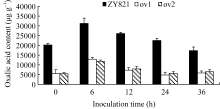
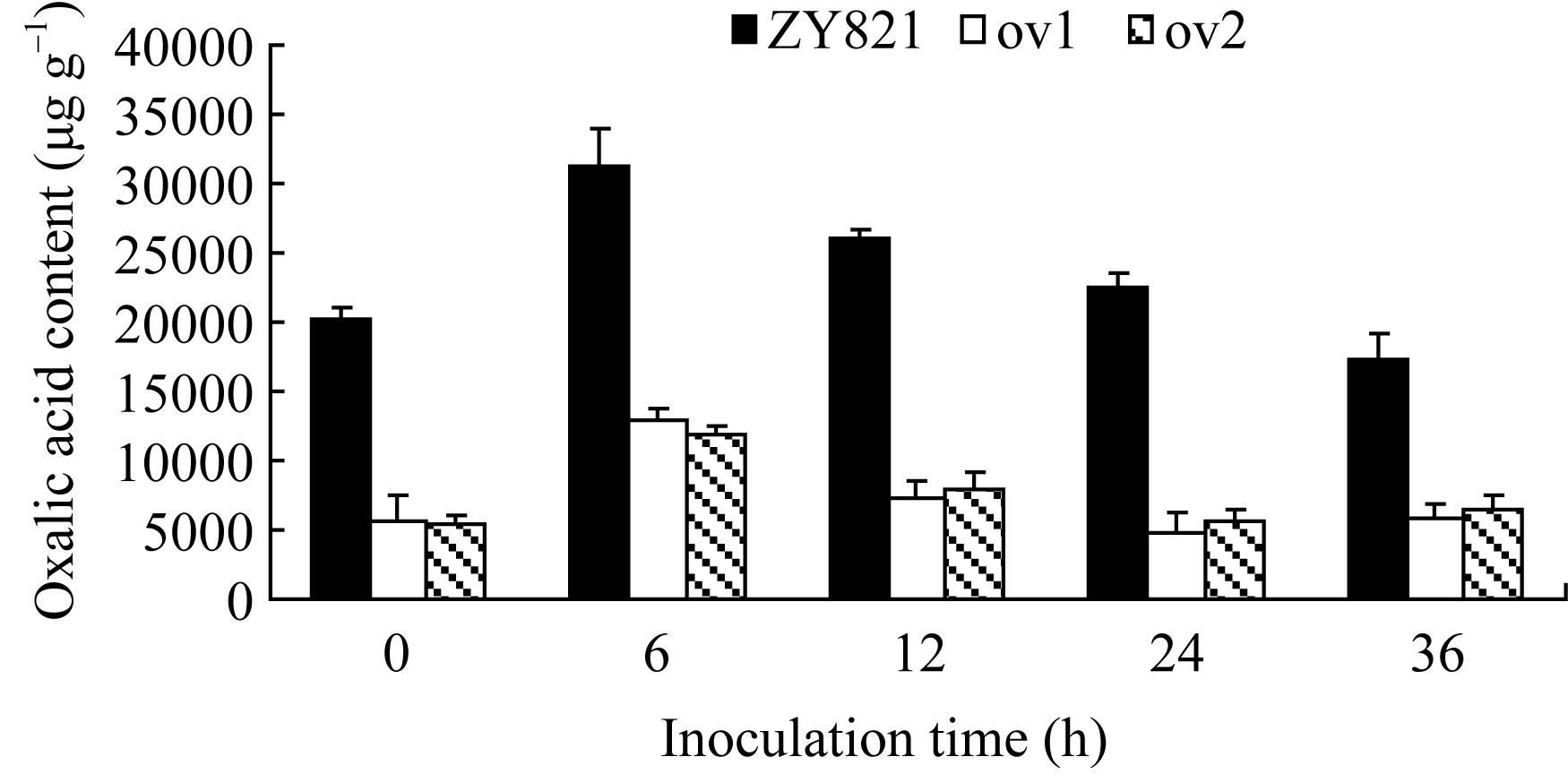
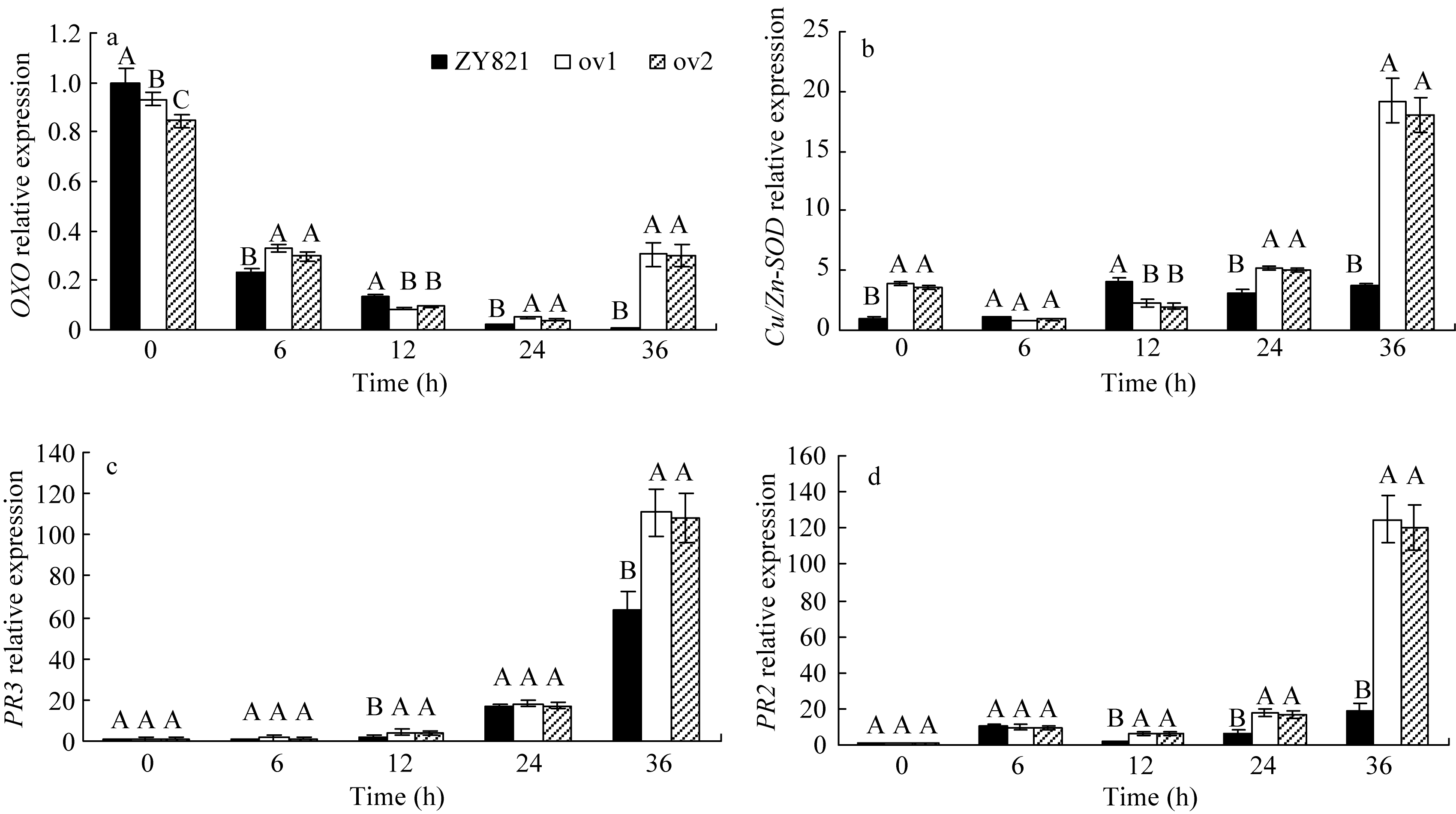
图4 接种菌核病菌的叶片中酶编码基因 OXO(a)、 Cu/Zn-SOD(b)、 PR3 (c)和 PR2(d)的相对表达标有不同字母的材料间差异极显著( P< 0.01)。Fig. 4 Relative expression of OXO(a), Cu/Zn-SOD(b), PR3(c), and PR2(d) in leaves inoculated by S. sclerotiorumBars superscripted by different letters are significantly different among the lines at P< 0.01.
表2 Table 2 表2(Table 2)
表2 酶编码基因相对表达量动态变化方差分析 Table 2 Dynamics analysis of relative expression of the genes encoding four enzymes in the leaf inoculated by S. sclerotiorum
来源 Source
自由度 df
F值 F-value
OXO
Cu/Zn-SOD
PR2
PR3
材料 Material
2
10.042**
1224.750**
2707.901**
787.454**
时间 Time
4
469.507**
4447.958**
11453.726**
35573.894**
材料×时间 Material × time
8
13.002**
878.185**
7979.590**
594.885**
表2 酶编码基因相对表达量动态变化方差分析 Table 2 Dynamics analysis of relative expression of the genes encoding four enzymes in the leaf inoculated by S. sclerotiorum
**表示在 P < 0.01水平差异极显著。**Significant difference at P < 0.01.
ColcombetJ, HirtH. Arabidopsis MAPKs: a complex signaling network involved in multiple biological processes. Biochem J, 2008, 413: 217-226[本文引用:1][JCR: 4.654]
[2]
朱斌, 梁颖. 植物MAPK C族基因的研究进展. 生物技术通报, 2012, (11): 27-28ZhuB, LiangY. Progress of study on plant’s MAPK genes of group C. Biotech Bull, 2012, (11): 27-28 (in Chinese with English abstract)[本文引用:1]
[3]
RohilaJ S, YangY N. Rice mitogen-activated protein kinase gene family and its role in biotic and abiotic stress response. J Integr Plant Biol, 2007, 49: 751-759[本文引用:1][JCR: 3.75][CJCR: 0.7555]
[4]
谷令坤. 玉米根系ZmOSMAPK1基因分离、功能鉴定及信号转导作用. 山东农业大学博士学位论文, 2006GuL K. Isolation and Function Identification of a Novel Mitogen-Activated Protein Kinase Gene, ZmOSMAPK1, in Maize Roots and Role in Plant Signal Transduction. PhD Dissertation of Shand ong Agricultural University, Tai’an, China2006 (in Chinese with English abstract)[本文引用:1]
[5]
李方球, 官春云. 油菜菌核病抗性鉴定、抗性机理及抗性遗传育种研究进展. 作物研究, 2001, 15(3): 85-92LiF Q, GuanC Y. Research progress of identification, mechanism and genetic breeding of Brassica napus against Sclerotinia sclerotiorum. Crop Res, 2001, 15(3): 85-92 (in Chinese with English abstract)[本文引用:1][CJCR: 0.7044]
[6]
万华方, 刘瑶, 梅家琴, 丁一娟, 梁颖, 曲存民, 卢坤, 李加纳, 钱伟. 人工合成高抗菌核病甘蓝型油菜几种关键酶编码基因的表达与其抗性的关系. 中国农业科学, 2012, 45: 4543-4551WanH F, LiuY, MeiJ Q, DingY J, LiangY, QuC M, LuK, LiJ N, QianW. Relationship between the expression of genes encoding resistance-related enzymes and the resistance to Sclerotinia sclerotiorumin resynthesized Brassica napus with high level of resistance. Sci Agric Sin, 2012, 45: 4543-4551 (in Chinese with English abstract)[本文引用:1][CJCR: 1.889]
[7]
MeiJ Q, QianL W, DisiJ O, YangX R, LiQ F, LiJ N, FrauenM, CaiD, QianW. Identification of resistant sources against Sclerotina sclerotiorum in Brassica species with emphasis on B. oleracea. Euphytica, 2011, 177: 393-399[本文引用:1][JCR: 1.643]
[8]
俞乐, 彭新湘, 杨崇, 刘拥海, 范燕萍. 反相高效液相色谱法测定植物组织及根分泌物中草酸. 分析化学, 2002, 30: 1119-1122YuL, PengX X, YangC, LiuY H, FanY P. Determination of oxalic acid in plant tissue and root exudate by reversed phase high performance liquid chromatography. Chin J Anal Chem, 2002, 30: 1119-1122 (in Chinese with English abstract)[本文引用:1][JCR: 0.769][CJCR: 1.29]
[9]
Shivani Y A K, SrivastavaD P, Singh D K A. Isolation of oxalic acid tolerating fungi and decipherization of its potential to control Sclerotinia sclerotiorum through oxalate oxidase like protein. World J Microbiol Biotechnol, 2012, 28: 3197-3206[本文引用:1][JCR: 1.262]
[10]
AndreasW, IrmgardZ S, SimoneT, AndreasK. Reactive oxygen intermediates and oxalic acid in the pathogenesis of the necrotrophic fungus Sclerotinia sclerotiorum. Plant Pathol, 2008, 120: 317-330[本文引用:1][JCR: 2.729]
[11]
RaiJ N, DhawanS. Studies on purification and identification of toxic metabolite produced by Sclerotinia sclerotiorum causing white rot disease of crucifers. Ind Phytopathil, 1976, 29: 407-411[本文引用:1]
[12]
刘胜毅. 草酸法筛选油菜抗菌核病材料的效果及其影响因素. 植物保护学报, 1998, 25: 56-59LiuS Y. Oxalic screening of rape resistance to the effect of the material and its influencing factors. Acta Phytophyl Sin, 1998, 25: 56-59 (in Chinese with English abstract)[本文引用:1]
[13]
吴纯仁, 刘后利. 油菜菌核病的致病机制: Ⅲ. 罹病组织内草酸毒素积累和分布的初步分析. 植物病理学报, 1991, 21: 135-140WuC R, LiuH L. Pathogenesis of Sclerotinia sclerotiorum: III. The oxalate accumulation of toxins in diseased tissues and preliminary analysis of the distribution. Acta Phytopathol Sin, 1991, 21: 135-140 (in Chinese with English abstract)[本文引用:1][CJCR: 1.088]
[14]
赵丹丹, 臧新, 田保明, 顾建伟. 菌核菌及油菜菌核病相关研究进展. 河南农业科学, 2010, (2): 120-122ZhaoD D, ZangX, TianB M, GuJ W. Research progress of Sclerotium fungus and Sclerotinia sclerotiorum. Henan Agric Sin, 2010, (2): 120-122 (in Chinese with English abstract)[本文引用:1]
[15]
LeonJ, LawtonM A, RaskinI. Hydrogen peroxide stimulates salicylic acid biosynthesis in tobacco. Plant Physiol, 1995, 108: 1673-1678[本文引用:1][JCR: 6.555]
[16]
AlvarezM E, PenndllR I, MeijerP J. Reactive oxygen intermediates mediate a systemic signal networks in the establishment of plant immunity. Cell, 1998, 92: 773-784[本文引用:1][JCR: 31.957]
[17]
ThompsonC, DunwellJ M, JohnstoneC E, LayV, RayJ, SchmittM, WatsonH, NisbetG. Degradation of oxalic acid by transgenic oilseed rape plants expressing oxalate oxidase. Euphytica, 1995, 85: 169-172[本文引用:1][JCR: 1.643]
[18]
DongX B, JiR Q, GuoX L, FosterS J, ChenH, DongC H, LiuY Y, HuQ, LiuS Y. Expressing a gene encoding wheat oxalate oxidase enhances resistance to Sclerotinia sclerotiorum in oilseed rape (Brassica napus). Planta, 2008, 288: 331-340[本文引用:1][JCR: 3.347]
[19]
ChippsT J, GilmoreB, MyersJ R, StotzH U. Relationship between oxalate, oxalate oxidase activity, oxalate sensitivity, and white mold susceptibility inPhaseolus coccineus. Phytopathology, 2005, 95: 292-299[本文引用:1]
[20]
DonaldsonP A, AndersonT, LaneB G, DavidsonA L, SimmondsD H. Soybean plants expressing an active oligomeric oxalate oxidase from the wheat gf-2. 8 (germin) gene are resistant to the oxalate-secreting pathogen Sclerotinia sclerotiorum. Physiol Mol Plant Pathol, 2001, 59: 297-307[本文引用:1][JCR: 1.506]
[21]
LivingstoneD M, HamptonJ L, PhippsP M, GrabauE A. Enhancing resistance to Sclerotinia minor in peanut by expressing a barley oxalate oxidase gene. Plant Physiol, 2005, 137: 1354-1362[本文引用:1][JCR: 6.555]
[22]
LiangY, SrivastavaS, RahamanM H, StrelkovS E, KavN N. Proteome changes in leaves of Brassica napus L. as a result of Sclerotinia sclerotiorum challenge. J Agric Food Chem, 2008, 56: 1963-1976[本文引用:1][JCR: 2.906]
[23]
齐绍武, 官春云, 刘春林. 甘蓝型油菜品系一些酶的活性与抗菌核病的关系. 作物学报, 2004, 30: 270-273QiS W, GuanC Y, LiuC L. Relationship between some enzyme activity and resistance to Sclerotinia sclerotiorum of rapeseed cultivars. Acta Agron Sin, 2004, 30: 270-273 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[24]
王雅平, 刘伊强, 施磊, 潘乃穟, 陈章良. 小麦对赤霉病抗性不同品种的SOD活性. 植物生理学报, 1993, 19: 353-358WangY P, LiuY Q, ShiL, PanN S, ChenZ L. SOD activity of wheat varieties with different resistance to scab. Acta Phytophysiol Sin, 1993, 19: 353-358 (in Chinese with English abstract)[本文引用:1]
[25]
云兴福, 李荣禧. 用抗病体子叶SOD同功酶蛋白质诱导黄瓜对霜霉病抗性的研究. 植物病理学报, 1997, 27: 221-224YunX F, LiR X. Studies of induced resistance to downy mildew of cucumber with SOD isozyme protein in cotyledons. Acta Phytopathol Sin, 1997, 27: 221-224 (in Chinese with English abstract)[本文引用:1][CJCR: 1.088]
[26]
牛立元, 王鸿升, 石明旺. 小麦叶片SOD、POD活性与白粉病抗性关系. 河南职业技术师范学院学报, 2004, 32(4): 5-8NiuL Y, WangH S, ShiM W. Changes of SOD and POD activities in wheat leaves infected by wheat powdery mildew and their relations to resistance. J Henan Voction-Technical Coll, 2004, 32(4): 5-8 (in Chinese with English abstract)[本文引用:1]
[27]
杨鸯鸯, 李云, 丁勇, 徐春雷, 张成桂, 刘英, 甘莉. 甘蓝型油菜Cu/ZnSOD和FeSOD基因的克隆及菌核病菌诱导表达. 作物学报, 2009, 35: 71-78YangY Y, LiY, DingY, XuC L, ZhangC G, LiuY, GanL. Cloning of Cu/Zn-superoxide dismutase of Brassica napus and its induced expression by Sclerotinia sclerotiorum. Acta Agron Sin, 2009, 35: 71-78 (in Chinese with English abstract)[本文引用:1][CJCR: 1.667]
[28]
EgeaC, AlcazarM D, Cand eland M E. β-1, 3-glucanase and chitinase as pathogenesis-related proteins in the defense reaction of two Capsicum annuum cultivars infected with cucumber mosaic virus. Biologia Plant, 1996, 38: 437-443[本文引用:1][JCR: 1.692]
[29]
蔡新忠. 植物病程相关蛋白. 植物生理学通讯, 1995, 31: 129-136CaiX Z. Plant pathogenesis-related proteins. Plant Physiol, 1995, 31: 129-136 (in Chinese with English abstract)[本文引用:1][JCR: 6.555]
[30]
蓝海燕. 表达β-1, 3-葡聚糖酶及几丁质酶基因的转基因烟草及其抗真菌病的研究. 遗传学报, 2000, 27: 70-77LanH Y. The study of the anti-fungal disease about the expression of β-1, 3-glucanase and chitinase genes in transgenic tobacco. Acta Genet Sin, 2000, 27: 70-77 (in Chinese with English abstract)[本文引用:1]





{kind=link}
{kind=link}
{kind=link}
{kind=link}