 , 张周翀1, 何韵1, 唐量1
, 张周翀1, 何韵1, 唐量1 , 彭霖靖1, 曹林1, 徐亚楠1, 衣学文1,2, 李小伟1, 须藤隆一3
, 彭霖靖1, 曹林1, 徐亚楠1, 衣学文1,2, 李小伟1, 须藤隆一31. 上海大学环境与化学工程学院, 上海 200444;
2. 上海市金山区生态环境局, 上海 200540;
3. 东北大学, 日本仙台 980-8579
收稿日期: 2019-06-24; 修回日期: 2019-07-24; 录用日期: 2019-07-24
基金项目: 国家自然科学基金(No.21777092)
作者简介: 丁国际(1955-), 男, 教授(博士), E-mail:gjding@shu.edu.cn
通讯作者(责任作者): 唐量(1983—), 男, 副研究员, 博士生导师, 主要研究方向为:环境科学与技术. E-mail:Tang1liang@shu.edu.cn
摘要: 蛭形轮虫常被用作生物处理性能良好的指示生物.本文以一种常见的蛭形轮虫—旋轮虫(Philodina sp.)为研究对象,着重探讨了旋轮虫及其分泌物对活性污泥处理性能和细菌的作用方式,以明确旋轮虫在污水生物处理中的作用机制.结果表明,旋轮虫及其分泌物均在一定程度上提高了活性污泥对COD、氨氮、总磷的去除率,且两者的提高效果相近,表明旋轮虫分泌物是旋轮虫对活性污泥的主要作用形式,即非直接捕食性的间接作用是其主要作用形式.旋轮虫分泌物对絮凝性细菌的絮凝性、异养硝化-好氧反硝化菌的同步硝化和脱氮效能均有促进作用,进一步表明旋轮虫提高活性污泥活性的作用是通过其分泌物提高细菌活性的结果.此外,通过使用根癌农杆菌KYC55从旋轮虫分泌物中检测出AHLs信号分子的类似物,表明旋轮虫对细菌的作用机制可能是其分泌物误导或触发细菌的群体感应所致.本文结果为研究微型动物在污水生物处理中的作用机制提供了新的研究思路.
关键词:旋轮虫分泌物污水生物处理作用群体感应
Preliminary study on the function of Philodina sp. in biological wastewater treatment
DING Guoji1
, ZHANG Zhouchong1, HE Yun1, TANG Liang1 , PENG Linjing1, CAO Lin1, XU Yanan1, YI Xuewen1,2, LI Xiaowei1, SUDO Ryuichi3 1. School of Environmental and Chemical Engineering, Shanghai University, Shanghai 200444;
2. Jinshan District Ecological Environment Bureau in Shanghai, Shanghai 200540;
3. Tohoku University, Sendai 980-8579
Received 24 June 2019; received in revised from 24 July 2019; accepted 24 July 2019
Abstract: The bdelloid rotifers are often used as indicator organisms in biological wastewater treatment. In this paper,a common bdelloid rotifer,Philodina sp.,was selected as a research object,and the effects of Philodina sp. and its secretion on the performance of activated sludge and the bacteria behavior were studied to determine the mechanism of Philodina sp. in biological wastewater treatment. The results show that both Philodina sp. and its secretion increased COD,ammonia nitrogen and total phosphorus remvoal by activated sludge,in similar pace,suggesting that the secretion of Philodina sp. is the main factor affecting the activated sludge. The Philodina sp. secretion can promote the flocculation of bacteria and the nitrogen removal by simultaneous heterotrophic nitrifying-aerobic denitrifying bacteria. In addition,the analogs of AHLs signaling molecules were detected in the secretion of Philodina sp. by using Agrobacterium tumefaciens KYC55,implying that the influence of on bacteria may be due to the secretion of Philodina sp. induces quorum sensing of bacteria. The results of this study provide a new idea for investigating the function of microfauna in biological wastewater treatment.
Keywords: Philodina sp.secretionbiological wastewater treatmentfunctionquorum sensing
1 引言(Introduction)在污水生物处理系统中, 除了细菌、真菌等微生物, 还往往存在包括原生动物和微型的后生动物在内的微型动物.有关微型动物在生物处理中的作用一直是研究的焦点.首先受到大量关注的是微型动物对水质和处理性能的指示作用, 这是因为微型动物的体型远远大于微生物, 因而可以直接在显微镜下进行观察和计数, 从而能够快速地对生物处理系统的处理水质和处理性能做出初步判断.这方面的研究内容较为丰富, 如直接调查微型动物的种类和数量与处理水质指标间的相关性, 以及研究在非正常运行状况下微型动物群落的变化特点等.例如, 陈声贵等(2003)在2001—2002年期间, 系统地研究了高碑店污水厂四系列曝气池活性污泥中微型动物种群动态与水质变化的相互关系, 发现游泳类纤毛虫总数与出水BOD5值呈正相关, 固着类纤毛虫的多样性值与出水总氮呈正相关, 爬行类纤毛虫的多样性值与出水硝态氮和出水总磷都呈正相关, 微型动物种群动态规律与污水处理效果关系密切.刘娟等(2009)系统研究了丝状菌膨胀与非丝状菌膨胀2种典型污泥状态下原生动物的群落结构特征及其演变规律, 发现伴随丝状菌的大量增殖, 原生动物总量相应减少, 匍匐型纤毛虫及有壳类肉足虫数量迅速上升, 占据明显的优势地位;非丝状菌污泥膨胀对原生动物总量及种群结构影响较小, 伴随粘性菌胶团的大量出现, 各功能类群的比例变化较小, 但原生动物总量持续增加, 其中, 菌食性纤毛虫呈线性增加.随着水体富营养化问题日渐受到重视, 有关污水硝化脱氮性能与微型动物的关系也一并受到了人们的关注.齐嵘等(2013)调查分析了人工氨氮配水处理化过程中原生动物的种群结构特征, 发现表壳虫丰度与亚硝酸盐氮浓度呈正相关, 累枝虫丰度与硝酸盐氮浓度呈线性相关;Zhou等(2008)发现, 匍匐型纤毛虫和有壳变形虫数量与污水处理厂出水氨氮和总氮显著负相关, 这两种原生动物对硝化作用具有很好的指示作用.随着现代科技的进步, 如图像识别技术等的不断发展, 对微型动物的种类鉴定和计数将变得更加简便和精确, 有关微型动物的生物指示作用的研究也有望得到更广泛的开展(孙晓霞等, 2014;梁晓彤等, 2017).
在污水生物处理生态系统中, 微型动物与微生物相比处于最高的生态位, 具有微生物所不具备的功能和作用, 会对生物处理性能产生特殊的影响.这些功能和作用包括减少剩余污泥量(Ghyoot et al., 2000;梁鹏等, 2003), 通过捕食悬浮颗粒物从而提高处理水的透明度(Lapinski et al., 2003), 促进絮凝性细菌的絮凝性(Weitere et al., 2005; Ding et al., 2017), 提高悬浮污泥的沉降性能(艾恒雨等, 2006), 促进细菌生长(Simek et al., 1997; Ding et al., 2017), 以及促进细菌活性(Gurijala et al., 1990)等.
微型动物对水质的指示作用机制可以从任一种微型动物都有适宜自己生长的环境方面予以解释, 微型动物对剩余污泥的减量作用可以在较大程度上用微型动物的直接捕食功能进行解释.但有关微型动物对微生物的絮凝性、活性等的作用机制仍然处于探讨中.很多****尝试从微型动物对细菌的捕食或捕食压角度解释这些机制.例如, Hahn等(2001)认为在水生生境中, 细菌食性的原生动物的捕食会影响细菌群落的形态结构、分类组成和生理状态, 原生动物捕食可导致细菌在群落和个体水平上的变化;在群落水平上, 捕食引起的形态变化已被观察到, 这些变化朝着变得较大抑或较小的细菌群落发展, 抑或同时朝着两个方向发展;形态学的变化伴随发生分类学上的群落结构和细菌活性的变化.在一项对新开设的污水厂开展的为期5周的检测硝化细菌的演替研究中, Pajdak-Stó等(2010)认为硝化菌种群的增大可能是由于匍匐性细菌食性的纤毛虫的捕食压力增大所致, 而强劲的捕食压对氨氮的去除率并无负面影响.不过, 上述对这些机制的解释在一定程度上仍然是一些推测, 因为仍有一系列的问题未被探明:这种捕食机制是否是由微型动物的直接捕食引起?如果是直接捕食引起的, 未被捕食的细菌是如何知道它们的同伴被捕食了从而采取对抗措施?细菌是如何感知到捕食压并采取相应的对策?已有不少研究表明, 细菌种群的絮凝特性、生长密度等表型是受细菌群体感应调控的(Fuqua et al., 1994; Schuster et al., 2013; Yang et al., 2016), 那么细菌对捕食压的感知是否可能是通过感知捕食者分泌的信息物质的途径获得?细菌感知了捕食者分泌的信息物质后的一些行为或形态的改变是否与细菌的群体感应机制有关?
蛭形轮虫(Bdelloid Rotifer)是一种在活性污泥、生物膜等基质表面上营附着生活的无性繁殖的淡水小型无脊椎动物.污水生物处理系统中常见的蛭形轮虫主要包括轮虫属(Rotaria)和旋轮虫属(Philodina)轮虫.在污水生物处理系统中, 蛭形轮虫体形较大, 常处于生态位的最高位, 而且在数量上也常成为优势种类, 被用作生物处理性能良好的指示生物(张晓宁等, 2010; Chen et al., 2012).
综上所述, 微型动物在污水生物处理中具有一些微生物没有的、特殊的有益作用, 这些作用可以分为直接作用和间接作用, 直接作用的机制可以用微型动物的直接捕食进行解释, 而非直接捕食的间接作用机制目前仍不明确.基于此, 本文以污水生物处理系统中常见的一种蛭形轮虫—旋轮虫为研究对象, 着重研究旋轮虫分泌物对活性污泥处理性能和细菌生长特性等的作用, 以明确旋轮虫的间接作用方式;此外, 对旋轮虫分泌物是否含有群体感应信号分子的类似物进行初步检测, 以期为揭示微型动物在污水生物处理中的作用机制提供新的研究思路.
2 材料与方法(Materials and methods)2.1 菌种本文用于旋轮虫分泌物对絮凝性影响实验的絮凝性细菌和用于旋轮虫分泌物对异养硝化-好氧反硝化细菌影响实验的异养硝化-好氧反硝化细菌均分离纯化自某城市污水厂的活性污泥.9株絮凝性细菌分别为Bacillus cereus PLJ01、Bacillus amyloliquefaciens PLJ08、Lysinibacillus boronitolerans PLJ09、Micrococcus luteus PLJ10、Exiguobacterium indicum PLJ13、Bacillus amyloliquefaciens PLJ17、Bacillus megaterium PLJ18、Brevundimonas vesicularis PLJ21、Bacillus amyloliquefaciens PLJ27.2株异养硝化-好氧反硝化细菌分别为Bacillus subtilis CL1和Gordonia terrae CL6.
2.2 培养基2.2.1 牛肉膏蛋白胨培养基牛肉浸膏5 g, 鱼粉蛋白胨10 g, 氯化钠5 g, 蒸馏水1 L, 调节pH为7.0~7.2.
2.2.2 发酵培养基酵母浸膏0.01 g, 葡萄糖25 g, 七水硫酸镁0.2 g, 磷酸氢二钾2 g, 磷酸二氢钾1 g, 氯化铵1 g, 蒸馏水1000 mL, 调节pH为7.0~7.2.
2.2.3 异养硝化培养基葡萄糖1.667 g, 硫酸铵0.25 g, 氯化钠0.3 g, 磷酸氢二钾1 g, 硫酸镁0.3 g, 硫酸亚铁0.03 g, 1000 mL去离子水, 用氢氧化钠及盐酸调节pH为7.0~7.2.
2.3 细菌培养将9株絮凝细菌和2株异养硝化-好氧反硝化细菌分别接种于牛肉膏蛋白胨培养基中, 30 ℃、150 r·min-1下振荡培养24 h, 然后在4 ℃、12000 r·min-1下离心20 min, 去除上清液, 沉淀用无菌水重悬, 稀释, 制备OD600为0.6~0.8的菌悬液, 备用.
2.4 旋轮虫旋轮虫(Philodina sp.)(图 1)分离自上海市某城市污水厂, 并经继代培养后供实验所用.
图 1(Fig. 1)
 |
| 图 1 旋轮虫显微镜照片 Fig. 1Microscopic image of rotifer |
2.5 旋轮虫分泌物用400目尼龙滤网过滤培养的旋轮虫, 用去离子水反复3次清洗截留在滤网上的旋轮虫, 并将滤网截留的旋轮虫倒入锥形瓶;然后用去离子水调节锥形瓶中旋轮虫密度至100 ind·mL-1, 将锥形瓶放置在30 ℃、150 r·min-1的摇床中振荡24 h;最后取出摇床中的锥形瓶, 用0.22 μm滤膜反复2次过滤锥形瓶中的溶液, 过滤后的滤液即为含有轮虫分泌物的溶液.
2.6 实验设计2.6.1 旋轮虫对活性污泥处理性能的影响设置3个SBR法实验组, 其中1组为对照组(C组)、1组为添加旋轮虫组(R组)、1组为添加旋轮虫分泌物组(RS组), 每个实验组设置2个平行.实验组均使用1 L锥形瓶作为活性污泥法反应器, 反应器中加入活性污泥100 mL和模拟废水800 mL.C组加入100 mL无菌水, R组加入100 mL由无菌水和旋轮虫(100 ind·mL-1)配制的溶液, RS组加入100 mL旋轮虫分泌物(旋轮虫100 ind·mL-1)溶液.反应器一个周期内曝气9 h、沉淀1.5 h、闲置1.5 h.按时取样测定COD、氨氮和TP浓度.
2.6.2 旋轮虫分泌物对絮凝性的影响设置2个实验组, 其中1组为对照组(C组)、1组为添加旋轮虫分泌物组(RS组), 每个实验组设置3个平行.絮凝细菌分离纯化自某城市污水厂的活性污泥.将扩大培养后的絮凝细菌菌悬液以1%的接种量接种至2倍浓度发酵培养液.对照组加50%无菌水, 实验组加50%分泌物溶液, 按时取样测定絮凝活性.
2.6.3 旋轮虫分泌物对异养硝化-好氧反硝化细菌的影响设置2个实验组, 其中1组为对照组(C组)、1组为添加旋轮虫分泌物组(RS组), 每个实验组设置3个平行.异养硝化-好氧反硝化细菌分离纯化自某城市污水厂的活性污泥.将扩大培养后的异养硝化-好氧反硝化细菌菌悬液以1%的接种量接种至2倍浓度异养硝化培养液, 对照组加50%无菌水, 实验组加50%分泌物, 按时取样测定氨氮和TN.
2.6.4 旋轮虫分泌物对细菌群体感应的影响在15 mL离心管中依次加入2 mL去离子水、2 mL过夜培养的KYC55(OD600≈0.6)、50 μL X-gal、50 μL旋轮虫分泌物, 以去离子水作对照, 28 ℃摇床培养12 h, 比较其颜色变化.
2.7 实验测定方法2.7.1 菌株生长量的测定菌株生长量用吸光度表示, 采用分光光度法测定, 波长为600 nm(沈萍等, 2007).发酵液在4 ℃、12000 r·min-1下离心20 min, 去除上清液, 沉淀用0.6%无菌生理盐水漂洗3次, 等体积重悬、混匀, 600 nm处测定吸光度.
2.7.2 絮凝活性测定方法称取0.5 g高岭土于锥形瓶中, 加入90 mL蒸馏水、5 mL的1%CaCl2溶液及5 mL各细菌的发酵液, 置于室温摇床内(200 r·min-1, 5 min), 然后取出静置5 min, 再吸取上清液, 在550 nm处测定其吸光度(以a表示).以不加细菌培养液(加5 mL新鲜培养基)为对照组(吸光度以b表示), 来确定絮凝物质的絮凝程度, 并通过如下公式计算絮凝活性(R)的大小:
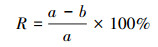 | (1) |
3 结果与讨论(Results and discussion)3.1 旋轮虫及其分泌物对活性污泥处理性能的影响为了明确旋轮虫对活性污泥处理性能的作用方式, 设置3个实验组, 分别为对照组(C组)、在对照组基础上添加旋轮虫的旋轮虫组(R组)和在对照组基础上添加旋轮虫分泌物的旋轮虫分泌物组(RS组).实验所考察的活性污泥的处理性能指标分别为COD、氨氮和总磷.
3.1.1 对COD去除性能的影响C组、R组和RS组中活性污泥在48 h内对COD的去除率如图 2a所示.在实验时间为12 h时, 3个实验组的COD去除率均超过了50%.但与C组的COD去除率相比, R组和RS组分别高出15.5%和22.5%, 均明显高于C组, 表明无论是添加旋轮虫还是添加旋轮虫分泌物均有助于活性污泥对COD的去除.
图 2(Fig. 2)
 |
| 图 2 旋轮虫及其分泌物对活性污泥COD(a)、氨氮(b)和总磷(c)去除性能的影响 Fig. 2Effects of rotifer and rotifer secretion on the removal efficiency of COD(a), ammonia nitrogen(b) and phosphorus(c) by activated sludge |
对比12 h时R组和RS组的COD去除率, 发现RS组的COD去除率明显高于R组, 这个结果提示旋轮虫有助于提高活性污泥COD处理性能的作用方式很可能主要是通过旋轮虫分泌物, 即旋轮虫的间接作用是主要的作用方式.至于R组的COD去除率低于RS组的原因可能有两种:一种可能是由于旋轮虫以细菌为食, 旋轮虫对细菌的捕食在一定程度上造成了细菌数量的减少, 从而降低了其提高活性污泥对COD处理性能的作用;二是RS组添加的旋轮虫分泌物的量不一定与R组旋轮虫实际产生的分泌物量完全相等, 即有可能略大于R组.
实验时间在24~48 h时, 在相同的实验时间内, C组、R组和RS组3组之间的COD去除率差异明显小于12 h时的结果, R组和RS组在24、36和48 h时的总平均COD去除率仅比C组分别高1.06%和3.67%, 表明添加旋轮虫及其分泌物后尽管仍有提高COD去除性能的作用, 但提高的程度有限.这或许与3个实验组的有机物去除性能均处于良好的状态有关, 即在生物处理系统性能处于良好状态时, 旋轮虫的作用有限.
3.1.2 对氨氮去除性能的影响如图 2b所示, 3个实验组中氨氮去除率的变化趋势与COD相似, 在整个实验期间, 3个实验组对氨氮的去除率随着时间的增加总体均呈现增加的趋势;在处理时间为24 h时, 3个实验组对氨氮的去除率有明显的提高, 且处于较高的水平.
在整个实验期间, R组和RS组的氨氮去除率均高于C组, 当实验时间为12 h时, R组和RS组对氨氮的去除率分别比C组高8.81%和9.61%;在实验时间24~48 h内, R组和RS组对氨氮的平均去除率分别比C组高6.63%和9.95%, 表明投加旋轮虫及其分泌物均能有效地提高活性污泥对氨氮的去除率.比较R组和RS组的氨氮去除率发现, RS组的去除性能更佳, 其原因也许与对COD的去除相似, 即R组中旋轮虫对细菌的捕食可能在一定程度上减少了细菌数量, 也可能是因为两组所含有的有效分泌物量并不等量.
3.1.3 对总磷去除性能的影响3个实验组对污水中总磷的去除效果见图 2c.在整个实验期间, 3个实验组中总磷的去除率出现了波动的状况, 这一点不同于COD和氨氮的去除率所呈现的增加趋势.不过, 比较C组、R组和RS组对总磷的去除率发现, 总体上仍均呈现RS组最高、R组次之、C组最低的趋势.表明旋轮虫及其分泌物均有助于提高活性污泥去除磷的性能.当实验时间为12 h时, R组和RS组对总磷的去除率分别比C组高0.93%和3.52%;在实验时间在24~48 h内, R组和RS组对总磷的平均去除率分别比C组高1.92%和1.46%, 表明投加旋轮虫及其分泌物对提高活性污泥的总磷去除率有一定的效果.
归纳3个实验组对COD、氨氮、总磷的去除效果, 均显示出旋轮虫及其分泌物均有助于提高活性污泥对3种主要污染物的去除性能, 提示旋轮虫对活性污泥的主要作用途径可能通过其分泌物的形式, 即旋轮虫的间接作用是主要的作用, 作用形式为分泌物.
3.2 旋轮虫分泌物对细菌的絮凝性和硝化脱氮的影响3.1节明确了旋轮虫对活性污泥的间接作用, 即通过其分泌物对活性污泥产生影响.为了进一步明确旋轮虫分泌物对活性污泥中细菌的影响, 本文考察了旋轮虫分泌物对分离自活性污泥的9株细菌的絮凝性及2株异养硝化-好氧反硝化细菌的硝化脱氮性的影响.
3.2.1 对细菌絮凝性的影响从活性污泥中分离出生长速度相对较高的9株细菌供本实验使用.将9株细菌按絮凝性的高低平均分成3组, 即絮凝性低、中、高3组, 每一组分别设置1个对照组和1个旋轮虫分泌物添加组.由图 3可见, 在细菌培养时间为36 h时, 添加旋轮虫分泌物后, 细菌的絮凝性均得到了提高, 表明旋轮虫分泌物有助于提高细菌的絮凝性.按絮凝性由低至高的顺序, RS组的絮凝性较C组分别提高11.64%、10.2%、6.27%.细菌的絮凝性在存在旋轮虫分泌物的条件下均得到了明显提高, 提高后的絮凝性大小排序与原絮凝性排序相同.不过如果将添加旋轮虫分泌物后细菌增加的絮凝性与原有的絮凝性的比值进行排序, 发现低、中、高3组的比值分别为67.2%、78.0%、85.3%, 即菌株的原有絮凝性越低时絮凝性的提高幅度越大.这表明当活性污泥的絮凝性较差时可以直接通过添加旋轮虫分泌物的方法来改善活性污泥的絮凝性.
图 3(Fig. 3)
 |
| 图 3 旋轮虫分泌物对细菌絮凝性的影响 Fig. 3Effect of secretion on bacterial flocculation |
本课题组曾探讨过旋轮虫分泌物对2株具有较高絮凝活性的细菌Brevundimonas vesicularis LW13和Bacillus cereus LW19的絮凝性的影响, 明确了旋轮虫分泌物具有提高这2株细菌絮凝性的作用(Ding et al., 2017).本文有关旋轮虫分泌物对9株细菌絮凝性影响的研究结果证实了旋轮虫分泌物的正面作用, 也再次提示旋轮虫可以通过其分泌物对细菌的絮凝性产生影响, 即不是通过直接捕食, 而是通过间接作用产生影响, 并且这种影响也许是一种常见的现象.
3.2.2 对异养硝化-好氧反硝化细菌硝化脱氮性能的影响异养硝化-好氧反硝化细菌可将水中的氨氮直接转化成氮, 从而提高脱氮反应效率;同时, 还能去除污水中的COD, 降低污水处理厂的建设运行成本.本文以2株分离自活性污泥的异养硝化-好氧反硝化细菌(Bacillus subtilis和Gordonia terrae)为研究对象, 探讨旋轮虫分泌物对该菌株的硝化和脱氮功能的影响.图 4显示了旋轮虫分泌物对菌株的氨氮和总氮去除率的影响.由图可知, 未添加分泌物时, C组中2株细菌对氨氮和总氮的平均去除率分别高达67.79%和67.30%, 且对氨氮和总氮的去除率很接近, 表明2株菌不仅具有很好的同步硝化和脱氮的性能, 且性能优异.
图 4(Fig. 4)
 |
| 图 4 分泌物对细菌去除氨氮和总氮的影响 Fig. 4Effect of secretion on biological removal of ammonia nitrogen and total nitrogen |
比较RS组与C组对氨氮和总氮的去除率结果可知, 在实验时间为48 h时, 添加旋轮虫分泌物后, 对氨氮和总氮的去除率分别提高了9.83%和9.58%, 且提高的幅度相近.此结果提示旋轮虫分泌物不仅对细菌的絮凝性有促进作用, 对异养硝化-好氧反硝化细菌的硝化和脱氮功能也有明显的促进作用, 并且再次提示旋轮虫对活性污泥净化水质性能的改善或促进作用的主要方式是通过其分泌物对活性污泥中的细菌产生影响.
3.3 旋轮虫对细菌的作用机制初探本文3.1节和3.2节的研究结果表明, 旋轮虫分泌物可以影响活性污泥的处理性能和细菌种群的行为, 即这种影响与旋轮虫的直接捕食无关, 而是通过分泌物的形式产生影响, 提示这种影响的机制可能是因为分泌物中含有某些化学信息物质.为了揭示这种可能性, 本文初步探讨了旋轮虫分泌物中是否含有群体感应信号分子或类信号分子.
群体感应是指微生物群体在其生长过程中, 由于群体密度的增加, 导致其生理和生化特性的变化.变化的原因在于当环境中微生物种群密度达到阈值, 环境中信号分子的浓度也达到一定的水平, 通过包括受体蛋白在内相关蛋白的信号传递, 诱导信号最终传递到胞内, 引起特定基因的表达, 调控微生物群体的生理特征.群体感应参与调控细菌的多种群体行为, 如EPS合成、生物发光、生物薄膜形成、抗生素生成、分泌毒性物质等(Federle et al., 2003;Okada et al., 2015).因此, 当细菌周围的环境中有类似群体感应信号分子的物质存在时或者细菌的信号分子被干扰时, 细菌生活形态将被改变.到目前为止, 有关细菌群体感应的文献主要是报道细菌种群间的群体感应, 而尚未见到有关微型动物对细菌的群体感应影响的报道.
群体感应机制中细胞间通信由信号分子介导, 如信号分子酰化高丝氨酸内酯(AHLs).本文采用根癌农杆菌KYC55检测AHLs, 由于该菌本身不产生AHLs, 当有外源AHLs信号分子或类似物存在时, KYC55表达lacZ基因生成半乳糖苷酶, 水解X\gal产生蓝色.为了检测旋轮虫分泌物中是否含有AHLs信号分子的类似物, 将分泌物的溶液加入到含有X\gal的根癌农杆菌KYC55的培养液中, 观察未加分泌物的对照组(C组)和添加了旋轮虫分泌物的分泌物组(RS组)培养液中颜色的变化情况.
如图 5所示, 添加旋轮虫分泌物的RS组的培养液变成了蓝色, 而对照组仍是原来的颜色, 提示旋轮虫分泌物中存在AHLs信号分子的类似物, 由此可能误导或触发细菌的群体感应, 使得细菌种群的行为发生变化.本文从研究旋轮虫分泌物是否含有群体感应信号分子的类似物的角度初步探讨了旋轮虫对细菌的间接作用机制, 为了进一步明确这种机制, 后续仍需进行深入研究.
图 5(Fig. 5)
 |
| 图 5 轮虫分泌物AHL类物质检测 Fig. 5Detection of AHL substances in rotifer secretion |
4 结论(Conclusions)1) 在实验时间为12 h时, 旋轮虫及其分泌物的投加使活性污泥对COD的去除率分别提高了15.5%和22.5%, 对氨氮的去除率分别提高了8.81%和9.61%, 对总磷的去除率分别提高了0.93%和3.52%, 显示了旋轮虫和旋轮虫分泌物均在一定程度上具有提高活性污泥处理性能的作用;并且由于旋轮虫及其分泌物的作用效果相近, 提示旋轮虫对活性污泥的主要作用方式可能是其分泌物, 因此, 旋轮虫对活性污泥的主要作用机制可能是非捕食性的间接作用机制.
2) 当实验时间为36 h时, 按絮凝性由低至高的顺序, 轮虫分泌物的添加使细菌的絮凝性分别提高了11.64%、10.2%、6.27%, 因此, 轮虫分泌物具有提高细菌絮凝性的作用, 且作用程度大小与菌株的原有絮凝性呈现反比的趋势.此结果提示当活性污泥的絮凝性较差时可以通过直接添加旋轮虫分泌物的方法来改善活性污泥的絮凝性.
3) 添加旋轮虫分泌物后, 在实验时间为48 h时异养硝化-好氧反硝化细菌对培养液中氨氮和总氮的去除率分别提高了9.83%和9.58%, 且提高的幅度相近.提示旋轮虫分泌物不仅对细菌的絮凝性有促进作用, 对异养硝化-好氧反硝化细菌的硝化和脱氮功能也有明显的促进作用.
4) 添加旋轮虫分泌物的根癌农杆菌KYC55的培养液变成蓝色, 而对照组仍是原来的颜色, 提示旋轮虫的作用机制在于旋轮虫分泌物中可能存在AHLs信号分子的类似物, 由此误导或触发细菌的群体感应, 使得细菌种群的行为发生变化.
参考文献
| 艾恒雨, 汪群慧, 谢维民. 2006. 生物捕食污泥减量化系统的污泥特性[J]. 给水排水, 2006, 32(s1): 72–75. |
| Chen L J, Liu Q, Peng Z, et al. 2012. Rotifer community structure and assessment of water quality in Yangcheng Lake[J]. Chinese Journal of Oceanology and Limnology, 30(1): 47–58.DOI:10.1007/s00343-012-0150-y |
| 陈声贵, 许木启, 曹宏, 等. 2003. 活性污泥微型动物种群动态与水质净化效能的关系[J]. 动物学报, 2003, 49(6): 775–786. |
| Ding G, Li X, Lin W, et al. 2017. Enhanced flocculation of two bioflocculation-producing bacteria by secretion of Philodina erythrophthalma[J]. Water Research, 112: 208–216.DOI:10.1016/j.watres.2017.01.044 |
| Federle M J, Bassler B L. 2003. Interspecies communication in bacteria[J]. The Journal of Clinical Investigation, 112(9): 1291–1299.DOI:10.1172/JCI20195 |
| Fuqua W C, Winans S C. 1994. A LuxR-LuxI type regulatory system activates Agrobacterium Ti plasmid conjugal transfer in the presence of a plant tumor metabolite[J]. Journal of Bacteriology, 176: 2796–2806.DOI:10.1128/jb.176.10.2796-2806.1994 |
| Ghyoot W, Verstraete W. 2000. Reduced sludge production in a two-stage membrane-assisted bioreactor[J]. Water Research, 34: 205–215.DOI:10.1016/S0043-1354(99)00138-4 |
| 国家环境保护总局. 2002. 水和废水监测分析方法(第四版, 增补版)[M]. 北京: 中国环境出版社: 1–826. |
| Gurijala K R, Alexander M. 1990. Effect of growth rate and hydrophobicity on bacteria surviving protozoan grazing[J]. Applied and Environmental Microbiology, 56(6): 1631–1635. |
| Hahn M W, Hofle M G. 2001. Grazing of protozoa and its effect on populations of aquatic bacteria[J]. FEMS Microbiology Ecology, 35: 113–121.DOI:10.1111/j.1574-6941.2001.tb00794.x |
| 梁晓彤, 徐践. 2017. MATLAB图像处理技术在农业病虫害识别中的应用分析[J]. 南方农业, 2017, 11(21): 117–122. |
| Lapinski J, Tunnacliffe A. 2003. Reduction of suspended biomass in municipal wastewater using bdelloid rotifers[J]. Water Research, 37(9): 2027–2034.DOI:10.1016/S0043-1354(02)00626-7 |
| 梁鹏, 黄霞, 钱易, 等. 2003. 污泥减量化技术的研究进展[J]. 环境污染治理技术与设备, 2003, 4(1): 44–52. |
| 刘娟, 杨敏, 齐嵘, 等. 2009. 污泥膨胀状态下原生动物群落结构分析[J]. 环境工程学报, 2009, 3(2): 229–233. |
| Okada M, Nakamura Y, Hayashi S, et al. 2015. Chemical structure and biological activity of a quorum sensing pheromone from Bacillus subtilis subsp.natto[J]. Bioorganic & Medicinal Chemistry Letters, 25(19): 4293–4296. |
| Pajdak-Stós A, Fia?kowska E, Fyda J, et al. 2010. Resistance of nitrifiers inhabiting activated sludge to ciliate grazing[J]. Water Science and Technology, 61(3): 573–580.DOI:10.2166/wst.2010.868 |
| 齐嵘, 刘娟, 杨敏, 等. 2013. 无机废水硝化过程中原生动物群落结构特征及其变化规律[J]. 环境科学学报, 2013, 33(1): 53–57. |
| 沈萍, 陈向东. 2007. 微生物学实验(第4版)[M]. 北京: 高等教育出版社: 1–275. |
| 孙晓霞, 孙松. 2014. 海洋浮游生物图像观测技术及其应用[J]. 地球科学进展, 2014, 29(6): 748–755. |
| Schuster M, Sexton D J, Diggle S P, et al. 2013. Acyl-homoserine lactone quorum sensing:from evolution to application[J]. Annual Review of Microbiology, 67(1): 43–63.DOI:10.1146/annurev-micro-092412-155635 |
| Simek K, Vrba J, Pernthaler J, et al. 1997. Morphological and compositional shifts in an experimental bacterial community influenced by protists with contrasting feeding modes[J]. Applied and Environmental Microbiology, 63(2): 587–595. |
| Weitere M, Bergfeld T, Rice S A, et al. 2005. Grazing resistance of Pseudomonas aeruginosa biofilms depends on type of protective mechanism, developmental stage and protozoan feeding mode[J]. Environmental Microbiology, 7(10): 1593–1601.DOI:10.1111/j.1462-2920.2005.00851.x |
| Wu C Y, Ushiwaka S, Horii H, et al. 2006. Boosting nitrification by membrane-attached biofilm[J]. Water Science and Technology, 54(9): 121–128.DOI:10.2166/wst.2006.864 |
| Yang J, Wu D, Li A, et al. 2016. The addition of N-hexanoyl-homoserine lactone to improve the microbial flocculant production of Agrobacterium tumefaciens strain F2, an exopolysaccharide bioflocculant-producing bacterium[J]. Applied Biochemistry and Biotechnology, 179(5): 728–739.DOI:10.1007/s12010-016-2027-6 |
| Zhou K, Xu M, Liu B, et al. 2008. Characteristics of microfauna and their relationships with the performance of an activated sludge plant in China[J]. Journal of Environmental Sciences, 20(4): 482–486.DOI:10.1016/S1001-0742(08)62083-5 |
| 张晓宁. 2010. 原后生动物在生活污水处理中的指示作用[J]. 广东化工, 2010, 37(11): 119–120.DOI:10.3969/j.issn.1007-1865.2010.11.060 |
