 , 汪涛1,3
, 汪涛1,3 , 任晓1,2,3, 邵志江1,2,3, 郑斌1,2,3
, 任晓1,2,3, 邵志江1,2,3, 郑斌1,2,31. 中国科学院、水利部成都山地灾害与环境研究所, 成都 610041;
2. 中国科学院大学, 北京 100049;
3. 中国科学院山地表生过程与生态调控重点实验室, 成都 610041
收稿日期: 2018-11-17; 修回日期: 2018-12-21; 录用日期: 2018-12-21
基金项目: 国家重点研发计划(No.2017YFD0800105);国家水污染治理与控制专项(No.2017ZX07101-001);国家自然科学基金项目(No.41371462)
作者简介: 刘莲(1994-), 女, E-mail:1181859683@qq.com
通讯作者(责任作者): 汪涛, E-mail:wangt@imde.ac.cn
摘要: 植物是影响沉积物反硝化作用的重要因素之一,国内外已有不少研究探讨了植物对河口或湖泊沉积物反硝化速率的影响,但关于植物类型对自然沟渠沉积物反硝化速率及其相应功能基因的影响研究不多.因此,本文以7种常见沟渠植物为研究对象,通过室内盆栽试验,利用改进的乙炔抑制法和实时荧光定量PCR技术研究了不同植物对自然沟渠沉积物反硝化速率和相应功能基因(nirS和nirK)拷贝数的影响.结果表明,培养至第180 d时,不同植物生长条件下沉积物反硝化速率在2.85~13.20 μg·m-2·h-1之间,不同植物间反硝化速率差异显著,且大型挺水植物>浮水植物>小型挺水植物.不同植物之间沉积物中nirS基因拷贝数在2.70×108~5.02×108 copies·g-1之间,nirK基因拷贝数在3.97×105~6.91×105 copies·g-1之间,与培养初期相比,培养180 d后沉积物中nirK、nirS基因拷贝数明显增多.7种植物中,美人蕉沉积物中的反硝化功能基因拷贝数较高,狐尾藻较低,整体来看,基因拷贝数大小顺序为:挺水植物>浮水植物.nirS基因拷贝数与NO3--N含量、TN含量及反硝化速率之间均呈显著性的正相关关系(p < 0.05),但nirK基因拷贝数与沉积物碳氮含量及反硝化速率之间相关性不显著.
关键词:植物类型沟渠沉积物反硝化速率nirKnirS
Effects of plant species on denitrification and functional genes in ditch sediment
LIU Lian1,2,3
, WANG Tao1,3 , REN Xiao1,2,3, SHAO Zhijiang1,2,3, ZHENG Bin1,2,3 1. Institute of Mountain Hazards and Environment, Chinese Academy of Science, Chengdu 610041;
2. University of Chinese Academy of Sciences, Beijing 100049;
3. Key Laboratory of Mountain Environment Evolvement and Regulation, Chengdu 610041
Received 17 November 2018; received in revised from 21 December 2018; accepted 21 December 2018
Abstract: The influence of plant on the denitrification of sediment in the estuary or lake have been reported by many studies. However, the effects of plant species on the denitrification of sediment and function genes in the natural ditches is still not clear. Here, we explored the effects of seven ditch plants on denitrification and functional genes (nirS, nirK) in ditch sediment via pot experiments by using the chloramphenicol-amended acetylene inhibition procedure and real time-PCR. The results show that the denitrification rates of the sediment under different plant species ranged from 2.85 μg·m-2·h-1 to 13.2 μg·m-2·h-1 after 180 days. The denitrification rates were as follows:Macrophytes > floating plants > small emerged plants, and there was a significant difference among different plant species. The gene abundances of nirS ranged from 2.70×108 copies·g-1 to 5.02×108 copies·g-1, and the gene abundances of nirK ranged 3.97×105 copies·g-1 to 6.91×105 copies·g-1. Furthermore, the gene abandunces of nirK and nirS increased significantly after 180 days of incubation. The denitrification functional gene copies was the highesin the sediment of Canna indica, while the lowest in Myriophyllum spicatum. The gene abundances of emerged plants were higher than that of floating plants. Moreover, pearson correlation analysis showed that there was a significant positive correlation between nirS and NO3--N content, TN content, and denitrification rate (p < 0.05), but the correlation between nirK and sediment physicochemical properties, denitrification rate were not significant.
Keywords: plant speciesditch sedimentdenitrificationnirKnirS
1 引言(Introduction)氮素不仅是植物生长必需的营养元素, 也是水体富营养化的主要元素(武姣娜等, 2018).沟渠作为连接农田与江河湖泊的小型湿地单元, 因其具有消减氮素污染物浓度、净化水质的功效, 受到了越来越多国内外****的广泛关注(Mulholland et al., 2008; Canfield et al., 2010; Pierobon et al., 2013).氮素在沟渠中的去除方式主要包括沉积物的吸附作用、微生物的硝化和反硝化脱氮作用和植物的吸收作用等(Senzia et al., 2003; 徐红灯等, 2007).这些去除方式中, 反硝化脱氮作用最终将水体中的氮素以气体形式排出, 是水体氮素去除最彻底的途径(Seitzinger et al., 2006).沉积物反硝化作用的影响因素众多, 譬如C/N、植物、温度等(Annelies et al., 2011; McCracki et al., 2012; 李筱宛等, 2016).植被是影响反硝化速率最重要的因素之一, 它不仅可为反硝化作用提供场所, 而且可为反硝化微生物提供合适的碳源(Kalscheur et al., 2012; Zulkifly et al., 2012).
反硝化作用是一种微生物异化氧化还原过程, 该过程发生的每一步都需要特异的酶参与完成, 其中亚硝酸还原酶参与的反应是反硝化过程的关键步骤(贺纪正等, 2013).已有研究表明, 通过编码反硝化功能基因可以更好地了解微生物对反硝化过程的调控机制(郭丽芸等, 2011)因此, 应用微生物分子生态学技术将编码亚硝酸还原酶的功能基因nirS和nirK作为靶标基因用来研究淡水沉积物等环境中反硝化细菌群落结构和多样性也成为了沉积物反硝化作用研究的热点(Braker et al., 2000; Heylen et al., 2006).尽管如此, 目前大多研究主要集中于江河湖口地区(Francis et al., 2013; Dang et al., 2008; Zheng et al., 2015), 而关于沟渠沉积物反硝化功能基因的研究鲜有报道.此外, 关于反硝化功能基因与环境因子的关系方面的研究也有很多, 分别研究了有机质(谢佳等, 2017)、施肥(郑燕等, 2012; 陈斌泽, 2013; 曾希柏等, 2014)和生物炭(刘杏认等, 2018)等因子对反硝化功能基因的影响, 但尚未有植物类型与沟渠沉积物反硝化功能基因的相关研究报道.
川中丘陵区位于长江上游腹地, 地势起伏.由于降雨径流的长期冲刷, 形成了众多的排水沟渠, 这些沟渠大多具有高差跌落、小型积水的特点, 加上川中丘陵区属亚热带季风性湿润气候, 雨量丰沛、气温适宜的气候条件十分适宜植物的生长, 导致沟渠内水生植物生长旺盛.植物的旺盛生长可能有利于促进沟渠沉积物的反硝化脱氮作用.已有不少研究探讨了不同沟渠植物对氮素的富集率与净化效率(任光前等, 2016; Mathieu et al., 2017), 并筛选出了适应川中丘陵区自然沟渠的高富集氮磷植物; 也有研究报道了有无植被对川中丘陵区自然沟渠沉积物反硝化速率的影响(龙虹竹等, 2016).但植物类型对沟渠沉积物反硝化作用效率及其相应的功能基因的影响尚不清楚.因此, 本文拟将7种生态沟渠常见水生植物作为研究对象, 探讨不同植物对沟渠沉积物反硝化速率及相应功能基因的影响, 为长江上游农田面源污染生态沟渠治理技术提供一定的科学依据.
2 材料与方法(Materials and methods)2.1 研究区概况本研究依托中国科学院盐亭紫色土农业生态试验站(31°16′N, 105°27′E).该站位于绵阳市盐亭县林山乡, 地处涪江支流弥江、湍江的分水岭上, 为丘陵地带, 该区域属亚热带湿润季风性气候, 无霜期294 d, 平均气温17.3 ℃, 多年平均降雨量825 mm左右, 年内降雨分配不均, 夏季多雨, 冬季少雨.川中丘陵区排水沟渠众多, 沟渠内植物生长茂盛.
2.2 试验设计与样品采集本研究主要通过室内盆栽试验进行, 采用随机试验设计的方法, 选取了7种生态沟渠常见植物作为研究对象, 分别是浮水植物:狐尾藻(Myriophyllum L)、铜钱草(Hydrocotyle chinensis Craib), 挺水植物:再力花(Thalia dealbata Fraser)、美人蕉(Canna indica L)、水蜡烛(Dysophylla yatabeana Makino)、茭白(Zizania latifolia Stapf)和水竹(Phyllostachys heteroclada Oliver), 同时设置了无植物对照处理, 每种处理3次重复.盆栽试验所用培养容器为高40 cm, 半径30 cm的圆柱体聚乙烯塑料桶, 在盆栽桶中加入约20 cm厚的沟渠沉积物, 约占盆栽桶体积的1/2, 所需沉积物均取自农田源头沟渠.沉积物pH值为7.73, NH4+-N含量为5.79 mg·kg-1, NO3--N含量为3.20 mg·kg-1, DOC含量为153.15 mg·kg-1, 有机质含量为22.6 g·kg-1, 有机碳含量为13.1 g·kg-1, 全氮含量为1.52 g·kg-1.
2017年3月将植株幼苗移栽至盆栽中, 为防止试验过程中出现死苗而导致试验失败的情况, 每盆移栽5株生长状况较好的植物幼苗, 且盆栽种植密度要稍大于田间条件下的种植密度, 以提高植物成活率.一周后在每个培养盆中随机选取3~4点, 利用土钻垂直采取0~20 cm深度的沉积物, 将每个处理中的3个重复的沉积物样合并为一个沉积物样品, 测定其中的nirK和nirS基因的拷贝数.待植株正常生长180 d后, 2017年9月再次采集沉积物样品, 每份样品分为两部分, 一部分密封于5 mL灭菌离心管中, 存入液氮罐中运回实验室, 利用实时荧光定量PCR技术(Real-time PCR)检测沉积物中反硝化功能基因nirS和nirK基因的表达量, 另一部分装入聚乙烯袋中密封带回实验室储于4 ℃以下的冰箱中, 用于反硝化速率及沉积物碳氮含量的测定.
2.3 沉积物碳氮含量的测定沉积物含水率的测定方法是称取一定重量的新鲜沉积物样品于105 ℃的烘箱中烘干直至恒重, 损失的水重量与烘干沉积物重量的比值即为沉积物的含水率.沉积物NH4+-N和NO3--N含量的具体测定方法如下(鲁如坤, 1999):称取5 g新鲜沉积物样品于离心管中, 加入25 mL 1 mol·L-1 KCl用于浸提沉积物NH4+-N和NO3--N, 拧紧离心管盖子摇匀沉积物样品, 放入振荡机里以25 ℃、280 r·min-1的条件振荡2 h, 待振荡结束后, 取出样品后利用0.45 μm的微孔滤膜过滤沉积物溶液, 再利用德国生产的连续流动分析仪(AA3, SEAL, Germany)测定滤液中的NH4+-N和NO3--N浓度(mg·L-1), 所得水样数据根据如下公式换算得到沉积物NH4+-N和NO3--N含量(mg·kg-1).
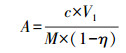 | (1) |
沉积物全氮(TN)含量利用元素分析仪(Elementar Vario MACRO)测定.有机碳(TOC)含量的测定则是先采用烧失量法测定出有机质含量, 再采用转换系数0.58将其换算成有机碳(Page et al., 1982).碳氮比(C/N)用TOC与TN的比值表示.
2.4 反硝化速率的测定反硝化速率采用改进的乙炔抑制法测定, 该方法是在传统的乙炔抑制法的基础上, 通过添加氯霉素来抑制反硝化酶的重新合成(Roley et al., 2012).具体实验方法为:称取(30±0.5) g沉积物于250 mL培养瓶中, 再加入对应的沉积物上覆水样75 mL, 氯霉素0.12 g, 并塞紧橡胶塞以防漏气.将培养瓶连接到真空泵和充氦气的装置上, 从采气口将培养瓶抽成真空, 注入1.01×105 Pa的高纯度氦气冲洗5 min, 以排除培养瓶中的空气, 同时使每个培养瓶内的压力都维持在大气压水平, 最后再添加10 mL乙炔气体, 实现8%~10%的C2H2顶空浓度.前期工作准备就绪后, 将培养瓶放入恒温培养箱中以采集沉积物时的现场采样温度培养4 h, 分别在0、2、4 h抽取10 mL气体, 利用Agilent7890A(USA)和Agilent7890B(USA)气相色谱仪对N2O进行分析.反硝化速率的计算公式如下:
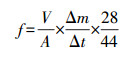 | (2) |
2.5 反硝化功能基因拷贝数的测定2.5.1 沉积物DNA的提取沉积物DNA的提取采用Fast DNA SPIN Kit For Soil(aidlab DN27, China)的试剂盒方法, 每个沉积物样品分别做3次重复, 每份样品0.5 g, 严格按照试剂盒的试验步骤进行.将提取的DNA采用SmartSpec Plus spectrophotometer(ScanDrop100, Germany)进行纯化, 最后将DNA样品保存于-20 ℃冰箱中以备用.
2.5.2 荧光实时定量PCR(Real-time PCR)nirK基因和nirS基因扩增引物序列及扩增片段大小见表 1.PCR反应体系(50 μL):AceQ? qPCR SYBR? Green Master Mix 10 μL, 10 μmol·L-1正向反向引物各0.40 μL, 1 μL DNA模板, 灭菌的ddH2O充足.反应程序为降落式(touchdown)PCR:95 ℃变性5 min, 95 ℃ 15 s, 67 ℃退火30 s(每个循环减少1 ℃, 直到58 ℃), 72 ℃延伸20 s, 10个循环; 然后95 ℃15 s, 57 ℃ 30 s, 72 ℃ 20 s, 35个循环, 最后72 ℃延伸7 min.
表 1(Table 1)
| 表 1 PCR引物 Table 1 Primers used in this study | ||||||||||||||||||
表 1 PCR引物 Table 1 Primers used in this study
| ||||||||||||||||||
扩增完成后, 分别以nirS基因的重组质粒DNA和nirK基因的重组质粒DNA为标准品, 分别作出各基因的标准曲线, 通过测定出待测样品的阈值循环数, 从标准曲线上计算出该样品的起始浓度, 实现对样本基因nirS和nirK拷贝数的绝对定量.每个反应3个重复, 平均拷贝数转换为每克干土样中的基因拷贝数.拷贝数(X0)的计算方法如下:
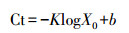 | (3) |
2.6 数据分析试验数据的统计分析采用SPSS21.0和Excel(2010), 采用LSD法比较不同处理间的差异, 采用Sigmaplot10.0作图.利用Pearson相关系数检验沉积物碳氮含量、反硝化速率分别与反硝化功能基因(nirS、nirK)拷贝数之间的相关性及显著性水平, p < 0.05表示差异性显著, p < 0.01表示差异性极显著, p > 0.05表示差异性不显著.
3 结果(Results)3.1 不同植物周边沉积物的碳氮含量分析沟渠沉积物的碳氮含量因植物种类的不同而有所差异.由表 2可见, 培养至180 d时, 沉积物中的NH4+-N、NO3--N、TN、TOC及C/N的均值分别为1.84~18.00 mg·kg-1、1.22~7.99 mg·kg-1、1.55~2.35 g·kg-1、9.86~18.47 g·kg-1和5.48~8.10.不同植物之间沉积物NH4+-N含量差异显著(表 2).其中, 沉积物NH4+-N含量大小顺序:狐尾藻 > 水蜡烛 > 水竹 > 铜钱草 > 再力花 > 美人蕉 > 茭白 > 空白.沉积物NO3--N含量:空白 > 美人蕉 > 狐尾藻 > 再力花 > 水蜡烛 > 水竹 > 茭白 > 铜钱草.整体上来说, 植物根系周围沉积物中的NH4+-N含量和NO3--N含量有着相反的关系, NH4+-N含量较高的植物, NO3--N含量便会表现出较低的趋势.不同植物之间沉积物TOC差异较为显著, 但TN和C/N差异不大.
表 2(Table 2)
| 表 2 不同植物沟渠沉积物的碳氮含量 Table 2 Carbon and nitrogen contents in ditch sediments with different plants | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
表 2 不同植物沟渠沉积物的碳氮含量 Table 2 Carbon and nitrogen contents in ditch sediments with different plants
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
3.2 不同植物周边沉积物的反硝化速率分析比较培养一周后和180 d后不同植物的沉积物反硝化速率变化发现(图 1), 总体上, 不同植物的沉积物反硝化速率在1.85~13.20 μg·m-2·h-1之间, 平均值为(5.28±1.02) μg·m-2·h-1.培养一周后, 不同植物的沉积物反硝化速率在1.85~5.90 μg·m-2·h-1之间, 平均值为(3.76±0.95) μg·m-2·h-1.培养至180 d时, 不同植物的沉积物反硝化速率在2.85~13.20 μg·m-2·h-1之间, 平均值为(6.33±0.28) μg·m-2·h-1.从图 1可以看出, 随着小型湿地系统的逐步稳定, 沉积物的反硝化速率逐渐增强.此外, 不同植物的沉积物反硝化速率大小关系表现为:美人蕉 > 铜钱草 > 狐尾藻 > 再力花 > 水竹 > 茭白 > 水蜡烛 > 空白, 整体上来看:大型挺水植物 > 浮水植物 > 小型挺水植物 > 无植物, 且它们之间有显著性的差异(p < 0.05).
图 1(Fig. 1)
 |
| 图 1 不同植物的沉积物反硝化速率 Fig. 1Denitrification rates of sediments wiht different plants |
3.3 反硝化功能基因拷贝数3.3.1 nirS基因拷贝数的变化培养一周后和180 d后采集的所有沉积物样品中均检测到了nirS基因的存在, 得到不同植物的沉积物nirS基因拷贝数在2.62×104~5.02×108 copies·g-1之间(图 2a).培养一周后, nirS基因拷贝数整体较低, 在2.62×104~1.18×107 copies·g-1范围内, 不同植物间nirS拷贝数大小顺序为:水蜡烛 > 茭白 > 空白 > 美人蕉 > 再力花 > 铜钱草 > 狐尾藻 > 水竹.培养至180 d时, 各植物周围沉积物的nirS基因拷贝数在2.70×108~5.02×108 copies·g-1之间, 从图中可以看出, nirS基因拷贝数大小顺序为:空白 > 水蜡烛 > 再力花 > 美人蕉 > 水竹>茭白 > 铜钱草 > 狐尾藻, 大体上表现为挺水植物 > 浮水植物.方差分析表明, 同一类型不同种类的植物沉积物nirS基因拷贝数之间有显著性的差异(p < 0.05).与培养一周后测定的沟渠沉积物nirS基因拷贝数相比, 所有植物的沟渠沉积物nirS基因拷贝数均有所增加, 其中水竹增加的幅度最大, nirS基因拷贝数增加了约3.55×108 copies·g-1, 茭白nirS基因拷贝数增加的幅度最小.
图 2(Fig. 2)
 |
| 图 2 不同植物沉积物的nirS(a)及nirK(b)基因拷贝数变化 Fig. 2Changes in abundance of nirS(a) and nirK(b) genes in sediments of different plants |
3.3.2 nirK基因拷贝数的变化培养一周后和180 d后采集的所有沟渠沉积物样品中均检测到了nirK基因的存在, nirK基因拷贝数在2.14×104~6.91×105 copies·g-1范围内.培养一周后, nirK基因拷贝数在2.14×104~1.09×105 copies·g-1范围内.方差分析结果表明, 不同植物间的nirK基因拷贝数差异性不显著.培养至180 d时, nirK基因拷贝数在3.97×105~6.91×105 copies·g-1范围内, 不同植物的沟渠沉积物nirK基因拷贝数之间的大小关系为:茭白 > 水蜡烛 > 美人蕉 > 空白 > 狐尾藻 > 水竹>再力花 > 铜钱草(图 2b), 大体上有挺水植物 > 浮水植物的关系.方差分析表明, 铜钱草与水竹、再力花除外的几种植物间有显著性的差异(p < 0.05).与培养一周后的测定结果相比, 所有植物的沟渠沉积物nirK基因拷贝数均增加了一个数量级, 美人蕉增加的幅度最大, 再力花沉积物中nirK基因拷贝数增加幅度最小.
4 讨论(Discussions)4.1 与其它类型水体同类研究的比较反硝化功能基因可以直观地反映出反硝化微生物的变化, 从而进一步了解反硝化作用的发生过程.目前已有很多****对不同水体类型中沉积物反硝化功能基因拷贝数进行了相关研究(Guan et al., 2014; Yin et al., 2017).李凤娥(2017)在黄河入海口检测出沉积物中nirS基因拷贝数在4.34×104~2.62×105 copies·g-1之间, nirK基因拷贝数在4.17×103~9.72×105 copies·g-1之间.程建华等(2016)在铜陵市河流沉积物中检测出nirS基因拷贝数在1.69×107~8.55×109 copies·g-1之间, nirK基因拷贝数在4.45×106~1.51×108 copies·g-1之间.本研究测得沟渠沉积物中nirS基因拷贝数在3.36×102~5.45×108 copies·g-1之间, nirK基因拷贝数在2.14×104~7.56×105 copies·g-1之间.与黄河入海口相比, 本文的反硝化功能基因拷贝数较高, 这可能与水体中的溶解氧含量有关.有研究表明, 在高浓度的溶解氧条件下, 反硝化细菌的脱氮能力会受到抑制, 从而会对其反硝化细菌拷贝数造成影响(Lefebvre et al., 2006).本研究的溶解氧平均含量(5.81 mg·L-1)低于黄河入海口的溶解氧平均含量(7.04 mg·L-1), 因而nirS和nirK基因拷贝数较黄河入海口高.与程建华等在铜陵市河流中得出的研究结果相比, 本文的结果较低.程建华等的研究结果指出以重金属为主要污染物的河流沉积物nirS基因拷贝数显著高于受氮、磷污染的河流, 说明反硝化功能基因拷贝数与水体污染物的类型也有一定关系.本文以沟渠为研究对象, 较河流、海水等水体而言, 沟渠毗邻农田及村镇生活区域, 水体污染物主要来源于农田径流、灌溉水及高氮磷含量的生活废水.加上川中丘陵区地势起伏, 区域内沟渠高差跌落, 具有明显的干湿交替过程(汪涛等, 2016; 龙虹竹等, 2016).因此, 在各种因素的综合影响下, 沟渠沉积物的反硝化功能基因拷贝数较其它水体类型会有所差异.也正因沟渠自身的特点, 目前关于植物类型与反硝化功能基因的关系方面的研究主要在湖泊等较大面积流域中开展(陈登等, 2018), 而在沟渠中还未见相关报道.
4.2 沉积物碳氮含量对nirS、nirK基因拷贝数的影响反硝化微生物的活动是在多种环境因子的综合作用下进行的(Mosier et al., 2010).有研究表明, 氮转化过程的功能微生物丰度和群落结构受沉积物中铵态氮、硝态氮和有机质含量的影响较大(王超等, 2012; Zhang et al., 2014).但也有许多研究发现, 不同环境的土壤中反硝化细菌群落与NO3--N含量具有很弱的相关性(Smith et al., 2007; Chen et al., 2014), 甚至有可能与NO3--N含量变化无关(Kong et al., 2010).另有研究发现, 辽河口沉积物(李博超, 2017)、黄河入海口沉积物(李凤娥, 2017)中的nirK基因拷贝数与其它环境因素也无明显的相关性.出现这些不同情况的原因可能是因为亚硝酸还原细菌的拷贝数变化受到不同理化性质的共同影响(张敏等, 2015).本文进一步探讨了川中丘陵区沟渠沉积物碳氮含量与反硝化功能基因拷贝数的相互关系(表 3), nirK和nirS基因拷贝数与沉积物的NH4+-N含量均呈负相关关系, 与沉积物中的C/N比值呈正相关关系, 但它们之间的相关性并不显著, 说明NH4+-N含量和C/N比有可能对nirS和nirK基因拷贝数有影响, 但并不能用于解释它们之间的相互关系.此外, 结果发现, nirS基因拷贝数与沉积物的NO3--N、TN含量均呈显著性的正相关关系(p < 0.05), 随着沉积物中NO3--N和TN含量的增大, nirS型反硝化微生物的总量也会随之增多.不同研究区域的环境因子不同, 因而沉积物理化性质与反硝化功能基因拷贝数之间的关系也不一致, 还需从RNA水平以及酶水平进行更为深入的研究, 以便更科学地了解沟渠植物对反硝化过程的影响机理, 对于减轻沟渠水体富营养化具有重要意义.
表 3(Table 3)
| 表 3 nirS和nirK基因拷贝数与沉积物碳氮含量的相关性分析 Table 3 The correlationship between abundance of nirS/nirK genes and carbon or nitrogen contents of sediments | ||||||||||||||||||||
表 3 nirS和nirK基因拷贝数与沉积物碳氮含量的相关性分析 Table 3 The correlationship between abundance of nirS/nirK genes and carbon or nitrogen contents of sediments
| ||||||||||||||||||||
4.3 植物类型对沉积物反硝化速率的影响不同类型水生植物在沟渠系统中所处的地位不同, 对沟渠沉积物反硝化作用产生的影响也不同(Knops et al., 2002; 张力等, 2014).Annelies等(2011)研究了大型挺水植物、浮水植物和沉水植物对沟渠反硝化作用的影响, 发现大型挺水植物和浮水植物对反硝化速率的影响较为明显.于洪刚等(2017)通过研究也发现大型挺水植物反硝化速率较其它类型水生植物高.本研究得到的反硝化速率大小在2.85~13.20 μg·m-2·h-1, 表现为:美人蕉 > 铜钱草 > 狐尾藻 > 再力花 > 水竹>茭白 > 水蜡烛 > 空白, 整体上有大型挺水植物 > 浮水植物 > 小型挺水植物的关系.大型挺水植物反硝化速率普遍高于其它类型水生植物, 可能与其强大的地下根茎和密集的地上植株有关.大型挺水植物地下强大的根系可为反硝化微生物提供良好的附着表面, 地上密集的植株枯萎凋落后会沉入水体, 为反硝化微生物提供充足的碳源, 由此大型挺水植物可以促进反硝化微生物的活动, 增强反硝化作用(王博等, 2009).从反硝化功能基因拷贝数的角度来看, 杨柳青(2017)通过研究发现, 反硝化功能基因拷贝数越大, 反硝化作用越容易发生.在本文中, 从植物培养初期到培养至180 d, 所有植物的沉积物反硝化速率显著增加, 反硝化功能基因(nirS和nirK)拷贝数也显著增加.不同植物之间的反硝化功能基因拷贝数的关系大体上有:挺水植物 > 浮水植物.Pearson相关性分析发现, nirK基因拷贝数与反硝化速率间相关性不显著(r=0.513, p > 0.05), 但nirS基因拷贝数与反硝化速率呈显著性的正相关关系(r=0.651, p < 0.05), 说明不同植物之间反硝化速率存在差异的原因也与反硝化功能基因拷贝数有关, 反硝化功能基因拷贝数越大, 反硝化速率越强.总的来说, 不同水生植物都是通过自身的特性改变所处介质环境中的物理和化学条件, 影响了环境中的反硝化微生物, 从而进一步造成了不同植物的沉积物反硝化速率之间的差异性(张力等, 2014).
5 结论(Conclusions)1) 盆栽试验培养至180 d时, 不同植物的沉积物反硝化速率在2.85~13.20 μg·m-2·h-1之间; 与培养初期相比, 反硝化速率有所增加, 整体上表现为:大型挺水植物 > 浮水植物 > 小型挺水植物.
2) 不同植物nirS基因拷贝数在2.70×108~5.02×108 copies·g-1之间, nirK基因拷贝数在3.97×105~6.91×105 copies·g-1之间; 与培养初期相比, 培养180 d后沉积物中nirK、nirS基因拷贝数明显增多.7种植物的基因拷贝数大小顺序整体为:挺水植物 > 浮水植物.
3) nirS基因拷贝数与沉积物NO3--N含量、TN含量及反硝化速率之间均呈显著性的正相关关系(p < 0.05), 但nirK基因拷贝数与沉积物碳氮含量及反硝化速率之间相关性不显著.
参考文献
| Annelies J V, Wilco J J, Jeroen J M, et al. 2011. Effects of aquatic vegetation type on denitrification[J]. Biogeochemistry, 104(1/2/3): 267–274. |
| Braker G, Fesefeldt A, Witzel K P. 1998. Development of PCR primer stems for amplification of nitrite reductase genes(nirK and nirS) to detect denitrifying, bacteria in environment samples[J]. Applied &Environmental Microbiology, 64: 3769–3775. |
| Braker G, Zhou J, Wu L, et al. 2000. Nitrite reductase genes (nirK and nirS) as functional markers to investigate diversity of denitrifying bacteria in pacific northwest marine sediment communities[J]. Applied and Environmental Microbiology, 66(5): 2096–2104.DOI:10.1128/AEM.66.5.2096-2104.2000 |
| Canfield D, Glazer A, Falkowski P. 2010. The evolution and future of earth′s nitrogen cycle[J]. Science, 330: 192–196.DOI:10.1126/science.1186120 |
| Chen Y N, Zhou W, Li Y P, et al. 2014. Nitrite reductase genes as functional markers to investigate diversity of denitrifying bacteria during agricultural waste composting[J]. Applied Microbiology and Biotechnology, 98: 4233–4243.DOI:10.1007/s00253-014-5514-0 |
| 程建华, 窦智勇, 孙庆业. 2016. 铜陵市河流沉积物中硝化和反硝化微生物分布特征[J]. 环境科学, 2016, 37(4): 1362–1370. |
| 陈斌泽. 2013.长期施肥条件下青藏高原高寒草甸土壤中反硝化细菌丰度和群落结构的季节性变化研究[D].兰州: 兰州大学http://cdmd.cnki.com.cn/Article/CDMD-10730-1016190459.htm |
| 陈登, 蔡启佳, 田翠翠. 2018. 3种沉水植物根际对沉积物中典型氮循环微生物功能基因丰度的影响[J]. 云南农业大学学报(自然科学), 2018, 33(2): 314–323. |
| Dang H Y, Wang C, Li J, et al. 2008. Diversity and distribution of sediment nirS-encoding bacterial assemblages in response to environmental gradients in the Eutrophied Jiaozhou Bay, China[J]. Microbial ecology, 58(1): 161–169. |
| Francis C A, OMullan G D, Cornwell J C, et al. 2013. Transitions in nirS-type denitrifier diversity, community composition, and biogeochemical activity along the Chesapeake Bay estuary[J]. Frontiers in Microbiology, 4: 237. |
| Guan X Y, Zhu L L, Li Y X. 2014. Composition and variation of sediment bacterial and nir S-harboring bacterial communities at representative sites of the Bohai Gulf coastal zone, China[J]. World Journal of Microbiology and Biotechnology, 30: 1291–1300.DOI:10.1007/s11274-013-1553-4 |
| 郭丽芸, 时飞, 杨柳燕. 2011. 反硝化菌功能基因及其分子生态学研究进展[J]. 微生物学通报, 2011, 38(4): 583–590. |
| Heylen K, Gevers D, Vanparys B, et al. 2006. The incidence of nirS and nirK and their genetic heterogeneity in cultivated denitrifiers[J]. Environmental Microbiology, 8(11): 2012–2021.DOI:10.1111/emi.2006.8.issue-11 |
| 贺纪正, 张丽梅. 2013. 土壤氮素转化的关键微生物过程及机制[J]. 微生物学通报, 2013, 40(1): 98–108. |
| Kalscheur K N, Rojas M, Peterson C G, et al. 2012. Algal exudates and stream organic matter influence the structure and function of denitrifying bacterial communities[J]. Microbial Ecology, 64: 881–892.DOI:10.1007/s00248-012-0091-1 |
| Kong A, Hristova K, Scow K, et al. 2010. Impacts of different N management regimes on nitrifier and denitrifier communities and N cycling in soil microenvironments[J]. Soil Biology and Biochemistry, 42: 1523–1533.DOI:10.1016/j.soilbio.2010.05.021 |
| Knops J M H, Bradley K L, Wedin D A. 2002. Mechanisms of Plant Species Impacts on Ecosystem Nitrogen Cycling[J]. Ecology Letters, 5(3): 454–466.DOI:10.1046/j.1461-0248.2002.00332.x |
| Lefebvre S, Marmonier P, Peiry J L. 2006. Nitrogen dynamics in rural streams: differences between geomorphologic units[C]//Annales de Limnologie-International Journal of Limnology[J]. EDP Sciences, 42(1): 43-52 |
| 刘杏认, 赵光昕, 张晴雯, 等. 2018. 生物炭对华北农田土壤N2O通量及相关功能基因丰度的影响[J]. 环境科学, 2018, 39(8): 3816–3824. |
| 鲁如坤. 1999. 土壤溶液化学分析方法(第一版)[M]. 北京: 中国农业科技出版社: 308–311. |
| 龙虹竹, 汪涛, 田琳琳, 等. 2016. 川中丘陵区农业源头沟渠反硝化速率特征及其影响因素[J]. 重庆师范大学学报(自然科学版), 2016, 33(4): 166–172. |
| 李博超. 2017.辽河口沉积物反硝化相关功能基因丰度和多样性分析[D].大连: 大连海事大学http://cdmd.cnki.com.cn/Article/CDMD-10158-1017851261.htm |
| 李凤娥. 2017.黄河入海口沉积物反硝化细菌分布特征及其影响因素研究[D].济南: 山东农业大学http://cdmd.cnki.com.cn/Article/CDMD-10434-1017075077.htm |
| 李筱宛, 张亚平, 万宇, 等. 2016. 上覆水不同C/N比条件下沉积物环境反硝化及氨氧化功能基因丰度变化特征[J]. 南京大学学报(自然科学), 2016, 52(1): 27–35. |
| Mathieu N K, Zhu B, Diana K M. 2017. Assessing the influence of different plant species in drainage ditches on mitigation of non-point source pollutants (N, P, and sediments) in the Purple Sichuan Basin[J]. Environmental Monitoring and Assessment, 189: 267.DOI:10.1007/s10661-017-5965-8 |
| McCrackin M L, Elser J J. 2012. Denitrification kinetics and denitrifier abundances in sediments of lakes receiving atmospheric nitrogen deposition (Colorado, USA)[J]. Biogeochemistry, 108(1/3): 39–54. |
| Mosier A C, Francis C A. 2010. Denitrifier abundance and activity across the San Francisco Bay estuary[J]. Environmental Microbiology Reports, 2: 667–676.DOI:10.1111/j.1758-2229.2010.00156.x |
| Mulholland P, Helton A, Poole G, et al. 2008. Stream denitrification across biomes and its response to anthropogenic nitrate loading[J]. Nature, 452(7184): 202–205.DOI:10.1038/nature06686 |
| Page A L, Miller R H, Keeney D R. 1982. Methods of soil analysis: part 2, Chemical and microbiological properties[M]. Madison, Wisconsin: American Society of Agronomy and Soil Science Society of America: 539–579. |
| Pierobon E, Castaldelli G, Mantovani S, et al. 2013. Nitrogen removal in vegetated and unvegetated drainage ditches impacted by diffuse and point sources of pollution[J]. Clean-Soil Air Water, 41: 24–31.DOI:10.1002/clen.201100106 |
| 任光前, 刘守江, 王彦, 等. 2016. 几种沟渠植物对村镇生活污水氮、磷去除效率的对比研究[J]. 资源开发与市场, 2016, 32(5): 599–603. |
| Roley S S, Tank J L, Williams M A, et al. 2012. Hydrologic connectivity increases denitrification in the hyporheic zone and restored floodplains of an agricultural stream[J]. Journal of Geophysical Research: Biogeosciences, 117: 0148–0227. |
| Rysgaard S, Risgaard P N, Nielsen L P, et al. 1993. Nitrification and denitrification in lake and estuarine sediments measured by the 15N dilution technique and isotope Pairing[J]. Applied and Environmental Microbiology, 59(7): 2093–2098. |
| Seitzinger S, Harrison J, Bohlke J, et al. 2006. Denitrification across landscapes and waterscapes: asynthesis[J]. Ecological Applications, 16(6): 2064–2090.DOI:10.1890/1051-0761(2006)016[2064:DALAWA]2.0.CO;2 |
| Senzia M A, Mashauri D A, Mayo A W. 2003. Suitability of constructed wetlands and waste stabilization ponds in waste water reatment: nitrogen transformation and removal[J]. Physics and Chemistry of the Earth, 28: 1117–1124.DOI:10.1016/j.pce.2003.08.033 |
| Smith C J, Nedwell D B, Dong L F, et al. 2007. Diversity and abundance of nitrate reductase genes (narG and napA), nitrite reductase genes (nirS and nirK), and their transcripts in estuarine sediments[J]. Applied and Environmental Microbiology, 73: 3612–3622.DOI:10.1128/AEM.02894-06 |
| Throback I N, Enwall K, Jarvis A, et al. 2004. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacterica with DGGE[J]. Fems Microbiology Ecology, 49(3): 401–417.DOI:10.1016/j.femsec.2004.04.011 |
| 汪涛, 龙虹竹, 赵原, 等. 2016. 川中丘陵区自然沟渠水体氮磷污染状况评价[J]. 山地学报, 2016, 34(2): 150–156. |
| 王博, 叶春, 杨勋, 等. 2009. 腐解黑藻生物量对高硝态氮水体氮素的影响[J]. 环境科学研究, 2009, 22(10): 1198–1203. |
| 王超, 单保庆. 2012. 子牙河水系水和沉积物好氧氨氧化微生物分布特征[J]. 环境科学学报, 2012, 32(12): 2943–2950. |
| 武姣娜, 魏晓东, 李霞, 等. 2018. 植物氮素利用效率的研究进展[J]. 植物生理学报, 2018, 54(9): 1401–1408. |
| 徐红灯, 席北斗, 翟丽华. 2007. 沟渠沉积物对农田排水中氨氮的截留效应研究[J]. 农业环境科学学报, 2007, 26(5): 1924–1928.DOI:10.3321/j.issn:1672-2043.2007.05.064 |
| 谢佳, 马晓航, 代嫣然, 等. 2017. 有机质对城市湿地微生物丰度的影响[J]. 生物技术通报, 2017, 33(10): 217–224. |
| Yin G Y, Hou L J, Liu M, et al. 2017. Effects of multiple antibiotics exposure on denitrification process in the Yangtze Estuary sediments[J]. Chemosphere, 171: 118–125.DOI:10.1016/j.chemosphere.2016.12.068 |
| 于洪刚, 杨希晨, 孟琦, 等. 2017. 富营养化水体中附植生物夏季反硝化脱氮作用研究[J]. 绿色科技, 2017, 20: 36–37. |
| 杨柳青. 2017.石灰性潮土N2O产生过程及相关功能基因丰度和表达[D].北京: 中国农业大学http://cdmd.cnki.com.cn/Article/CDMD-10019-1017164663.htm |
| Zhang X L, Agogué H, Dupuy C, et al. 2014. Relative abundance of ammonia oxidizers, denitrifiers, and anammox bacteria in sediments of hyper-nutrified estuarine tidal flats and in relation to environmental conditions[J]. Clean-Soil, Air, Water, 42(6): 815–823.DOI:10.1002/clen.v42.6 |
| 张敏, 陶然, 杨扬, 等. 2015. 环境因素对不同工艺人工湿地反硝化功能基因丰度的影响[J]. 环境工程学报, 2015, 9(6): 2645–2652. |
| 曾希柏, 王亚男, 王玉忠, 等. 2014. 施肥对设施菜地nirK型反硝化细菌群落结构和丰度的影响[J]. 应用生态学报, 2014, 25(2): 505–514. |
| 郑燕, 侯海军, 秦红灵, 等. 2012. 施氮对水稻土N2O释放及反硝化功能基因(narG/nosZ)丰度的影响[J]. 生态学报, 2012, 32(11): 3386–3393. |
| Zulkifly S, Hanshew A, Young B E, et al. 2012. The epiphytic microbiota of the globally widespread macroalga Cladophora glomerata (Chlorophyta, Cladophorales)[J]. American Journal of Botany, 99: 1541–1552.DOI:10.3732/ajb.1200161 |
