 , 何义亮
, 何义亮 , 袁其懿, 杨俊, 张波
, 袁其懿, 杨俊, 张波上海交通大学, 环境科学与工程学院, 上海 200240
收稿日期: 2018-11-11; 修回日期: 2019-01-03; 录用日期: 2019-01-03
基金项目: 水体污染控制与治理科技重大专项(No.2017ZX07207002)
作者简介: 吴韵斐(1994-), 女, E-mail:wuyunfei312@163.com
通讯作者(责任作者): 何义亮, E-mail:ylhe@sjtu.edu.cn
摘要: 饮用水水源地中抗生素抗性基因会威胁生态安全和人类健康.为了探究抗生素抗性基因(Antibiotic Resistance Genes,ARGs)在金泽水源型水库中的赋存特征,以及水库净化措施对抗生素抗性基因的净化作用,选择微生物活性较高的夏季时期采集水样,采用高通量荧光定量PCR和16S rRNA实时荧光定量PCR进行研究.结果表明,该水库检出62种ARGs;多重抗药类、氨基糖苷类和磺胺类ARGs为水库内占主导的抗性基因.抗生素抗性基因绝对丰度从水库的预处理区至生态净化区,再至输水区呈逐渐降低的趋势,表明水库净化措施对削减ARGs含量有一定作用.抗性基因丰度与可移动基因元件(Mobile Genetic Elements,MGEs)丰度存在显著相关性(p < 0.05),表明MGEs对ARGs的水平转移、传播和富集具有重要作用.
关键词:水源型水库抗生素抗性基因赋存特征水库净化措施高通量定量PCR
Study on antibiotic resistance genes characteristics in a drinking water reservoir
WU Yunfei
, HE Yiliang , YUAN Qiyi, YANG Jun, ZHANG Bo School of Environmental Science and Engineering, Shanghai Jiao Tong University, Shanghai 200240
Received 11 November 2018; received in revised from 3 January 2019; accepted 3 January 2019
Abstract: Antibiotic resistance genes (ARGs) in drinking water source may threaten the ecological system and human health. Water samples were collected in summer and then were analyzed using high-throughput quantitative PCR (HT-qPCR) and real-time quantitative PCR (RT-qPCR), in order to investigate the characteristics of ARGs in Jinze drinking water reservoir and the impact of the water purification processes of the reservoir on ARGs. The results show that 62 target ARGs were detected in the reservoir. Multidrug resistance genes, aminoglycoside resistance genes and sulfonamide resistance genes were the dominant ARGs in the reservoir. The absolute abundances of ARGs in the reservoir show a decline trend from the pretreatment area to the ecological purification area and the water delivery area, indicating that the purification processes in the reservoir played a certain role in the attenuation of ARGs. The abundance of ARGs was significantly correlated to the abundance of mobile genetic elements (MGEs) (p < 0.05), implicating that MGEs play an important role in the horizontal transfer, dissemination and accumulation of ARGs.
Keywords: drinking water reservoirantibiotic resistance genesoccurrence characteristicswater purification processeshigh-throughput quantitative PCR
1 引言(Introduction)抗生素长期应用于人和动物疾病的治疗, 并添加于饲料中作为生长促进剂(Sarmah et al., 2006).然而, 由于抗生素不能被机体完全吸收和代谢, 大多以原形或代谢产物的形式, 通过尿液和粪便排出体外, 并通过不同途径进入到环境中(Roman Hirsch et al., 1999).环境中残留的抗生素不仅对环境造成污染, 还会诱导抗生素抗性细菌和抗生素抗性基因(Antibiotic Resistance Genes, ARGs)的产生, 降低抗生素对人和动物病原菌感染的治疗能力(Qiao et al., 2018).国内外已有研究在医疗废水(Rodriguez-Mozaz et al., 2015)、养殖场污水(Chen et al., 2015a)、污水处理厂(Yang et al., 2013)、地表水(邹世春等, 2009; Chen et al., 2013a)等水体环境中检测到抗生素抗性基因.抗生素抗性基因具有可复制、可传播、环境持久等特征, 其在环境中的残留和传播, 将严重威胁人类健康和生态安全(文汉卿等, 2015), 被认为是一种新型环境污染物.
饮用水源地水环境与人类健康安全密切相关, 目前, 大多数研究集中于对水源地氮(宋国强等, 2009)、磷(Jiang et al., 2016)、持久性有机污染物(李霞等, 2015)、重金属(Jiang et al., 2017)等污染物的污染特征进行探索, 而对饮用水源地抗生素抗性基因的研究报道相对较少.此外, 大量研究仅仅针对少数几种至几十种ARGs, 且以磺胺类ARGs和四环素类ARGs为主.饮用水源地抗生素抗性基因污染会对人类健康造成威胁, 因此, 对于抗生素抗性基因在水源地中的赋存情况进行系统而全面的研究是十分必要的.此外, 生态净化工艺逐渐融入水源型水库的建设以满足水资源的可持续发展.然而目前大部分研究集中于生态净化工艺对氮、磷等常规污染物的去除(郑立国等, 2013), 也有试验研究证实人工湿地等生态净化工艺对抗生素及抗生素抗性基因有一定的去除效率(Liu et al., 2013; Chen et al., 2016), 但各种净化工艺在实际应用中对抗生素抗性基因的去除效率还未明确, 特别是应用于水库的净化工艺, 对抗生素抗性基因的净化作用有待研究, 以此为水库的污染控制提供依据.
金泽水库是上海重要的水源型水库, 也是长三角地区首座应用生态净化措施与物理净化措施相结合的生态型水库.本研究以金泽水库为研究对象, 选择微生物活性较高的夏季(6—8月), 采集水样并使用高通量荧光定量PCR和实时荧光定量PCR方法, 检测水体中九大类283种抗生素抗性基因, 对其多样性和丰度进行研究, 并分析水库净化措施对ARGs的净化作用, 以此评价水库净化措施对ARGs的净化效果, 为水库ARGs污染控制提供依据.
2 材料与方法(Materials and methods)2.1 样品采集与处理金泽水库是上海市黄浦江上游水源水库, 向上海西南五区供水, 惠及约670万人口.水库位于青浦区金泽镇西部、太浦河北岸, 其原水取自太浦河, 占地面积约2.7 km2, 总容量910万m3, 供水规模为351万m3·d-1.整个水库可以分为3个区域:预处理区、生态净化区、输水区(图 1).预处理区包含取水闸、引水河道; 生态净化区包含李家荡和乌家荡两大库区, 输水区为输水闸所在处.
图 1(Fig. 1)
 |
| 图 1 金泽水库采样点 Fig. 1Sampling sites in Jinze Reservoir |
本研究于2018年夏季(6、7和8月)进行采样, 每月一次, 在水库范围内共设置6个采样点(图 1).其中, 采样点S1位于水库取水闸外, 太浦河上; S2位于水库的引水河道; S3位于引水河道的终端; S4、S6分别位于李家荡和乌家荡; S5位于水库输水闸.S1、S2、S3属于预处理区, S4、S6属于生态净化区, S5属于输水区.
在各采样点距离水面0.5 m处采集10 L水样, 并立即运回实验室, 用0.22 μm混合纤维素滤膜(Millipore, 美国)过滤, 各样品在滤膜上过滤的体积都为1 L, 将滤膜于-20 ℃保存.
2.2 水样DNA提取水样DNA使用FastDNA? Spin Kit for Soil试剂盒(MP Biomedicals, USA)提取.将滤膜剪成小块放入试剂盒提供的Lysing Matrix E tube离心管中, 按照试剂盒操作说明, 提取水样总DNA, 并于-20 ℃保存.提取的DNA样品用1%琼脂糖凝胶电泳、Nanodrop2000微量分光光度计(Thermo Scientific, USA)进行纯度和浓度的检测.根据测定的DNA样品浓度, 用无菌水将所有样品统一稀释至40 ng·μL-1, 用于高通量荧光定量PCR实验.
2.3 高通量荧光定量PCR高通量荧光定量PCR在SmartChip Real-Time PCR system(WaferGen Inc., USA)高通量荧光定量反应平台进行.使用的296对基因引物已在先前的研究中被有效验证(An et al., 2018; 张丹丹等, 2018), 包括283对抗生素抗性基因引物, 1对16S rRNA基因引物, 8对转座子基因引物, 4对整合子基因引物.荧光定量试剂LightCycler 480 SYBR? Green Ι Master Mix (Roche, USA).定量体系体积100 nL, 体系中各试剂终浓度为:1×LightCycler 480 SYBR? Green Ι Master Mix, 1 μg·μL-1牛血清蛋白(BSA), 5 ng·μL-1 DNA模板, 1 μmol·L-1上下游引物, Nuclease-free PCR-Grade water.
定量PCR反应条件:95 ℃预变性10 min; 95 ℃变性30 s, 60 ℃退火延伸30 s, 40个循环; 程序自动升温进行熔解曲线分析.qPCR反应得到的数据通过Cycler预先设定的筛选条件(PCR扩增效率介于1.8~2.2)进行导出.根据SmartChip Real-Time System平台的精确度和灵敏度, 确定循环次数CT=31为仪器的检测限.
2.4 16S rRNA基因实时荧光定量PCR本实验使用Roche 480Ⅱ实时荧光定量PCR仪(Roche, USA)测定所有样品中的16S rRNA基因, 采用标准质粒外标法对其丰度进行绝对定量(李丽等, 2011).标准质粒原始浓度为1.39×1010 copies·L-1, 将以10倍浓度梯度稀释的质粒用于制作绝对定量标准曲线, 标准曲线范围为1.39×102~1.39×109 copies·L-1.
PCR反应体系体积20 μL, 其中, 10 μL 2×LightCycler 480 SYBR? Green Ι Master Mix (Roche, USA), 上、下游引物(10 μmol·L-1)各1 μL, 1 μL DNA模板, 7 μL无菌水.使用无菌水代替样品进行阴性对照实验.
PCR反应条件:95 ℃预变性5 min; 95 ℃变性30 s, 60 ℃退火1 min, 72 ℃延伸30 s, 40个循环; 仪器自动添加溶解曲线程序.
2.5 数据分析根据Looft等(2012)的方法, 基因的相对拷贝数由公式(1)计算, 根据黄福义等(2016)的方法, 绝对丰度由公式(2)计算.
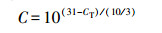 | (1) |
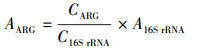 | (2) |
利用Microsoft Excel 2010进行计算分析, 采用SPSS 20.0进行相关性分析, 使用Origin Pro 9.0制图.
3 结果与讨论(Results and discussion)3.1 抗生素抗性基因的多样性按照基因对应抗生素类型, 将283种使用的抗性基因引物分为九大类ARGs.九大类ARGs分别为氨基糖苷类抗生素(Aminoglycoside)抗性基因(36种)、β-内酰胺类抗生素(β-Lactamase)抗性基因(52种)、氯霉素类抗生素(Chloramphenicol)抗性基因(3种)、大环内酯类-林肯酰胺类-链阳性菌素B类抗生素(MLSB)抗性基因(45种)、多重抗药类抗生素(Multidrug)抗性基因(52种)、磺胺类抗生素(Sulfonamide)抗性基因(7种)、四环素类抗生素(Tetracycline)抗性基因(39种)、万古霉素类抗生素(Vancomycin)抗性基因(32种)和其他类抗生素(Others)抗性基因(17种).此外, 可移动基因元件(mobile genetic elements, MGEs)(12种)分为8种转座子基因和4种整合子基因.
图 2显示了3个采样时期各采样点检测到的抗生素抗性基因的类型及种类数, 九大类ARGs和MGEs在水样中均有检出.所有水样中检测出的基因种类数为54~71种, 其中, 6月S6样点检出的种类数最多(71种), 7月S6样点检出的种类数最少(54种).夏季时期, 该水源地平均检出62种ARGs, 其中6月所有采样点平均检出64种, 7月平均检出59种, 8月平均检出62种.有研究对其它地表水体中这九大类ARGs进行了高通量检测, 发现长江下游某河口型水库(江月等, 2017)夏季时期平均检测到118种抗性基因, 福建省敖江下游(张丹丹等, 2018)平均检测到108种抗性基因, 都大于该水库抗性基因检出的种类数, 说明金泽水库抗性基因污染程度相对较低.
图 2(Fig. 2)
 |
| 图 2 检测到的抗生素抗性基因种类数 Fig. 2Profiles of detected antibiotic resistance genes |
S1在3个采样时期共检出ARGs198种, S2共检出189种, S3共检出175种, S4共检出179种, S5共检出179种, S6共检出191种, 库外(S1)检出的抗生素种类数相比库内(S2~S6)更多, 表明水库的净化作用在一定程度上降低了ARGs的种类.
3.2 抗生素抗性基因丰度九大类抗生素抗性基因在水样中均有检出, 但在含量水平上存在差异.该水源地ARGs的绝对丰度如图 3a所示, ARGs绝对丰度为n.d.~7.97×109 copies·L-1, MGEs绝对丰度为n.d.~1.78×1010 copies·L-1.九大类ARGs中, 多重抗药类ARGs呈现最高绝对丰度(2.71×109~2.01×1010 copies·L-1), 其次为氨基糖苷类ARGs(8.02×108~5.81×109 copies·L-1)和磺胺类ARGs(3.86×108~4.23×109 copies·L-1), 氯霉素类ARGs呈现最低绝对丰度(0~1.28×108 copies·L-1).
图 3(Fig. 3)
 |
| 图 3 抗生素抗性基因丰度 (a.绝对丰度, b.相对丰度) Fig. 3Abundance of antibiotic resistance genes (a.Absolute abundance, b.Relative abundance) |
Klappenbach等(2001)研究表明, 基于rRNA数据库, 每个细菌平均约有4个16S rRNA基因的拷贝, 因此, 利用相对丰度可以在单个细菌水平上评估ARGs的赋存情况.该水源地各样品ARGs的相对丰度如图 3b所示, ARGs相对丰度为n.d.~5.69×10-2, MGEs相对丰度为n.d.~1.11×10-1.九大类ARGs中, 相对丰度较高的3类为多重抗药类ARGs(2.28×10-2~1.12×10-1)、氨基糖苷类ARGs(4.44×10-3~3.24×10-2)、磺胺类ARGs(2.48×10-3~2.36×10-2).抗性基因相对丰度结果与绝对丰度一致, 多重抗药类ARGs、氨基糖苷类ARGs和磺胺类ARGs为水库内主要的抗生素抗性基因.
不同种类ARGs的含量水平存在差异, 可能与该地区抗生素的使用情况有关, 抗生素残留的环境选择压力, 诱导产生ARGs.金泽水库原水取自太浦河, 而太浦河沿线及水库周边地区多为农田、城镇村及水产或畜禽养殖用地.根据调查(李昊洋等, 2017), 水库上游太浦河沿线有重要的畜禽养殖基地, 规模化养殖及散户养殖数量较大, 主要养殖品种为猪和家禽, 且太浦河沿线用地中水域用地占多数, 水产养殖成为该地区的重要支柱产业; 此外, 太浦河沿线耕地面积总计达81.72万亩, 各乡镇农村人口约69.7万, 城镇人口约39.7万; 对于太浦河及水库周边地区点源污染的调查发现, 水库上游地区至水库周边共有污水处理厂30座.抗生素被广泛用于人和动物疾病的治疗, 或添加于饲料中以促进动物生长, 在中国用于畜禽养殖的抗生素以喹诺酮类抗生素和β-内酰胺类抗生素为主(Zhang et al., 2015), 氨基糖苷类和磺胺类抗生素在水产养殖业中被广泛应用(Liu et al., 2017), 人类疾病的治疗以β-内酰胺类抗生素为主.然而, 抗生素无法被机体完全吸收, 畜禽养殖中产生的动物粪便含有大量抗生素, 渗入土壤并通过地表径流进入水环境; 居民使用的抗生素同样通过排泄物排出体外, 使得大量抗生素存在于生活污水中, 而抗生素在污水处理厂中不能被完全去除(徐维海等, 2007), 由污水处理厂出水排入河流中; 而水产养殖业中使用的抗生素更是被直接添加到水环境中.
27种(51.92%)多重抗药类抗性基因(52种)被检出, 其中5种检出率为100%.随着抗生素的过度使用, 细菌的抗药性逐渐由单一抗药性发展为多重抗药性(高盼盼等, 2009), 多重抗药抗性基因会使携带这种基因的微生物对多种抗生素具有耐药性, 从而使其成为“超级细菌”(庄榆佳等, 2017).由于金泽水库周边及上下游地区有大规模水产养殖区和畜禽养殖区及大面积城镇村, 养殖业产生的含抗生素废水及居民产生的生活污水通过各种途径进入水体, 在残留于水体中的各种抗生素诱导下, 产生多重抗药基因, 对生态安全和人体健康造成极大威胁.
氨基糖胺类ARGs和磺胺类ARGs的丰度相比其他几种ARGs处于较高水平.18种(50%)氨基糖苷类抗性基因(36种)被检出, 其中9种检出率为100%.各样品氨基糖胺类ARGs的绝对丰度均值为5.13×104~3.18×108 copies·L-1, 相对丰度均值为2.73×10-7~2.00×10-3.7种磺胺类抗性基因中, 仅sul(Ⅱ)和dfrA1被检出, 且检出率均为100%, 其余磺胺类抗性基因均未检出, 与在德国和澳大利亚水体中的检出情况一致(Stoll et al., 2012), 表明这两种基因在水体中的存在具有普遍性.其中, 各样品sul(Ⅱ)绝对丰度和相对丰度分别为2.94×108~3.72×109 copies·L-1和1.89×10-3~2.07×10-2; dfrA1绝对丰度和相对丰度分别为5.69×107~5.11×108 copies·L-1和3.14×10-4~2.85×10-3.氨基糖苷类和磺胺类抗生素被广泛应用于水产养殖业中(Liu et al., 2017).Luo等(2010)对中国海河流域抗生素及抗生素抗性基因含量进行了检测, 结果表明, 磺胺类抗生素抗性基因丰度与磺胺类抗生素浓度存在显著正相关, 抗生素的持续存在选择性地诱导了相应的抗性基因.因此, 本研究中氨基糖苷类ARGs和磺胺类ARGs丰度相对较大, 可能与这两类抗生素在该地区养殖业中的大量使用, 导致其在环境中的残留有关, 进而诱导产生抗生素抗性基因.
β-内酰胺类ARGs和四环素类ARGs在该水库中的检出浓度相比其他几类ARGs处于中等水平.52种β-内酰胺类ARGs有25种被检出, 绝对丰度和相对丰度的均值分别为9.71×104~3.54×108 copies·L-1和5.41×10-7~2.08×10-3.39种四环素类抗生素抗性基因中检出18种, 其中, tetG-01和tetG-02检出率为100%, 其余检出率均小于50%.各样品四环素类ARGs绝对丰度和相对丰度的均值分别为5.00×104~1.22×108 copies·L-1和2.66×10-7~7.49×10-4.Ling等(2013)对河流水体中tet基因含量进行了检测, 其中, tetG检出率最高, tetE和tetS未检出, 本研究结果与其一致, 但在该河流中tet基因的相对丰度(4.3×10-6~1.37×10-1)高于金泽水库, 表明水库系统相比河流受tet抗性基因污染的影响相对较小.
3.3 水库净化措施对抗生素抗性基因的净化作用金泽水库作为一个生态型水库, 不仅采用了常规物理净化措施, 更是应用了生态净化措施.整个水库可以分为3个区域:预处理区、生态净化区和输水区(图 1).预处理区包括进口取水闸前设置的拦污排, 设有回转式格栅清污机的取水闸, 以及作为入库沉淀区的引水河道, 同时, 引水河道的两岸放置了微纳米充氧设备, 有增氧、强化净化效果的作用.生态净化区, 即水库的主要区域, 由李家荡库区和乌家荡库区两大库区组成, 近1700 m2的生态小岛横卧于两大库区之间; 库区堤坝设计为设置生态砾石床的生态护坡, 有利于微生物的富集从而净化水质; 库内生态净化区还设有人工浮岛, 人工浮岛和生态护坡都种植水生植物.原水经取水闸引水, 流经预处理区和生态净化区后, 到达输水区.ARGs在预处理区、生态净化区和输水区的分布情况如图 4所示, 除β-内酰胺类ARGs和万古霉素ARGs以外, 其他各类ARGs和MGEs绝对丰度从预处理区到生态净化区, 再至输水区呈逐渐降低趋势, 其中, 氨基糖苷类ARGs、磺胺类ARGs绝对丰度从预处理区至输水区显著降低(p < 0.05), 表明水库净化措施对抗生素抗性基因, 尤其是氨基糖苷类ARGs和磺胺类ARGs的绝对丰度具有削减作用.
图 4(Fig. 4)
 |
| 图 4 抗生素抗性基因分布 (图中显著性水平p < 0.05) Fig. 4Distribution of antibiotic resistance genes |
ARGs的赋存受抗生素污染水平、微生物含量、水生植物种类等多种因素的影响.水库取水闸前的拦污排及格栅清污机能够有效去除水中悬浮颗粒物, 引水河道有沉降颗粒物的作用, 由于部分抗生素(如氟喹诺酮类及四环素类)易吸附于固相颗粒物上, 且有相关研究表明抗生素含量与ARGs含量显著相关(Xu et al. , 2015).因此, ARGs丰度在经过预处理区后有所降低, 可能是由于随着颗粒物的去除和沉降, 水体中的抗生素含量相应减少, 减弱了抗生素对ARGs的选择压力, 使得AGRs丰度也随之降低.生态净化区种植芦苇、水葱、美人蕉、鸢尾等水生植物, 有****(Chen et al., 2013b)通过研究栽种有不同水生植物的人工湿地系统对抗生素抗性基因的去除效果, 发现ARGs在不同系统中去除效率的差异可能与水生植物的种类有关; Fang等(2017)研究表明, 芦苇是降低ARGs污染的重要水生植物种类.因此, 生态净化区人工浮岛和生态护坡种植的水生植物, 可能在一定程度上起到降低水体中ARGs丰度的作用.鉴于水生植物在去除ARGs方面的效果, 本研究认为, 可以通过改变水库生态净化区栽种的水生植物的种类, 以达到最佳净化效果, 例如, 多栽种芦苇等对ARGs有削减效果的水生植物.另一方面, 由于ARGs通常由微生物携带, 微生物的去除有利于ARGs的去除(Huang et al., 2015), 而16S rRNA可以用来表征细菌生物量大小; 也有研究证实, 抗生素抗性基因拷贝数与16S rRNA基因拷贝数显著相关(Wu et al., 2010; Chen et al., 2013b).然而本研究中, 16S rRNA丰度在3个区域内无显著差异(p > 0.05)(图 4), 因此, 可以推测从预处理区到输水区抗生素抗性基因丰度的降低, 并不是由于微生物含量降低引起的, 而是生态净化区水生植物发挥了重要作用.此外, 人工浮岛的土壤或基质的吸附作用及生物降解是其去除ARGs的两个主要机制(Chen et al., 2015b), 且不同的基质类型对ARGs的去除效率也不同(Liu et al., 2013), 因此, ARGs丰度从预处理区到输水区的依次降低, 也可能与生态净化区的人工浮岛的基质吸附和植物根系富集的微生物的降解作用有关.本研究认为, 可以在水库各区域再布设一定数量的人工浮岛, 并对人工浮岛的基质类型进行改善, 已达到更好的净化效果.
综合上述研究结果, 水库中的人工浮岛和水生植物等净化措施在削减AGRs丰度上发挥了一定的作用, 如何对净化工艺进行改善, 以达到最佳的净化效果和水库管理模式还有待进一步的研究.
3.4 抗生素抗性基因和可移动元件的相关性分析可移动基因元件(MGEs)包括整合子基因和转座子基因, 可以捕获和表达多种抗性基因(Gillings et al., 2008).本研究中, 除intⅠ2和tnpA-03在所有样品中均未检出外, 其余均有检出, 且MGEs绝对丰度(5.65×109~2.47×1010 copies·L-1)和相对丰度(3.18×10-2~1.38×10-1)都显著高于其他各类ARGs的丰度(p < 0.05).梁惜梅等(2013)研究发现, int1与总ARGs的丰度存在显著相关性; Zhu等(2017)研究发现, ARGs绝对丰度与整合子和转座子分别呈显著正相关.本研究对各类抗生素抗性基因和可移动元件之间的相关性进行了研究(表 1), 结果表明, 氨基糖苷类ARGs、多重抗药类ARGs、磺胺类ARGs与MGEs存在显著相关关系(p < 0.01), 氯霉素类ARGs、大环内酯类-林肯酰胺类-链阳性菌素B类抗生素、四环素类ARGs与MGEs存在显著相关性(p < 0.05), 总ARGs丰度与MGEs也存在显著相关关系(p < 0.01).先前的研究也证实, 转座子和整合子基因能够携带ARGs, 使ARGs在不同细菌间传播(Galimand et al., 2005; Poole et al., 2006; Ma et al., 2017), 因此, 水体中ARGs的传播可能依赖于MGEs, 推测MGEs对这几类ARGs在水库中的迁移、传播和富集发挥重要作用, 能够促进AGRs的水平转移从而加剧水环境中抗生素抗性基因的污染.
表 1(Table 1)
| 表 1 抗性基基因丰度与可移动基因元件的相关性 Table 1 Correlation analysis between ARGs and MGEs | ||||||||||||||||||||||||
表 1 抗性基基因丰度与可移动基因元件的相关性 Table 1 Correlation analysis between ARGs and MGEs
| ||||||||||||||||||||||||
4 结论(Conclusions)1) 夏季时期, 金泽水库平均检出62种抗生素抗性基因, 与其他地区相比处于低污染水平.
2) 九大类ARGs中, 多重抗药类ARGs、氨基糖苷类ARGs和磺胺类ARGs呈现最高的绝对丰度和相对丰度, 为水库中占主导的ARGs.
3) 抗生素抗性基因丰度从水库预处理区至生态净化区, 最后至输水区呈现逐渐减小的趋势; 水库净化措施对削减水中的ARGs具有一定作用, 为达到最佳的净化效果, 水库各净化工艺的改善措施和管理模式还有待进一步的研究.
4) ARGs和可移动基因元件之间存在显著正相关关系, 表明MGEs能够促进ARGs的水平转移过程, 并促进抗性基因的迁移、传播和富集, 加剧污染, 其具体机制还需进一步研究.
参考文献
| An X L, Chen Q L, Zhu D, et al. 2018. Impact of wastewater treatment on the prevalence of integrons and the genetic diversity of integron gene cassettes[J]. Applied and Environmental Microbiology, 84(9): e02766–02717. |
| Chen B, Hao L, Guo X, et al. 2015a. Prevalence of antibiotic resistance genes of wastewater and surface water in livestock farms of Jiangsu Province[J]. Environmental Science and Pollution Reserch, 22(18): 13950–13959.DOI:10.1007/s11356-015-4636-y |
| Chen B, Liang X, Huang X, et al. 2013a. Differentiating anthropogenic impacts on ARGs in the Pearl River Estuary by using suitable gene indicators[J]. Water Research, 47(8): 2811–2820.DOI:10.1016/j.watres.2013.02.042 |
| Chen H, Zhang M. 2013b. Effects of advanced treatment systems on the removal of antibiotic resistance genes in wastewater treatment plants from Hangzhou, China[J]. Environmental Science & Technology, 47(15): 8157–8163. |
| Chen J, Wei X D, Liu Y S, et al. 2016. Removal of antibiotics and antibiotic resistance genes from domestic sewage by constructed wetlands:Optimization of wetland substrates and hydraulic loading[J]. Science of the Total Environment, 565: 240–248.DOI:10.1016/j.scitotenv.2016.04.176 |
| Chen J, Liu Y, Su H, et al. 2015b. Removal of antibiotics and antibiotic resistance genes in rural wastewater by an integrated constructed wetland[J]. Envrionmental Science Pollution Research, 22(3): 1794–1803.DOI:10.1007/s11356-014-2800-4 |
| Fang H, Zhang Q, Nie X, et al. 2017. Occurrence and elimination of antibiotic resistance genes in a long-term operation integrated surface flow constructed wetland[J]. Chemosphere, 173: 99–106.DOI:10.1016/j.chemosphere.2017.01.027 |
| Galimand M, Sabtcheva S, Courvalin P, et al. 2005. Worldwide disseminated armA aminoglycoside resistance methylase gene is borne by composite transposon Tn1548[J]. Antimicrobial Agents and Chemotherapy, 49(7): 2949–2953.DOI:10.1128/AAC.49.7.2949-2953.2005 |
| 高盼盼, 罗义, 周启星, 等. 2009. 水产养殖环境中抗生素抗性基因(ARGs)的研究及进展[J]. 生态毒理学报, 2009, 4(6): 770–779. |
| Gillings M, Boucher Y, Labbate M, et al. 2008. The evolution of class 1 integrons and the rise of antibiotic resistance[J]. Journal of Bacteriology, 190(14): 5095–5100.DOI:10.1128/JB.00152-08 |
| 黄福义, 安新丽, 陈青林, 等. 2016. 梅花鹿养殖场抗生素抗性基因分布特征[J]. 环境科学, 2016, 37(11): 4402–4409. |
| Huang X, Liu C, Li K, et al. 2015. Performance of vertical up-flow constructed wetlands on swine wastewater containing tetracyclines and tet genes[J]. Water Research, 70: 109–117.DOI:10.1016/j.watres.2014.11.048 |
| 江月, 陈奕涵, 何义亮. 2017. 长江下游某水源型水库抗生素抗性基因污染研究[J]. 环境科学学报, 2017, 38(3): 884–891. |
| Jiang L G, Liang B, Xue Q, et al. 2016. Characterization of phosphorus leaching from phosphate waste rock in the Xiangxi River watershed, Three Gorges Reservoir, China[J]. Chemosphere, 150: 130–138.DOI:10.1016/j.chemosphere.2016.02.008 |
| Jiang X, Xiong Z, Liu H, et al. 2017. Distribution, source identification, and ecological risk assessment of heavy metals in wetland soils of a river-reservoir system[J]. Environmental Science Pollution Research, 24(1): 436–444.DOI:10.1007/s11356-016-7775-x |
| Klappenbach J A, Saxman P R, Cole J R, et al. 2001. rrndb:the ribosomal RNA operon copy number database[J]. Nucleic Acids Research, 29(1): 181–184.DOI:10.1093/nar/29.1.181 |
| 李昊洋, 周宏伟, 陆沈钧. 2017. 太浦河沿线污水环境污染源调查[J]. 水利技术监督, 2017, 25(4): 52–56.DOI:10.3969/j.issn.1008-1305.2017.04.020 |
| 李丽, 赵成萍, 李宏, 等. 2011. 质粒制备绝对定量PCR标准曲线方法的建立[J]. 农业生物技术学报, 2011, 19(6): 1157–1162.DOI:10.3969/j.issn.1674-7968.2011.06.024 |
| 李霞, 王东红, 王金生, 等. 2015. 北京市枯水季和丰水季水源水中持久性有机污染物的水平分析[J]. 环境科学学报, 2015, 35(2): 437–442. |
| 梁惜梅, 聂湘平, 施震. 2013. 珠江口典型水产养殖区抗生素抗性基因污染的初步研究[J]. 环境科学, 2013, 34(10): 4073–4080. |
| Ling Z, Yang Y, Huang Y, et al. 2013. A preliminary investigation on the occurrence and distribution of antibiotic resistance genes in the Beijiang River, South China[J]. Journal of Environmental Sciences, 25(8): 1656–1661.DOI:10.1016/S1001-0742(12)60223-X |
| Liu L, Liu C, Zheng J, et al. 2013. Elimination of veterinary antibiotics and antibiotic resistance genes from swine wastewater in the vertical flow constructed wetlands[J]. Chemosphere, 91(8): 1088–1093.DOI:10.1016/j.chemosphere.2013.01.007 |
| Liu X, Steele J C, Meng X Z. 2017. Usage, residue, and human health risk of antibiotics in Chinese aquaculture:A review[J]. Environmental Pollution, 223: 161–169.DOI:10.1016/j.envpol.2017.01.003 |
| Looft T, Johnson T A, Allen H K, et al. 2012. In-feed antibiotic effects on the swine intestinal microbiome[J]. Proceedings of the National Academy of Sciences, 109(5): 1691–1696.DOI:10.1073/pnas.1120238109 |
| Luo Y, Mao D, Rysz M, et al. 2010. Trends in antibiotic resistance genes occurrence in the Haihe River, China[J]. Environmental Science & Technology, 44(19): 7220–7225. |
| Ma L, Li A D, Yin X L, et al. 2017. The prevalence of integrons as the carrier of antibiotic resistance genes in natural and man-made environments[J]. Environmental Science & Technology, 51(10): 5721–5728. |
| Poole T L, Callaway T R, Bischoff K M, et al. 2006. Macrolide inactivation gene cluster mphA-mrx-mphR adjacent to a class 1 integron in Aeromonas hydrophila isolated from a diarrhoeic pig in Oklahoma[J]. Journal of Antimicrobial Chemotherapy, 57(1): 31–38.DOI:10.1093/jac/dki421 |
| Qiao M, Ying G G, Singer A C, et al. 2018. Review of antibiotic resistance in China and its environment[J]. Environmental International, 110: 160–172.DOI:10.1016/j.envint.2017.10.016 |
| Rodriguez-Mozaz S, Chamorro S, Marti E, et al. 2015. Occurrence of antibiotics and antibiotic resistance genes in hospital and urban wastewaters and their impact on the receiving river[J]. Water Research, 69: 234–242.DOI:10.1016/j.watres.2014.11.021 |
| Roman Hirsch T T, Klaus Haberer, Karl-Ludwig Kratz. 1999. Occurrence of antibiotics in the aquatic environment[J]. Science of the Total Environment, 225: 109–118.DOI:10.1016/S0048-9697(98)00337-4 |
| Sarmah A K, Meyer M T, Boxall A B. 2006. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment[J]. Chemosphere, 65(5): 725–759.DOI:10.1016/j.chemosphere.2006.03.026 |
| 宋国强, 殷明, 张卫东, 等. 2009. 丹江口水库入库河流总氮通量监测[J]. 环境科学与技术, 2009, 32(12): 135–137.DOI:10.3969/j.issn.1003-6504.2009.12.031 |
| Stoll C, Sidhu J P S, Tiehm A, et al. 2012. Prevalence of clinically relevant antibiotic resistance genes in surface water samples collected from Germany and Australia[J]. Environmental Science & Technology, 46(17): 9716–9726. |
| Wu N, Qiao M, Zhang B, et al. 2010. Abundance and diversity of tetracycline resistance genes in soils adjacent to representative swine feedlots in China[J]. Environmental Science & Technology, 44(18): 6933–6939. |
| 文汉卿, 史俊, 寻昊, 等. 2015. 抗生素抗性基因在水环境中的分布、传播扩散与去除研究进展[J]. 应用生态学报, 2015, 26(2): 625–635. |
| Xu J, Xu Y, Wang H, et al. 2015. Occurrence of antibiotics and antibiotic resistance genes in a sewage treatment plant and its effluent-receiving river[J]. Chemosphere, 119: 1379–1385.DOI:10.1016/j.chemosphere.2014.02.040 |
| 徐维海, 张干, 邹世春, 等. 2007. 典型抗生素类药物在城市污水处理厂中的含量水平及其行为特征[J]. 环境科学, 2007, 28(8): 1779–1783.DOI:10.3321/j.issn:0250-3301.2007.08.023 |
| Yang Y, Li B, Ju F, et al. 2013. Exploring variation of antibiotic resistance genes in activated sludge over a four-year period through a metagenomic approach[J]. Environmental Science & Technology, 47(18): 10197–10205. |
| 张丹丹, 郭亚平, 任红云, 等. 2018. 福建省敖江下游抗生素抗性基因分布特征[J]. 环境科学, 2018, 39(6): 2600–2606. |
| Zhang Q Q, Ying G G, Pan C G, et al. 2017. Comprehensive evaluation of antibiotics emission and fate in the river basins of China:source analysis, multimedia modeling, and linkage to bacterial resistance[J]. Environmental Science & Technology, 49(11): 6772–6782. |
| 郑立国, 杨仁斌, 王海萍, 等. 2013. 组合型生态浮床对水体修复及植物氮磷吸收能力研究[J]. 环境工程学报, 2013, 7(6): 2153–2159. |
| Zhu Y G, Zhao Y, Li B, et al. 2017. Continental-scale pollution of estuaries with antibiotic resistance genes[J]. Nature Microbiology, 2: 16270.DOI:10.1038/nmicrobiol.2016.270 |
| 庄榆佳, 赵忆, 苏建强, 等. 2017. 抗生素抗性基因在养殖废水中的分布与去除[J]. 环境化学, 2017, 36(11): 2311–2318.DOI:10.7524/j.issn.0254-6108.2017041801 |
| 邹世春, 朱春敬, 贺竹梅, 等. 2009. 北江河水中抗生素抗性基因污染初步研究[J]. 生态毒理学报, 2009, 4(5): 655–660. |
