 ,1,*, 王李花3, 傅明楷4, 张兆群1,2,5
,1,*, 王李花3, 傅明楷4, 张兆群1,2,5An Upper Miocene “Hipparion fauna” locality sandwiched by basalt in Hanjiaying, Nei Mongol
WANG Qian1,2, LIU Yan,1,*, WANG Li-Hua3, Mikael FORTELIUS4, ZHANG Zhao-Qun1,2,5通讯作者: *liuyan@ivpp.ac.cn
收稿日期:2020-10-21网络出版日期:2021-04-20
| 基金资助: |
Corresponding authors: *liuyan@ivpp.ac.cn
Received:2020-10-21Online:2021-04-20

摘要
内蒙古乌兰察布市集宁区韩家营村附近发现了一处夹在两层玄武岩之间的三趾马动物群化石地点。通过K-Ar同位素测年,确定了上、下两层玄武岩的年代分别为6.8和7.2 Ma。与中国北方其他三趾马动物群相比,韩家营三趾马动物群与山西保德、内蒙古四子王旗以及甘肃临夏盆地的三趾马动物群非常相似。保德第43、44、49地点与韩家营的动物群组成类似,表明这几个地点的时代约为7.0 Ma而不是5.5 Ma。内蒙古四子王旗乌兰花地点与韩家营的化石类群也十分相像,证实了该动物群年龄约为7 Ma。对比甘肃临夏盆地的动物群,柳树组上部的杨家山三趾马动物群与韩家营的更接近。韩家营动物群的组成和牙齿结构表明其属于中国晚中新世的西部生物区系Gazella dorcadoides动物群。
关键词:
Abstract
Here we report a Hipparion fauna locality discovered in between two basalt layers near Hanjiaying Village, Jining District, Wulanchabu City, Nei Mongol. K-Ar isotopic dating of the lower and upper level of the basalt constrains the age of the fauna from 7.2 to 6.8 Ma. Compared with classical Hipparion fauna from northern China, the Hanjiaying fauna is closer to those from Baode of Shanxi, Siziwang Banner of Nei Mongol and the Linxia Basin of Gansu. It is similar to the Loc. 43, 44 and 49 from Baode by faunal composition, confirming their age to be ~7.0 Ma rather than 5.5 Ma. The high similarity with the fossils from Wulanhua, Siziwang Banner, Nei Mongol, verified the age of Wulanhua fauna at about 7 Ma. Compared with faunas from the Linxia Basin, Gansu, it is more similar to those from the upper part of the Liushu Formation, especially the Yangjiashan fauna. Based on the faunal composition and their tooth morphology, the Hanjiaying fauna could be included in the “Gazella dorcadoides” fauna, which is supposed to be at the west paleobiome in northern China during the Late Miocene.
Keywords:
PDF (3422KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
王倩, 刘艳, 王李花, 傅明楷, 张兆群. 内蒙古韩家营玄武岩夹层中三趾马动物群化石. 古脊椎动物学报[J], 2021, 59(2): 125-137 DOI:10.19615/j.cnki.1000-3118.210323
WANG Qian, LIU Yan, WANG Li-Hua, Mikael FORTELIUS, ZHANG Zhao-Qun.
Neogene terrestrial depositions in China are well developed and produced a great number of mammal fossils. Among them, the Late Miocene Hipparion faunas have long been extensively explored (Schlosser, 1924) and researchers have documented the highest diversification and richness of mammal fossils with more than 140 genera during the Neogene (Zhang et al., 2006). Due to lack of absolute dating materials, the sequencing and biochronology of the faunas were mainly inferred by evolutionary levels of mammals and magnetostratigraphic correlation (Tedford et al., 1991). The discontinuity of the fossil records and short depositional sequences weakened the biochronologic interpretations, hence absolute dating constraints are badly needed. Recently we found a fossil rich locality near Hanjiaying Village, Jining District, Nei Mongol. The fossil horizon is located right in between two basalt layers, which provides ideal conditions for the precise dating of the Hipparion fauna. Detailed analysis of the fauna will improve our understanding of the biochronology of Chinese Late Miocene.
Institutional abbreviation IVPP V or V, specimen prefix of the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China.
1 Geological settings
The fossil locality is situated at the hillside west of Hanjiaying village, about 18 km west of Jining District, Wulanchabu City, Nei Mongol (41°00.825′ N, 112°53.234′ E) (Fig. 1A). Fossils are produced from orange-red mudstone and silty mudstone, and the strata are basically horizontal. The stratigraphic section (Fig. 1B) is described from top to bottom as following:Fig. 1
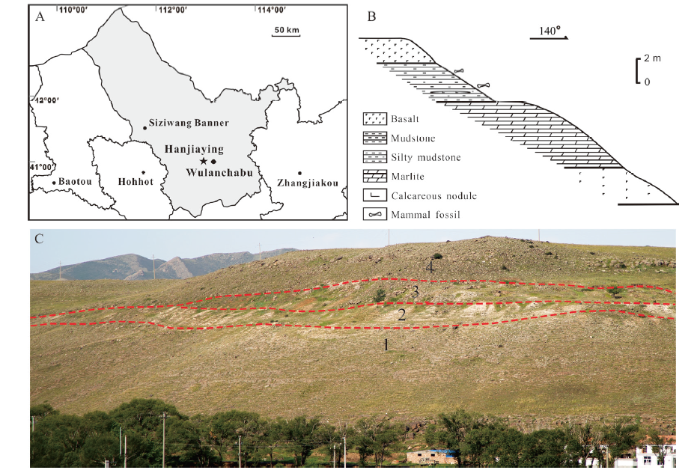 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1The geographic location and stratigraphic profile of the Hanjiaying locality
A. location of fossil site (star) in Wulanchabu City, Nei Mongol; B. stratigraphic section of the Hanjiaying fossil locality; C. the photograph of Hanjiaying profile: 1. dark grey basalt layer; 2. grayish-white marlite layer; 3. orange-red mudstone layer; 4. caesious basalt layer
4. Caesious, purplish-grey fine-grained basalt with vasicular and amygdaloidal structure, bedded with multiple eruptive cycles ~15 m
3. Orange-red mudstone. Fossils are fragmentary and fragile. Orange-red mudstone with fine sand, silt and calcareous nodules. Fossils are well preserved. Yellow, green mudstone 3.25 m
2. Grayish-white marlite, massive or nodular structure 5.47 m
1. Dark grey basalt with vasicular and amygdaloidal structure ~29 m
The basalt samples from the bottom first layer and the topmost fourth layer were tested in the K-Ar laboratory of the Institute of Geology, China Earthquake Administration (Table 1). The age of the Hanjiaying fossil locality is 7.2-6.8 Ma, corresponding to MN 12 (8.0-6.6 Ma ) (Steininger, 1999) of the European land mammal age, which was then near the boundary of the Bahean and Baodean age of the Late Miocene (Zhang et al., 2013).
Table 1
Table 1K-Ar isotopic dating data of the basalt from the Hanjiaying fossil locality in Jining District
| Sample number | K (%) | Sample weight (mg) | 40Ar (10-11mol/g) | 38Ar (10-14mol) | 40Ar/38Ar | 38Ar/36Ar | Age (Ma) |
|---|---|---|---|---|---|---|---|
| HJY-01 | 1.60 | 31.40 | 2.014 | 8.087 | 11.22 | 86.31 | 7.24±0.19 |
| HJY-02 | 1.11 | 33.50 | 1.322 | 8.095 | 8.661 | 92.14 | 6.85±0.21 |
新窗口打开|下载CSV
2 Mammalian fossils
Fossils collected from the Hanjiaying locality represent 12 species in total: Amphi-machairodus giganteus, Rhinocerotinae gen. et sp. indet., Aceratheriinae gen. et sp. indet., Hipparion coelophyes, Palaeotragus microdon, Samotherium sp., Dorcadoryx palaeosinensis, Gazella sp., Oioceros sp., Bovidae gen. et sp. indet., Ochotona lagreli, and Erinaceidae gen. et sp. indet.Most of the above-mentioned mammals are the same as or close to the members of the Baode Fauna, such as Amphimachairodus giganteus, Hipparion coelophyes, Palaeotragus microdon, Samotherium sp., Dorcadoryx palaeosinensis, and Gazella sp. Amphimachairodus lived from the Late Miocene to the Pleistocene (Liu, 2004). Hipparion coelophyes lived in the Baodean in North China (Liu, 2013). Dorcadoryx lived from the Late Miocene to the Early Pliocene (MN 11-14) in Mongolia and North China, and Gazella appeared in the early Late Miocene (MN 9) and was widely distributed in North China in the late Late Miocene (Baodean) (Chen and Zhang, 2009).
The main characters of the representative taxa from the Hanjiaying fauna are described as following.
Carnivora, Felidae Amphimachairodus giganteus Kretzoi, 1929: There found a juvenile skull fragment (IVPP V 27533.1) and a partial right P4 (V 27533.2) (Fig. 2). The infraorbital foramen is large and round; the diastema of I3 and C is short; the I2 cone is shaped, serrated at the medial and lateral edges, with two lingual styles and a cingulum in between them; the upper canine is laterally flat and serrated at the anterior and posterior edges; there is a parastyle anterior to the main cusp on the P3, with serrated crista; the P4 is long and narrow, with two cusps increasingly larger anterior to the main cusp.
Fig. 2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Photographs of Amphimachairodus giganteus from Hanjiaying, Nei Mongol
A. fragmentary skull (IVPP V 27533.1); B. fragmentary right P4 (V 27533.2)
A1, B1. labial view; A2, B2. occlusal view; B3. lingual view
Perssodactyla, Equidae Hipparion coelophyes Sefve, 1927: The materials include three mandibles (IVPP V 27534.1-3), four maxillae (V 27534.4-7) and isolated teeth (V 27534.8-75) (Fig. 3). The cheek tooth row is about 120-140 mm in length; the average height of the crown is about 53 mm (Tables 2, 3). It is relatively small in size; the lingual wall of incisors is weakly grooved; the protocone is moderately long with pointed at ends when there is little wear, and it is labially rounded and lingually flattened; the plications are moderate to strong, with bifid pli caballins and sometimes single or complex; the hypoconal groove is deep, and the hypoconal constriction is present when not significantly worn; the labial wall of the lower teeth at the protoconid and hypoconid is flattened; the metaconid is rounded and the metastylid is subangular; the protostylid and pli caballinid are developed; the hypoconulid of the m3 is bifid and hammar-shape with a narrow neck when deeply worn.
Fig. 3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Photographs of Hipparion coelophyes from Hanjiaying, Nei Mongol
A. mandibles (IVPP V 27534.1); B. fragmentary left maxilla (V 27534.4); C. right M1/2 (V 27534.30); D. left p2-3 (V 27534.8-9); E. right p3/4 (V 27534.10); F. right m1/2 (V 27534.11)
A1, B1, C1, D1, E1, F1. occlusal view; A2, B2, C2, D2, E2, F2. labial view
Table 2
Table 2Measurements of upper teeth of Hipparion coelophyes from Hanjiaying (mm)
| V 27534.4 | V 27534.5 | V 27534.6 | V 27534.30 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | W | L | W | L | W | L | W | |||||
| P2 | 27.7 | 20.8 | 26.6 | 19.7 | ||||||||
| Pr | 5.7 | 4.0 | 8.4 | 5.8 | ||||||||
| P3 | 22.7 | 22.1 | 19.4 | 20.6 | 17.9 | 19.6 | ||||||
| Pr | 6.6 | 4.0 | 7.1 | 4.7 | 7.4 | 4.6 | ||||||
| P4 | 21.7 | 19.8 | 19.5 | 21.7 | 19.3 | 22.5 | ||||||
| Pr | 7.2 | 4.1 | 7.5 | 4.7 | 7.4 | 4.4 | ||||||
| M1 | 19.8 | 15.0 | 17.0 | 20.9 | 17.8 | 21.6 | ||||||
| Pr | 6.5 | 3.2 | 7.9 | 5.1 | 7.5 | 4.6 | ||||||
| M2 | 19.0 | 19.0 | 17.3 | 20.3 | 18.5 | 20.2 | 25.4 | 22.8 | ||||
| Pr | 5.8 | 3.1 | 6.3 | 4.2 | 6.8 | 4.3 | 7.3 | 3.5 | ||||
| M3 | 20.7 | 18.2 | 21.5 | 18.3 | ||||||||
| Pr | 6.2 | 4.3 | 6.4 | 3.5 | ||||||||
新窗口打开|下载CSV
Table 3
Table 3Measurements of lower teeth of Hipparion coelophyes from Hanjiaying (mm)
| V 27534.1 | V 27534.3 | V 27534.2 | V 27534.8 | V 27534.9 | V 27534.10 | V 27534.11 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| left | right | L | W | L | W | L | W | L | W | L | W | L | W | |||||||||
| L | W | L | W | |||||||||||||||||||
| p2 | 24.2 | 13.3 | 24.4 | 13.5 | 25.2 | 12.2 | 29.9 | 13.7 | ||||||||||||||
| p3 | 23.0 | 14.2 | 22.4 | 14.2 | 19.4 | 13.1 | 24.6 | 14.5 | 24.2 | 15.8 | ||||||||||||
| p4 | 21.6 | 13.2 | 22.0 | 13.4 | 18.7 | 13.0 | 19.0 | 12.8 | ||||||||||||||
| m1 | 21.4 | 12.7 | 21.6 | 12.8 | 16.7 | 12.3 | 16.4 | 12.1 | 25.2 | 13.3 | ||||||||||||
| m2 | 20.3 | 11.7 | 20.9 | 11.8 | 20.8 | 11.5 | 18.9 | 11.4 | ||||||||||||||
| m3 | 21.8 | 9.8 | 21.3 | 9.7 | 24.8 | 10.0 | 24.9 | 9.8 | ||||||||||||||
| i1 | 13.7 | 8.8 | 13.8 | 8.7 | 11.7 | 9.7 | 11.4 | 9.0 | ||||||||||||||
| i2 | 14.5 | 8.0 | 14.4 | 8.2 | 11.9 | 8.9 | 11.9 | 9.0 | ||||||||||||||
| i3 | 12.9 | 7.3 | 12.6 | 7.3 | 11.5 | 7.7 | 11.4 | 7.5 | ||||||||||||||
新窗口打开|下载CSV
Artiodactyla, Giraffidae Palaeotragus microdon Koken, 1985: The species is represented by a right mandible fragment with m2-3 (IVPP V 27535.1) and isolate teeth (V 27535.2-3) (Fig. 4). It is a small sized Palaeotragus; the crown height is moderately hypsodont; the enamel has longitudinal grain; it is the simple structure on the M2 with separate protocone and hypocone, parastyle and mesostyle developed, median pillar absent, the measurement as 25.9 mm (length)×27.8 mm (width)×9.4 mm (height); there is a basal pillar between protoconid and hypoconid developed on m3, basal pillar prior to hypoconulid present, the measurement as 33.7 mm×16.8 mm×15.1 mm.
Fig. 4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Photographs of Giraffidae from Hanjiaying, Nei Mongol
A-C. Palaeotragus microdon: A. fragmentary right mandible with m2-3 (IVPP V 27535.1); B. right M2 (V 27535.2); C. right m3 (V 27535.3); D. Samotherium sp. fragmentary left mandible with p3-m2 (V 27536)
A1, B, C1, D1. occlusal view; A2, C2, D2. labial view
Samotherium sp.: The left mandible with p3-m2 (V 27536) (Fig. 4). The measurements are 25.0 mm×19.9 mm×18.8 mm on p3; 30.8 mm×24.8 mm×20.5 mm on p4; 34.9 mm× 30.6 mm×15.4 mm on m1 and 45.6 mm×25.3 mm×24.3 mm on m2. The crown height is moderately hypsodont; the lower cheek teeth lack cingulum, the metaconid and entoconid are lingually convex; the lower premolars are large with strong anterior and weak posterior lobes; the prefossette is enclosed and the parastylid is developed on p3 and p4; the hypoconid is weak and the entoconid is absent on p3; the hypoconid is developed and the entoconid is weak on p4; the parastylid and entostylid are developed on m2.Artiodactyla, Bovidae Dorcadoryx palaeosinensis Schlosser, 1903: The materials include a right mandible fragment with p2-m1, half m2 and m3 (IVPP V 27537.1), a left mandible fragment with p2-3 (V 27537.2) and isolated teeth (V 27537.3-5) (Fig. 5A, B). The measurements of IVPP V 27537.1 are 6.7 mm×4.2 mm×8.7 mm on p2; 8.9 mm×6.0 mm×10.8 mm on p3; 11.4 mm×6.6 mm×11.3 mm on p4; 11.9 mm×8.6 mm×11.8 mm on m1 and 18.9 mm×8.2 mm× 13.4 mm on m3, and the measurements of V 27537.3 are 14.5 mm×13.9 mm×16.7 mm on M2. The mandible is strongly built; the cheek teeth are high crowned; the premolar row is short in relation to the length of molar row; the styles on the upper molar are developed, the anterolabial rib is distinct, and the lingual basal pillar is weak; the paraconid on the p4 is not fused with the metaconid.
Fig. 5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Photographs of Bovidae from Hanjiaying, Nei Mongol
A, B. Dorcadoryx palaeosinensis: A. fragmentary right mandible with p2-m1, half m2, m3 (IVPP V 27537.1); B. left M2 (V 27537.3); C-G. Gazella sp: C. fragmentary right maxilla with P2-M2 (V 27538.1); D. fragmentary right mandible with p2-m3 (V 27538.3); E. fragmentary left mandible with p3-m3 (V 27538.4); F-G. horn cores (V 27538.15-16); H-K. Oioceros sp. horn cores (V 27539.1-4); L. Bovidae gen. et sp. indet. horn core (V 27540)
A1, B1, C1, D1, E1. labial view; A2, B2, C2, D2, E2. occlusal view
Gazella sp.: The materials include two maxillae (V 27538.1-2), seven mandibles (V 27538.3-9), isolated teeth (V 27538.10-14), and some horn cores (V 27538.15-23) (Fig. 5C-G; Table 4). It is relatively small in size; the supraorbital foramen is small; the average total length of the horn cores is 101 mm, with no torsion and they are curved backwards and diverge outwards; the slightly elliptical cross section measures 27-28 mm (anteroposterior diameter)× 24-26 mm (transverse diameter) at the base; the cheek teeth crown has a high crown; the styles on upper molar are developed, and the ribs are weak; the lower premolar row (20.3-22.0 mm) / molar row (36.6-40.2 mm) ratio is about 53%-58%; the anterior and posterior valley of the p4 are unclosed; the labial basal pillar is developed on the lower molars.
Table 4
Table 4Measurements of upper and lower teeth of Gazella sp. from Hanjiaying (mm)
| V 27538.1 | P2 | P3 | P4 | M1 | M2 | |
|---|---|---|---|---|---|---|
| L | 8.4 | 7.5 | 7.3 | 9.6 | 11.4 | |
| W | 5.7 | 6.7 | 8.5 | 10.5 | 11.0 | |
| V 27538.3 | p2 | p3 | p4 | m1 | m2 | m3 |
| L | 4.8 | 7.5 | 8.9 | 9.7 | 11.3 | 17.2 |
| W | 3.5 | 4.4 | 5.3 | 7 | 7.5 | 7.6 |
| V 27538.4 | p3 | p4 | m1 | m2 | m3 | |
| L | 7.8 | 9.3 | 8.9 | 11.1 | 17.0 | |
| W | 4.4 | 5.0 | 7.7 | 8.2 | 7.4 |
新窗口打开|下载CSV
Oioceros sp.: The materials include six horn core fragments (V 27539.1-6) (Fig. 5H-K). They are relatively small in size; the horn cores have torsion of half circuits, inclined backwards, and are elliptical in cross section measuring 18.9-20.1 mm×14.6-15.8 mm at the base.
Bovidae gen. et sp. indet.: The materials include a horn core (V 27540) (Fig. 5L). The horn core has a weak torsion with posterolateral curvature and is elliptical in cross section. It measures 22.2 mm×16.5 mm at the base.
Lagomorpha, Ochotonidae Ochotona lagreli Schlosser, 1924: The materials include two mandible fragments (IVPP V 27541.1-2) (Fig. 6). The occlusal surface of the p3 is subtriangular in shape, measures 1.8 mm in length and 2.0 mm in width, the anteroconid is nearly rhombus, the anterolabial and anterolingual folds are about equal in depth and closely connected by a narrow bridge, the posterolabial fold posterolingually is about half the width of the talonid, and the lingual edge of the talonid is straight. The p4, m1 and m2 are very similar in morphology; the trigonid and talonid are nearly the same oval shape, and only the m1 and m2 have a shallow depression on anterolabial side of the two lobes. The m3 is short with only one lobe and approximately parallelogram-shaped.
Fig. 6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Photographs of Ochotona lagreli in occlusal view from Hanjiaying, Nei Mongol
A. fragmentary left mandible with p3-m1 (IVPP V 27541.1); B. fragmentary right mandible with m2-3 (V 27541.2)
3 Comparison with other Hipparion faunas from North China
By the faunal composition, the Hanjiaying fauna is a typical Hipparion fauna. There recorded a species of the Felidae. This kind of saber tooth cat is widely recorded in Eurasia during the Late Miocene. There is one species of three-toed horse, two species of the Giraffidae and four species of the Bovidae. The Rhinocerotidae are also richly represented, unfortunately no diagnostic anatomical parts were preserved. Pikas are also very common elements from many Late Miocene localities. However, no deer or pigs, which are also common in many Hipparion faunas, have been found from this locality.Compared with mammals from locality 30 (~5.7 Ma), localities 43, 44 and 49 (~6.5-7.0 Ma) of Baode, Shanxi, the representative localites of the Hipparion fauna (Kaakinen et al., 2013), the Hanjiaying fauna has the same species Amphimachairodus giganteus and Palaeotragus microdon, the same genera Gazella and Dorcadoryx. Brachydont Honanotherium schlosseri appears at locality 49 of Baode, while mesodont Samotherium appears at locality 30 and Hanjiaying fauna (Kaakinen et al., 2013). Hipparion coelophyes has only been recorded in localities 43 and 44. H. plocodus has only been found in locality 49. H. forstenae and H. dermatorhinum both appear at locality 30 (Bernor et al., 1990).
Compared with the Wulanhua fauna (~7.0 Ma) from Siziwang Banner of Nei Mongol (Deng et al., 2011), the Hanjiaying fauna has the same species Palaeotragus microdon and Dorcadoryx palaeosinensis, and the same genera Amphimachairodus, Samotherium and Gazella. Without Hipparion coelophyes, the Wulanhua fauna has H. teilhardi instead, which shows some primitive characters, such as the strong crown curvature of the upper tooth crown and the rounded rectangle shape of the protocone. Otherwise Sinohippus robustus, Chilotherium anderssoni, Cervavitus novorossiae, Gazella gaudryi and Plesiaddax depereti only appear in the Wulanhua fauna.
The Hanjiaying fauna has the same species Palaeotragus microdon and the same genera Dorcadoryx, Gazella and Ochotona as the Bahe fauna (11-7 Ma), but these only appear at the top of the Bahe Formation; the same species Palaeotragus microdon, and the same genus Gazella and Hipparion as the Lantian fauna (~6.6 Ma) from Lantian, Shaanxi (Liu et al., 1978; Zhang et al., 2002, 2013; Liu, 2013). The Bahe fauna has Hipparion weihoense, H. chiai, Dorcadoryx orientalis, Schansitherium, Shaanxispira, Protoryx, Tetralophodon, which are not present in the Hanjiaying fauna. The two species of hipparionine from the Bahe fauna suggest an earlier age than that of the Hanjiaying fauna. Also, Dorcadoryx orientalis from the Bahe fauna may be an early form of the genus Dorcadoryx (Chen, 2005). Compared to the Lantian fauna, Samotherium, Dorcadoryx and Oioceros were recovered from the Hanjiaying fauna. The Lantian fuana contains a large number of deer and pig materials (Zhang et al., 2013).
Compared with faunas from the Liushu Formation of the Linxia Basin in Gansu (Deng et al., 2004a, 2013), the Hanjiaying fauna has the same species Amphimachairodus giganteus, Hipparion coelophyes and Palaeotragus microdon, and genus Gazella as the Yangjiashan fauna, the upper part of the Liushu Formation (~8 Ma). It does not share any members with the Qingbushan fauna, the top of the Liushu Formation (~6.5 Ma). Compare to the Qingbushan fauna, the Yangjiashan fauna shows a closer similarity with the Hanjiaying fauna.
The Hanjiaying fauna has the same genera of Hipparion, Palaeotragus, Samotherium and Gazella as the Lamagou fauna (~7.8 Ma) from Fugu, Shaanxi; the same genera Hipparion, Samotherium, and Gazella as the Taohuapo fauna (~6.7 Ma) from Lingtai County, Gansu (Xue et al., 1995, 2006). Among them, Hipparion chiai from the Lamagou fauna shows its earlier age. Except for the genera shared with the Hanjiaying fauna, others from the Taohuapo fauna are very different.
4 Discussion
Based on the distribution of Chinese Neogene mammalian faunas, Early Miocene and Middle Miocene faunas are ecologically different in a west-east direction, with dense forests in eastern areas contrasted with open grasslands in western areas. However, since the early Late Miocene, the dense eastern forests were sparser and many had turned into woodlands, so the paleobiome boundary between eastern and western China was not as easy to distinguish (Tong et al., 1996; Qiu and Li, 2004; Deng et al., 2011). The later Late Miocene shows the apparent differentiation of a humid and closed fauna in the east and a dry and open fauna in the west (Kurtén, 1952; Zhang, 2006). The components of the Hanjiaying fauna, with Amphimachairodus giganteus, Palaeotragus microdon, Samotherium etc., show that this fauna should belong to the Gazella dorcadoides fauna, which were distributed at the west part of northern China (Kurtén, 1952).The Hipparion red clay from the Baode area of Shanxi, which produced the classical Baode fauna, has long been investigated and studied. Based on detailed stratigraphic work, Kaakinen et al. (2013) concluded an ecological boundary shifting within the time-space continuum, contrary to the temporal or spatial explanation of the faunal differences for the Baode localities. The Baode area was then located at the east of paleobiome boundary at ~7.0 Ma represented by locality 49, at west of the boundary at ~5.7 Ma represented by locality 30 due to the shifting of the eastern boundary (Kaakinen et al., 2013). Located northeastern to Baode, the Jining area should have been in the eastern ecolological regime according the northeast-southwest ecological gradient suggested by isotopic data (Passey et al., 2009) at ~7 Ma. The higher latitude and high elevation of the Jining area, which is in the Mongolian Plateau, may explain its western ecological characteristics. The high similarity of the faunal composition at the locality 49 and the Hanjiaying locality verifies the geological age of locality 49 at about 7.0 Ma, rather than 5.5 Ma as previously thought (Deng et al., 2004b).
Based on the faunal components, Deng et al. (2011) inferred the Wulanhua fauna from Siziwang Banner of Nei Mongol at about 7 Ma. This region was a typical steppe and is located to the west of the paleobiome boundary between eastern and western China during the Late Miocene. The Wulanhua fauna has many similarities with the Hanjiaying fauna, which confirms its age estimation at about 7 Ma. These two localities are close geographically and all belong to the western faunas.
The age of the Hanjiaying fauna is between those of the Bahe (11-7 Ma) and Lantian faunas (~6.6 Ma) (Zhang et al., 2002). The components of the Hanjiaying fauna are not significantly similar to the Bahe and Lantian fauna, which corresponds to the habitat differences because of the long distance between Jining and Lantian geographically.
The Hanjiaying fauna is closer to the Yangjiashan fauna (~8 Ma) than is it to the Qingbushan fauna (~6.5 Ma) from Linxia Basin, Gansu (Deng et al., 2013). This phenomenon shows there are large differences between the members of the MN12 and MN13 in North China faunas.
There are few similarities between the Hanjiaying fauna and the Lamagou fauna from Fugu, Shaanxi, and the Taohuapo fauna from Lingtai, Gansu (Xue et al., 1995, 2006). This may correspond to the differences of their age and location respectively.
5 Conclusion
Hipparion faunas first occurred during the early Late Miocene and lived through the whole Late Miocene, but the numerical dating is still in its infancy. The discovery of the Hanjiaying fauna helped fill this gap in the timeline and provides strong evidence for the paleomagnetic dating of other faunas. By K-Ar isotopic dating of the basalt layers it is sandwiched in between, the age of the Hanjiaying fauna is 7.2-6.8 Ma. The Hanjiaying fauna is a typical Hipparion fauna, dominated by Hipparion and Gazella, but does not include the Hyaenidae and Cervidae. There are some other faunas, like those in Baode localities 43, 44, 49 and the Wulanhua fauna, which have members similar to the Hanjiaying fauna. Based on the taxa and their tooth pattern, the Hanjiaying fauna should belong to the Gazella dorcadoides fauna, located west of the boundary between the eastern and western ecological regimes in North China during the Late Miocene.参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 3]
[本文引用: 2]
[本文引用: 4]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 3]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}