 ,1,2, 刘文晖1,2,3, 白炜鹏1,2,3
,1,2, 刘文晖1,2,3, 白炜鹏1,2,3Cladistic approach on chronological relationship of the Pleistocene mammalian faunas from China
DONG Wei,1,2, LIU Wen-Hui1,2,3, BAI Wei-Peng1,2,3收稿日期:2019-01-17网络出版日期:2020-01-20
| 基金资助: |
Received:2019-01-17Online:2020-01-20
作者简介 About authors
dongwei@ivpp.ac.cn

摘要
在中国境内有很多地点产出更新世哺乳动物群,为哺乳动物演化、生物年代学、古生态学、古环境学、古动物地理学等的研究提供了很好的材料。在生物年代学方面,动物群组合面貌和绝灭率曾经是用来推断动物群年龄的方法。在此基础上又发展出根据Brainerd-Robinson法则对动物群的二元相似性系数进行排序,通过二元相似性系数确定动物群的古老系数判断动物群的年代顺序等方法。而二元相似性系数的确定取决于一个种类在一个动物群中的存在与否,类似于支序系统学中性状矩阵的建立是根据一个特征在一个种类中存在与否那样。因此本文尝试使用支序系统学方法来研究动物群之间的演化关系,选择了一些经过二元相似性系数排序的动物群进行支序系统学分析,将这些结果与使用其他生物年代学方法得出的结果进行比较,结合一些动物群绝对年龄的测年成果,对那些尚未进行绝对年龄测年的动物群进行年龄推断,得出的绝对年龄如下:辽宁大连的古龙山动物群为16~20 ka; 辽宁本溪的山城子动物群为20~30 ka; 安徽东至人遗址的华龙洞动物群为150~400 ka; 河南南召人遗址的杏花山动物群为150~400 ka; 陕西洛南人遗址的东河动物群为500~700 ka; 湖北郧西人遗址的白龙洞动物群为500~850 ka; 湖北郧县的梅铺动物群为500~850 ka; 广西田东人遗址的么会洞动物群为1.2~1.8 Ma; 广西柳城巨猿洞动物群为1.2~1.5 Ma; 内蒙古赤峰初头朗动物群为1.6~1.9 Ma; 安徽繁昌人字洞动物群为1.9~2.4 Ma。
关键词:
Abstract
There are many localities yielding the Pleistocene mammalian faunas in China. It offers excellent material for the study of mammalian evolution, biochronology, paleoecology, paleoenvironment, paleozoogeography, etc. Faunal assemblage characters and taxonomic extinction rates were widely used for determining the faunal ages in biochronology. Faunal binary similarity coefficients sequenced according to Brainerd-Robinson’s rule and antiquity coefficients were further developed methods in biochronology for dating the faunal ages. The faunal binary similarity coefficients are based on the presence or absence of a taxon in a fauna. It is similar to the presence or absence of a character of a species in cladistic analyses for phylogeny, and all faunas have a special ancestor-descendant relationship. The present work is an attempt to find the relationship of the faunas with cladistic methods by selecting three groups of faunas sequenced by faunal binary similarity coefficients according to Brainerd-Robinson’s rule and antiquity coefficients, to compare the results with different methods, and then to estimate the ages of the faunas not yet dated by physical or chemical methods. The estimations are as follow: Gulongshan in Dalian, Liaoning Province: 16-20 ka; Shanchengzi at Benxi, Liaoning Province: 20-30 ka; Hualongdong at Dongzhi Man site, Anhui Province: 150-400 ka; Xinghuashan at Nanzhao Man site, Henan Province: 150-400 ka; Donghe at Luonan Man site, Shaanxi Province: 500-700 ka; Bailongdong at Yunxi Man site, Hubei Province: 500-850 ka; Meipu at Yunxian, Hubei Province: 500-850 ka; Mohui at Tiandong Man site, Guangxi Autonomous Region: 1.2-1.8 Ma; Juyuandong at Liucheng, Guangxi Autonomous Region: 1.2-1.5 Ma; Chutoulang at Chifeng, Nei Mongol Autonomous Region: 1.6-1.9 Ma; Renzidong at Fanchang, Anhui Province: 1.9-2.4 Ma.
Keywords:
PDF (576KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
董为, 刘文晖, 白炜鹏. 中国境内部分更新世哺乳动物群的支序系统学分析及生物年代学推断. 古脊椎动物学报[J], 2020, 58(1): 67-81 DOI:10.19615/j.cnki.1000-3118.190525
DONG Wei, LIU Wen-Hui, BAI Wei-Peng.
1 Introduction
Cladistics is a method of hierarchical analysis in which a branching diagram or cladogram is used to group entities as nested sets based on special common similarities (Young, 1986). It is widely used for searching phylogenetic relationship of given groups of organisms (Hennig, 1965; Ashlock, 1974; Nelson and Platnick, 1991; de Pinna, 1991). It is also used in other disciplines other than phylogeny of organisms, such as reconstruction of ancient continents evolution (Young, 1986), determination of the ages of divergent nodes of different clades (Ashlock, 1974). In short, any group of individuals that can be hypothesized to have a common ancestor, and to which a set of shared characteristics may or may not apply, can be compared in pairs (Platnick and Cameron, 1977). Thus, cladistics is also used in textual, linguistic, comparative mythology and folktale (Tehrani, 2013; d'Huy, 2013; Ross et al., 2013), stemmatology (Robinson and O’Hara, 1996; Kapitan, 2017), and even in astrophysics (Fraix-Burnet et al., 2006).The Pleistocene mammalian faunas are very numerous in China and they offer very useful material for the study of mammalian evolution, biochronology, paleoecology, paleoenvironment, paleozoogeography, and so on. Some faunas have been dated with physical or chemical methods for absolute ages, but some others have not yet and in need of dating. During the study of the mammalian fauna from Xiaogushan site (Dong et al., 2010), the faunal binary similarity coefficients sequenced according to Brainerd-Robinson’s rule, a method proposed by Chen (1983, 2005), was introduced to find the relationship between Xiaogushan fauna and some selected faunas, especially for biochronological sequence of the faunas. The method was developed further into antiquity coefficients to find the biochronological sequence of the faunas during the establishment of biochronological framework of Homo erectus horizons in China (Dong, 2016) and the study of the Early Pleistocene fauna from Chutoulang in northeastern China (Dong et al., 2017). The faunal binary similarity coefficients are based on the presence or absence of a taxon in a fauna. It is similar to the presence or absence of a character of a species in cladistic analyses for phylogeny in mathematic sense, and all faunas have their special ancestor-descendant relationship. It is therefore possible to use cladistic methods to find the affinity of the faunas and to interpret biochronological sequences, zoogeographical boundaries and paleoenvironmental evolution of the faunas. Here we show a test of application of cladistic methods to find the relationship of three groups of the Pleistocene faunas and to learn the effectiveness of the methods, and give our estimated ages on the faunas not yet dated by physical or chemical methods.
2 Material and methods
Three groups of the Pleistocene mammalian faunas analyzed by binary similarity coefficients in above mentioned three studies (Dong et al., 2010, 2017; Dong, 2016) were selected here again for cladistic analyses.The first group includes Xiaogushan fauna (Dong et al., 2010) and nine faunas with comparable geographic and chronological significances, e.g. Gulongshan fauna (Zhou et al., 1990) from Dalian, Liaoning; Miaohoushan fauna (Zhang et al., 1986) and Shanchengzi fauna (Zhang et al., 1986) from Benxi, Liaoning; Yushu fauna (IVPP, 1959) from Yushu area, Jilin; Yanjiagang fauna (NET et al., 1987) from Haerbin, Heilongjiang; Dingcun fauna (Pei, 1958) from Xiangfen, Shanxi; Upper Cave fauna (Pei, 1940) from Zhoukoudian, Beijing; Lingjing fauna (Li and Dong, 2007; Zhang et al., 2009) from Xuchang, Henan; and Salawusu (Sjara-osso-gol) fauna (Qi, 1975; Tong et al., 2008) from Erdos, Nei Mongol. The ages of the faunas in this group range in the Late Pleistocene except that of Miaohoushan which is of the late stage of the Middle Pleistocene. The selected faunas total 10 and referred taxa 116 (Appendix I).
The second group are faunas from Homo erectus horizons in China, e.g. Danawu horizon from Yuanmou Man site (Lin et al., 1978), Mohui horizon from Tiandong (Wang et al., 2007), Gongwangling horizon from Lantian Man site (Hu and Qi, 1978), Longgudong horizon from Jianshi Man site (Zheng, 2004), Quyuanhekou horizon from Yunxian Man site (Ji, 1999), Meipu horizon from Yunxian (Xu, 1978), Donghe horizon from Luonan (Xue et al., 1999), Bailongdong horizon from Yunxi Man site (Wu et al., 2009), Longtandong horizon from Hexian Man site (Huang et al., 2012), Chenjiawo horizon from Lantian (Qi, 1989), Qizianshan horizon from Yiyuan Man site (Lü et al., 1989), Xinghuashan horizon from Nanzhao Man site (Qiu et al., 1982), Huludong horizon from Nanjing Man site (Wu et al., 2002) and Hualongdong horizon from Dongzhi Man site (Gong et al., 2014). Since there are 9 layers yielding Homo erectus material (Li and Ji, 1981) and the hominin horizons at Locality 1 of Zhoukoudian Site were defined according to three cycles of sedimentations related to environmental changes (Xu et al., 1999), Layers 8-11, Layers 6-7 and Layers 3-5 were considered as lower, middle and upper Homo erectus horizons respectively. The ages of the faunas in this group range from the middle stage of the Early Pleistocene to the middle stage of the Middle Pleistocene. Because some faunas were collected with screen washing technique for small mammals, but the others not, and in order to reach balanced results we just select the taxa of large mammals from these faunas. The faunas from all Homo erectus horizons from China total 17 and referred taxa 123 (Appendix II).
The third group are all Early Pleistocene faunas with the number of the taxa above 30 from northeastern, northern, northwestern, central and southern parts of China such as Chutoulang (Zhang, 1989; You and Zhang, 1989; Dong et al., 2017), Nihewan (s.s.) (Teilhard de Chardin and Piveteau, 1930; Qiu, 2000; Tong et al., 2011), Gongwangling (Hu and Qi, 1978), Longdan (Qiu et al., 2004), Renzidong (Jin and Liu, 2009), Longgupo (Huang et al., 1991), Longgudong (Zheng, 2004), Juyuandong (Gigantopithecus Cave) (Pei, 1987; Han, 1987; Han and Xu, 1989) and Dadong (Jin et al., 2009). A French fauna from upper Saint-Vallier (LD3) about 1.5 km northeast of Saint-Vallier Village (45°11'16.94"N, 4°49'47.69"E, 360 m above sea level) (Guérin et al., 2004) was also selected as a classic European Villafranchian fauna for comparison. The ages of the faunas in this group range from the early stage of the Early Pleistocene to the middle stage of the Early Pleistocene. The selected faunas total 10 and referred taxa 317, small mammals included (Appendix III).
All three groups were then gathered together as a single assembled group ranging from the early stage of the Early Pleistocene to the late stage of the Late Pleistocene for cladistic analyses. Since the faunas from Gongwangling and Longgudong are present in both the second and third groups, the faunas in assembled group total 35 and referred taxa 418, small mammals included (Appendix IV).
All extinct taxa, including regional extinct ones, were considered as a presumed archaic fauna and those of extant taxa as a presumed modern fauna in every group.
The presence or absence of a species, including conformis species, in a fauna is considered as equivalent to the presence or absence of a character in a taxon in phylogenetic cladistic analyses and marked by “1” (present) or “0” (absent) in the data matrices in the present work. The question mark “?” in the data matrix indicates that the taxon of the fauna is likely close to the determined species, and it is equivalent to the missing character in phylogenetic cladistic analyses.
The results of cladistic analyses were then compared with those of binary faunal similarity coefficients sequenced according to Brainerd-Robinson’s rule (Chen, 1983, 2005) and antiquity coefficients (Dong, 2016), extinction rates, as well as dated ages, to test the effectiveness of the analyses on faunas and estimate the ages of the faunas not yet dated by any physical or chemical methods.
3 Results of cladistic analyses
For the first group of the faunas, the heuristic tree search tried 902 rearrange-ments and found 244 best trees in which one optimal tree with 10 nodes was retained, in which 9 nodes have dichotomous branches and only one has three (Fig. 1).Fig. 1
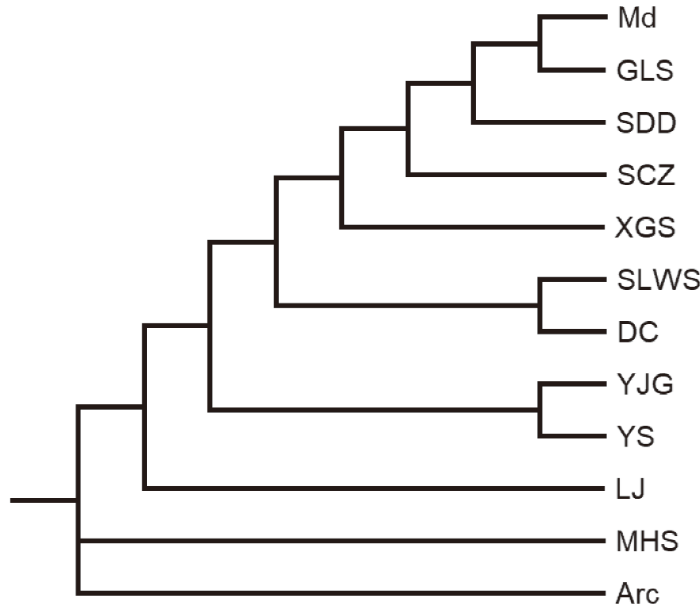 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Optimal tree for selected Late Pleistocene mammalian faunas from China
Arc. Presumed archaic fauna; DC. Dingcun at Xiangfen; GLS. Gulongshan in Dalian; LJ. Lingjing in Xuchang; Md. Presumed modern fauna; MHS. Miaohoushan at Benxi; SCZ. Shanchengzi at Benxi; SDD. Upper Cave at Zhoukoudian; SLWS. Salawusu (Sjara-osso-gol) in Ordos;XGS. Xiaogushan at Haicheng; YJG. Yanjiagang in Harbin; YS. Yushu at Yushu area
For the second group of the faunas, the heuristic tree search tried 48498 rearrangements and found 275 best trees in which 10 optimal trees were retained. The strict and semi-strict consensus trees of the retained optimal trees are the same. Both of them have 11 nodes with dichotomous branches, one node with three branches and another with four (Fig. 2, right). The Adams consensus tree has some minor differences from the strict and semi-strict consensus trees. It has two nodes with three branches and the rest 14 nodes have dichotomous branches (Fig. 2, left).
Fig. 2
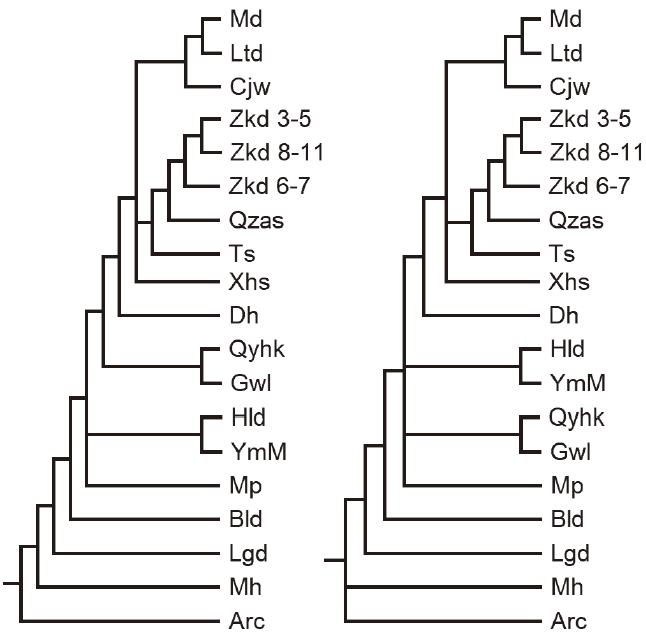 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Adams (left) and strict consensus (right) trees for the faunas from Homo erectus horizons in China
Arc. Presumed archaic fauna; Bld. Yunxi Man locality at Bailongdong; Cjw. Lantian Man locality at Chenjiawo; Dh. Luonan Man locality at Donghe; Gwl. Lantian Man locality at Gongwangling; Hld. Dongzhi Man locality at Hualongdong; Lgd. Jianshi Man locality at Longgudong; Ltd. Hexian Man locality at Longtandong; Md. Presumed modern fauna; Mh. Tiandong Man locality at Mohui Cave; Mp. Yunxian Man locality at Meipu; Qyhk. Yunxian Man locality at Quyuanhekou; Qzas. Yiyuan Man localities at Qizianshan; Ts. Nanjing Man locality at Huludong; Xhs. Nanzhao Man locality at Xinghuashan; YmM. Yuanmou Man locality at Danawu; Zkd 3-5, Zkd 6-7, Zkd 8-11. Layers 3-5, 6-7, 8-11 at Locality 1 of Zhoukoudian Peking Man Site
For the third group of the faunas, the heuristic tree search tried 3737 rearrange-ments and found 620 best trees in which seven optimal trees were retained. The strict and semi-strict consensus trees of the seven optimal trees retained are the same, each tree has only three nodes: the first one has three branches, the second one has 9 branches and only the last one has dichotomous branches (Fig. 3, right). The Adams consensus tree is more distinct. It has seven nodes, in which four nodes have dichotomous branches, two nodes have three branches and one has four (Fig. 3, left).
Fig. 3
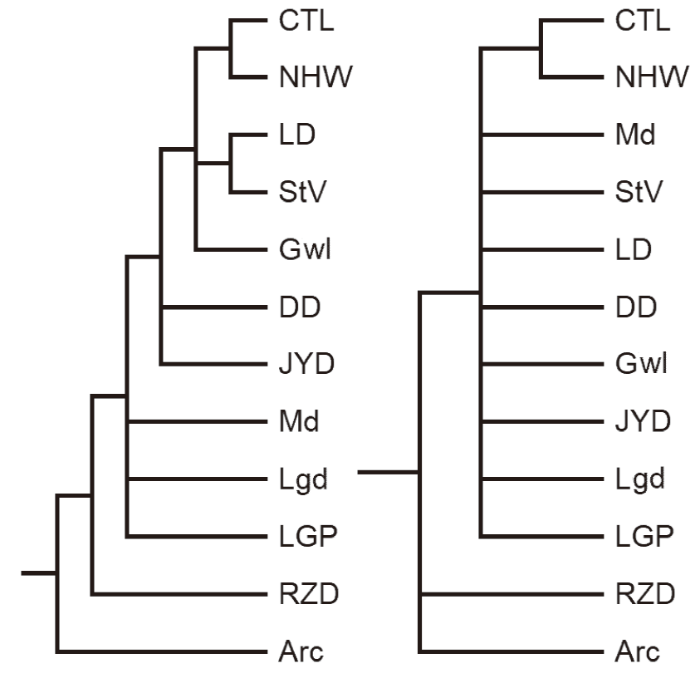 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Adams (left) and strict consensus (right) trees for selected Early Pleistocene mammalian faunas from China
Arc. Presumed archaic fauna; CTL. Chutoulang in Chifeng; DD. Dadong at Chongzuo; Gwl. Gongwangling at Lantian; JYD. Juyuandong (Gigantopithecus Cave) at Liucheng; LD. Longdan in Linxia Basin; Lgd. Longgudong at Jianshi; LGP. Longgupo at Wushan; Md. Presumed modern fauna; NHW. Xiashagou in Nihewan Basin; RZD. Renzidong at Fanchang; StV. Saint Vallier in France for comparison
For the assembled group of the faunas, the heuristic tree search tried 1452544 rearrange-ments and found 995 best trees in which 169 optimal trees were retained. The strict and semi-strict consensus trees of 169 trees are very similar to each other; the former has one more node with five branches compared with the latter. The semi-strict consensus tree has 25 nodes, in which 20 nodes have dichotomous branches, one has three, two have four and the other two have five (Fig. 4, right). The Adams consensus tree has 29 nodes, in which 24 nodes have dichotomous branches, three nodes have three branches and two have four (Fig. 4, left). Miaohoushan (MHS) fauna of the first group appears in the clade formed by many faunas from the second group, and some faunas from the second group, e.g. Mohui (Mh) and Longgudong (Lgd), appear on the stem formed by many faunas from the third group.
Fig. 4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Adams (left) and semi-strict consensus (right) trees for selected Pleistocene mammalian faunas from China
See Figs. 1-3 for abbreviations of the localities
4 Discussion
4.1 The first group of faunas
The result of cladistic analysis on the first group of the Pleistocene faunas is chronologically and geographically significant (Fig. 1; Table 1). The branches of the crown clade such as Xiaogushan (XGS), Shanchengzi (SCZ), Upper cave (SDD), Gulongshan (GLS) are geographically close to each other and chronologically related (Fig. 1). It is the same for the clade by Yanjiagang (YJG) and Yushu (YS). For the clade by Dingcun (DC) and Salawusu (SLWS), they are separated by a moderate distance of about 360 km, but chronologically very close to each other (Table 1). The result is very close to that by binary faunal similarity coefficients sequenced according to Brainerd-Robinson’s rule (Dong et al., 2010). It is also close to that by antiquity coefficients and extinction rates (Table 1). The cladistic analysis appears effective for this group of the faunas. Gulongshan and Shanchengzi faunas have not yet dated by any physical or chemical methods. Furthermore, the result of the analyses of the first group is also very close to that with assembled group. Based on the cladistic analyses, Gulongshan fauna is closest to presumed modern faunas (Figs. 1, 4), it is the same by binary faunal similarity coefficients sequenced according to Brainerd-Robinson’s rule (Dong et al., 2010). But it is slightly older than Upper Cave fauna by antiquity coefficients and extinction rates (Table 1). Shanchengzi fauna is placed on the stem between the branches of Xiaogushan and Upper Cave faunas by cladistic analyses (Fig. 1), in accordance with the results by binary faunal similarity coefficients sequenced according to Brainerd-Robinson’s rule and antiquity coefficients (Dong et al., 2010). Miaohoushan (MHS) fauna is close to the presumed archaic fauna of the first group (Fig. 1) or close to the Middle Pleistocene faunas (Fig. 4), and it is the oldest fauna in the first group with all methods (Table 1). Lingjing (LJ) fauna appears as the second oldest fauna in the first group with all methods (Fig. 1; Table 1). For the faunas without numeric dating, we estimate the age of Gulongshan (GLS) fauna roughly equivalent to that of Upper Cave one, as 16-20 ka. We estimated the age of Shanchengzi fauna as 20-30 ka, evidently older than that of Upper Cave fauna, overlaps that of Yanjiagang fauna, partly overlaps that of Yushu fauna and younger than that of Xiaogushan fauna (Table 1), although the extinction rates indicate its age would be between those of Yushu and Xiaogushan.Table 1
Table 1Comparison of the results of the 1st group faunas with different methods
| Cladistics | B.-R.’s rule1) | Antiquity1) | Extinction rate1) | Dated age | Estimated age |
|---|---|---|---|---|---|
| GLS | GLS | SDD (100) | SDD (13.64%) | GLS: 16-20 ka | |
| SDD | SDD | GLS (120) | GLS (17.65%) | SDD: 18 ka (Huang, 1991) | |
| SCZ | SCZ | SCZ (126) | YJG (26.67%) | SCZ: 20-30 ka | |
| XGS | XGS | SLWS (128) | SLWS (27.27%) | YJG: (27.0±0.6) ka (Wei et al., 1986) | |
| SLWS | YS | YJG (132) | DC (30.00%) | YS: 27-40 ka (Sun et al., 1981) | |
| DC | YJG | DC (144) | YS (33.33%) | XGS: 30-80 ka (Zhang et al., 2010) | |
| YJG | SLWS | YS (146) | SCZ (35.9%) | SLWS: 61-68 ka (Yin and Huang, 2004) | |
| YS | DC | XGS (156) | XGS (37.50%) | DC: 100 ka (Huang, 1991) | |
| LJ | LJ | LJ (162) | LJ (52.94%) | LJ: 105-125 ka (Li et al., 2017) | |
| MHS | MHS | MHS (174) | MHS (53.13%) | MHS: <400 ka (NET et al., 1987) |
新窗口打开|下载CSV
4.2 The second group of faunas
The results of the cladistic analyses for the second group (Fig. 2) show that Hexian Man fauna (Ltd) is closest to the presumed modern fauna of the group and they form a clade with Chenjiawo fauna. Three faunas of Peking Man horizons at Zhoukoudian form a reasonable clade, but surprisingly the faunas from upper and lower horizons are more closely related than the middle one, the result is the same as that according to B.-R.’s rule (Dong, 2016). They form a clade successively with the faunas from Yiyuan Man site (Qzas), and then Nanjing Man site (Ts). The faunas from Gongwangling (Gwl) and Yunxian Man site (Qyhk) also form a reasonable clade. But the clade by the faunas from Yuanmou Man site (YmM) and that of Dongzhi Man site (Hld) is unexpected. It results from the numerous undeterminable species in Hld fauna whose genera are the same with those of YmM, and the taxon number of Hld is not sufficient on the other hand. Therefore, it does not indicate their close relationship in biochronology. It is relatively complicated in comparing the cladistic results with those by binary faunal similarity coefficients sequenced according to Brainerd-Robinson’s rule, antiquity coefficients and extinction rates (Table 2). It likely results from the zoogeographic differences of the faunas that make the fauna compositions very varied. We gave our estimated ages of the faunas not yet dated by physical or chemical methods in Table 2 based on the results by different biochronological methods.Table 2
Table 2Comparison of the results of the 2nd group faunas with different methods
| Cladistics | B.-R.’s rule1) | Antiquity1) | Extinction rate1) | Dated age | Estimated age |
|---|---|---|---|---|---|
| Ltd | Gwl | Hld (81) | Mp (33.33%) | Hld: 150-400 ka | |
| Cjw | Ltd | Ts (87) | Hld (36.36%) | Zkd 3-5: 230-310 ka (Zhao et al., 1985);>400 ka (Shen et al., 2001) | |
| Zkd 3-5 | Mh | Xhs (88) | Xhs (40.00%) | Xhs: 150-400 ka | |
| Zkd 8-11 | Lgd | Zkd 3-5 (89) | Ts (43.75%) | Ts: 350 ka (Chen et al., 1996); 500 ka (Wang et al., 2002) | |
| Zkd 6-7 | Bld | Qzas (90) | Bld (46.43%) | Ltd: 150-190 ka (Chen et al., 1987); 300 ka (Huang P H et al., 1995) | |
| Qzas | Qyhk | Cjw (92) | Zkd 3-5 (47.06%) | Qzas: 320-420 ka (Han et al., 2016) | |
| Ts | Mp | Zkd 6-7 (94) | Cjw (50.00%) | Zkd 6-7: 380 ka (Zhao et al., 1985) | |
| Xhs | Dh | Ltd (95) | Qzas (53.85%) | Cjw: 650 ka (An and Ho, 1989) | |
| Dh | Xhs | Bld (97) | Dh (57.89%) | Dh: 500-700 ka | |
| Qyhk | Zkd 8-11 | Dh (98) | Ltd (58.06%) | Zkd 8-11: 462 ka (Zhao et al., 1985); 750 ka (Shen et al., 2009) | |
| Gwl | Zkd 3-5 | Mp (99) | Zkd 6-7 (59.09%) | Bld: 500-850 ka | |
| Mp | Zkd 6-7 | Zkd 8-11 (102) | Zkd 8-11 (55.26%) | Mp: 500-850 ka | |
| Hld | Ts | Qyhk (104) | Qyhk (65.22%) | Qyhk: 936 ka (de Lumley et al., 2008) | |
| YmM | Qzas | Gwl (106) | Lgd (72.50%) | Gwl: 1.15 Ma (An and Ho, 1989); 1.7 Ma (Zhu et al., 2015) | |
| Bld | Cjw | Lgd (114) | Gwl (75.00%) | Lgd: 2.14 Ma (Gao and Cheng, 2004) | |
| Lgd | Hld | Mh (117) | Mh (80.00%) | Mh: 1.2-1.8 Ma | |
| Mh | YmM | YmM (120) | YmM (80.77%) | YmM: 700 ka (Hyodo et al., 2002); 1.7 Ma (Zhu et al., 2008) |
新窗口打开|下载CSV
4.3 The third group of faunas
The strict and semi-strict consensus trees illustrate that Nihewan (NHW) and Chutoulang (CTL) faunas are the closest, Renzidong (RZD) fauna is the basal fauna in the third group, and the other faunas are generally very close to each other (Fig. 3, right). Dadong (DD) is closely related to Gigantopithecus Cave (JYD), Longgupo is closely related to Longgudong (Fig. 3, left; Fig. 4), just as they are closely related in zoogeography. The close relationship between JYD and DD (dated as 1.2 Ma, see Jin et al., 2009) is also supported by antiquity sequence and B.R.’s rule and we estimate the age of JYD as 1.2-1.5 Ma (Table 3). The European Villafranchian fauna from Saint-Vallier (StV) is the closest to that of Longdan (LD). It is also in accordance with the sequences by antiquity coefficients and extinction rates (Table 3). It might indicate that Longdan fauna and Saint-Vallier fauna were in the same zoogeographic region (Palaeoarctic) as well as similar biochronological period. But in terms of sequenced binary similarity coefficients according to B.-R.’s rule (Table 3), Longdan fauna is also very similar to Chutoulang and Juyuandong faunas. The close relationship between Chutoulang and Longdan is also found in binary faunal similarity coefficients and they are both in Palaeoarctic region. But with JYD of Oriental region, it can be interpreted only by their biochronological context (Table 3). Nevertheless, NHW and CTL form a clade with LD first and then StV in the assembled group (Fig. 4). The age of CTL is very likely older than DD, younger than LD and StV, equivalent or slightly younger than NHW. We estimate the age of CTL as 1.6-1.9 Ma. RZD is the closest branch to the presumed archaic one in both the third and assembled groups (Figs. 3-4). Its oldest status in the third group is also supported by B.-R.’s rule and Antiquity sequence but its extinction rate is lower than those of CTL, NHW, StV and LD, we estimate its age as 1.9-2.4 Ma (Table 3).Table 3
Table 3Comparison of the results of the 3rd group faunas with different methods
| Cladistics | B.-R.’s rule1) | Antiquity1) | Extinction rate1) | Dated age | Estimated age |
|---|---|---|---|---|---|
| NHW | Gwl | Gwl (198) | DD (61.1%) | JYD: 1.2-1.5 Ma | |
| CTL | Lgd | DD (202) | Gwl (63.4%) | DD: 1.2 Ma (Jin et al., 2009) | |
| LD | DD | JYD (208) | Lgd (65.9%) | Gwl: 1.15 Ma (An and Ho, 1989); 1.7 Ma (Zhu et al., 2015) | |
| StV | JYD | CTL (209) | LGP (69.6%) | CTL: 1.6-1.9 Ma | |
| Gwl | CTL | Lgd (232) | JYD (71.4%) | NHW: 1.7-2.2 Ma (Liu et al., 2012) | |
| JYD | LD | LD (234) | RZD (72.5%) | LGP: 1.12-1.96 Ma (Huang W B et al., 1995) | |
| DD | NHW | StV (238) | CTL (76.7%) | Lgd: 2.14 Ma (Gao and Cheng, 2004) | |
| Lgd | StV | NHW (242) | NHW (85.4%) | LD: 2.16-2.55 Ma (Qiu et al., 2004) | |
| LGP | LGP | LGP (258) | StV (85.7%) | StV: 1.95-2.58 Ma (Sen, 2004) | |
| RZD | RZD | RZD (266) | LD (96.8%) | RZD: 1.9-2.4 Ma |
新窗口打开|下载CSV
5 Conclusion
Cladistic analyses are feasible methods for searching relationship of faunas. It is effective for comparing the faunas with large number of taxa and within the same zoogeographic province as the first group of the faunas for biochronological sequence. With the faunas having insufficient number of taxa or from different zoogeographic provinces, it is recommended to refer the results by other biochronological methods such as antiquity coefficients and extinction rates for comprehensive biochronological interpretation.We estimate the ages of the faunas not yet dated by physical or chemical methods based on the results of present cladistic analyses and refer with those of sequences according to Brainerd-Robinson’s rule, antiquity coefficients and extinction rates as follow:
Gulongshan in Dalian, Liaoning Province: 16-20 ka
Shanchengzi at Benxi, Liaoning Province: 20-30 ka
Hualongdong at Dongzhi Man site, Anhui Province: 150-400 ka
Xinghuashan at Nanzhao Man site, Henan Province: 150-400 ka
Donghe at Luonan Man site, Shaanxi Province: 500-700 ka
Bailongdong at Yunxi Man site, Hubei Province: 500-850 ka
Meipu at Yunxian, Hubei Province: 500-850 ka
Mohui at Tiandong Man site, Guangxi Autonomous Region: 1.2-1.8 Ma
Juyuandong at Liucheng, Guangxi Autonomous Region: 1.2-1.5 Ma
Chutoulang at Chifeng, Nei Mongol Autonomous Region: 1.6-1.9 Ma
Renzidong at Fanchang, Anhui Province: 1.9-2.4 Ma.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 3]
[本文引用: 2]
[本文引用: 5]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 6]
[本文引用: 3]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 3]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}