 ,1,2, 郑晓廷3,4, 王孝理3,4, 王元青1,2, 毕顺东5, 孟津6,1
,1,2, 郑晓廷3,4, 王孝理3,4, 王元青1,2, 毕顺东5, 孟津6,1Evidence of diphyodonty and heterochrony for dental development in euharamiyidan mammals from Jurassic Yanliao Biota
MAO Fang-Yuan,1,2, ZHENG Xiao-Ting3,4, WANG Xiao-Li3,4, WANG Yuan-Qing1,2, BI Shun-Dong5, MENG Jin6,1收稿日期:2018-03-30网络出版日期:2019-01-20
| 基金资助: |
Corresponding authors: *maofangyuan@ivpp.ac.cn
Received:2018-03-30Online:2019-01-20

摘要
贼兽是一已绝灭的哺乳动物型类,它们与多瘤齿兽和相关类群的系统关系尚存争议,很大的原因在于这类动物的牙齿和头骨形态的特征呈哺乳动物冠群和似哺乳动物之间的镶嵌状态。通过显微CT和平板CL技术,对辽宁省侏罗纪燕辽生物群4种真贼兽的8件标本的牙齿形态、磨损状况和牙齿替换信息进行观察和描述,并与贼兽已发表属种牙齿替换进行对比,对二出齿这一被认为可能与哺乳和亲代养育行为演化有关的重要特征在贼兽中的存在与否进行了讨论。结果显示,所有贼兽的颊齿数恒定,在任何萌出的臼齿下均无齿胚保存,且无替换证据,可以确定为单出齿。前臼齿具一次替换,为典型的哺乳动物二出齿。所有真贼兽只有一对下门齿,一对上门齿,仅玲珑仙兽例外,其在增大的I2内侧,有一残留的I1。神兽和仙兽上下颌骨中,都不具有门齿牙胚,而树贼兽和翔齿兽的上下颌骨中,都具有替换牙胚。其中上门齿齿胚具2-3个齿尖,位于萌出的乳门齿背后方;下门齿齿胚根部始于m2下方,齿尖向前延伸至乳门齿的后背方;牙胚随着不同个体发育阶段向前推进的位置不同,较成熟的个体,齿根有逐渐愈合的现象。门齿的替换应晚于最后臼齿的完全萌出,显示了明显的滞后性,相比其他贼兽的门齿替换应为异时发育。牙胚的形态、相对颊齿大小和后端位置与神兽和仙兽已萌出的门齿更为相近,而较树贼兽和翔齿兽已萌出的门齿显得更大且更复杂,因此神兽和仙兽的门齿更可能为恒齿,贼兽类的门齿则更可能为二出齿。树贼兽和翔齿兽极度膨大的前臼齿很可能加长和推迟门齿的替换过程,甚至影响了替换的发生,同时也在一定程度上抑制了最后臼齿的萌出和功能作用。树贼兽和翔齿兽的这些特征显示它们可能具有某种特殊的食性,更倾向于强调前臼齿对食物的摄取和咀嚼处理。而贼兽具有的典型的哺乳动物二出齿特征、稳定的齿式和高度分化的牙齿等,增加了其为哺乳动物的可能性。
关键词:
Abstract
Evidences for tooth replacement of known euharamiyidans are reported based on eight specimens of four species from the Jurassic Yanliao Biota, Liaoning Province, China. Tooth morphologies, eruptional and wear condition, and tooth germs are directly observed and/or revealed by Micro CT or slab CL scan. The euharamiyidan dentition has definite number of cheek teeth and monophyodont molars that are related to precise occlusion. Incisor germs are found in three specimens of Arboroharamiya but not in Shenshou lui and Xianshou linglong. The incisor germs in the upper jaw, presumably I2, have a large crown with two or three cusps; those in the lower jaw, interpreted as the permanent i2, are positioned dorsal to the root of the erupted incisor, interpreted as di2. Comparing dental development within various ontogenetic stages, the incisor tooth germs in Arboroharamiya and Vilevolodon would replace the deciduous incisors in a much later time than when ultimate molars became fully erupted and functional, if it did happen. The available evidence indicates presence of diphyodonty in the loci of the ultimate lower premolar and incisor, which are common mammalian features potentially related to lactation and parental care. The prolonged or delayed eruptions of incisors and ultimate molars in Arboroharamiya and Vilevolodon are probably associated with the specialization of dentition, with emphasis on the P4/p4 chewing function, which should be an autapomorphy and represent a heterochronic shift of tooth replacement in terms of ontogenetic timing comparing to other “haramiyidians”. The heterochronic incisor replacement is probably owing to developmental suppression related to expansion of the premolars, and may have evolved independently multiple times within mammaliaforms.
Keywords:
PDF (5590KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
毛方园, 郑晓廷, 王孝理, 王元青, 毕顺东, 孟津. 侏罗纪燕辽生物群贼兽类牙齿发育双出齿和异时发育的证据. 古脊椎动物学报[J], 2019, 57(1): 51-76 DOI:10.19615/j.cnki.1000-3118.180803
MAO Fang-Yuan, ZHENG Xiao-Ting, WANG Xiao-Li, WANG Yuan-Qing, BI Shun-Dong, MENG Jin.
1 Introduction
With the discovery of several “haramiyidans” from the Jurassic Yanliao Biota in northeastern China (Zheng et al., 2013; Zhou et al., 2013; Bi et al., 2014; Meng et al., 2017; Luo et al., 2017; Han et al., 2017), our knowledge about this group has been greatly improved, although their taxonomy and phylogenetic position still remain contentious (Meng, 2014; Meng et al., 2014, 2018; Luo et al., 2015a, b, 2017; Han et al., 2017). Various hypotheses have been proposed for allotherians recently; some clustered haramiyidans with multituberculates and placed them within Mammalia (Luo et al., 2002, 2007a, b, 2011; Luo and Wible, 2005; Rowe et al., 2008; Ji et al., 2009; Meng et al., 2011, 2018; Zheng et al., 2013; Bi et al., 2014; Krause et al., 2014), but others considered them as non-mammalian cynodonts (Averianov et al., 2011; Luo et al., 2015a, b, 2017; Huttenlocker et al., 2018). The unstable phylogenetic positions of haramiyidans mainly stemmed from their peculiar and mosaic craniodental and postcranial features, which were often interpreted inconsistently.In dental structures, mammals differ from nonmammalian cynodonts in their greatly reduced number of successional teeth per tooth locus and increased complexity from continuous replacements of dentition to a derived pattern of a limited dentition composed of only two generations of heteromorphous teeth (Luo et al., 2004; Kielan-Jaworowska et al., 2004; Rodrigues et al., 2011). Diphyodonty (two generations in dental replacement) has been regarded as a major apomorphy of mammals and correlated with determinate growth pattern of the skull and is probably associated with lactation and parental care (Pond, 1977; Tyndale-Biscoe and Renfree, 1987; Jenkins, 1990; Zeller, 1999, Luo et al., 2004; Kielan-Jaworowska et al., 2004). Diphyodonty thus has a profound impact on mammalian growth patterns and on development of numerous apomorphies in dentition and skull, such as reduced replacement of the postcanines, development of the precise molar occlusion, and formation of the dentary-squamosal temporomandibular joint (Brink, 1956; Hopson and Crompton, 1969; Ziegler, 1971; Hopson, 1971, 1973; Pond, 1977; Kermack and Kermack, 1984; Gow, 1985; Crompton and Hylander, 1986; Luckett, 1993; Luo, 1994; Crompton, 1995; Zhang et al., 1998), and also allows mammals to diversify their feeding habits (Ross et al., 2007; Luo, 2007). However, it is rare to have both deciduous and permanent teeth preserved in Mesozoic mammals (Clemens, 1966; Lillegraven, 1969; Fox, 1981; Archibald, 1982; Cifelli, 1994; Cifelli and de Muizon, 1998; Han and Meng, 2016), although this has been changed at least for euharamiyidans due to the new discoveries (Luo et al., 2017; Han et al., 2017).
Following previous efforts in describing morphologies of the dentition, mandibles, auditory bones, tooth enamel microstructures, and some postcrania of euharamiyidans (Zheng et al., 2013; Bi et al., 2014; Meng et al., 2014, 2017, 2018; Luo et al., 2017; Han et al., 2017; Mao et al., 2016, 2017), here we present additional evidence of tooth replacement in most euharamiyidans known from the Yanliao Biota. Our work is based on direct observation of specimens with exposed tooth germ and on embedded tooth germs that are revealed by computed laminography (CL) and computed tomography (CT) scanning. The new evidence suggests presence of diphyodonty in euharamiyidans. While it will take time to amass data to resolve the discrepancy between competing phylogenetic hypotheses about “haramiyidans”, multituberculates, and/or allotherians, it is helpful to continue deepening our knowledge about the morphology of euharamiyidans. Our finding of potential diphyodonty in euharamiyidans provides an additional piece of evidence for mammalness of the peculiar group.
2 Materials and methods
Eight specimens of four euharamiyidan species from the Jurassic Yanliao Biota were X-ray computed tomography (CT) and computed laminography (CL) scanned to explore tooth eruption and replacement in the group, including the holotypes of two Arboroharamiya species, an unpublished Arboroharamiya sp., the holotype of Xianshou linglong (IVPP V 16707), and the four specimens of Shenshou lui (Holotype, LDNHMF2001, Lande Museum of Natural History, Tangshan, Hebei Province, China; Paratype 1, WGMV-001, Museum of Wuyishan Mountain, Fujian Province; Paratype 2, 3, JZT-CK005, JZT-D061, Jizantang Paleontological Museum, Chaoyang, Liaoning). Of these specimens the incisor germs in the holotypes of A. jenkinsi (STM 33-9, Tianyu Museum of Nature, Pinyi, Shandong Province, China), A. allinhopsoni (HG-M 017, the Paleontology Center, Bohai University, Jinzhou, Liaoning Province, China), and an unpublished specimen of Arboroharamiya sp. (IVPP V 18954, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing, China) were exposed from the jawbones so that direct observations are also available.Fig. 1
 新窗口打开|下载原图ZIP|生成PPT
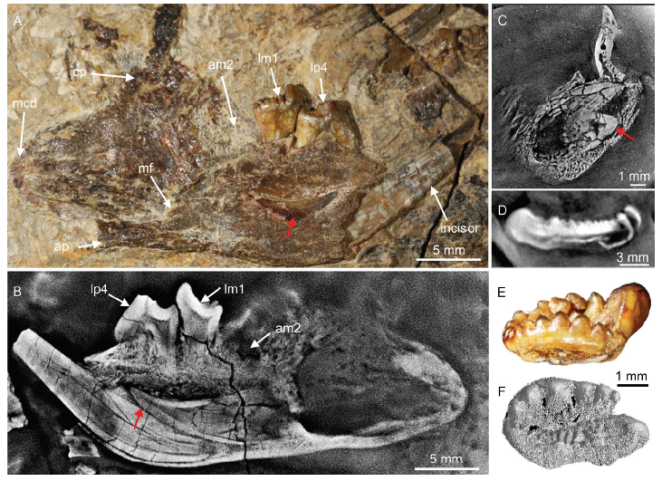
新窗口打开|下载原图ZIP|生成PPTFig. 1Tooth replacement and identification of Arboroharamiya jenkinsi (holotype, STM 33-9)
A, B. the left mandible in medial and optic view (A) and in CL view (B), showing that a tooth germ is dorsal to the root of the enlarged incisor in the dentary bone; C. upper incisor germ within a fragmentary maxilla, revealed by CL scan; D. lm2 in a CL image slice; E. rm2 in labial view; F. reconstruction of the rM2 from CL scan. Red arrows point to tooth germs
The CT scan was carried out using the 225 kV micro-computerized tomography and scanned with beam energy of 120 kV and a flux of 120 μA at a resolution of 9.72 μm per pixel for the upper teeth for paratype 3 of Shenshou lui (JZT-D061), and these were done using a 360° rotation with a step size of 0.5° and an unfiltered alininium reflection target. A total of 720 transmission images were reconstructed in a 2048 by 2048 matrix of 1536 slices using a two-dimensional reconstruction software developed by the Institute of High Energy Physics, CAS (IHEP). The computed laminography scanner was developed by IHEP and has been installed at the Key Laboratory of Vertebrate Evolution and Human Origins, Chinese Academy of Sciences, where the scanning was carried out. Specimens were scanned by CL scanner with a beam energy of 70 kV and a flux of 70 μA at a resolution of 10.89 μm per pixel for the upper incisor germ of STM 33-9, 28.46 μm per pixel for the lower jaw of STM 33-9, 10.9 μm per pixel for the teeth of HG-M 017, 17.70 μm per pixel for the skull of V 18954, using a 720° rotation with a step size of 0.5° and getting a total of 720 image slices; 100 kV and a flux of 50 μA at a resolution of 28.1 μm per pixel for the skull of HG-M 017, 24.91 μm per pixel for the skull of LDNHMF2001, 29.8 μm per pixel for the skull of WGMV-001, 28.59 μm per pixel for the skull of JZT-D061; 90 kV and a flux of 50 μA at a resolution of 19.4 μm per pixel for the lower jaw of JZT-D061, 21.97 μm per pixel for the skull of JZT- CK005 using a 360° rotation with a step size of 1° and getting a total of 360 image slices. These image slices, each with a size of 2048 by 2048 pixels, were reconstructed using a modified Feldkamp algorithm developed by the IHEP. The CT/CL data were rendered to produce three-dimensional reconstructions of specimens using the VGstudio Max 3.02 (Volume Graphics).
Fig. 2
 新窗口打开|下载原图ZIP|生成PPT
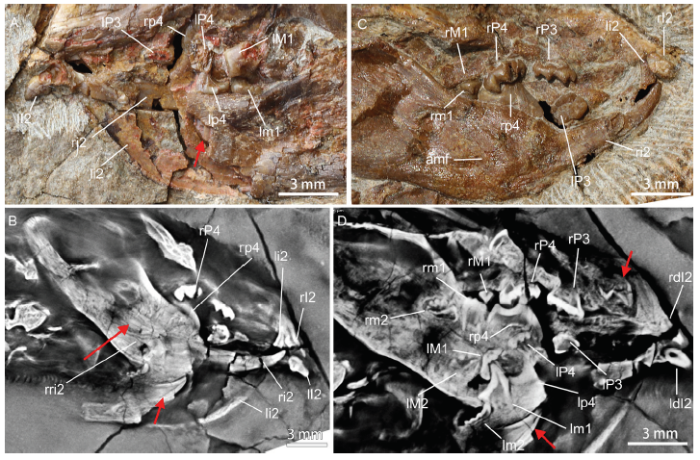
新窗口打开|下载原图ZIP|生成PPTFig. 2Tooth condition of Arboroharamiya allinhopsoni (HG-M 017, holotype)
A. lateral view of the left partial skull, where the lower incisor germ is exposed at the breakage, posterodorsal to the root of the erupted incisor; B. CL image showing the left and right lower incisor germs in the jaws; C. optic image of the lateral view of partial right skull, showing the barely worn cheek teeth; D. CL image showing the two-cusped upper incisor germ in the maxilla, the unerupted two ultimate lower molars (m2), and the unerupted left ultimate upper molar (M2). Red arrows point to tooth germs
Optical images are taken using a Canon digital camera with a macro lens and a Zeiss microscope (SteREO Discovery V. 20) with a digital imaging system (AxioVision SE64 Rel. 4.9). For SEM imaging, the specimens were un-coated and imaged using a Hitachi S4700 scanning electron microscope. Most SEM photographs were taken at 3 to 5 kV voltage and magnifications are commonly between 10-30 times for the general shape of the tooth.
Abbreviations am2, alveolus for m2; amf, anterior extremity of the masseteric fossa; ap, angular process; cp, coronoid process; l, left; mcd, mandibular condyle; mf, mandibular foramen; r, right; rr, right root.
3 Descriptions
Arboroharamiya jenkinsi By removing matrix and a small piece of bone of lower jaw from the holotype (STM 33-9), a tooth germ was exposed posterodorsal to the root of the erupted lower incisor and ventral to p4 and m1 (Fig. 1A). The tooth germ is completely in the dentary bone and has only the crown formed. The germ crown measures about 6 mm long, with its tip beneath the anterior root of the p3; its distal end is widely open and extends to the level below m2, anterior to the edge of masseteric fossa. We interpret the tooth germ as the permanent (successive) incisor to the enlarged one that is in function. Although the incisor germ coexists with the erupted one (presumably the deciduous), there seems no sign of replacement: the root of the deciduous one is nearly closed, and the germ crown is deeply in the dentary (Fig. 1B). The germ crown bears enamel with pigmentation being darker than the erupted incisor. The germ crown has a sharp dorsal and ventral edge, respectively; its medial surface is nearly flat, whereas its lateral surface is convex. The erupted left incisor has a large part of the crown broken and the rest of the tooth does not seem to have much enamel coverage. The tooth thickens distally, and the thickest region is ventral to p4 within the dentary; then it gradually thins distally and ends at the level of m2. A maxillary fragment is between the two lower jaws, from which CL scan reveals a sizable upper tooth germ. The upper tooth germ has two distinct cusps, with the mesial one being higher but narrower than the distal one (Fig. 1C). The orientation of the germ is tentative because the isolated preservation of the maxillary fragment.Fig. 3
 新窗口打开|下载原图ZIP|生成PPT
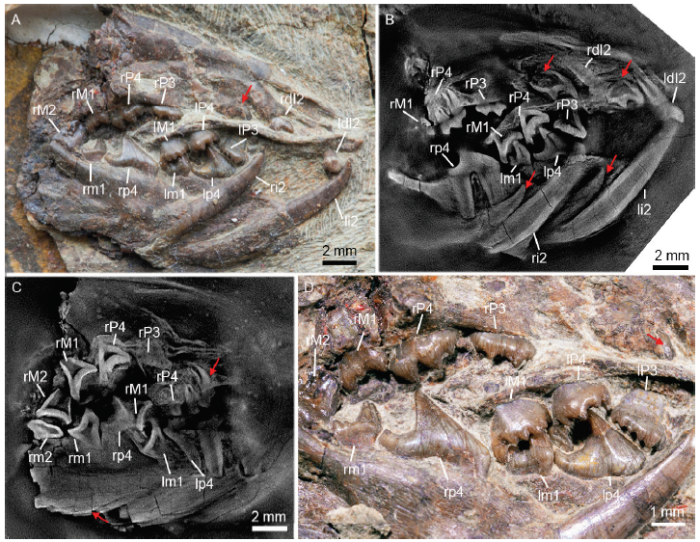
新窗口打开|下载原图ZIP|生成PPTFig. 3Tooth condition of Arboroharamiya sp. (IVPP V 18954, new specimen)
A, D. optic images of the upper incisor germ (A) with the closer view (D) showing the dentition and the exposed mesial cusp; B, C. CL images showing the three cusped upper incisor germ (I2), two cusped upper incisor (dI2), and the fully formed ultimate molars. Red arrows point to tooth germs
P3, P4/p4, and M1/m1 are fully erupted, and have root tips closed, or nearly so. The sole lower premolar (p4) is hypertrophic and its roots extend ventrally to the lateral side of the tooth germ and the root of the erupted incisor. M1/m1 and P4/p4 show wear facets on the crowns. P3 bears little or no wear (Meng et al., 2014), whereas wear facets on P4/p4 are clear; this indicates that either P4 erupted earlier than P3 or functioned as the main tooth again p4 in chewing, or both. CL images reveal two unexposed teeth, of which one is identified as the right P3 (not illustrated) and the other as the left m2 (Fig. 1D). The CL images also show that the m2 has only the crown formed but not the root, similar to the condition of the exposed right m2 (Fig. 1E). CL scan reveals no tooth germ under the premolar or molar loci in the mandible (Fig. 1B).
Two split molars are embedded in the split slabs; both were identified as M2 by Meng et al. (2014). However, the lengths of these two teeth are different; the two corresponding cusp rows measure (L/W) 4.74/4.26 mm (with width estimated) and 3.47/3.9 mm long, respectively. The number of the cusps is also different between these two teeth, as revealed by CL scan. The cusps of the longer one are notably more than the short one. The longer tooth has been partially reconstructed, but because the CL scan data do not have the necessary resolution, the rendered tooth morphology is not fully resolved (Fig. 1F). However, the outline of the tooth is recognizable, which we identify as the right M2. Similar to m2, M2 does not have root preserved. In contrast, the short tooth has clear impression of a long root with part of the crown missing. Thus, the shorter tooth is more likely the left M1, which is similar to the isolated M1 in size (L/W: 3.67/3.2, Meng et al., 2014). All the data show that M2/m2 of A. jenkinsi do not have the root formed and probably have not fully erupted, although m2 does bear wear (Meng et al., 2014).
Arboroharamiya allinhopsoni Similar tooth germ has been found in each mandible of the holotype specimen of Arboroharamiya allinhopsoni (HG-M 017, Fig. 2A, B), as revealed by CL images (Fig. 2C, D). The tooth germ is posterodorsal to the root of the deciduous incisor and ventral to p4 and m2. The root of the deciduous one is closed. The tooth germ in deeply embedded in the dentary, showing no sign of replacement, and is morphologically similar to that of A. jenkinsi (Fig. 2C). The roots of the enlarged p4 extend ventrally and end laterally to the root of the erupted incisor. CL scan also shows that there is also a sizable tooth germ within the maxilla, posterodorsal to the erupted upper incisor. The tooth germ has at least two distinct cusps, a high mesial one and a small distal one (or more), differing from the deciduous incisor that has only one major mesial cusp and several miniscule distal cuspules. Again, there is no tooth germ in any cheek tooth locus.
P3, P4/p4, and M1/m1 are fully erupted, and have root end closed, or nearly closed. The crown view of most cheek teeth is invisible because the upper and lower teeth are preserved in occlusal position, except for the left P3 and P4 (Han et al., 2017, extended data fig. 4; Fig. 2C). Both P3 and P4 show little wear facet. The crowns of the ultimate upper molars are formed but not fully erupted. The roots of these molars are absent and most part of the crown appears still in the jaw bone. These suggest that the holotype of A. allinhopsoni represents an individual animal that is slightly younger than the one represented by the holotype of A. jenkinsi. However, the holotype of A. allinhopsoni is significantly smaller than that of A. jenkinsi.
Arboroharamiya sp. An unpublished specimen (IVPP V 18954) has been tentatively assigned to Arboroharamiya sp. because of its similarity to A. allinhopsoni (Fig. 3). This specimen has the ultimate molars fully erupted and their roots formed, which indicate that the specimen represents an adult individual. Still, the specimen contains incisor germs, and the large mesial cusp of the right upper incisor germ was actually exposed posterodorsal to the root of the erupted upper incisor (Fig. 3A). CL scan revealed that the upper incisor germs are tricuspid with almost closed root (Fig. 3B). The posterior ends of the lower incisor germs are broken; thus where the germ root ends and whether or not the root is closed remain unknown (Fig. 3C). The tips of the lower germs extend anterior to the anterior root of p4. We consider that both the upper and lower incisor germs are at the ontogenetic stage about to erupt. The root of the functioning lower incisor is closed but shrunk; it extends to the level of m1 and shows the trend of shedding (Fig. 3B). The m2s are blocked by jawbone, but CL scan shows that the left m2 is occluded with the M2, while the right m2 is popped out and the left m2 is damaged. Because the crown views of cheek teeth are invisible, the wear facets of them are unknown (Fig. 3D). All evidence suggests that V 18954 represents an individual animal that is older than the two holotypes of Arboroharamiya; thus, it forms a convincing evidence that the incisor germ of Arboroharamiya would erupt at a time much later than the eruption of the ultimate molars, or the replacement of the incisors is considerably prolonged or delayed.
Xianshou linglong Xianshou linglong (IVPP V 16707) is quite different from other euharamiyidans in that it has two pairs of upper incisors. The lateral one, identified as I2 (Bi et al., 2014; Han et al., 2017), is enlarged and possesses three cusps, with the size of cusps decreasing distally (Fig. 4A). A very small incisor, identified as the left I1, is medial to I2 and comparable in size to the smallest distal cusp of I2 (Fig. 4A, B). CL scan reveals a short and curved root of I1, which was later exposed by unintended preparation. No incisor germ is in the maxilla (Fig. 4B). The right I1 was not preserved. The root of the lower incisor is not completely closed and proportionally longer and more robust than that of Arboroharamiya; it extends distally to the level below m2 with a prominent expansion under the position of p4 and m1 (Fig. 4D). The roots of p4 extend ventrally to the lateral side of the root of the incisor, similar to that of Arboroharamiya.
Fig. 4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Tooth condition of Xianshou linglong (IVPP V 16707, holotype)
A, B. optic image (A) and CL image (B) showing the three cusped second upper incisor (I2) and the single cusped first upper incisor (I1); C. CL image showing the small single cusped I1; D. CL image showing the wear condition of the cheek teeth
All cheek teeth of the holotype of X. linglong are fully formed and erupted (Fig. 4D). M1/m1, P4/p4 and P3 all show clearly occlusal wear on the crowns. The wear condition of M2/m2 is not clear because they are in occlusal position. The development stage of the last molars shows that the holotype of X. linglong represents an individual that is older than those represented by the holotype specimens of the two species of Arboroharamiya.
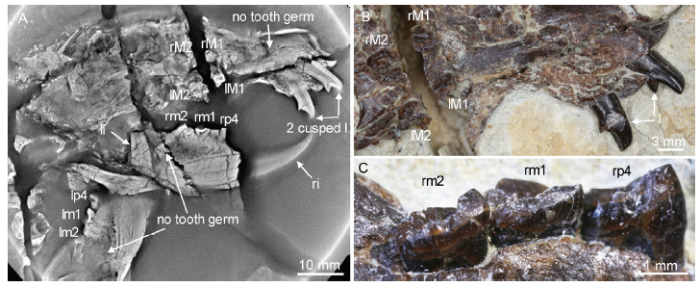
Shenshou lui Four specimens have been assigned to Shenshou lui (Bi et al., 2014) of which none has the incisor germ, as revealed by CL scan (Figs. 5-8). All specimens are from adult individual animals, judging from their fully erupted ultimate molars with closed roots. However, these specimens show different degree of tooth wear, indicating slightly different ontogenetic stages. Ranging from the youngest to the oldest, these specimens are WGMV-001, JZT-D 061, LDNHMF 2001, and JZT-CK 005 (Figs. 5-8). Because their ultimate molars show little or no wear, we consider that JZT-CK 005 and WGMV-001 represent animals with a similar age. Of the four specimens, the root of the enlarged lower incisor has a closed end and extends distal to the ultimate molar (m2), except for JZT-D061 in which the root appears ended under the distal root of m2. S. lui differs from other euharamiyidans from the Yanliao Biota in having a relatively small and more molariform p4; its roots end dorsal to the root of the lower incisor. CL images also show that the upper incisors of paratype 2 (JZT-CK 005) and paratype 3 of S. lui (JZT-D 061) possesses three cusps, although they are not so well developed compared to those of the tricuspid I2 of Xianshou linglong (IVPP V 16707) and Arboroharamiya sp. (V 18954).
Fig. 5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Tooth condition of Shenshou lui (LDNHMF 2001, holotype)
A. CL image showing the two cusped upper incisors, absence of tooth germ, and wear condition of cheek teeth; B, C. optic images showing wear facets of the upper incisors (B) and the left lower cheek teeth (C)
Fig. 6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Tooth condition of Shenshou lui (WGMV-001, paratype 1)
A, B. CL images showing the two cusped upper incisors, closed root of the lower incisor, absence of tooth germ, and wear condition of the cheek teeth; C, D. optic images showing the wear condition of the right lower cheek teeth (C) and the left lower cheek teeth (D)
4 Discussion
Tooth replacement in mammaliaforms is phylogenetically significant and in many cases related to feeding behavior (Winge, 1941; Ziegler, 1971; Luckett, 1985, 1993) and allows mammals to diversify their feeding habits (Ross et al., 2007; Luo, 2007). Although the definition and phylogenetic interest of tooth replacement vary through time (Rowe, 1993), mammals were commonly considered to be distinguishable from other vertebrates in having a diphyodont antemolar dentition that has been associated with lactation and prolonged parental care (Hopson, 1973; Pond, 1977; Tyndale-Biscoe and Renfree, 1987; Jenkins, 1990; Zeller, 1999, Luo et al., 2004; Kielan-Jaworowska et al., 2004). The diphyodont dentition has a profound impact on mammalian growth patterns and on development of numerous mammalian apomorphies in dentition and skull, such as reduced replacement of the postcanines, development of the precise molar occlusion, and formation of the dentary-squamosal temporomandibular joint (Brink, 1956; Hopson and Crompton, 1969; Ziegler, 1971; Hopson, 1971, 1973; Pond, 1977; Kermack and Kermack, 1984; Gow, 1985; Crompton and Hylander, 1986; Luckett, 1993; Luo, 1994; Crompton, 1995; Zhang et al., 1998).In nonmammalian cynodonts, three tooth replacement patterns have been recognized (Abdala et al., 2013): 1) an alternate postcanine replacement, as in Thrinaxodon and several cynodonts (Parrington, 1936; Crompton, 1963; Osborn and Crompton, 1973; Gow, 1980; Crompton and Luo, 1993); 2) a sequential postcanine replacement with variable generations in a mesiodistal direction, as in some diademodontids and traversodontids (Fourie, 1963; Crompton, 1963, 1972; Hopson, 1971; Osborn, 1974a; Liu and Sues, 2010); 3) a sequential addition at the distal end of tooth row without any replacement, as in some tritylodontids and gomphodonts (Ku?hne, 1956; Hopson, 1965, 1971; Clark and Hopson, 1985; Cui and Sun, 1987). Compared to the continuous replacements of dentition in diapsids, these cynodont patterns reflect reduced generations of tooth replacement per tooth locus, but are still plesiomorphic to mammals (Kielan-Jaworowska et al., 2004).
In studying tooth replacement of fossil species, Abdala et al. (2013) suggested that tooth replacement could be achieved by scanning an ontogenetic series. However, fossil species of early mammals are often represented by limited number of specimens. Conventionally, several features have been used as evidences for tooth replacement in fossil species. These include the groove in the jaw that communicate with pits housing permanent teeth in which replacement teeth developed and assumed dental lamina housed (Crompton and Luo, 1993; Parrington, 1971), the residual space or sign such as replacement pits or “alveolar gap” that was left by the predecessor tooth, and traces for loss of functioning tooth, such as shedding, reabsorbing, and erosion of roots that can be observed in jaws (Greenwald, 1988; Abdala et al., 2013). Tooth wear gradience in dentition and presence of the tooth germ within jawbone as revealed by CT scan are informative to interpret tooth eruption sequence and patterns, such as alternate or sequential replacement (Abdala et al., 2013; Lopatin and Averianov, 2015). Continuous growth of the jaw and skull in ontogeny and “indeterminate” growth pattern (or “extended” adult growth) were often considered in association with sequential tooth replacement or related to replacement of larger succeeding teeth or additional teeth to the tooth row (Edmund, 1960; Osborn, 1974b; Crompton and Luo, 1993; Zhang et al., 1998; O'Meara and Asher, 2016).
For many decades, it is difficult to discuss tooth replacement among “haramiyidans”, because all members of the group were represented by isolated teeth (Owen, 1871; Hennig, 1922; Simpson, 1928; Parrington, 1947; Peyer, 1956; Hahn, 1973; Clemens, 1980; Jenkins et al., 1983; Sigogneau-Russell et al., 1986; Hahn et al., 1989; Sigogneau-Russell, 1989; Butler and MacIntyre, 1994; Heinrich, 1999, 2001; Butler, 2000; Butler and Hooker, 2005; Maisch et al., 2005; Hahn and Hahn, 2006; Clemens, 2007; Martin et al., 2010a; Averianov et al., 2011; Clemens and Martin, 2014). The Late Triassic Haramiyavia clemmenseni has the lower jaw preserved (Jenkins et al., 1997), and CT scan in a recent study revealed that the holotype of the species has completed tooth replacement and the ultimate molars are fully erupted (Luo et al., 2015b). The condition in Haramiyavia can be used as a primitive state in understanding tooth replacement in “haramiyidans”, particularly those represented by skeletal specimens from the Jurassic Yanliao Biota.
All known “haramiyidans” with dentition preserved (except for Haramiyavia and Cifelliodon) lack the canine and have a reduced number of incisors, premolars and molars; thus, our discussion will focus on two regions of the dentition, the cheek teeth (premolar and molars) and the incisors. The tooth replacement is related to ontogenetic stages in mammals so that we will first consider the ontogenetic variations represented by the specimens we studied and those reported in various publications.
Ontogenetic variation The CL scan data and optic images are used to display the condition of incisor germ in preservation. The abrasive wear level of tooth, the tooth formation and root closure of teeth, and the eruption degree of molars are used as age marks to show ontogenetic differences among specimens examined. Among reported specimens and some unpublished ones available to us, the youngest known individual of euharamiyidans is represented by the specimen of an unnamed eleutherodont (BMNH 3258). As described by Meng et al. (2017: extended data fig. 4), the specimen “has the lower permanent premolar half-erupted to replace the deciduous premolar that is only represented by root alveoli at the ultimate premolar locus; M1 crown present but the roots not yet formed.” In contrast, the oldest known individual of euharamiyidans is represented by paratype 2 of Shenshou lui, in which M2 and m2 were fully erupted and deeply worn (Fig. 7).
Fig. 7
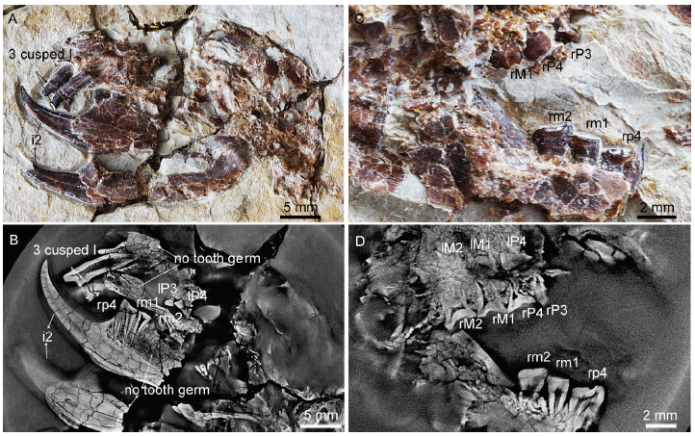 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 7Tooth condition of Shenshou lui (JZT-CK005, paratype 2)
A, B. optic (A) and CL (B) images showing the three cusped upper incisor and no teeth germ under any tooth locus; C, D. optic (C) and CL (D) images showing the deep wear facet of the cheek teeth
Of the three specimens of Arboroharamiya, the holotype of A. allinhopsoni and the specimen assigned to Arboroharamiya sp. are much smaller than A. jenkinsi, as shown by their sizes of teeth, mandibles, and postcrania. For instance, the tibial length of A. allinhopsoni is 21.5 mm (Han et al., 2017) and that of Arboroharamiya sp. (IVPP V 18954) is 21.7 mm (personal observation), whereas it is 49.6 mm in A. jenkinsi (STM 33-9, Zheng et al., 2013). However, both HG-M 018 and STM 33-9 are considered younger than V 18954 because the roots of their ultimate molars have not been formed. The crown of m2 in the holotype of A. jenkinsi may not be fully erupted, but it bears wears (Meng et al., 2014), indicating that m2 was partly in function. In the holotype of A. allinhopsoni the crown of m2 is not fully formed (Han et al., 2017), and it is difficult to see its cusp because of the preservation condition. From the general shape of the crown, we think that the holotype of A. allinhopsoni represents an individual slightly younger than that of A. jenkinsi. In contrast, the specimen of Arboroharamiya sp. (V 18954, Fig. 3) represents an adult; its ultimate molars already erupted and have closed roots. The holotype of V. diplomylos (BMNH 2942) was also considered as from a relatively young adult animal, probably older than those represented by the holotypes of A. allinhopsoni and A. jenkinsi but younger than V 18954, because the ultimate molars of A. allinhopsoni and A. jenkinsi are not fully formed and erupted. However, the femoral length of BMNH 2942 is 21.5 mm (Luo et al., 2017), comparable with that of A. allinhopsoni but notably smaller than STM 33-9. In BMNH 2942 and V 18954, the roots of the ultimate molars are formed, with the ends being closed. In addition, M2 and m2 already show occlusal wears in BMNH 2942 (Luo et al., 2017:fig. 1c, d), indicating that these teeth were in full function (Luo et al., 2017: supplementary information:15). The wear facet for the cheek teeth of V 18954 is not visible because of the preservation condition, but the root of the upper incisor germ is almost closed, suggesting that it may represent the oldest individual among the specimens that possess incisor germs.
In contrast, there is no incisor tooth germ in Shenshou lui and Xianshou linglong, as we reported above and compared in Fig. 9. All specimens assigned to S. lui represent adult animals, judging from their deeply worn teeth and fully erupted ultimate molars. The holotype of X. linglong also represents an adult individual in which the ultimate molars have roots and fully erupted, although it appears younger than the specimens of S. lui because it is lightly worn. The holotype of X. songae was considered as “a juvenile specimen”, using the individual dental age stages (IDAS; Anders et al., 2011) (Luo et al., 2017). The IDAS is established on eutherians with three molars, whereas euharamiyidans have only two molars, as in multituberculates. Moreover, if tooth eruption is prolonged or delayed in some euharamiyidans, then whether the ontogenetic stages of these species can be judged using conventional criterion or the IDAS becomes questionable. M2 of the holotype of X. songae specimen is nearly fully erupted (Bi et al., 2014: extended data fig. 6) so that the individual could be considered as a sub-adult individual.
The holotype of Maiopatagium furculiferum (BMNH 2940) represents an adult individual, similar in ontogenetic stage to the paratype 2 of S. lui (JZT-CK 005), but the lower jaw and teeth are not preserved. This species was thought to be most similar to S. lui (Meng et al., 2017), although their occlusal patterns are significantly different (Meng et al., 2017; Han et al., 2017).
In general, however, the known specimens of Shenshou, Xianshou, Vilevolodon, and Arboroharamiya represent individual animals of similar age or with slightly different ages, except for BMNH 3258; thus, presence or absence of the tooth germ in these specimens is probably not age-related but may well be associated with the specialization of the dentition, as discussed below.
Cheek Teeth It is clear that cheek tooth eruption and replacement of “haramiyidans” do not belong to any of the three tooth replacement patterns in non-mammalian cynodonts, as outlined by Abdala et al. (2013). Instead, euharamiyidans show a typical mammalian pattern of cheek tooth dentition that is differentiated into incisors, premolars, and molars; the canine was presumably lost compared to the condition of Haramiyavia and Cifelliodon (Huttenlocker et al., 2018); the latter was interpreted to be similar to the holotype of Hahnodon (Sigogneau-Russell, 1991). All specimens of euharamiyidans (not including Megaconus) clearly show a common cheek tooth formula: P2-M2/p1-m2. The number of cheek teeth is definite regardless of various ontogenetic stages. Definitive number of cheek teeth in euharamiyidans premises for precise tooth occlusion.
As evidenced by BMNH 3258 that represents by far the youngest individual, at least the lower premolar is diphyodont and that replacement of dp4 by p4 took place before or about the same time along with the eruption of m1. All evidence also convincingly shows that all molars are monophyodont in euharamiyidans. Presence of two pairs of upper and lower monophyodont molars that erupt in a mesiodistal sequence is shared by euharamiyidans and multituberculates among mammaliaforms; it must be a derived condition in relation to the dentition with three or more molars (or molariforms) in either non-mammaliaform cynodonts, Haramiyavia, or any mammaliaforms with a triconodont tooth pattern or its derivatives, such as symmetrodontans and therians. Whether the two-pair molar condition evolved independently in euharamiyidans and multituberculates or is a synapomorphy for the two groups depends on their phylogenetic placements, an issue that remains open.
The cheek teeth of euharamiyidans display some significant differences in premolar and molar morphologies, cusp shape, and size. These differences are potentially related to tooth replacement. In Shenshou lui, the cheek teeth are relatively simple and proportionally small with their roots ending dorsal to the root of the lower incisor (Fig. 9A). In contrast, the cheek teeth of arboroharamiyidans (including Vilevolodon) are proportionally much large and have a hypertrophic A1/a1 on p4 and broadly basined P4. The roots, particularly those of p4, are long and extend to the lateral side of the root of the erupted incisor and the incisor germ. The condition in Xianshou appears to be intermediate between S. lui and arboroharamiyidans (Fig. 9B). Enlarged cheek teeth are stronger and have larger chewing area of the crown; thus, tooth enlargement potentially enhances efficiency of food processing. In particular, arboroharamiyidans show a trend of emphasizing P4/p4 function by increasing their size and complexity; this is probably correlative to the delayed eruption of the ultimate molars, as seen in several specimens (Fig. 9C, D); the function of the ultimate molars perhaps became less important in advanced arboroharamiyidans that have enlarged and complex ultimate premolars as the main functional unit for food processing.
Incisors The incisor replacement of euharamiyidans appears more complicated than the cheek teeth. Among the reported Yanliao euharamiyidans, all species have one pair of functional lower incisors. Among them Shenshou lui and Xianshou linglong do not have incisor germ in both lower and upper jawbones (Fig. 9A, B). However, S. lui has only one pair of upper incisors but X. linglong has an extra miniscule upper incisor, identified as I1, in addition to the enlarged tricuspid I2. In contrast, Arboroharamiya jenkinsi, A. allinhopsoni, Arboroharamiya sp., and Vilevolodon diplomylos have one pair of upper incisors as well as upper and lower incisor germs in jawbones (Fig. 9C, D). In S. lui and X. linglong, the root of the lower incisor extends more posteriorly than the root of the erupted incisor of arboroharamiyidans (including Vilevolodon); in the latter taxa, the root of the erupted incisor and the tooth germs are on the medial side of the roots of m1 and p4 and the distal end of the germ roughly levels with the distal end of the incisor in S. lui and X. linglong. The upper incisors are not preserved in the holotype of X. songae, whereas the lower incisor germ was not observed through the breakage on the medial surface of the left lower jaw (Bi et al., 2014), but this needs to be confirmed with CT scan. The upper incisor of Maiopatagium furculiferum is single cusped, differing from the double-cusped upper incisor of S. lui and tricuspid one in Xianshou. We would predict that the holotype of M. furculiferum does not contain the incisor tooth germ.
The upper incisor germ in arboroharamiyidans possesses two or three sizable cusps and the crown appears larger than that of the erupted one but is similar to the upper incisors of Shenshou and Xianshou. The lower incisor germs in arboroharamiyidans are similar in shape, relative size, and position in relation to the ultimate molars, although some differences exist. The size differences of the lower tooth germs in arboroharamiyidans are apparently related to the body size of these individuals, with the largest being that of A. jenkinsi. The relative position of the germ is probably related to different ontogenetic stages. Using the relative position of the germ tip as a reference point, the ontogenetic stages of may range from the youngest to oldest, respectively, in the following sequence: A. allinhopsoni, A. jenkinsi, Vilevolodon, and Arboroharamiya sp. (V 18954). In V 18954, the ultimate molars are completely formed and erupted, while its lower incisor germ extends anterior to p4; both the upper and lower germs are almost erupted, with the upper ones partly visible (Fig. 3).
Several possibilities have been proposed in interpreting the peculiar incisor germs in Vilevolodon (Luo et al., 2017). In particular, the authors thought that “the prolonged and ongoing incisor replacement in Vilevolodon is plesiomorphic” and that “the prolonged replacement is plesiomorphic, but a slower growth of permanent incisor would be unique to Vilevolodon, thus an autapomorphy of Vilevolodon”. We endorse the interpretation for prolonged incisor replacement in Vilevolodon and add that this feature is shared with Arboroharamiya. Whether this represents a derived condition depends on how one interprets the incisor replacement in Shenshou lui and Xianshou linglong. Luo et al. (2017), however, identified the erupted lower incisor and the incisor germ as the first and second generations deciduous incisor 1, denoted as (di-1a) and (i-1b), respectively. For replacement pattern of incisors and canines, Luo et al. (2017: supplementary information:78, ch. 198) coded Vilevolodon as having more than one replacement. With our observation of the specimens, we think that there is no evidence for on-going or multiple generations of incisor replacement in Vilevolodon and Arboroharamiya. The only evidence perhaps came from Luo et al.'s (2017) interpretation of the upper incisors in X. linglong, where the miniscule tooth identified as I1 (Bi et al., 2014; this study) was considered to be typical of the topographic relationship of a replacing successor to a deciduous predecessor already erupted and that “these 'I2' and 'I1' may be two successive replacing incisors of the same incisor locus: the 'I2' is a predecessor, and 'I1' is the emerging apex of a successor in the process of eruption.” These authors then coded X. linglong as having one pair of upper incisors, instead of two. As we show above, the miniscule I1 is a complete tooth with its root in the premaxilla; it is absolutely not the emerging apex of a successor in the process of eruption. It is best interpreted as a vestigial incisor.
In discussing tooth replacement, the polyphyodont dentition (more than two generations of teeth) could be demonstrated if the residual sign of the predecessor and eruptional evidence of succeeding tooth coexisting with the functional one at the same tooth locus. For instance, the coexistence of replacement teeth germ, old remnant roots for the predecessor, together with the functioning tooth at the same locus of Thrinaxodon liorhinus constitute the evidence for polyphyodonty of the taxon (Abdala et al., 2013). By far, there is no evidence supporting the polyphyodonty in “haramiyidans”.
Moreover, the interpretation for on-going incisor replacement in Vilevolodon is difficult to interpret in terms of evolutionary transformation within “haramiyidans”. Based on CT scan data, Luo et al. (2017) found that in the holotype of the Late Triassic Haramiyavia clemmenseni, the incisors and canines are permanent teeth and the ultimate molars are fully erupted. Thus, basal haramiyidans do not have the sustained incisor replacement as seen in Vilevolodon, and now in Arboroharamiya, among the haramiyidan clade. Luo et al. (2017: supplementary information) interpreted that “the ongoing incisor replacement may be a derived feature of eleutherodontids that evolved from within the haramiyidan clade…Placed in the phylogenetic context, late replacement of incisors in the adult stage (IDAS) is both a convergent trait with Sinoconodon, and also unique to eleutherodontids among haramiyidans.” Luo et al. (2017: supplementary information) further concluded that “By the presence of the prolonged (or delayed) incisor replacement in young or full adult stages of the skull, Vilevolodon is different from (and more plesiomorphic than) Morganucodon and docodontans.” These interpretations are contradictory to each other. On the one hand, it was considered as a derived feature within “haramiyidans”, perhaps as a reversal from the condition of Haramiyavia and evolved convergently with that of Sinoconodon; on the other hand, it was considered as more plesiomorphic than the diphyodont dentition in Morganucodon and docodontans.
We would argue for an alternative interpretation for such a prolonged replacement. The primitive condition of haramiyidan clade should be that of Haramiyavia, which probably had a diphyodont dentition, as in Morganucodon (Luo et al., 2017) and docodonts (Luo et al., 2015a, 2017; Meng et al., 2015; Panciroli et al., 2017), although there may be more than one replacement for the upper canine of Haldanodon (Martin et al., 2010b) and resorption of anterior postcanines in mature individuals of Morganucodon, Eozostrodon, and Dinnetherium (Mills, 1971; Parrington, 1971; Crompton and Luo, 1993). We also recognize that the condition in Vilevolodon and Arboroharamiya differs from that of multituberculates in which the incisors completed replacement before or around the same time at eruption of m2 (Miao, 1986; Greenwald, 1988; Luo et al., 2017). In all known specimens of Vilevolodon and Arboroharamiya in which m2 is partly or fully erupted, the incisor germ is still positioned in the jaw bone. We agree with Luo et al. (2017) that the residual germs with combination of fully erupted premolars and molars in Arboroharamiya and Vilevolodon would represent a heterochronic shift of tooth replacement for ontogenetic timing pattern, relative to that of Shenshou and Xianshou and typical mammaliaform pattern, which would be unique.
A key issue, however, is whether the erupted incisors in Vilevolodon and Arboroharamiya on the one hand, and those in Shenshou lui and Xianshou linglong on the other, belong to the same generation. For S. lui and X. linglong, the incisors can be interpreted either as permanent (successive) teeth, homologous to the tooth germs in the jaw bones of Vilevolodon and Arboroharamiya, or as deciduous teeth, homologous to the erupted incisors in the latter taxa. If interpreted as permanent incisors, then one may assume that the dentitions of S. lui and X. linglong, favorably diphyodont, have a tooth replacement pattern primitively similar to that of Haramiyavia, in which the incisor replacement had completed when the ultimate molars erupted, which is the typical mammalian diphyodont replacement (Luo et al., 2002). If the erupted incisors in all known euharamiyidans belong to the same deciduous generation, the absence of the incisor germs in S. lui and X. linglong may be resulted from suppression of the permanent tooth germs at an ontogenetic stage earlier than that in Vilevolodon and Arboroharamiya.
Fig. 8
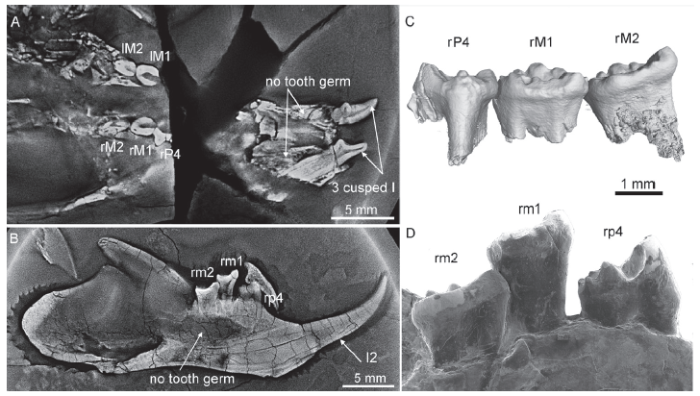 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 8Tooth condition of Shenshou lui (JZT-D061, paratype 3)
A, B. CL images showing three cusped upper incisors and no tooth germ under any tooth locus; C, D. CT reconstruction (C) and SEM (D) images showing wear facets of the cheek teeth
Of the two potential interpretations, we prefer the first one that the incisors in S. lui and X. linglong are the successive teeth, a primitive condition similar to that of Haramiyavia. This interpretation is supported by the fact that the upper incisors in S. lui and X. linglong are larger and more complex than the erupted one of Arboroharamiya and Vilevolodon, and the lower incisors are robust with root extending distal to m2, similar in shape, relative size, and position with the tooth germ seen in Arboroharamiya and Vilevolodon. Such tooth germs in Vilevolodon and Arboroharamiya are an evidence for a prolonged replacement of the permanent incisor, which is better interpreted as derived from a diphyodont dentition of Haramiyavia but not an evidence of on-going replacement of incisors that is more primitive than the condition in Morganucodon and docodontans. In addition, in all the specimens of Vilevolodon and Arboroharamiya, the strong roots of cheek teeth extend to the lateral side of the tooth germ, which limits the space for further expansion of the tooth germ and replacement. This indicates that the prolonged replacement of the permanent incisor is probably associated with the hypertrophic p4 and specialized occlusal pattern in Vilevolodon and Arboroharamiya in which P4/p4 function has been emphasized, whereas the permanent incisor and ultimate molars became less important in food processing; thus, their replacement and eruption have been prolonged, delayed, or even suppressed.
Phylogenetic implications However, presence of the diphyodont dentition alone is not diagnostic for mammals. This is because a diphyodont dentition exists not only in mammals but also in stem mammaliaforms, such as Morganucodon and docodonts (Mills, 1971; Parrington, 1971, 1973, 1978; Kermack et al., 1973, 1981; Gow, 1985; Crompton and Luo, 1993; Nowotny et al., 2001; Rougier et al., 2014; Luo, 2004; Luo et al., 2015a, 2017; Meng et al., 2015; Panciroli et al., 2017), although there may be more than one replacement for the upper canine of Haldanodon (Martin et al., 2010b). In addition, there is evidence for resorption of anterior postcanines in mature individuals of Morganucodon, Eozostrodon and Dinnetherium (Mills, 1971; Parrington, 1971; Crompton and Luo, 1993). Sinoconodon appears to be the only stem mammaliaform that has polyphyodont tooth replacement (Luo, 1994; Luo and Wu, 1994, 1995; Zhang et al., 1998; Luo et al., 2004). This conclusion is based on different size and degree of eruption of functional teeth in a collection of Sinoconodon skulls that presumably represents various ontogenetic stages (Crompton and Luo, 1993; Zhang et al., 1998). Of the 11 examined specimens with the skull length ranging from 22 mm to 62 mm, Zhang et al. (1998) found only two that displayed convincing evidence of incisor replacement. To gather additional evidence, we have CT scanned one specimen of Sinoconodon (IVPP V 4726) that has a skull length of 35.5 mm (slightly broken at the posterior end); the diastema between the upper canine and first post-canine locus is 5.2 mm, which indicate that this individual is probably belong to the middle-late ontogenetic stage (Stage E-F) of Zhang et al. (1998). If Sinoconodon did have polyphyodont teeth, V 4726 would have tooth germs in the jawbones. However, our CT scan found no tooth germ in any tooth locus of the specimen; this raises doubt about the polyphyodont replacement of Sinoconodon.
On the other hand, tooth replacement is also complex among mammals. For instance, the molariform teeth of eutriconodonts show replacement (Jenkins and Schaff, 1988; Kielan-Jaworowska and Dashzeveg, 1998; Wang et al., 2001; Meng et al., 2003; Lopatin and Averianov, 2015; Martin et al., 2015), and some species have the entire dentition replaced and show at least three tooth generations (Lopatin and Averianov, 2015). Cheek tooth replacement is uncertain in “symmetrodontans” (Cifelli, 1999; Ensom and Sigogneau-Russell, 2000; Gill, 2004; Sweetman, 2008). In North American spalacotheriids deciduous canine and premolars were retained late in life and may never have been replaced; thus, their dentitions perhaps were monophyodont (Cifelli, 1999; Kielan-Jaworowska et al., 2004). This has been supported by the spalacolestine Lactodon from the Early Cretaceous Jehol Biota, in which there is no sign of cheek tooth replacement even though this taxon possesses deciduous-like antemolars (Han and Meng, 2016). New CT scan data (unpublished) further confirmed that there is no tooth germ at any tooth locus, including incisors and canines, of Lactodon. Thus, presence of the diphyodonty in euharamiyidans, does not constitute a sufficient evidence for the group's mammalian affinity.
Fig. 9
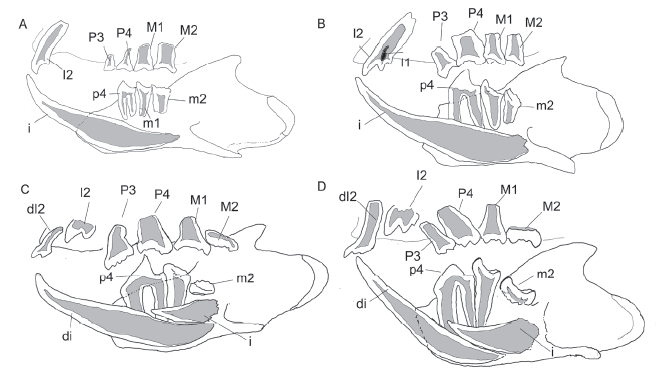 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 9Diagrams comparing reconstruction of tooth development of euharamiyidans
A. Shenshou lui; B. Xianshou linglong; C. Arboroharamiya allinhopsoni; D. A. jenkinsi The reconstruction of S. lui is based on LDNHMF 2001, WGMV-001, JZT-D 061, and JZT-CK 005; X. linglong on V 16707A; A. allinhopsoni on HG-M 017; A. jenkinsi on STM 33-9 The upper teeth of A. jenkinsi were preserved in isolation so that the reconstruction should be viewed as an interpretation of the authors, with references of tooth developments in other species. Not to scale
5 Conclusion
Tooth eruption and replacement of eight specimens of euharamiyidans are explored with assistance of micro-CT and slab CL scan, and discussion is made in comparison with specimens published in other studies. There is no evidence supporting polyphyodonty in any “haramiyidans” and tooth replacement in known specimens of euharamiyidans shows typical mammalian diphyodont replacement and definitive cheek tooth formula. The delayed development of incisor and ultimate molars, associated with enlargements of premolars, should be autapomorphic for Vilevolodon and Arboroharamiya, and represents a heterochrony in tooth development comparing to other “haramiyidans”. It is clear that tooth replacement in mammaliaforms is more complex than what we previously knew. Presence of the diphyodont dentition alone is not diagnostic for mammals. However, the reduced but definite tooth number, monophyodont molars that indicate precise tooth occlusion, and diphyodont premolars and incisors are mammal-like; these feature in combination lend strong support for euharamiyidans as a group of mammals.Acknowledgments
We thank Xie S H (IVPP) for specimen preparation; Yin P F and Hou Y M (IVPP) for CL and CT scan of the specimens. We thank Li H J, Gao Z J, Ding X H (Jizantang Paleontological Museum), Lu J H (Lande Museum of Natural History), Sun D Y (Museum of Wuyishan Mountain), and Han G (Bohai University, Hainan Tropical Ocean University) for providing studied specimens. We are grateful to Prof. Luo Z X (University of Chicago) and Averianov A O (Zoological Institute, Russian Academy of Sciences) for constructive comments during the course of the work, and Prof. Abdala F (University of the Witwatersrand) and Li C K (IVPP) for reviewing this paper and their help to improve this manuscript. This work was supported by the National Natural Science Foundation of China (Nos. 41688103, 41404022), and the Strategic Priority Research Program (B) of the Chinese Academy of Science (Grant Nos. XDB26000000, XDB18000000).参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 4]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 7]

The phylogeny of Allotheria, including Multituberculata and Haramiyida, remains unsolved and has generated contentious views on the origin and earliest evolution of mammals. Here we report three new species of a new clade, Euharamiyida, based on six well-preserved fossils from the Jurassic period of China. These fossils reveal many craniodental and post-cranial features of euharamiyidans and clarify several ambiguous structures that are currently the topic of debate. Our phylogenetic analyses recognize Euharamiyida as the sister group of Multituberculata, and place Allotheria within the Mammalia. The phylogeny suggests that allotherian mammals evolved from a Late Triassic (approximately 208 million years ago) Haramiyavia-like ancestor and diversified into euharamiyidans and multituberculates with a cosmopolitan distribution, implying homologous acquisition of many craniodental and postcranial features in the two groups. Our findings also favour a Late Triassic origin of mammals in Laurasia and two independent detachment events of the middle ear bones during mammalian evolution.
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 3]
DOIURL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 8]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 3]
DOIPMID [本文引用: 2]
Haramiyida was a successful clade of mammaliaforms, spanning the Late Triassic period to at least the Late Jurassic period, but their fossils are scant outside Eurasia and Cretaceous records are controversial. Here we report, to our knowledge, the first cranium of a large haramiyidan from the basal Cretaceous of North America. This cranium possesses an amalgam of stem mammaliaform plesiomorphies and crown mammalian apomorphies. Moreover, it shows dental traits that are diagnostic of isolated teeth of supposed multituberculate affinities from the Cretaceous of Morocco, which have been assigned to the enigmatic 'Hahnodontidae'. Exceptional preservation of this specimen also provides insights into the evolution of the ancestral mammalian brain. We demonstrate the haramiyidan affinities of Gondwanan hahnodontid teeth, removing them from multituberculates, and suggest that hahnodontid mammaliaforms had a much wider, possibly Pangaean distribution during the Jurassic-Cretaceous transition.
DOIURL [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIPMID [本文引用: 1]
Previously known only from isolated teeth and lower jaw fragments recovered from the Cretaceous and Palaeogene of the Southern Hemisphere, the Gondwanatheria constitute the most poorly known of all major mammaliaform radiations. Here we report the discovery of the first skull material of a gondwanatherian, a complete and well-preserved cranium from Upper Cretaceous strata in Madagascar that we assign to a new genus and species. Phylogenetic analysis strongly supports its placement within Gondwanatheria, which are recognized as monophyletic and closely related to multituberculates, an evolutionarily successful clade of Mesozoic mammals known almost exclusively from the Northern Hemisphere. The new taxon is the largest known mammaliaform from the Mesozoic of Gondwana. Its craniofacial anatomy reveals that it was herbivorous, large-eyed and agile, with well-developed high-frequency hearing and a keen sense of smell. The cranium exhibits a mosaic of primitive and derived features, the disparity of which is extreme and probably reflective of a long evolutionary history in geographic isolation.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 4]
DOIURL
DOIURL
DOIURL
DOIURL
DOIURL
DOIURL [本文引用: 13]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL
Five mammalian taxa based on teeth and jaw fragments are reported from a bonebed of the Late Jurassic (Oxfordian) Qigu Formation at the Liuhuanggou site in the southern Junggar Basin. The mammals recovered to date comprise a new eleutherodontid haramiyid, the docodonts <i>Dsungarodon</i> and <i>Tegotherium</i>, an undetermined amphilestid triconodont, and a new species of the stem zatherian <i>Nanolestes</i> and represent the most diverse Late Jurassic mammal assemblage of Asia. The Liuhuanggou mammal assemblage is dominated by docodonts. <i>Acuodulodon</i> Hu et al., 2007 from the upper part of the Shishugou Formation (Oxfordian) of the Wucaiwan area in the central Junggar Basin is a junior synonym of <i>Dsungarodon</i> Pfretzschner and Martin, 2005. <i>Tegotherium</i> has been reported from the Late Jurassic Shar Teeg locality in Mongolia. With the exception of the common occurrence of <i>Nanolestes</i>, the mammalian assemblage from the Late Jurassic of the Guimarota coal mine (Portugal) is quite different from that of the Late Jurassic Qigu Formation. The Guimarota assemblage is dominated by five genera of dryolestidans and several genera of multituberculates, which have not been reported from the Qigu assemblage. The known Late Jurassic mammalian assemblages of Asia are similar to the Middle Jurassic assemblages known from Asia and elsewhere in the world.
DOIURL [本文引用: 1]
DOIURL [本文引用: 8]
[本文引用: 1]
DOIPMID [本文引用: 1]
The transference of post-dentary jaw elements to the cranium of mammals as auditory ossicles is one of the central topics in evolutionary biology of vertebrates. Homologies of these bones among jawed vertebrates have long been demonstrated by developmental studies; but fossils illuminating this critical transference are sparse and often ambiguous. Here we report the first unambiguous ectotympanic (angular), malleus (articular and prearticular) and incus (quadrate) of an Early Cretaceous eutriconodont mammal from the Jehol Biota, Liaoning, China. The ectotympanic and malleus have lost their direct contact with the dentary bone but still connect the ossified Meckel's cartilage (OMC); we hypothesize that the OMC serves as a stabilizing mechanism bridging the dentary and the detached ossicles during mammalian evolution. This transitional mammalian middle ear narrows the morphological gap between the mandibular middle ear in basal mammaliaforms and the definitive mammalian middle ear (DMME) of extant mammals; it reveals complex changes contributing to the detachment of ear ossicles during mammalian evolution.
DOIURL
DOIURL [本文引用: 2]
DOIURL [本文引用: 3]
DOIURL [本文引用: 5]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIPMID
PMID
[本文引用: 1]
London: Palaeontographical Society.
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 4]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 4]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 5]
DOIURL [本文引用: 3]
DOIURL [本文引用: 1]
DOIURL [本文引用: 3]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}