 ,1,2,*
,1,2,*New skulls of ctenodactyloids from the Early Oligocene of Ulantatal, Nei Mongol, China
XU Ran-Cheng1,2,3, LI Qian,1,2,*通讯作者: *liqian@ivpp.ac.cn
收稿日期:2019-08-20网络出版日期:2020-10-20
| 基金资助: |
Corresponding authors: *liqian@ivpp.ac.cn
Received:2019-08-20Online:2020-10-20

摘要
记述了4件采自内蒙古乌兰塔塔尔早渐新世地层的近乎完整的、上下颌咬合在一起的梳趾鼠类头骨化石,基于牙齿特征将新材料归于乌兰塔塔尔丘齿鼠(Bounomys ulantatalensis
关键词:
Abstract
Keywords:
PDF (24187KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
徐冉成, 李茜. 内蒙古乌兰塔塔尔地区早渐新世梳趾鼠类头骨新材料. 古脊椎动物学报[J], 2020, 58(4): 305-327 DOI:10.19615/j.cnki.1000-3118.200413
XU Ran-Cheng, LI Qian.
Summary
Ctenodactyloids are a group of rodents that is characterized by the combination of a protrogomorphous or hystricomorphous skull and a sciurognathous mandible. The earliest known ctenodactyloid rodent is Cocomys, reported from China during the Early Eocene, and ctenodactyloids were widely distributed in East and Central Asia during the Eocene and Oligocene. The four genera of living ctenodactyloids are restricted to areas in North and East Africa. The many known Paleogene ctenodactyloids are based on fragmentary specimens, while only fragmentary skulls are known for Tataromys, Yindirtemys and Bounomys in Oligocene ctenodactyloids, which provide only limited information.Here we report four complete skulls with articulated jaws of ctenodactyloids that were found from the Early Oligocene of Ulantatal, Nei Mongol, China. These specimens were recovered from the light brownish yellow clayey siltsand of the 12-meter level of the Ulantatal Main Section. Based on the teeth morphology, the new specimens are referred to Bounomys ulantatalensis (Huang, 1985). The new materials described herein add important data on the skull morphology of Bounomys and provide new and reliable evidence for the understanding of the skull evolution of the ctenodactyloid rodents.

(Figs. 2-10; Tables 1, s1-2)
Tataromys spp. (partim) Huang, 1982, p. 340
Tataromys ulantatalensis Huang, 1985, p. 28; Russell and Zhai, 1987, p. 292
Leptotataromys gracilidens (partim) Huang, 1985, p. 32; Russell and Zhai, 1987, p. 292
Yindirtemys ulantatalensis Vianey-Liaud et al., 2006, p. 146
Holotype IVPP V 7341, a left lower jaw fragment with p4-m3, from the upper part of the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol, China.
Paratype IVPP V 7342, a lower jaw fragment with dp4-m3.
New materials IVPP V 26125-26128, four skulls with articulated jaws, from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol, China.
Emended diagnosis A medium-sized ctenodactyloid, skull length is 37-43 mm, with hystricomorphous skull and sciurognathous mandible. Differs from the Eocene ctenodactyloids Cocomys or Exmus in having a hystricomorphous skull; the anterior end of the nasal situated behind the anterior edge of the upper incisors; the posterodorsal process of premaxilla long and narrow; the dorsal branch of zygomatic process of maxilla more posterior than the ventral one; the palate comparatively narrow; the postorbital process weak; the postorbital constriction not prominent; possessing ossified external acoustic canal; the auditory bulla, mastoid and epitympanic recess more inflated, and the longitudinal length of the auditory bulla about 1/3 of the skull length; the head of malleus swollen and extending forward; the anterior process of malleus absent; the dorsal part of the supraoccipital more extended; the laminated paracondylar process of exoccipital attached to the auditory bulla posteriorly; the premaxilla possessing a laterodorsal crest; the slightly S-shaped temporal crest situated on the parietal; the nuchal crest more developed; infraorbital foramen enlarged; the incisive foramen larger, about 2/3-3/4 length of the upper diastema; the anterior margin of choana distinctly behind M3, more posteriorly; the sphenopalatine foramen above M2 and M3, enlarged and slightly posteriorly; the lacrimal foramen completely situated in the lacrimal; the optic canal merged with the sphenorbital fissure; the coronoid process of mandible reduced; the ventral masseteric crest extending forward to below ml; single mental foramen below p4; medial pterygoid fossa large and deep. Differs from the other Oligocene ctenodactyloids Tataromys or Yindirtemys in the palate comparatively wider; the major palatine foramen not as narrow as in Tataromys or Yindirtemys; the anterior margin of sphenopalatine foramen at the level of junction of M1 and M2, comparatively anteriorly; the buccinator foramen closer to the masticatory foramen. Differs from extant ctenodactylids in malleus and incus not fused.
Differs from Bounomys bohlini in the teeth being significantly larger, about 1.5 times of B. bohlini; the main cusps of the cheek teeth swollen, and lophs relatively higher and developed; on the upper molars anterior cingulum joining protoloph, mesosinus curving posterolabially; on M3 metacone usually possessing crochet connected to paracone, and metaloph tortuous; sinus separated from posterosinus; on lower molars usually having a large central basin closed by longitudinal anterior arm and transverse posterior arm of entoconid; hypoconulid more swollen, and arm of hypoconulid relatively developed.
Discussion From the Eocene to the Oligocene, the ctenodactyloids skull had undergone significant changes, and the most distinct of these changes is the zygomasseteric structure’s transition from the protrogomorphous to hystricomorphous, which is mainly manifested as follows: the infraorbital foramen is enlarged; the forming of the premaxillary laterodorsal crest; the dorsal branch of the zygomatic process of the maxilla moving posteriorly; the coronoid process of mandible is reduced; the ventral masseteric crest extending forward. These changes probably reflect the enhancement of the masticatory function of the Oligocene ctenodactyloids. In addition, the auditory bulla, mastoid and epitymanic recess are significantly inflated; the head of the malleus is swollen and extends forward, and the anterior process of the malleus is absent. The change in the structure of the auditory region may be beneficial to enhance their low-frequency hearing.
The Eocene-Oligocene transition was one of the most pronounced climatic events of the Cenozoic era, marking a dramatic shift from a ‘‘greenhouse’’ to an ‘‘icehouse’’ world, with vegetation changing from warm-humid forest to dry-temperate forest-steppe. Related studies have shown that the hystricomorphous skull is better at grinding coarse grasses than other zygomasseteric structures, and the rodents living in arid areas are usually good at low-frequency hearing, which can improve their fitness. Therefore, the changes of the zygomasseteric structure and auditory region in ctenodactyloids from the Eocene to Oligocene probably result from the global climate change during the corresponding period.
1 简介
梳趾鼠类是一类具有始啮型或豪猪型头骨及松鼠型下颌的啮齿动物,因其现生种类的脚趾上具有用于梳理毛发的梳状硬毛而得名。现生种类仅有梳趾鼠科(Ctenodactylidae), 该科包括4属5种,高阶元分类上多将该科和相关化石种类归入梳趾鼠超科(Ctenodactyloidea)。一般认为梳趾鼠超科在早始新世初期起源于亚洲,目前公认最早的化石代表是发现于中国湖南衡东岭茶组的岭茶钟健鼠Cocomys lingchaensis (李传夔,1975; Li et al., 1989; Wible et al., 2005; Li and Meng, 2015; 李茜、童永生,2019)。梳趾鼠科是在中始新世时从梳趾鼠超科中分出的一支,现有的系统发育分析普遍认同该科为一单系(Vianey-Liaud et al., 2010; Li and Meng, 2015)。梳趾鼠类作为始新世以来动物群中的优势类群,凭借其数量多、分布广、多样性高、演化快等特点,成为划分与对比地层的“关键化石”(Bohlin, 1946; Wood, 1977; 黄学诗,1985; Qiu and Qiu, 1995; 童永生,1997; Wang, 1997; Vianey-Liaud et al., 2006; Schmidt-Kittler et al., 2007; Rodrigues et al., 2014; Oliver and Daxner-H?ck, 2017)。长期以来,梳趾鼠类的头骨化石一直比较匮乏,在梳趾鼠科中仅对个别属种有少量的关于头骨形态的报道和研究(Bohlin, 1946; Wang, 1997)。目前对梳趾鼠类化石的认识主要集中在牙齿形态上,对头骨的形态及其演化过程还很不清楚。近年来,在内蒙古阿拉善左旗乌兰塔塔尔地区渐新世地层中发现了一些保存完好、上下颌咬合在一起的梳趾鼠类头骨,这些材料为全面了解这一类群头骨的形态特征,以及探讨梳趾鼠类头骨的演化过程提供了新的、可靠的证据。
新材料均来自内蒙古阿拉善盟阿拉善左旗乌兰塔塔尔地区,该地区位于阿左旗巴彦浩特镇北北西方向约42 km (图1)。该地点最初由宁夏地质局区测队地质填图时发现,之后多位****对这一地区的化石和地层做了相关研究和报道(黄学诗,1982, 1985, 1998; 王伴月、王培玉,1991; Vianey-Liaud et al., 2006, 2010)。最近,Zhang et al. (2016)建立了该区域内渐新世地层序列,同时对化石产出层位做了详细记录,为进一步的生物地层划分和对比提供了基础。Zhang et al. (2016)实测的4条剖面从东向西依次为:克克阿木剖面、上井剖面、乌兰塔塔尔主剖面和稍增图剖面, 新材料均出自乌兰塔塔尔主剖面12 m处的浅棕黄色粉砂岩中,对应乌兰塔塔尔组中下部,时代为早渐新世。
Fig. 1
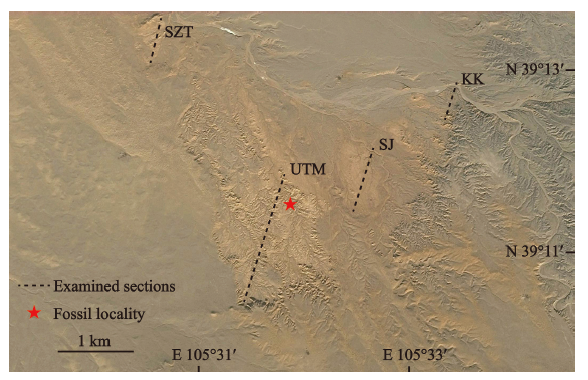 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Google map showing the four sections in the Ulantatal gulley
bbreviations: KK. Kekeamu section 克克阿木剖面;SJ. Shangjing section 上井剖面;SZT. Shaozengtu section 稍增图剖面;UTM. Ulantatal Main Section 乌兰塔塔尔主剖面(After Zhang et al., 2016)
2 材料和方法
新材料为4件上下颌咬合在一起的梳趾鼠类头骨,进行对比的标本包括:Cocomys lingchaensis头骨3件(IVPP V 7402, V 7403, V 7404>); Exmus mini带下颌的头骨2件(V 7429, V 7430); Tataromys plicidens带下颌的头骨1件(V 10534.1); Yindirtemys ambiguous带下颌的残破头骨1件(RV 46001), 腭骨1件(Sh.785); Bounomys bohlini带下颌的残破头骨1件(V 7348), 残破头骨1件(V 7349); Sayimys obliquidens带下颌的残破头骨1件(T.b. 279a, b)。由于未获取到现生梳趾鼠Ctenodactylus gundi的相关标本,其形态信息主要来自Schrenk (1989)的插图及相关描述。头骨各构造的描述主要基于Wible et al. (2005), 同时也参考了Wahlert (1974)和Meng et al. (2003)的部分术语,牙齿形态描述采用Wang (1997)的术语。头骨测量的定位和方法采用了王伴月、邱占祥(2018)的建议。头骨测量使用数码显示游标卡尺,牙齿测量使用体视显微镜Zeiss Discovery. V8, 测量精度均为0.01 mm。电镜照片使用中国科学院脊椎动物演化与人类起源重点实验室的JSM-00 SEM拍摄。头骨内部形态数据通过该实验室的225 Micro CT进行高精度扫描获取,扫描和重建方法主要参考Wang et al. (2019), 扫描电压100 kV, 电流100 μA, 分辨率29.01-31.37 μm。三维模型使用Mimics Research 18.0重建。
缩写 IVPP, 中国科学院古脊椎动物与古人类研究所;IVPP RV, IVPP标本馆重新研究过的脊椎动物化石标本编号;IVPP V, IVPP标本馆脊椎动物化石编号;Sh, 中瑞西北考察(1927-1933年)期间瑞典古生物学家博格·步林(Birger Bohlin)在甘肃沙拉果勒地区采集的化石标本编号;T.b, 步林在甘肃塔奔布鲁克(Taben-buluk)地区采集的化石标本编号。
3 系统描述

(图2-10; 表1, 附表1-2)
Tataromys spp. (partim) Huang, 1982, p. 340
Tataromys ulantatalensis Huang, 1985, p. 28; Russell and Zhai, 1987, p. 292
Leptotataromys gracilidens (partim) Huang, 1985, p. 32; Russell and Zhai, 1987, p. 292
Yindirtemys ulantatalensis Vianey-Liaud et al., 2006, p. 146
正模 IVPP V 7341, 一段左下颌骨具p4-m3。
副模 IVPP V 7342, 一段左下颌骨具dp4-m3。
新标本 IVPP V 26125-26128, 4件上下颌咬合的头骨。
产地和层位 内蒙古阿拉善左旗乌兰塔塔尔,下渐新统乌兰塔塔尔组。
修订特征 中等大小的梳趾鼠类,头骨枕鼻长37-43 mm, 具有豪猪型头骨及松鼠型下颌。头骨顶部较平;鼻骨前缘位于上门齿前缘之后;前颌骨背后突窄长;上颌颧突背支较腹支更靠后;腭部相对较窄;眶后突存在但较弱;眶后缩不明显;有骨质外耳道;鼓上隐窝、乳突及听泡均膨大,听泡纵长约占头长的1/3; 锤骨头向前膨大,锤骨前突退化缺失,锤骨与砧骨不愈合;上枕骨颅顶部分面积较大;外枕骨髁旁突呈薄片状贴附于听泡后面;前颌骨具侧背嵴;颞嵴位于顶骨且呈S形;项嵴发达;眶下孔较大;门齿孔亦较大,约为上齿隙长的2/3-3/4; 后鼻孔位于M3之后,较靠后;蝶腭孔较大,位于M2和M3背侧,位置稍靠后;泪孔位于泪骨内;视神经孔和蝶眶裂相融合。下颌骨冠状突较弱,腹侧咬肌嵴更靠前,向前延伸至m1下方,颏孔仅1个,且位于p4下方,翼内肌窝大而深。
相比渐新世其他梳趾鼠类Tataromys和Yindirtemys, Bounomys腭部相对较宽,且大腭孔没有Tataromys和Yindirtemys中那么狭长;蝶腭孔稍靠前,前缘位于M1/M2交界处背侧;颊肌神经孔与咬肌神经孔距离较近。
Bounomys ulantatalensis和B. bohlini的不同之处在于:个体明显大,臼齿大小约为后者的1.5倍;颊齿主尖也更膨大,齿脊相对较高;上臼齿前边尖不孤立,前齿带与原脊相连,且中凹弯向后颊侧;M3的后尖常具前刺连接到前尖,后脊较曲折;后凹与内凹相分隔;下臼齿由发达的下内尖前臂和横向的后臂封闭成大的中央盆(凹); 下次小尖更膨大,下次小尖臂较发育。
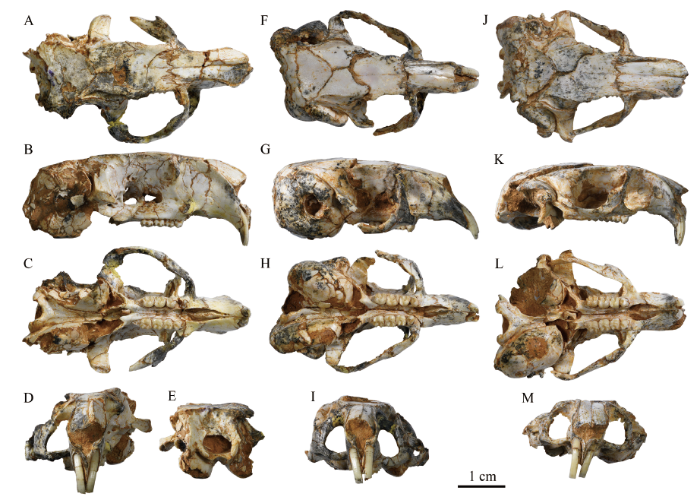
描述 4件标本总体保存较好,尤其是V 26128, 除了右侧听泡和左侧眶部稍有破损、顶骨稍有破损变形、两侧鼓上隐窝隆起部破损变形外,整个头部骨骼保存近乎完整。而V 26125左侧眶部保存较好,V 26126右侧鼓上隐窝隆起保存较好,V 26127右侧听小骨保存相对较好。因此,主要依据V 26128, 并参考其他标本对B. ulantatalensis的头骨形态进行描述。另外,头骨各枕骨接近完全愈合,以及颊齿都已经为恒齿表明这4件标本均为成年个体。
鼻骨(nasal) 4件标本都保存了鼻骨,其中V 26128保存最为完好。背面观(图2A, B),左右鼻骨合起来近似铲形,向前渐宽,在鼻-前颌骨缝的最前端达到最宽;鼻-额缝呈锯齿状。另外,鼻骨背面布满细丝状痕迹,可能与吻部肌肉附着有关。前面观(图3A, B), 外鼻孔(external nasal aperture)较大,呈橄榄形。
Fig. 2
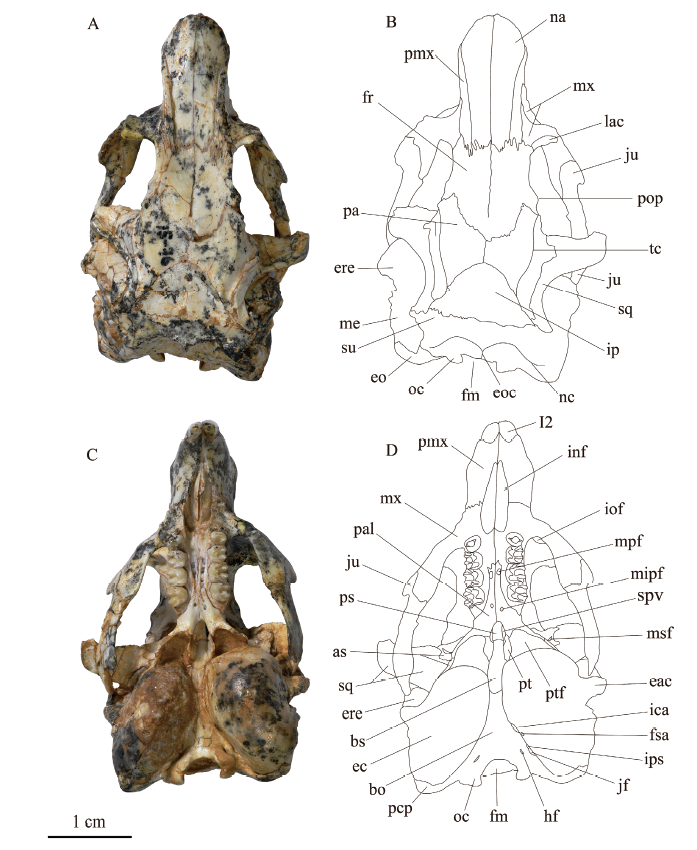 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Skull of Bounomys ulantatalensis (IVPP V 26128) from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol in dorsal (A, B) and ventral (C, D) views
Abbreviations: as. alisphenoid 翼蝶骨; bo. basioccipital 基枕骨; bs. basisphenoid 基蝶骨;
eac. external acoustic canal 外耳道; ec. ectotympanic 外鼓骨; eo. exoccipital 外枕骨; eoc. external occipital crest 枕外嵴; ere. epitympanic recess eminence 鼓上隐窝隆起; fm. foramen magnum 枕骨大孔;
fr. frontal 额骨; fsa. foramen for stapedial artery 镫骨动脉孔; hf. hypoglossal foramen 舌下神经孔;
I2. upper second incisor 上第二门齿; ica. foramen for internal carotid artery 颈内动脉孔; inf. incisive foramen 门齿孔; iof. infraorbital foramen 眶下孔; ip. interparietal 间顶骨; ips. foramen for inferior petrosal sinus 岩下窦沟孔; jf. jugular foramen 颈静脉孔; ju. jugal 颧骨; lac. lacrimal 泪骨; me. mastoid exposure 岩骨乳突暴露部; mipf. minor palatine foramen 小腭孔; mpf. major palatine foramen 大腭孔; msf. masticatory foramen 咬肌神经孔; mx. maxilla 上颌骨; na. nasal 鼻骨; nc. nuchal crest 项嵴; oc. occipital condyle 枕髁; pa. parietal 顶骨; pal. palatine 腭骨; pcp. paracondylar process 髁旁突; pmx. premaxilla 前颌骨;
pop. postorbital process 眶后突; ps. presphenoid 前蝶骨; pt. pterygoid 翼骨; ptf. pterygoid fossa 翼窝;
spv. sphenopalatine vacuity 蝶腭空缺; sq. squamosal 鳞骨, su. supraoccipital 上枕骨; tc. temporal crest 颞嵴
Fig. 3
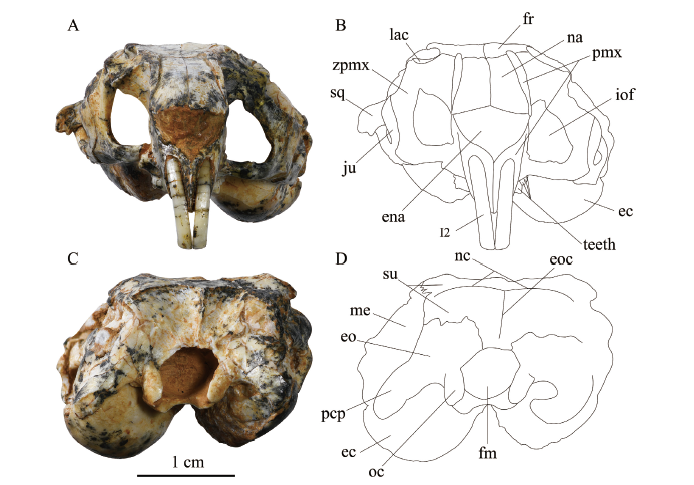 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Skull of Bounomys ulantatalensis (IVPP V 26128) from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol in rostral (A, B) and occipital (C, D) views
Abbreviations: ena. external nasal aperture 外鼻孔; zpmx. zygomatic process of maxilla 上颌颧突;other abbreviations see
前颌骨(premaxilla) 4件标本都保存了前颌骨,其中V 26128保存最完好。背面观(图2A-B), 前颌骨面突(facial process of premaxilla)具有细长的后背突(posterodorsal process of premaxilla), 向后延伸到额-鼻缝附近或稍后一点,后端微微拓宽;背后突在吻部背外侧形成明显的侧背嵴(premaxillary laterodorsal crest)。
上颌骨(maxilla) 4件标本都有保存,其中V 26128右侧上颌骨保存最为完好。上颌骨具面突、颧突、腭突以及眶突。V 26128和V 26125侧面观(图4), 上颌骨面突构成了后1/3吻部侧壁;颧突背支位于腹支背后方;上颌眶突构成了眼眶的底板和前腹壁;眶突后缘分别与泪骨、额骨和腭骨相接;蝶腭孔(sphenopalatine foramen)呈卵圆形,较大,由上颌骨、额骨和腭骨共同围成,蝶腭孔左右相通,用于传导上颌神经蝶腭支和蝶腭血管(Wahlert, 1974); 蝶腭孔之前的小孔是前齿槽孔(anterior alveolar foramen)。前面观(图3A, B), 眶下孔较大,高度适中,大致呈正立的圆角三角形,完全由上颌骨围成。腹面观(图2C, D), 上颌腭突与前颌骨围成一对较大的门齿孔,合起来呈水滴形;腭沟从门齿孔向后延伸且逐渐变浅,到大腭孔(major palatine foramen)附近消失。
Fig. 4
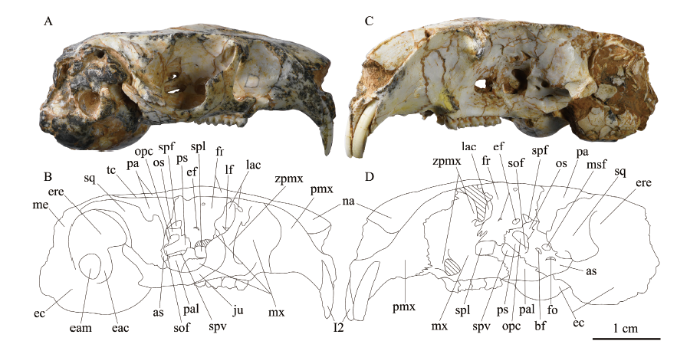 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Skulls of Bounomys ulantatalensis from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol
A, B. IVPP V 26128 in right lateral view; C, D. V 26125 in left lateral view
Abbreviations: bf. buccinator foramen 颊肌神经孔; eam. external acoustic meatus 外耳门;
ef. ethmoidal foramina 筛孔; fo. foramen ovale 卵圆孔; lf. lacrimal foramen 泪孔; opc. optic canal 视神经孔; os. orbitosphenoid 眶蝶骨; sof. sphenorbital fissure 蝶眶裂; spf. sphenofrontal foramen 蝶额孔; spl. sphenopalatine foramen 蝶腭孔; other abbreviations see Figs. 2, 3
腭骨(palatine) V 26128的腭骨水平突和垂直突保存最好。腹面观(图2C, D), 左右两块腭骨的水平突合起来大致呈楔形夹在两侧齿槽之间,前缘与M1次尖相对,后缘远在M3之后;后鼻孔切迹呈U形,无后鼻棘(posterior nasal spine); 腭骨后缘往后延伸形成翼窝(pterygoid fossa)前腹侧壁;大腭孔与M2原尖相对,小腭孔(minor palatine foramen)与M3后缘相对或稍后;另外,V 26125在大小腭孔之间存在数量众多的小孔,可能是滋养孔。侧面观(图4), 腭骨垂直突参与构成蝶腭孔的部分后缘和腹缘及蝶眶裂(sphenorbital fissure)的外腹侧缘。
泪骨(lacrimal) V 26128的右侧泪骨保存最好,具面突和眶突。背面观(图2A, B), 面突较细窄,位于上颌颧突背支根部后缘,内侧与额骨相接,前面与上颌骨相接。侧面观(图4A, B), 泪骨眶突较大,大致呈卵圆形;泪骨面突、眶突及上颌颧突根部在眼眶背前内侧共同围成一个凹窝;泪孔(lacrimal foramen)卵圆形,开口朝上,完全位于泪骨内,但不在前述的凹窝内,而是位于泪骨偏下部。
颧骨(jugal) 颧骨在V 26128中保存最为完好,呈不规则薄骨片,分为颧骨主体、背突、后腹突和后突。背面观(图2A, B), 颧骨主体较直。侧面观(图4A, B), 颧骨前段叠覆在上颌颧突之上,不与泪骨连接;背突位于颧骨前端,较弱;后腹突位于颧骨后腹部,朝向后腹侧;后突是颧骨的最后端,呈三棱锥状,较尖锐,背面与鳞骨颧突(zygomatic process of the squamosal)腹面外侧部分毗连。
额骨(frontal) 额骨背面部分在V 26125和V 26127中保存较好,额骨眶部在V 26125的左侧和V 26128的右侧保存较好。V 26125背面观(图5A), 额骨背面前缘由内向外分别与鼻骨、前颌骨、上颌颧突和泪骨面突相接;后缘与顶骨相接,骨缝大致呈U形。眶后突(postorbital process)位于顶-额缝附近,不发达。V 26125侧面观(图4C, D), 从前往后,额骨眶部分别与泪骨、上颌骨、腭骨、眶蝶骨、顶骨和翼蝶骨相接触;大致在视神经孔(optic canal)背后方,额骨、眶蝶骨、翼蝶骨三者在顶骨眶突前缘上汇聚于一点。位于眶部正中的小孔是前筛孔(rostral ethmoidal foramen)。前筛孔之后,视神经孔背前方有一个较大的半圆形孔,应当是后筛孔(caudal ethmoidal foramen)。紧邻后筛孔之后,在眶蝶骨和额骨骨缝间有一个三角形小孔,可能是蝶额孔(sphenofrontal foramen)。蝶额孔和后筛孔通常相融合,只在V 26125左侧是分隔的。另外,额骨还构成了蝶腭孔的整个背缘和部分后缘。
Fig. 5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Skulls of Bounomys ulantatalensis from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol
A-E. IVPP V 26125 in dorsal (A), left (reversed) (B), ventral (C), rostral (D), and occipital (E) views;
F-I. V 26126 in dorsal (F), right (G), ventral (H), and rostral (I) views;
J-M. V 26127 in dorsal (J), right (K), ventral (L) and rostral (M) views
顶骨(parietal) V 26128顶骨整体保存较好。背面观(图2A, B), 两块顶骨合起来大致呈蝶形,颞嵴(temporal crest)将顶骨分为背面部分和侧壁部分,背面部分较为扁平,侧壁部分向外侧倾斜。顶骨后外侧突逐渐尖灭于间顶-鳞骨缝中,不与枕骨接触。从背面可看到两条明显的嵴,一条是颞嵴,大致呈S形曲线,自额骨眶后突,向后延伸至顶骨后侧突末端;另一条是位于顶-鳞缝上的嵴,同样大致呈S形,自鳞骨颧突后缘背侧,向后延伸至顶骨后侧突末端与颞嵴汇合。
间顶骨(interparietal) 4件头骨间顶骨保存都较好。背面观(图2A, B), 间顶骨较大,呈三角形,位于顶骨之后。两侧缘(人字缝(lambdoid suture))微凸,后缘较曲折。
翼骨(pterygoid) V 26128腹面观(图2C, D), 翼骨参与构成翼窝内壁,前部叠覆在腭骨垂直突内侧;背缘与基蝶骨连接;腹缘向后伸出翼钩(pterygoid hamulus), 与听泡前突(rostral process of bulla)相连;翼骨和听泡之间存在半月形空缺,沟通了翼窝和基咽管(basipharyngeal canal)。
前蝶骨(presphenoid) V 26128腹面观(图2C, D), 前蝶骨呈短柱状,两侧是较大的蝶腭空缺(sphenopalatine vacuity), 前蝶-基蝶骨缝位于蝶腭空缺后缘之前。侧面观(图4A, B), 前蝶骨前缘与腭骨、眶蝶骨相接,但与眶蝶骨的骨缝已经愈合;前蝶孔(presphenoid foramen)较小,位于前蝶骨前端,该孔在V 26125右侧最明显。
眶蝶骨(orbitosphenoid) V 26125左侧观(图4C, D), 眶蝶骨前腹缘与腭骨相接,背前缘与额骨相接,后缘叠覆在翼蝶骨上,其中腹缘构成视神经孔(optic canal)和蝶眶裂(sphenorbital fissure)的背缘;视神经孔整体呈卵圆形,开口朝向前外腹侧,且左右视神经孔相通;毗邻于视神经孔后外腹侧的是蝶眶裂,蝶眶裂和视神经孔之间没有骨板将二者相分隔,只在背侧缘有一微弱突起代表着二者的分界。
基蝶骨(basisphenoid) V 26128腹面观(图2C, D), 基蝶骨腹面呈狭长的矩形,前半段腹面较隆凸,后半段较凹;基蝶骨侧缘前半段与翼骨基部相接,后半段与听泡内壁相毗连。侧缘还参与构成了蝶腭空缺和翼骨-听泡空缺的背内侧缘。
翼蝶骨(alisphenoid) V 26128腹面观(图2C, D), 翼窝较大且深,整体接近三角形,位于听泡之前,由腭骨、翼蝶骨、翼骨及听泡前壁包围;外翼嵴(ectopterygoid crest)由腭骨和翼蝶骨共同组成,外翼嵴外表面微凸且较为粗糙;内翼嵴(entopterygoid crest)由腭骨和翼骨构成;CT扫描显示,翼蝶骨颅基部后侧存在较大的中破裂孔(middle lacerate foramen)。V 26128和V 26125侧面观(图4), 翼蝶骨近似梯形,其前缘竖直,上半段与眶蝶骨相接,下半段构成蝶眶裂外侧壁;后缘微凸,大部分叠覆在听泡前外侧壁上。翼蝶骨上具有明显的3个孔:卵圆孔(foramen ovale)、颊肌神经孔(buccinator foramen)和咬肌神经孔(masticatory foramen)。卵圆孔位于外翼嵴外侧,卵圆形,开口朝向腹侧;咬肌神经孔和颊肌神经孔都位于卵圆孔背前方,其中偏外侧的是咬肌神经孔,偏内侧的是颊肌神经孔,这两个孔相邻且呈卵圆形。另外,卵圆孔内壁上存在一个较浅的切迹,从形态来看可能有脉管从此处横向通过,可能代表了副卵圆孔(foramen ovale accessorius)的遗迹,但已和卵圆孔融合为一个大孔。从前侧观,蝶眶裂外侧壁和腹侧壁相交处,有一个小孔,CT扫描显示该孔从蝶眶裂通向翼窝,可能是翼蝶管前口(rostral opening of alisphenoid canal)。
鳞骨(squamosal) 从4件标本侧面观(图4; 5B, G, K), 鳞骨前缘与顶骨和翼蝶骨相接,三者相交处即鳞骨最前端,鳞骨无关节后突。背面观(图2A, B), 鳞骨最后端尖灭于枕骨-鼓上隐窝隆起部缝中,鳞骨颧突整体接近鸟喙状,鳞骨表面没有明显的孔。腹面观(图2C, D; 5C, H, L), 下颌关节窝较宽且浅,其内侧面由鳞骨颧突和鼓上隐窝隆起部前壁(epitympanic recess eminence of petrosal)共同构成,外侧面由颧骨后突构成。
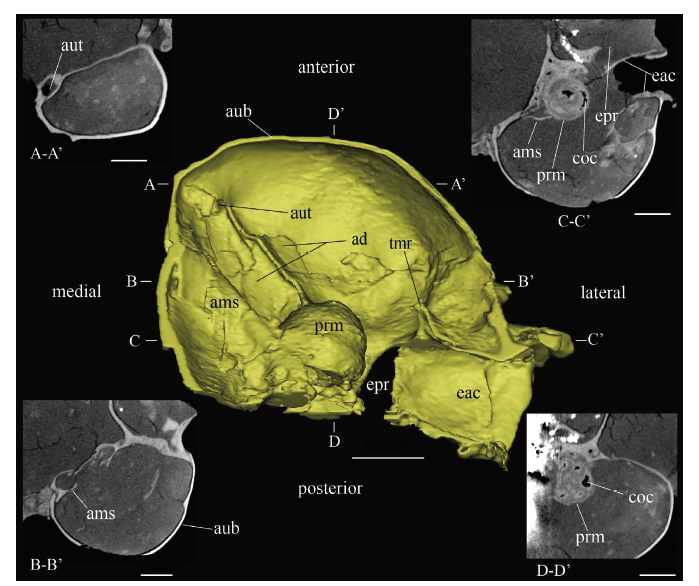
岩骨(petrosal) V 26126右侧观(图5G)并结合CT影像观察,鼓上隐窝(epitympanic recess)非常膨大,呈囊状紧贴在外耳门背侧,内部无隔板,后紧贴乳突。乳突(mastoid exposure of petrosal)亦膨大为较大的空腔,暴露面亦较大,横跨头骨枕面和侧面。乳突与鼓上隐窝隆起表面以明显的沟相隔,内部则以骨板相隔。乳突暴露面向下与听泡壁自然过渡,无明显界线,但二者在内部却存在矮嵴相隔。结合CT影像,可以获得听泡内岩骨的部分形态信息:V 26127腹侧观(图6), 岬(promontorium)呈圆丘形;左侧耳蜗(cochlea)从基圈到蜗顶顺时针旋转,匝数近3圈,蜗轴(modiolus)朝向前外侧,蜗轴与矢状面的夹角为32.7°, 与水平面的夹角为0°; 岬前内侧和腹侧分别为前内侧架(anteromedial shelf)和前凹(anterior depression), 前内侧架前后狭长,向腹侧凹;前凹较宽大,其腹面中间有一条斜嵴向前内侧延伸,将前凹分为内侧和外侧部分,该斜嵴和前内侧架在听泡前内侧角附近相会,并在相会处有咽鼓管(auditory tube)前口通向鼓室(tympanic cavity)之外。
Fig. 6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 63D-reconstruction and some CT images of partial structures of the left middle ear of Bounomys ulantatalensis (IVPP V 26127) from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol
The yellow model in the center of the picture is the ventral view of the middle ear cavity (some structures that obstruct the observation have been removed) and is surrounded by CT images of different parts of the middle ear cavity. A-A’. the transverse plane through the opening of the auditory tube; B-B’. the transverse plane through the anteromedial shelf; C-C’. the transverse plane through the promontorium;
D-D’. the sagittal plane through the promontorium Abbreviations: ad. anterior depression 前凹; ams. anteromedial shelf 前内侧架; aub. auditory bulla 听泡;
aut. auditory tube 咽鼓管; coc. cochlear canal 蜗管; eac. external acoustic canal 外耳道; epr. epitympanic recess 鼓上隐窝; prm. promontorium 岬; tmr. tympanic ring 鼓环。Scale bars = 2 mm
外鼓骨(ectotympanic) 听泡由外鼓骨单独构成,V 26126右侧观(图5G), 听泡很大,长度占头长的1/3, 高度接近头高;外鼓骨前部与岩骨鼓上隐窝前壁、鳞骨及翼蝶骨相接,后部与乳突外壁自然过渡;有明显的骨质外耳道(external acoustic canal)连通鼓室和外耳门(external acoustic meatus), 且外耳道前腹侧明显突出于听泡外侧,另外在外耳道突出部的腹侧有一个小孔或缺口,应为鼓窗(tympanic fenestra), 外耳门相对于膨大的听泡较小。V 26127 (图5L)和V 26128 (图2C, D)腹面观,极为膨大的一对听泡向前汇聚,几乎挤占了颅底的大部分空间;听泡背前内侧角的小孔是咽鼓管(欧氏管)前口,由岩骨及外鼓骨围成;在听泡内侧壁中部有一个小孔,朝向后腹侧,应为镫骨动脉孔(foramen for stapedial artery)。CT影像显示,听泡内壁光滑,无隔板。
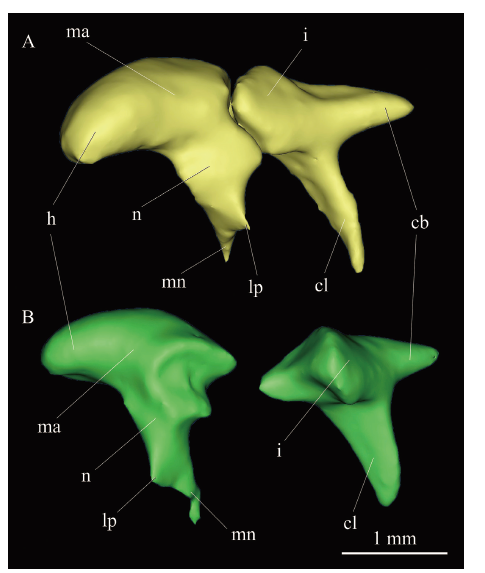
听小骨(ossicles) V 26126保存了右侧锤骨和砧骨(图5G), V 26127和V 26128保存了左侧的锤骨和砧骨(图7), 但发生了较大的位移,V 26127的锤骨和砧骨呈分离状态掉落在鼓上隐窝空腔内,V 26128的呈关联状态掉落在听泡内。由于保存的原因,所有标本均未发现镫骨(stapes)。CT影像显示,V 26126, V 26127和V 26128的锤骨和砧骨不愈合。
Fig. 7
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 73D-reconstruction of the ossicles of Bounomys ulantatalensis from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol
A. IVPP V 26128, lateral view of left malleus and incus in articular with the head of malleus forward; B. V 26127, posterolateral view of left malleus and anterolateral view of left incus Abbreviations: cb. crus breve 砧骨短脚; cl. crus longum 砧骨长脚; h. head of malleus 锤骨头; i. incus砧骨; lp. lateral process of malleus 锤骨侧突;
ma. malleus 锤骨; mn. manubrium 锤骨柄;
n. neck of malleus 锤骨颈
锤骨(malleus) 位于砧骨前外侧,近似T形,具有头、颈、柄和侧突4个基本结构(图7)。锤骨头向前膨大;颈部较粗长;侧突位于锤颈和锤柄相接处外侧,呈三角形薄骨片;无前突;在锤骨颈接近砧骨关节面内侧缘有一个小突起,呈短茎状,朝向内侧,可能是肌突(muscular process, 仅见于V 26127); 侧突之下是锤柄,呈细茎状,远端微微弯向外侧。
砧骨(incus) 位于锤骨后内侧,具有砧骨体、短脚和长脚(图7)。砧骨体与锤骨头相关联,锤骨关节面呈马鞍状,由两个锐角相交的面构成;短脚由砧骨体继续向后延伸所形成,呈锥形;长脚向腹侧突出,呈板状楔形。
枕骨复合体(occipital complex) 单一的枕骨复合体由不成对的基枕骨(basioccipital), 上枕骨(supraoccipital)和成对的外枕骨(exoccipital)愈合形成。现生的Ctenodactylus gundi的枕骨到成年阶段才愈合为一个整体(Schrenk, 1989), 4件新化石标本各枕骨也接近愈合,可能代表了成年个体。
V 26128和V 26127腹面观(图2C, D; 5L), 基枕骨整体近似Y形,较窄长;基枕骨腹面前半段呈纵向凹沟,后半段则隆凸形成龙骨突;枕髁与髁旁突(paracondylar process)之间存在切迹,枕动脉可能从此穿过(Wible et al., 2005); 基枕骨后部是枕髁,完全由基枕骨构成;枕髁间切迹呈U形,内缘存在凹缺,将枕髁分为枕叶和腹叶;基枕骨与外鼓骨之间是鼓枕裂(tympano-occipital fissure), 其内大致可以识别出4个狭缝状的孔,从后往前依次是:迷走神经耳支孔(foramen for auricular branch of vagus nerve, 图中不可见), 颈静脉孔(jugular foramen), 岩下窦沟孔(foramen for inferior petrosal sinus)和颈内动脉孔(foramen for internal carotid artery), 其中颈静脉孔和岩下窦沟孔在外部相融合,颈内动脉孔位于镫骨动脉孔背侧;基枕骨腹面紧靠枕髁前外侧的卵圆形孔是舌下神经孔(hypoglossal foramen), 较小。
V 26128和V 26127顶面观(图2A, B; 5J), 上枕骨顶面部分较为宽大扁平;前缘与间顶骨和鳞骨相接,前外侧角与鼓上隐窝隆起部存在很短的点状接触;外侧缘与外鼓骨相接;颅顶后缘是发达的项嵴(nuchal crest), 其正中位置明显后突,两侧则向前微微弯曲。
V 26128枕面观(图3C, D), 枕骨大孔位于枕部腹侧,卵圆形,其背缘由上枕骨构成,侧缘和腹缘分别由外枕骨和基枕骨构成;上枕骨和外枕骨的骨缝从枕骨大孔背外侧延伸到枕部外缘中部;外枕骨与基枕骨的骨缝愈合程度较深,不易辨认;枕髁关节旋转轴向与水平面的夹角较大;上枕骨枕面部分近似扇形;枕外嵴(external occipital crest)垂直,较发达,与项嵴相连,枕外嵴两侧是凹窝。但V 26125不同,其枕外嵴非常微弱,且两侧具发达的斜嵴,呈“八”字状,斜嵴之间是明显的凹窝;另外V 26125的上枕骨上存在数量较多的滋养孔,也与其他三件头骨不同(图5E)。4件标本的外枕骨髁旁突呈窄长的薄片状,贴附在听泡后表面(图3C, D)。
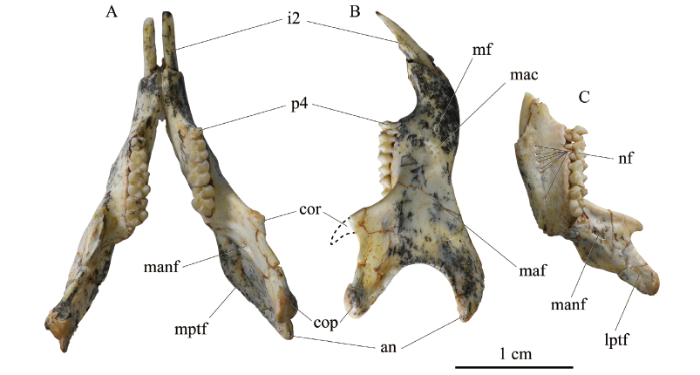
下颌骨(mandible) 4件新标本都保存了下颌骨,其中V 26128保存最完整,虽然冠状突缺失,但结合其他标本,基本可以复原整个下颌形态。下颌为松鼠颌型。V 26125-26128颊侧观(图8B; 9B, D, F), 下颌体(mandible body)较宽且底缘弯曲,下颌支(ramus)也较宽。齿隙(diastema)背缘较凹且长,下颌齿隙长与下颌骨长度之比平均为0.22 (附表2)。颏孔(mental foramen)位于p4下方,更靠近下颌体腹缘。冠状突(coronoid process)较细弱,其前缘与齿列纵轴的夹角约为45°。髁突(condyloid process)较发达;髁突关节面呈半圆形,较凸;髁突微低于冠状突。角突(angular process)较为发达。咬肌窝(masseteric fossa)较浅。腹侧咬肌嵴(ventral masseteric crest)明显且向前延伸到m1之下,背侧咬肌嵴(dorsal masseteric crest)较弱且短。V 26128舌侧观, 翼内肌窝(medial pterygoid fossa)呈半月形,较大且深,位于髁突腹缘和角突腹缘之间;V 26125舌侧观(图8C)翼外肌窝(lateral pterygoid fossa)较浅且小,位于髁突内侧面。下颌孔(mandibular foramen)开口朝后,位于冠状突下方。下颌体内侧面有一排滋养孔。
Fig. 8
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 8Mandibles of Bounomys ulantatalensis from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol
A. IVPP V 26128 in occlusal view; B. the right mandible of V 26128 in labial view;
C. the right mandible of V 26126 in lingual view
Abbreviations: an. angular process 角突; cop. condyloid process 髁突; cor. coronoid process 冠状突;
i2. lower second incisor 下第二门齿; lptf. lateral pterygoid fossa 翼外肌窝; mac. masseteric crest 咬肌嵴;
maf. masseteric fossa 咬肌窝; manf. mandibular foramen 下颌孔; mf. mental foramen 颏孔; mptf. medial pterygoid fossa 翼内肌窝; nf. nutricium foramina 滋养孔; p4. lower fourth premolar下第四前臼齿
Fig. 9
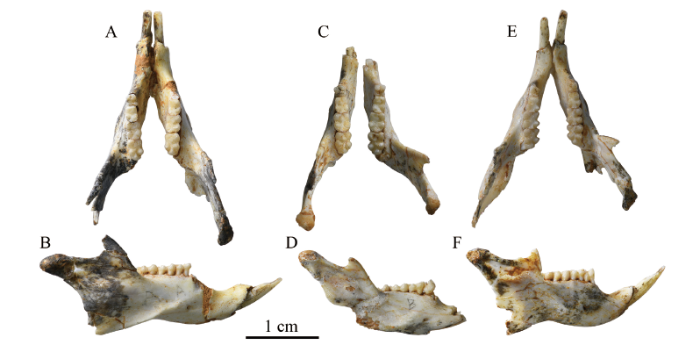 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 9Mandibles of Bounomys ulantatalensis from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol
A, C, E. mandibles of IVPP V 26125 (A), V 26126 (C), V 26127 (E) in occlusal view;
B, D, F. the right mandibles of V 26125 (B), V 26126 (D), V 26127 (F) in labial view
上门齿(I2)横断面呈圆角三角形,上门齿弧度比下门齿大,CT显示上门齿向后延伸到了P4的背侧;釉质层覆盖整个唇面和内、外侧面的1/3,釉质层表面具有密集的发丝状褶皱(图3A, B; 4A, B)。
上颊齿齿列P4到M3尺寸依次增大(表1)。
Table 1
Table 1Comparison of measurements among Bounomys (mm)
| B. ulantatalensis | B. ulantatalensis (Wang, 1997) | B. bohlini (Wang, 1997) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| V 26125 | V 26126 | V 26127 | V 26128 | Average | ||||||
| P4-M3 | L | 8.54 | 8.54 | 8.12 | 8.18 | 8.35 | / | 6.15-7.30 | ||
| P4 | L | 1.4 | 1.29 | 1.17 | 1.38 | 1.31 | 1.84 | 0.98-1.48 | ||
| W | 1.88 | 2.07 | 1.71 | 1.90 | 1.89 | 2.16 | 1.23-1.97 | |||
| M1 | L | 2.09 | 2.21 | 2.12 | 2.08 | 2.13 | 2.24-2.56 | 1.31-1.99 | ||
| W | 1.96 | 2.23 | 2.01 | 2.08 | 2.07 | 2.16-2.53 | 1.48-2.05 | |||
| M2 | L | 2.28 | 2.42 | 2.39 | 2.38 | 2.37 | 2.56-2.80 | 1.64-2.13 | ||
| W | 2.29 | 2.58 | 2.47 | 2.42 | 2.44 | 2.08-2.72 | 1.56-2.09 | |||
| M3 | L | 2.52 | 2.52 | 2.46 | 2.41 | 2.48 | 2.24-2.88 | 1.72-2.16 | ||
| W | 2.38 | 2.53 | 2.37 | 2.43 | 2.43 | 2.45-2.72 | 1.60-2.05 | |||
| p4-m3 | L | 9.07 | 9.24 | 8.61 | 8.55 | 8.87 | 9.12-10.08 | 7.46-7.71 | ||
| p4 | L | 1.45 | 1.49 | 1.25 | 1.26 | 1.36 | 1.60-1.76 | 1.48-1.56 | ||
| W | 1.32 | 1.41 | 1.21 | 1.25 | 1.30 | 1.28-1.65 | 1.31-1.31 | |||
| m1 | L | 2.08 | 2.2 | 2.27 | 2.21 | 2.19 | 2.32-2.88 | 1.89-2.01 | ||
| W | 1.53 | 1.79 | 1.82 | 1.77 | 1.73 | 1.76-2.40 | 1.44-1.56 | |||
| m2 | L | 2.52 | 2.58 | 2.45 | 2.55 | 2.53 | 2.56-3.36 | 2.09-2.46 | ||
| W | 1.95 | 2.22 | 2.12 | 2.13 | 2.11 | 2.08-2.61 | 1.64-1.97 | |||
| m3 | L | 2.9 | 2.9 | 2.77 | 2.78 | 2.84 | 2.72-3.52 | 2.13-2.38 | ||
| W | 1.87 | 2.14 | 2.08 | 2.15 | 2.06 | 2.05-2.61 | 1.56-1.80 | |||
新窗口打开|下载CSV
P4原尖锥形,前尖较原尖高;原脊和后脊完整,二者都与原尖相连,后脊比原脊低,形成封闭的三角凹;前齿带较低、较短,在冠面近1/2处与原脊相连,缺乏舌侧部分;后齿带比前齿带稍显著,与后脊中部相连,具有较短的舌侧支和较长的颊侧支(图10)。
Fig. 10
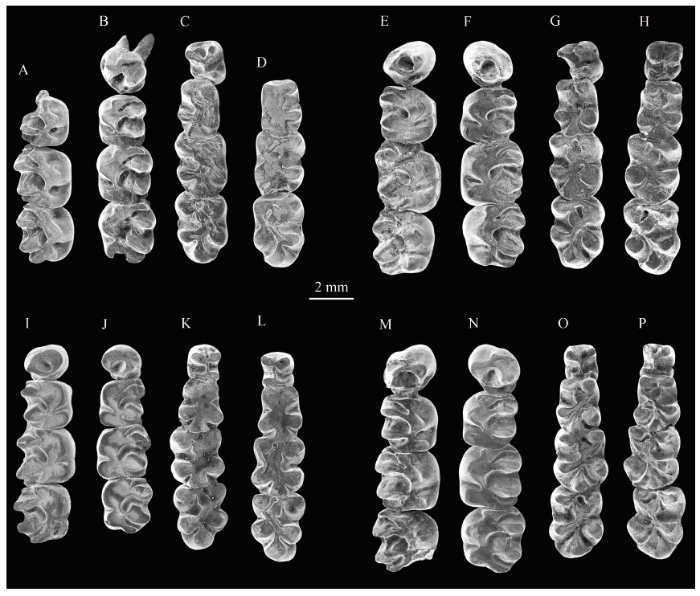 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 10Occlusal view of cheek teeth of Bounomys ulantatalensis from the Ulantatal Formation of Ulantatal area, Alxa Left Banner, Nei Mongol
A-D. IVPP V 26125: A. right M1-M3; B. left P4-M3; C. left p4-m3; D. right m1-m3;
E-H. V 26126: E. right P4-M3; F. left P4-M3; G. left p4-m3; H. right p4-m3;
I-L. V 26127: I. right P4-M3; J. left P4-M3; K. left p4-m3; L. right p4-m3;
M-P. V 26128: M. right P4-M3; N. left P4-M3; O. left p4-m3; P. right p4-m3
M1冠面方形;原尖呈V形,磨蚀后比前尖和后尖低;后尖比前尖略大;原脊和后脊都与原尖相连且二者中部均较低且窄。次尖明显,次尖与原尖之间无内脊相连;前齿带较长,长度超过牙宽的一半;前齿带舌侧端有一明显的前边尖;前沟较短;后齿带与后脊以短脊相连,后齿带与次尖之间无后沟。前凹窄长;中凹近似U形,明显比前凹和后凹开阔;后凹呈新月形,比前凹和中凹短;内凹呈斜V形,斜向后颊侧与后凹相对,中间隔以短脊。牙齿冠面可见两个明显的磨蚀面,前磨蚀面较浅、方向从前沟延伸到中凹,后磨蚀面较深、从内凹延伸到后凹;从次尖等主尖的釉质层上可以观察到方向为前舌侧-后颊侧的磨痕,与磨蚀面方向一致(图10)。
M2基本形态类似于M1,不同之处在于:个体较大;后脊较弯曲;后齿带更长;前凹呈V形,向颊侧的开口更大;后凹更为窄长(图10)。
M3大小接近M2, 与M1和M2所不同的是:冠面更接近梯形,舌侧比颊侧短;原脊较M1和M2中的更接近横向;后尖常具有前刺,并连接到前尖(3/4); 前沟和后沟较M1和M2中的更为明显;内凹更短(图10)。
下门齿(i2)横断面也呈圆角三角形,其向后延伸到m3后缘下方(图8A, B)。
同上颊齿一样,下颊齿齿列p4到m3尺寸也依次增大(表1)。
p4下原尖和下后尖后端相连(V 26125, V 26127, V 26128)或分离(V 26126); 通常下前凹呈V形,向前开口;V 26125存在下后脊Ⅰ(= mesial metalophid), 形成封闭的下三角凹;下次尖呈较低的横脊,和下内尖相连;下外脊纵向,靠近舌侧,下外脊前端只与下原尖连接(V 26126)或同时与下原尖和下后尖连接(V 26125, V 26127, V 26128) (图10)。
m1长大于宽,下三角座比下跟座窄;前齿带较弱;下原尖和下后尖都呈锥形,较膨大;下原尖后臂中段较膨大;下原尖后臂与下原尖之间具有明显的垂向沟;下三角凹封闭,较大;下外脊较短,斜向,位于臼齿近中线处;下内尖丘形,下内尖前臂向前颊侧延伸,与下原尖后臂相接;下内尖横臂较短,有时甚至缺失;V 26128磨损较轻,可以看到下内尖前臂和横臂与下外脊共同围成中央凹;下次尖呈锥形、侧扁;下次小尖呈丘形,下次小尖臂低窄且与下次尖臂相连;下次小尖颊侧具有较低的副脊,延伸到下次凹开口处;下外凹比下中凹和下后凹深、更为宽阔,且底部外缘具有低矮的齿带;下次凹比Yindirtemys中的更深更长。m1冠面可见一明显的磨蚀面,方向从下中凹延伸到下次凹;从下次尖等主尖的釉质层上可以观察到从前舌侧到后颊侧方向的磨痕(图10)。
m2形态类似于m1, 与m1不同的是:个体较大;下三角座宽度接近下跟座宽度;下原尖前后更侧扁;下三角凹较大;下次尖更接近半圆形;下后凹更深;下外凹更大;冠面可见两条磨蚀槽,前磨蚀槽较浅,后磨蚀槽较深(图10)。
m3与m1和m2所不同的是:个体最大;下三角座宽度大于或接近下跟座宽度;下原尖更侧扁,其前缘较凹;下后脊Ⅰ的中部向前凸;下次小尖颊侧无副脊;下次凹更陡(图10)。
4 对比与讨论
4.1 颊齿对比及讨论
梳趾鼠科是Gervais (1853)基于现生属Ctenodactylus建立的,亚洲陆续发现的化石种如Karakoromys, Tataromys, Leptotataromys, Yindirtemys和Sayimys等陆续被归入该科中。Wang (1997, 2001)对梳趾鼠类进行了系统分析,将梳趾鼠科(Ctenodactylidae)分为四个亚科:Tataromyinae, Karakoromyinae, Distylomyinae和Ctenodactylinae。乌兰塔塔尔新发现的化石材料为低冠丘形齿,明显不同于Distylomyinae的双叶型高冠齿及Ctenodactylinae的双叶或三叶型的颊齿类型。新材料也不同于Karakoromyinae, 区别主要在于新材料的P4后脊完整,上臼齿无内脊,后脊完整且和原尖相连,有前边尖;下臼齿下原尖后臂膨大,有中央凹,下次小尖膨大,下次凹较长、较平缓。而新材料与Tataromyinae则共享较多特征,比如:颊齿低冠,上臼齿具四条横脊,后脊完整并连接原尖,原尖呈V形,M1和M2缺失内脊,内凹较深并斜向后颊侧与后凹相对;下臼齿下原尖后臂完整,下三角凹通常封闭。因此,新材料明显不属于Karakoromyinae, Distylomyinae或Ctenodactylinae, 而应该是Tataromyinae的成员。
Wang (1997)所定义的Tataromyinae包含Tataromys, Yindirtemys, Bounomys三个属。Vianey-Liaud et al. (2006)和Schmidt-Kittler et al. (2007)分别建立了Alashania和Huangomys, 并将之归入Tataromyinae。最近,王伴月(2019)认为Alashania是Tataromys的晚出异名。
黄学诗(1985)依据乌兰塔塔尔的材料建立了两个Tataromys的种:T. ulantatalensis和T. bohlini。后来Wang (1997)将这两个种从Tataromys分离出来,转置于她建立的Bounomys中。不过,Vianey-Liaud et al. (2006)认为Wang (1997)建立Bounomys所选择的特征都是原始特征(如:颊齿低冠、齿尖膨大、齿脊较弱和腭骨较宽), 并且B. bohlini和B. ulantatalensis与Yindirtemys共享一些衍征(下中尖(即下原尖后臂)与其他的尖为新月形), 因此主张废弃Bounomys, 将所涉及两个种归入了Yindirtemys。 Schmidt-Kittler et al. (2007)也采纳了Vianey-Liaud的观点。王伴月(2019)认为Bounomys和Yindirtemys的演化方向完全不同,强调Bounomys仍然是一个有效属。
通过观察Bounomys和Yindirtemys的标本,发现二者之间既有相似性又存在明显区别。通过与Cocomys, Exmus, Protataromys这些较原始的梳趾鼠类和Tataromys, Yindirtemys, Sayimys中一些较进步的类群对比,Bounomys颊齿低冠、齿脊较弱、下内尖呈丘形以及下次小尖臂弱而低等特征确实显得较为原始。Vianey-Liaud et al. (2006)认为Bounomys的腭部较宽,并将该特征视为原始特征,但与以上3个原始类群相比,Bounomys的腭部实际上较窄。另外,Bounomys还具有一些不同于Yindirtemys的特征:Bounomys的下内尖通常具有两个弱臂,且和下外脊围成中央凹。被Vianey-Liaud et al. (2006)视为Bounomys和Yindirtemys共有衍征之一的新月形下次小尖,经再次对比后认为二者并非完全相同,前者的下次小尖更加圆胖、脊型化较弱、磨蚀面呈圆形或半月形,而在Yindirtemys中该尖则更加狭长、更加脊型化、磨蚀面呈狭长的新月形。因此,仍将Bounomys视为一个有效属。由此,根据新的厘定,Tataromyinae现在包含有Tataromys, Yindirtemys, Bounomys和Huangomys这4个属。
新材料不同于Tataromys之处在于,颊齿丘形;主尖膨大,脊较细弱;下原尖后臂中部强烈膨大呈新月形;下三角凹较大且封闭;下外脊位于臼齿近中线处。不同于Yindirtemys之处在于腭部相对较宽;颊齿主尖呈丘形,齿脊弱而低;上臼齿后尖无反前刺;p4下外脊较长;下臼齿下内尖膨大呈锥形,通常具2个弱臂。
Huangomys臼齿长/宽比较大,P4宽度超过M1宽度,颊齿主尖不膨大,齿脊较发育,下臼齿下原尖后臂不膨大,无下三角凹,下外脊更靠近舌侧,下次小尖与下内尖连接。这些特征与新标本完全不同。
Bounomys包含有两个种:B. bohlini和B. ulantatalensis。二者最明显的不同是颊齿大小的差异,B. ulantatalensis中M1的长度大概是B. bohlini中的1.5倍;另外比较显著的差异是,B. bohlini上臼齿的前齿带(前边脊)游离,中凹横向且平直,内凹和后凹常相通,下臼齿下内尖前臂较弱;而B. ulantatalensis上臼齿的前齿带不游离且与原尖相连,中凹弯向后颊侧,内凹和后凹分开,下臼齿下内尖前臂发达,和横向的后臂封闭成大的中央凹(盆); 其他细微差别如下:与B. bohlini相比,B. ulantatalensis颊齿主尖相对更膨大、脊相对较高;下臼齿中下内尖比下次尖大。通过对比,新材料颊齿特征更接近B. ulantatalensis, 此外,其大小也落在B. ulantatalensis的变异范围内(表1)。综上考虑,新材料应当归属于Bounomys ulantatalensis。
4.2 梳趾鼠类头骨对比及讨论
与始新世梳趾鼠类Cocomys和Exmus相比,Bounomys的特别之处主要在:1) 鼻骨前缘向前未超过上门齿最前面;2) 眶后突位置较靠后;3) 眶后缩不显著;4) 听泡、岩骨乳突和鼓上隐窝都很膨大;5) 存在骨质外耳道;6) 蜗管匝数较多,近3圈;7) 锤骨头向前膨大;8) 门齿孔、眶下孔和蝶腭孔都较大;9) 后鼻孔位于M3之后,较靠后;10) 下颌骨冠状突较弱等(见附表1-2)。除了在此报道的保存完好的Bounomys外,渐新世梳趾鼠类中只有Tataromys和Yindirtemys的头骨有过报道(Wang, 1997), 其中Yindirtemys保存较差,只残留了少量面颅部分。就目前已知的材料来看,这三个属头骨的基本形态非常相似,只有个别结构存在小的差异。
Bounomys与Tataromys相比,最主要的区别是:Bounomys腭孔没有Tataromys那样狭长;蝶腭孔稍靠前;额骨前筛孔较小,位置稍低,位于眶部正中(Tataromys中位于眶部中上); 蝶额孔呈三角形(Tataromys中呈半圆形); 颊肌神经孔与咬肌神经孔相邻(在Tataromys中则相距较远)。
Bounomys与Yindirtemys相区别的特征是:Bounomys眶突侧面无明显纵沟;蝶腭孔稍靠前;腭部较宽,腭桥前宽/上颊齿列长的比值范围主要在0.28-0.36 (V 26126因挤压变形其比值偏差较大,未包含在内), 而Yindirtemys腭部较窄,腭桥前宽与上颊齿列长的比值仅为0.11 (见附表2); Bounomys大腭孔没有Yindirtemys中那么狭长;下颌骨腹侧咬肌嵴前段没有Yindirtemys中发育。
由此,可以看到从始新世到渐新世梳趾鼠类头骨发生了较为明显的变化。整体来看,面颅缩窄变高,尤其是吻部根部明显缩窄,颅顶变平,颧弓向两侧拓宽。局部来看,鼻骨前缘后退;前颌骨背后突变细长,并具明显的侧背嵴;门齿孔变大;眶下孔明显扩大;上颌颧突背支后移;蝶腭孔变大;腭部变窄;后鼻孔后移;泪孔由泪骨边缘移入泪骨内;眶后突后移;眶后缩变得不明显;颞嵴由额骨移动到顶骨;顶骨和间顶骨缩小;头骨顶部颞孔消失;视神经孔和蝶眶裂融合为一个更大的孔;副卵圆孔从卵圆孔外部移到内部;鼓上隐窝和岩骨乳突膨大;蜗管匝数增加;听泡显著增大;出现了骨质外耳道;枕外嵴进一步发育;锤骨头向前膨大;锤骨前突缺失;上枕骨向头骨顶部拓展;下颌骨颏孔减少为1个,冠状突减弱,背侧咬肌脊减弱,腹侧咬肌脊向前延伸(特征及变化详见附表1-2)。
已知的中新世梳趾鼠类代表Sayimys的头骨保存的部分并不多(Bohlin, 1946), 通过现有的材料对比可以看到从渐新世到中新世梳趾鼠类头骨也有一定的变化,如:腭部水平面与脑颅底平面交角变大,使得头骨整体变拱;腭骨水平突变得更加窄长;后鼻孔位置前移;基咽管变狭窄;大腭孔前移;基蝶骨中轴部分变薄;听泡稍变小;枕外脊更加发达;下颌骨颏孔前移,腹侧咬肌嵴向前延伸更远等。从中新世到现代,梳趾鼠类头骨的基本形态没有太大的变化,只是在后鼻孔位置和大腭孔的大小等细微结构上有所变化。
始新世梳趾鼠类前颌骨顶部和侧壁圆滑过渡,而渐新世梳趾鼠类前颌骨背后突上形成显著的侧背嵴。在该嵴和眶下孔及门齿槽之间的吻部侧壁上有密集的细丝状痕迹,可能是吻部肌肉附着的痕迹。对比现生梳趾鼠Ctenodactylus vali的咀嚼肌分布(Hautier, 2010), 认为该嵴后半段系深层咬肌眶下部的起点。从始新世至渐新世,梳趾鼠类头骨的眶下孔进一步扩大,深层咬肌可由此穿过;与此同时,上颌颧突背支向后移,进一步增加了吻部背外侧嵴的长度和吻部侧壁的面积,因此可供更多的深层咬肌附着。此外,下颌骨腹侧咬肌嵴向前延伸可能也与深层咬肌在吻部的拓展和加强相对应。这些变化共同表明,从始新世至渐新世,梳趾鼠类颧咬结构由始啮型转变为了豪猪型。
Cox et al. (2012)对现生啮齿类三种颧咬肌结构的有限元分析表明,豪猪型颧咬结构对于臼齿的咀嚼运动更为高效,更有利于处理粗糙的草料类食物。从始新世到渐新世,梳趾鼠类颧咬结构从始啮型转变为豪猪型,使得其咀嚼效率变得更高。与此同时,颊齿脊形化程度相对加强,也更能适应对粗糙食物的处理。梳趾鼠类头骨及颊齿上的这些变化很可能是对渐新世以来全球变冷变干、植被由密林转为较为开阔的森林-草地这一重大环境变化的适应(Meng and McKenna, 1998; Zachos et al., 2001; Kraatz and Geisler, 2010; Sun et al., 2015; Harzhauser et al., 2016)。
从始新世到渐新世,梳趾鼠类除了咀嚼器官发生了较大变化外,耳区形态也发生了显著变化,包括:听泡、鼓上隐窝和乳突的膨大,锤骨头膨大并向前延长以及锤骨前突退化缺失。膨大的听泡常见于沙鼠(Meriones), 短耳沙鼠(Desmodillus), 更格卢鼠(Dipodomys), 非洲跳鼠(Jaculus)和巨泡象鼩(Macroscelides)等适应干旱环境的现生小哺乳动物,对它们的中耳形态及其功能的研究表明,听泡的膨大和听骨链刚度的降低(如中耳肌和锤骨前突的退化缺失)可以增强其低频听力,从而更好地适应干旱环境(Webster, 1962; Webster and Webster, 1980; Mason, 2016a, b)。南美的毛丝鼠(Chinchilla)也具有较大的听泡,这类动物生存于安第斯山干旱而开阔的环境中,不掘穴,对其耳区形态的相关功能分析也认为听泡和锤骨头的膨大,以及锤骨前突和镫骨肌的退化或缺失等特征有利于增强其低频听力(Mares and Lacher, 1987; Rosowski et al., 2006; Mason, 2013)。现生梳趾鼠(Ctenodactylus)生存于干旱的撒哈拉沙漠地区,不论是生境还是耳区形态,都与毛丝鼠非常类似,可能也具有敏锐的低频听力。渐新世梳趾鼠类耳区形态与以上适应干旱环境的小哺乳动物较为相似,尤其与现生梳趾鼠和毛丝鼠最为接近。另外,从始新世到渐新世,气候环境逐渐变冷变干,植被更加开阔(Prothero and Heaton, 1996; Zachos et al., 2001; Sun et al., 2015)。因此,渐新世梳趾鼠类膨大的听泡、乳突及鼓上隐窝、膨大并延长的锤骨头以及退化缺失的锤骨前突等特征很可能也有利于增强其低频听力,从而更好地适应渐新世更加干旱开阔的环境。
5 结论
内蒙古乌兰塔塔尔早渐新世的4件梳趾鼠类标本都属于乌兰塔塔尔丘齿鼠Bounomys ulantatalensis; 这些标本保存了几近完好的头骨及下颌,通过详细地描述为该类群提供了新的、更全面的解剖信息。从始新世到渐新世,梳趾鼠类头骨发生了明显的变化,颧咬结构由始啮型转变为豪猪型,具体表现为眶下孔扩大、前颌骨侧背嵴形成、上颌颧突背支后移、下颌骨冠状突减弱以及腹侧咬肌嵴更加前伸,这些结构的变化反映了渐新世梳趾鼠类咀嚼能力的增强。此外,耳区结构也发生了显著变化:乳突和听泡以及鼓上隐窝显著增大、锤骨头膨大并向前延长、锤骨前突退化缺失,这些特征很可能也有利于增强其低频听力。
从始新世至渐新世,全球气候发生剧变,地球由“温室”进入“冰室”,植被由密林转为较为开阔的森林-草地。相关研究表明豪猪型颧咬结构处理粗糙草料类食物的能力较其他颧咬结构更强,且低频听力在干旱开阔地区的啮齿类中有着良好的适应性。因此,梳趾鼠类颧-咬结构和耳区结构的变化,可能是对这一时期全球气候干旱化的一种积极适应。
致谢
张兆群、周伟、王平、李士杰、王永富、刘艳、王李花和王健参加乌兰塔塔尔野外工作,本文研究材料的获得有赖他们辛勤的工作;张兆群在标本鉴定等工作中提出宝贵意见;王伴月、白滨、李绿洲和熊武阳与笔者进行了有益的讨论;高伟拍摄标本照片;侯叶茂对标本进行CT扫描;司红伟协助完成显微照相;在CT数据处理过程中得到了董丽萍、侯叶茂、尹鹏飞、崔心东和郭林的帮助。在此向他们表示诚挚的谢意。Supplementary material can be found at the website of Vertebrate PalAsiatica (
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 3]
DOIURLPMID

The masticatory musculature of rodents has evolved to enable both gnawing at the incisors and chewing at the molars. In particular, the masseter muscle is highly specialised, having extended anteriorly to originate from the rostrum. All living rodents have achieved this masseteric expansion in one of three ways, known as the sciuromorph, hystricomorph and myomorph conditions. Here, we used finite element analysis (FEA) to investigate the biomechanical implications of these three morphologies, in a squirrel, guinea pig and rat. In particular, we wished to determine whether each of the three morphologies is better adapted for either gnawing or chewing. Results show that squirrels are more efficient at muscle-bite force transmission during incisor gnawing than guinea pigs, and that guinea pigs are more efficient at molar chewing than squirrels. This matches the known diet of nuts and seeds that squirrels gnaw, and of grasses that guinea pigs grind down with their molars. Surprisingly, results also indicate that rats are more efficient as well as more versatile feeders than both the squirrel and guinea pig. There seems to be no compromise in biting efficiency to accommodate the wider range of foodstuffs and the more general feeding behaviour adopted by rats. Our results show that the morphology of the skull and masticatory muscles have allowed squirrels to specialise as gnawers and guinea pigs as chewers, but that rats are high-performance generalists, which helps explain their overwhelming success as a group.
URLPMID [本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 9]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URLPMID [本文引用: 1]
DOIURLPMID [本文引用: 1]
Many species of small desert mammals are known to have expanded auditory bullae. The ears of gerbils and heteromyids have been well described, but much less is known about the middle ear anatomy of other desert mammals. In this study, the middle ears of three gerbils (Meriones, Desmodillus and Gerbillurus), two jerboas (Jaculus) and two sengis (elephant-shrews: Macroscelides and Elephantulus) were examined and compared, using micro-computed tomography and light microscopy. Middle ear cavity expansion has occurred in members of all three groups, apparently in association with an essentially 'freely mobile' ossicular morphology and the development of bony tubes for the middle ear arteries. Cavity expansion can occur in different ways, resulting in different subcavity patterns even between different species of gerbils. Having enlarged middle ear cavities aids low-frequency audition, and several adaptive advantages of low-frequency hearing to small desert mammals have been proposed. However, while Macroscelides was found here to have middle ear cavities so large that together they exceed brain volume, the bullae of Elephantulus are considerably smaller. Why middle ear cavities are enlarged in some desert species but not others remains unclear, but it may relate to microhabitat.
DOIURLPMID [本文引用: 1]
Anatomists and zoologists who study middle ear morphology are often interested to know what the structure of an ear can reveal about the auditory acuity and hearing range of the animal in question. This paper represents an introduction to middle ear function targetted towards biological scientists with little experience in the field of auditory acoustics. Simple models of impedance matching are first described, based on the familiar concepts of the area and lever ratios of the middle ear. However, using the Mongolian gerbil Meriones unguiculatus as a test case, it is shown that the predictions made by such 'ideal transformer' models are generally not consistent with measurements derived from recent experimental studies. Electrical analogue models represent a better way to understand some of the complex, frequency-dependent responses of the middle ear: these have been used to model the effects of middle ear subcavities, and the possible function of the auditory ossicles as a transmission line. The concepts behind such models are explained here, again aimed at those with little background knowledge. Functional inferences based on middle ear anatomy are more likely to be valid at low frequencies. Acoustic impedance at low frequencies is dominated by compliance; expanded middle ear cavities, found in small desert mammals including gerbils, jerboas and the sengi Macroscelides, are expected to improve low-frequency sound transmission, as long as the ossicular system is not too stiff.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
URLPMID [本文引用: 2]
[本文引用: 1]
[本文引用: 8]
DOIURL [本文引用: 2]
[本文引用: 2]
[本文引用: 9]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
DOIURLPMID [本文引用: 2]
Since 65 million years ago (Ma), Earth's climate has undergone a significant and complex evolution, the finer details of which are now coming to light through investigations of deep-sea sediment cores. This evolution includes gradual trends of warming and cooling driven by tectonic processes on time scales of 10(5) to 10(7) years, rhythmic or periodic cycles driven by orbital processes with 10(4)- to 10(6)-year cyclicity, and rare rapid aberrant shifts and extreme climate transients with durations of 10(3) to 10(5) years. Here, recent progress in defining the evolution of global climate over the Cenozoic Era is reviewed. We focus primarily on the periodic and anomalous components of variability over the early portion of this era, as constrained by the latest generation of deep-sea isotope records. We also consider how this improved perspective has led to the recognition of previously unforeseen mechanisms for altering climate.
DOIURL [本文引用: 3]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}