 ,1,2,3,*
,1,2,3,*Metatarsal II osteohistology of Xixianykus zhangi (Theropoda: Alvarezsauria) and its implications for the development of the arctometatarsalian pes
QIN Zi-Chuan1,2,3, ZHAO Qi1,2, XU Xing,1,2,3,*通讯作者: *xu.xing@ivpp.ac.cn
收稿日期:2019-03-26网络出版日期:2019-07-20
| 基金资助: |
Corresponding authors: *xu.xing@ivpp.ac.cn
Received:2019-03-26Online:2019-07-20

摘要
阿尔瓦雷斯龙类的一晚期演化支——张氏西峡爪龙(Xixianykus zhangi), 是体型最小的非鸟兽脚类恐龙之一。与其他阿尔瓦雷斯龙类类似,其胫跗骨和足部相对较长,并具有善于奔跑的兽脚类恐龙中常见的窄足型足部,指示其可能非常善于奔跑。对张氏西峡爪龙的第二跖骨进行了骨组织学研究,发现了两种罕见的骨组织学结构:放射状骨脉管和沙比纤维。认为这两种结构与其具有的窄足型足部有关,然而还需要对更多的兽脚类跖骨进行骨组织学研究来验证这一结论。
关键词:
Abstract
The late-branching alvarezsaurian Xixianykus zhangi is among the smallest known non-avialan theropods. With great similarity to its close relatives, it is highly cursorial as indicated by proportionally long lower segments of the hindlimbs and the presence of an arctometatarsalian pes-a highly modified structure that has been suggested to improve cursorial capability in theropods. Here we describe the osteohistology of the metatarsal II of the holotype of X. zhangi (XMDFEC V 0011). Two rarely reported histological features, radial vascular canals and Sharpey’s fibers, are presented in this study. We suggest that both features are related to the development of the arctometatarsalian pes; however, further investigations of metatarsal osteohistology in theropods are required for the validation of our interpretation.
Keywords:
PDF (3004KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
秦子川, 赵祺, 徐星. 张氏西峡爪龙(兽脚类:阿尔瓦雷斯龙类)第二跖骨的骨组织学及其对“窄足型”足部结构发育的启示. 古脊椎动物学报[J], 2019, 57(3): 205-213 DOI:10.19615/j.cnki.1000-3118.190425
QIN Zi-Chuan, ZHAO Qi, XU Xing.
1 Introduction
Alvarezsauria is an early-branching subgroup of maniraptoran theropods with a nearly global geographical distribution (Bonaparte, 1991; Perle et al., 1993; Karhu and Rautian, 1996; Novas, 1997; Chiappe et al., 1998; Hutchinson and Chiappe, 1998; Naish and Dyke, 2004; Martinelli and Vera, 2007; Alifanov and Barsbold, 2009; Longrich and Currie, 2009; Xu et al., 2010, 2011; Agnolin et al., 2012; Averianov and Sues, 2017). The late-branching members of this group are highly cursorial animals, as indicated by two features-extremely long lower segments of the hindlimbs and arctometatarsalian pes. The latter is characterized by a laterally compressed metatarsal III that contributes to a highly compacted metatarsus and has been suggested to improve cursorial capabilities (Holtz, 1995; Snively et al., 2004). Xixianykus zhangi is a late-branching alvarezsaurian theropod from the Upper Cretaceous Majiacun Formation of Xixia, Henan Province, China (Xu et al., 2010) and similar to other late-branching alvarezsaurians, it has an extremely modified arctometatarsalian pes that is featured by a significantly short metatarsal III producing an even more compacted metatarsus (Xu et al., 2010). Here, we study the pedal osteohistology of the holotype of X. zhangi (XMDFEC V 0011) to investigate the development of the highly modified pes.2 Material and method
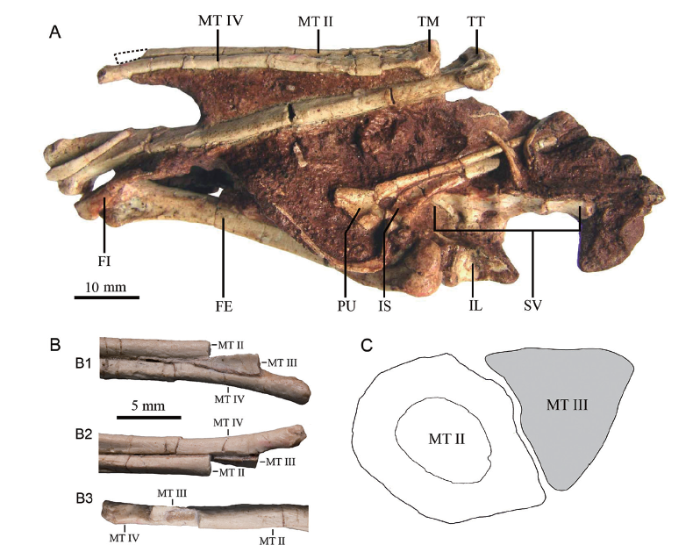
We sampled the right metatarsal II of the holotype of Xixianykus zhangi, XMDFEC V 0011 (Xixia Museum of Dinosaur Fossil Eggs of China). In this study, two ground sections were made based on the distal end of the right metatarsal II (Fig. 1).Fig. 1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1The holotype of Xixianykus zhangi (XMDFEC V 0011)and the sketch diagram for where the ground sections sliced
A. in ventral view (a dotted line indicates the sampled position); B. the dorsal (B1), ventral (B2) and medial (B3) views of its distal portion of right tarsometatarsus; C. the sketch diagram showing the spatial relation between the metatarsals II and III on the thin section (the metatarsal II colored in white is where we histologically sectioned). C is not scaled
Abbreviations: FE. femur; FI. flange; IL. ilium; IS. ischium; MT II. metatarsal II; MT III. metatarsal III;MT IV. metatarsal IV; PU. pubic; SV. sacral vertebrae; TM. tarsometatarsus; TT. tibiotarsus
Histological sections used in this study were prepared using standard techniques (Padian and Lamm, 2013). The distal end of the preserved portion of the right metatarsal II was embedded in resin, and the diaphyseal transverse sections were made using a diamond circular saw fitted with a diamond-tipped wafering blade. One surface of each ground section was polished with a wheel grinder/polisher (EXAKT400CS) by rough emery paper (500 grit), and then grounded using emery paper with finer grit (4000 grit) to produce a smooth texture ideal for the adhesion to a glass slide. The ground sections were then cut to a thickness of about 250 μm with a diamond circular saw before being ground down further to the desired final thickness of 80 μm (ground section I) and 50 μm (ground section II), leaving the exposed surface of the section smooth. The ground section I is more distally located than the ground section II, the distance between them is about 1-2 mm. Each slide was then cleaned in a water-filled ultrasonic cleaner to remove microscopic grit and finally capped with a glass coverslip. The completed two thin sections were studied in normal and polarized light, observed with Zeiss Axio Imager A2 microscope, and photographed with ZEN software from Zeiss.
3 Histological description
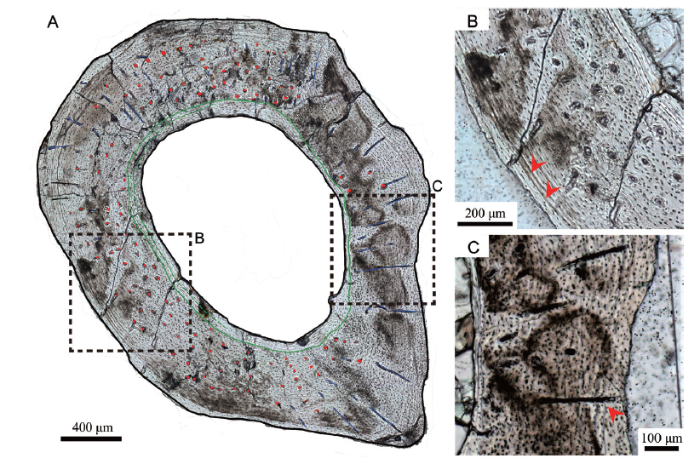
Both ground sections display compact primary cortices with sparse primary vascular canals (Figs. 2A, 3A). The medullary cavity is elliptical and filled with crystals (Fig. 3A). Two lines of arrested growth (LAGs) are identified (Fig. 2) and have been interpreted as a one-year growth record (Chinsamy, 2005). Though fibrolamellar bone occupies most of the cross section, the endosteal bone forms an uninterrupted band of lamellar bone laying alongside what used to be the endosteum. Longitudinal primary vascular canals and osteocyte lacunae dominate the bone cortex (Fig. 2B).Fig. 2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Microphotograph of the ground section I of metatarsal II of Xixianykus zhangi (XMDFEC V 0011) under normal light
A. whole cross section under the normal light (red dots, primary canals; blue lines, radial canals; green line, endosteal bone boundary); B. the medial portion of the slice is dominated by longitudinal canals (two LAGs are indicated by two red arrows); C. a radial canal lies across the two LAGs (indicated by a red arrow)
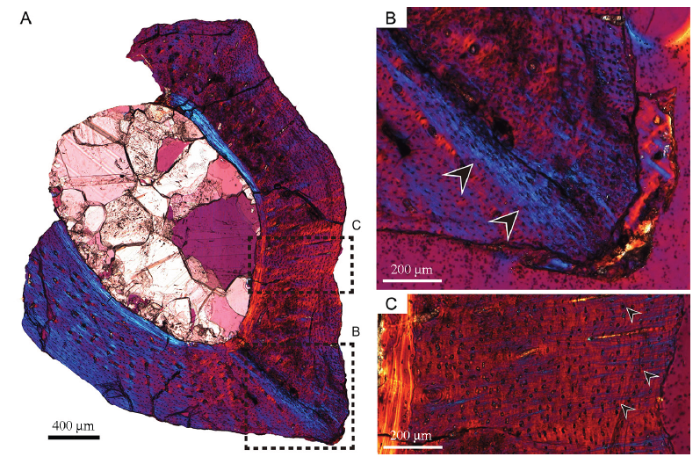
Fig. 3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Microphotograph of the ground section II of metatarsal II of Xixianykus zhangi (XMDFEC V 0011) under polarized light
A. whole cross section under polarized light;B. Sharpey’s fibers can be seen in the lateroventral corner of metatarsal II (indicated by the black arrows);C. sparsely distributed Sharpey’s fibers on the lateral side of metatarsal II (indicated by the small black arrows)
Ground section I, which has a thickness of 80 μm, displays radially arranged vascular canals in most of the cortex except for the ventral half of the lateral portion, and for the endosteal bone (Fig. 2A). These radial canals cross the growth marks without any morphological changes, indicating a multiyear forming structure (Fig. 2C). These radial canals are about 100-300 μm apart (Fig. 2C). Most of those in the lateral portion of the ground section are approximately 200 μm long, and those in the other portions of the ground section are variable in length and in general much shorter. Most radial canals are oriented perpendicularly to sub-perpendicularly to the periosteum. They reach close to the periosteum on the lateral and medial sides but are much deeper within the cortex and far from the periosteum on the dorsal side.
In ground section II, which has a thickness of 50 μm, Sharpey’s fibers can be seen on the lateral side when observed under polarized light (Francillon et al., 1990; Holger and Sander, 2013). Under polarized light equipped with a quartz interferometer plate, Sharpey’s fibers emerge as a blue-colored filamentous structure (Fig. 3A). Since the ground section I is thicker, Sharpey’s fibers are hardly observed.
Near the lateroventral portion, what appears to be a bundle of radial Sharpey’s fibers can be seen. This bundle is obliquely oriented to the ventral periosteal surface (with an angle of approximately 40°, Fig. 3B). More dorsally in the lateral portion of the ground section are more sparsely distributed, much shorter Sharpey’s fibers. These fibers are parallel to each other and to nearby radial canals, and they are nearly perpendicular to the periosteal surface (Fig. 3C).
4 Discussion
The main histological features observed in the metatarsal II of Xixianykus zhangi, including proportionally large medullary cavity, fibrolamellar-bone-dominated cortex, are similar to those in the long bones of other small-sized theropods, such as the Shuvuuia and Archaeopteryx (Erickson et al., 2001, 2009). For example, the medullary cavity is proportionally large, the cortex is mainly composed of fibrolamellar bone, longitudinal vascular canals dominate the cortex and are relatively sparse in distribution (Erickson et al., 2001, 2009).Two histological features, the radial vascular canals and the Sharpey’s fibers, are particularly interesting. Sharpey’s fibers were first reported in alveolar tooth sockets of mammals as part of the periodontal ligament that can be traced through dental tissues (Sharpey, 1848). Sharpey’s fibers also referred to connective tissue fibers that continuous with skeletal tissues and their enthesis. Sharpey’s fibers also function as the binding of cranial bones and attaching cranial muscles to bone periosteum (Pritchard et al., 1956; Jones and Boyde,1974; Nicaise et al., 1988; Hieronymus, 2006). Sharpey’s fibers have been reported in extinct taxa, and have been suggested that they functioned as same as in extant animals (Chinsamy, 2005; Francillon et al., 1990; Holger and Sander, 2013). Among Mesozoic theropods, Sharpey’s fibers have been reported in the tibia of the ceratosaurian Masiakasaurus knopfleri (Lee and O’Connor, 2013), the femur of an unnamed ceratosaurian (Canale, 2016), the hindlimb of the early-branching avialan Confuciusornis (Ricqlès et al., 2003), and the quill knobs of a few early-branching avialans (Wang, 2012). In the first three examples, Sharpey’s fibers were probably used for muscle attachment whereas in the last case, they have been suggested to represent the insertion sites of feathers to bone (Wang, 2012). Sharpey’s fibers were also suggested to be present in the metatarsals of some tyrannosaurids and in Allosaurus based on intermetatarsal rugose facets (Snively and Russell, 2003), though in the absence of any histological evidence.
Our observation represents the first histological evidence for the presence of Sharpey’s fibers in the pes of any Mesozoic theropod. Sharpey’s fibers in XMDFEC V 0011 include two different types: 1) obliquely oriented, densely-bunched ones in the lateroventral corner of the cross section of metatarsal II; and 2) parallelly arranged and sparsely distributed ones in the lateral portion of the cross section of metatarsal II. In living mammals, the Sharpey’s fibers found at the junction of cranial bones are sparser than those found at muscle attachment (Jones and Boyde, 1974). The densely-packed Sharpey’s fibers seen in the lateroventral corner of the cross section of metatarsal II of XMDFEC V 0011 were more likely to be used for a tendinous attachment, whereas the parallelly arranged and sparsely distributed ones were most likely part of a ligamentous sheath surrounding and binding the metatarsal bones together, as in living mammals where Sharpey’s fibers are used to bind cranial bones in a firm yet slightly movable manner (Pritchard et al., 1956).
In living birds, radial canals represent extremely rapid growth (Margerie et al., 2004; Montes et al., 2010), but they are also known in two other conditions such as in pathological bones and in bones associated with substantial shape changes (Erickson and Tumanova, 2000). Radial vascular canals were not reported in other small theropods such as Mahakala or Archaeopteryx (Erickson et al., 2009), nor other alvarezsaurians including Shuvuuia deserti, Albinykus Baatar, Xiyunykus pengi and Bannykus wulatensis (Erickson et al., 2001; Nesbitt et al., 2011; Xu et al., 2018). They are also unknown in giant theropods (Erickson et al., 2004, Horner and Padian, 2004) and were not found even in sauropods (Curry, 1999; Klein and Sander, 2008), presumably because these gigantic dinosaurs had lower maximum growth rates and bone deposition rates than those of living-birds (Margerie et al., 2004; Montes et al., 2010; Erickson, 2014).
However, radial vascular canals are known to be present in the ceratopsians Psittacosaurus mongoliensis (Erickson and Tumanova, 2000) and P. lujiatunensis (Zhao et al., 2013), and the hadrosaurian Maiasaura (Cubo et al., 2015). In these cases, the radial vascular canals were suggested to be related to substantial shape changes as a result of loading change (Erickson and Tumanova, 2000; Zhao et al., 2013; Cubo et al., 2015). Radial vascular canals were also reported in the early-branching sauropodomorpha Plateosaurus engelhardti but the interpretation of these structures was ambiguous (Klein, 2004).
Our report of the presence of radial vascular canals in XMDFEC V 0011 is the first one in any theropod. They differ from the radial vascular canals that were identified as resulting from a pathology, like those found in a giant dinosaur (identified as a possible ornithopod or sauropod) in the following criteria (Chinsamy and Deratzian, 2009): radial canals are 100-300 μm apart from each other rather than densely bunched; radial canals are oriented perpendicularly to the periosteum rather than parallelly; radial canals extend from the endosteum to the periosteum instead of limited to near the periosteum. Radial vascular canals in XMDFEC V 0011 are probably related to substantial shape changes, as suggested for other dinosaurs (Erickson and Tumanova, 2000; Zhao et al., 2013; Cubo et al., 2015). More specifically, the lateroventral portion of metatarsal II expands significantly laterally to buttress the ventrally thin metatarsal III to form the arctometatarsal condition.
The presence of Sharpey’s fibers and radial vascular canals in metatarsal II of Xixianykus shed new light on how the arctometatarsal condition forms. The former feature explains how the metatarsals connects in a firm but movable manner, which is important for cursorial capability; the latter feature explains the substantial shape change, seen at the histological level, that occurs during the development of the arctometatarsal condition. However, more histological investigations of theropod metatarsals, especially in the arctometatarsalian pes, are needed to confirm this interpretation.
Acknowledgements
We thank Ding Xiaoqing for preparing specimens, Zhang Shukang for guidance of manufacturing histological slices, and Chen Guanhan for photographs. We thank P. Martin Sander from University of Bonn for the discussion. We thank Alida Bailleul for the discussion and her comments on the manuscript. This study was supported by the National Natural Science Foundation of China (41688103 and 41120124002).参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 3]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 4]
PMID [本文引用: 3]

Did dinosaurs grow in a manner similar to extant reptiles, mammals or birds, or were they unique? Are rapid avian growth rates an innovation unique to birds, or were they inherited from dinosaurian precursors? We quantified growth rates for a group of dinosaurs spanning the phylogenetic and size diversity for the clade and used regression analysis to characterize the results. Here we show that dinosaurs exhibited sigmoidal growth curves similar to those of other vertebrates, but had unique growth rates with respect to body mass. All dinosaurs grew at accelerated rates relative to the primitive condition seen in extant reptiles. Small dinosaurs grew at moderately rapid rates, similar to those of marsupials, but large species attained rates comparable to those of eutherian mammals and precocial birds. Growth in giant sauropods was similar to that of whales of comparable size. Non-avian dinosaurs did not attain rates like those of altricial birds. Avian growth rates were attained in a stepwise fashion after birds diverged from theropod ancestors in the Jurassic period.
PMID [本文引用: 1]
How evolutionary changes in body size are brought about by variance in developmental timing and/or growth rates (also known as heterochrony) is a topic of considerable interest in evolutionary biology. In particular, extreme size change leading to gigantism occurred within the dinosaurs on multiple occasions. Whether this change was brought about by accelerated growth, delayed maturity or a combination of both processes is unknown. A better understanding of relationships between non-avian dinosaur groups and the newfound capacity to reconstruct their growth curves make it possible to address these questions quantitatively. Here we study growth patterns within the Tyrannosauridae, the best known group of large carnivorous dinosaurs, and determine the developmental means by which Tyrannosaurus rex, weighing 5,000 kg and more, grew to be one of the most enormous terrestrial carnivorous animals ever. T. rex had a maximal growth rate of 2.1 kg d(-1), reached skeletal maturity in two decades and lived for up to 28 years. T. rex's great stature was primarily attained by accelerating growth rates beyond that of its closest relatives.
DOIURL [本文引用: 3]
[本文引用: 2]
PMID [本文引用: 1]
Muscular reconstructions in vertebrate paleontology have often relied heavily on the presence of "muscle scars" and similar osteological correlates of muscle attachment, a practice complicated by the fact that approximately half of tendinous muscle attachments to bone in extant vertebrates do not leave readily interpretable scars. Microanatomical and histological correlates of tendinous muscle attachment are much less ambiguous. This study examines the microanatomical correlates of muscle attachment for the mandibular adductors in six species of diapsids. Most prominent tendinous or aponeurotic muscle attachments display a high density of extrinsic fibers (similar to Sharpey's fibers). There is also some indication that the density of extrinsic fibers at an attachment may be directly related to the amount of stress exerted on that attachment. The presence of comparable densities of extrinsic fibers in fossil tissue constitutes strong and readily interpretable positive evidence for the presence of adjacent fibrous connective tissue in life. Microanatomy and histology provide reliable data about muscle attachments that cannot be gleaned from gross observation alone. These additional data, when coupled with existing muscular reconstruction techniques, may be essential to the resolution of ambiguous character states, and will provide more severe tests for long-standing hypotheses of musculature in extinct diapsids. Increasing the accuracy and precision of muscular reconstructions lends greater strength to any phylogenetic, paleobiological, or paleoecological inferences that draw upon these reconstructions as important lines of evidence.Copyright 2006 Wiley-Liss, Inc.
DOIURL [本文引用: 2]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
PMID [本文引用: 2]
Microstructure-function relationships remain poorly understood in primary bone tissues. The relationship between bone growth rate and bone tissue type, although documented in some species by previous works, remains somewhat unclear and controversial. We assessed this relationship in a species with extreme adaptations, the king penguin (Aptenodytes patagonicus). These birds have a peculiar growth, interrupted 3 months after hatching by the austral winter. Before this interruption, chicks undergo extremely rapid statural and ponderal growth. We recorded experimentally (by means of fluorescent labelling) the growth rate of bone tissue in four long bones (humerus, radius, femur and tibiotarsus) of four king penguin chicks during their fastest phase of growth (3-5 weeks after hatching) and identified the associated bone tissue types ('laminar', 'longitudinal', 'reticular' or 'radial' fibro-lamellar bone tissue). We found the highest bone tissue growth rate known to date, up to 171 microm day(-1) (mean 55 microm day(-1)). There was a highly significant relationship between bone tissue type and growth rate (P<10(-6)). Highest rates were obtained with the radial microarchitecture of fibro-lamellar bone, where cavities in the woven network are aligned radially. This result supports the heuristic value of a relationship between growth rate and bone primary microstructure. However, we also found that growth rates of bone tissue types vary according to the long bone considered (P<10(-5)) (e.g. growth rates were 38% lower in the radius than in the other long bones), a result that puts some restriction on the applicability of absolute growth rate values (e.g. to fossil species). The biomechanical disadvantages of accelerated bone growth are discussed in relation to the locomotor behaviour of the chicks during their first month of life.
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 1]
[本文引用: 1]
PMID [本文引用: 1]
We present a hypothesis of tyrannosaurid foot function termed the "tensile keystone model," in which the triangular central metatarsal and elastic ligaments dynamically strengthened the foot. The tyrannosaurid arctometatarsus, in which the central metatarsal is proximally constricted, displays osteological correlates of distal intermetatarsal ligaments. The distal wedge-like imbrication of tyrannosaurid metatarsals indicates that rebounding ligaments drew the outer elements towards the middle digit early in the stance phase, unifying the arctometatarsus under high loadings. This suggests increased stability and resistance to dissociation and implies, but does not demonstrate, greater agility than in large theropods without an arctometatarsus.Copyright 2002 Wiley-Liss, Inc.
DOIURL [本文引用: 1]
Beijing: University of Chinese Academy of Sciences,
[本文引用: 2]
DOIURL [本文引用: 3]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIPMID [本文引用: 3]
A few dinosaurs are inferred to have undergone an ontogenetic shift from quadrupedal-to-bipedal posture, or vice versa, based on skeletal allometry. The basal ceratopsian Psittacosaurus lujiatunensis is considered to have been mainly bipedal as an adult. Here we infer a postural shift in this species based on a novel combination of limb measurements and histological data. The forelimb is strongly negatively allometric relative to the hindlimb, and patterns of vascular canal orientation provide evidence that growth of the hindlimb was particularly rapid during the middle part of ontogeny. Histology also makes it possible to determine the ontogenetic ages of individual specimens, showing that the forelimb-to-hindlimb ratio changed rapidly during the first or second year of life and thereafter decreased gradually. Occurrence of an ontogenetic shift from quadrupedality to bipedality was evidently widespread in dinosaurs, and may even represent the ancestral condition for the entire group.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}