 ,1,2, 白炜鹏1,2,3, 潘越1,2,3, 刘文晖4
,1,2, 白炜鹏1,2,3, 潘越1,2,3, 刘文晖4New material of Cervidae (Artiodactyla, Mammalia) from Xinyaozi Ravine in Shanxi, North China
DONG Wei,1,2, BAI Wei-Peng1,2,3, PAN Yue1,2,3, LIU Wen-Hui4通讯作者: dongwei@ivpp.ac.cn
收稿日期:2019-12-12网络出版日期:2020-07-20
| 基金资助: |
Corresponding authors: dongwei@ivpp.ac.cn
Received:2019-12-12Online:2020-07-20

摘要
20世纪80年代在桑干河盆地一带考察泥河湾层时,在山西省天镇县南高崖乡的台家坪、水冲口和大庄科的辛窑子沟一带发现了很多哺乳动物化石地点并出土了很多哺乳动物化石。最近对其中鹿科化石的研究鉴定出7个种:步氏麂(Muntiacus bohlini)、化德祖鹿相似种(Cervavitus cf. C. huadeensis)、山西轴鹿(Axis shansius)、华丽日本鹿(Nipponicervus elegans)、始麋鹿(Elaphurus davidianus predavidianus)、双叉麋鹿(E. bifurcatus )及赤鹿(Cervus (Elaphus) elaphus)。其中前6个种产自台家坪和水冲口化石地点的泥河湾组中,层位相当于欧洲的中、晚维拉方期,即早更新世,而赤鹿产自大庄科化石地点,层位目前还不确定,但可能也是泥河湾组。因此在辛窑子沟一带的早更新世目前发现的鹿科化石至少有6个种。其中的化德祖鹿相似种和山西轴鹿是从晚新近纪残留下来的种类,而步氏麂、华丽日本鹿、始麋鹿和双叉麋鹿是早更新世出现的新种类。如果大庄科地点的层位可以确定为早更新世的话,赤鹿的最早记录则可以回溯到早更新世,其鹿角的演化是始于无冰枝,然后出现冰枝。另外,在中国大陆和日本列岛都发现过麋鹿和日本鹿化石,进一步说明在早更新世期间海平面曾经下降很多,使得大陆的一些鹿类得以迁徙到列岛上。最后,数量较多的低冠鹿科化石标本指示桑干河盆地一带在早更新世存在一定范围的森林环境。
关键词:
Abstract
Many cervid specimens were uncovered during the field exploration for Nihewan beds at the beginning of the 1980s from Taijiaping, Shuichongkou and Dazhuangke localities along Xinyaozi Ravine at Nangaoya Township of Tianzhen County, Shanxi Province in North China. Recent studies on the cervid material identified seven species of Cervidae in total: Muntiacus bohlini, Cervavitus cf. C. huadeensis, Axis shansius, Nipponicervus elegans, Elaphurus davidianus predavidianus, E. bifurcatus from the Early Pleistocene deposits at Taijiaping and Shuichongkou localities, and Cervus (Elaphus) elaphus from uncertain horizon at Dazhuangke. At least the previous six species of cervids were from Nihewan Formation (Nihewanian or equivalent to European middle and late Villafranchian), i.e. the Early Pleistocene, in Sangganhe Basin area. Cervavitus cf. C. huadeensis and A. shansius were survivors from the Late Neogene; M. bohlini, N. elegans, E. davidianus predavidianus and E. bifurcatus are new forms of the Early Pleistocene. If Dazhuangke horizon can be dated as those of Shuichongkou and Taijiaping localities, the appearance of elaphoid cervids could be traced back to the Early Pleistocene, and the evolution of elaphoid antler would start from absence to presence of bez tine. The presence of Elaphurus and Nipponicervus in mainland China and Japanese archipelago implies further that the sea level was dropped down that these cervids could migrate from the mainland to the islands. The abundance of folivorous cervid specimens in the Xinyaozi Ravine area indicates the existence of a certain scale of forested environment in Sangganhe Basin area during the Early Pleistocene.
Keywords:
PDF (2350KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
董为, 白炜鹏, 潘越, 刘文晖. 山西天镇辛窑子早更新世鹿科化石新材料. 古脊椎动物学报[J], 2020, 58(3): 221-248 DOI:10.19615/j.cnki.1000-3118.200320
DONG Wei, BAI Wei-Peng, PAN Yue, LIU Wen-Hui.
1 Introduction
The field exploration for Nihewan beds at the beginning of the 1980s by the team of Wei Qi (1997) resulted in discovery of many cervid specimens from Taijiaping, Shuichongkou and Dazhuangke localities along Xinyaozi Ravine at Nangaoya Township of Tianzhen County, Shanxi Province in North China. The mammalian fossils collected along the ravine were called Xinyaozi fauna (Wei, 1997; Tong et al., 2011) and identified total four cervid taxa, i.e. Muntiacus bohlini, Elaphurus bifurcatus, Eucladoceros boulei and Rusa elegans (Wei, 1997). Although the localities are in Shanxi Province, they are very close to Xiashagou (=Hsia-sha-kou) localities (Teilhard de Chardin and Piveteau, 1930) in the Nihewan Basin yielding cervids such as E. bifurcatus, Cervus (Rusa) elegance etc. in Hebei Province. The fossils unearthed from the Shuichongkou deposits include Nyctereutes sp., Canis sp., Agriotherium sp., Ursus sp., Meles sp., Felis sp., Pachycrocuta sp., Postschizotherium intermedium, Plesiohipparion houfenense, Equus sanmeniensis, Axis sp. etc. (Qiu et al., 2002), as well as Spirocerus wongi (Bai et al., 2019). The faunal composition indicates an earlier age than that of the classical Nihewan fauna, likely ranging between 1.8 Ma and 2.6 Ma (Qiu et al., 2002). The fossils unearthed from Taijiaping deposits include Canis chihliensis palmidens, Nyctereutes sinensis, Pachycrocuta licenti, Homotherium sp., Megantereon nihewanensis, Lynx sp., Felis sp., Proboscidipparion sinense, Equus sanmeniensis, Coelodonta yanshanensis, ?Rhinoceros sp., Gazella sp., Elaphurus bifurcatus (Qiu, 2002), as well as E. davidianus predavidianus (Dong et al., 2019). The geologic age of the fauna can roughly be considered as contemporary with that of the classical Nihewan fauna (Qiu, 2002), with an estimated age of ca. 2.2-1.7 Ma (Liu et al., 2012), equivalent to from upper part of MNQ17 to lower part of NMQ18 (Woodburne, 2013). The deposits at Shuichongkou are therefore slightly earlier than those at Taijiaping. However, many specimens collected during the 1980s’ explorations have not yet been systematically studied. We reinvestigated the localities along Xinyaozi Ravine in 2017, studied cervid specimens from Taijiaping locality and identified two Elaphurus species (Dong et al., 2019). Here we systematically describe the rest specimens of cervids from Xinyaozi Ravine at Tianzhen and discuss on their taxonomy and evolutionary history. The antler measurement methods follow that of Dong et al. (2019), the dental terminology follows that of Dong (2004). The complete skulls with both antlers and dentitions are unfortunately absent in the collection. The taxonomic assignment of the antlers, upper and lower dentitions were based on their relative dimensions and morphological comparison with available identified specimens. The specimens described are housed at the Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences (IVPP).2 Systematic paleontology

(Figs. 1-2; Tables 1-3)
Cervulus cf. sinensis Teilhard de Chardin and Piveteau, 1930, p. 44-46
Cervulus bohlini sp. nov. Teilhard de Chardin, 1940, p. 79-88
Muntiacus bohlini Hooijer, 1951, p. 7
Muntiacus bohlini Wang and Wu, 1979, p. 541
Muntiacus bohlini Wei, 1997, p. 196
Material A nearly complete left antler with burr and broken pedicle (IVPP V 26267.1) from Shuichongkou; a right maxillary fragment with M1-3 (V 26267.2), a left mandibular fragment with p2-m2 (V 26267.3), a right mandibular fragment with p3-m3 (V 26267.4), and a right mandibular fragment with dp4-m2 (V 26267.5) from Taijiaping, Tianzhen, Shanxi.
Description The specimen V 26267.1 (Fig. 1) is a left mature but unshed antler (see Table 1 for measurements) from an adult individual. Its pedicle is broken but a small part remained just below the burr. The cross sections of the pedicle are oval and the surface of the pedicle is smooth. Although the burr is partially broken on the lateral side, it is moderately developed and composed of series of bony nodes. The brow tine is broken, and it might be very small based on the scar of the broken brow tine and close to but relatively high above the burr in munjacks (Fig. 1D). The main beam is thick, developed backward and slightly laterally at first and then curved medially at two thirds of its length. It is flattened near the base. Its cross sections are roughly semicircular or somewhat triangular near the base and become oval at the distal part. The ornamentation of the antler is formed by well-developed longitudinal grooves and crests on the surface of the antler crown.
Fig. 1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1A nearly complete antler (IVPP V 26267.1) of Muntiacus bohlini from Xinyaozi Ravine
A. lateral view; B. medial view; C. anterior-medial view; D. outline of anterior-medial view Dotted lines show missing tips and oblique lines show broken area
Table 1
Table 1Measurements of antler of Muntiacus bohlini from Xinyaozi Ravine and comparison (mm)
| Taxon | M. bohlini | M. lacustris | M. zhaotongensis | M. leilaoensis | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Locality | Shuichongkou | Xiashagou1) | Loc. 182) | Yushe3) | Juyuandong4) | Zhaotong5) | Leilao6) | ||||
| Max diameter of distal pedicle | 18.9 | 18.5 | 12-17.1 | 16-17 | 15 | 16.44-20.10 | 18.56-18.82 | ||||
| Min diameter of distal pedicle | 13.7 | 13 | 10-14.5 | 13.5 | 14.02-16.94 | 16.40-15.68 | |||||
| Thickness of burr | 4.4 | 4.9* | 4.3 | 4.56-6.32 | 5.96-7.22 | ||||||
| Max diameter of burr | 27.1 | 21.4 | 22-32 | 25? | 25.52-33.03 | 38.78-39.46 | |||||
| Min diameter of burr | 21.9 | 18.7 | 23.23-29.62 | 33.12-37.24 | |||||||
| Length of antler base | 17.5 | 14.7* | 16 | 16-31 | 7.38-18.48 | 12.24-22.32 | |||||
| Angle of bifurcation (°) | 50 | 40 | 40 | 50-65 | 35-40 | 40 | |||||
| Max diameter of proximal antler base | 21.2 | 24 | 20-26 | 17-24 | 19 | 16.54-24.36 | 23.38-30.12 | ||||
| Min diameter of proximal antler base | 18.3 | 23 | 16-23 | 14-23 | 15.6 | 11.79-16.53 | 21.22-22.16 | ||||
| Length of the main beam | >76.1 | 108 | 72-80 | 75-92 | 80 | 73.23-90.05 | 125.10 | ||||
| Max diameter of proximal main beam | 21.4 | 22 | 15-20 | 16.04-24.42 | 21.86-23.82 | ||||||
| Min diameter of proximal main beam | 12.4 | 14 | 10.5-14 | 11.68-16.72 | 14.58-14.92 | ||||||
| Max diameter of proximal brow tine | 17.2 | 9.6 | 14-26 | 7.26-9.47 | 9.74 | ||||||
| Min diameter of proximal brow tine | 11.3 | 9.5* | 8.4 | 12-18 | 4.59-5.89 | 8.68 | |||||
新窗口打开|下载CSV
The preserved right maxillary fragment (V 26267.2) is composed mainly of three molars (Fig. 2A). The M1, M2 and M3 are all composed of four main cusps; their length and width are close to each other. Accessory elements such as neocrista, entostyle and spur are developed; entocingulum is visible although undeveloped. Measurements of the upper cheek teeth are listed in Table 2.
Fig. 2
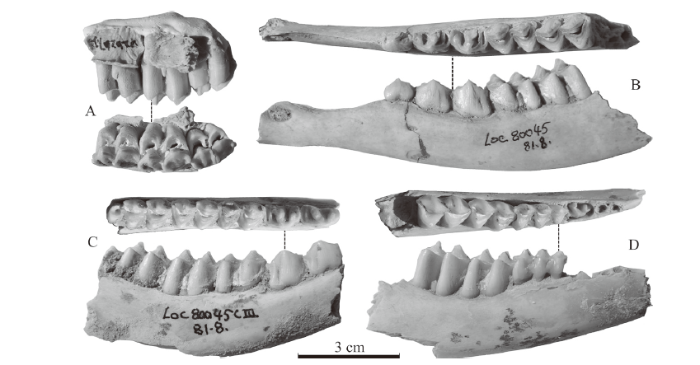 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Cheek teeth of Muntiacus bohlini from Xinyaozi Ravine
A. a right maxillary fragment with M1-3 (IVPP V 26267.2), lingual and occlusal views;
B. a left mandibular fragment with p2-m2 (V 26267.3), occlusal and buccal views;
C. a right mandibular fragment with p3-m3 (V 26267.4), occlusal and buccal views;
D. a right mandibular fragment with dp4-m2 (V 26267.5), occlusal and buccal views
Table 2
Table 2Measurements of the upper cheek teeth of Muntiacus bohlini from Xinyaozi Ravine and comparison (mm)
| M. bohlini | M. lacustris | Muntiacus sp. | M. zhaotongensis | |||||
|---|---|---|---|---|---|---|---|---|
| V 26267.2 | RV 4002 | Sanhe1) | OV 949 | Zhaotong2) | ||||
| M1 L | 14.18 | 11.22-12.16 | 10.5-10.7 | 10.26-10.86 | 9.88-10.61 | |||
| M1 W | 13.54 | 11.02-11.28 | 13.8-14.1 | 11.94-11.96 | 11.35-12.19 | |||
| M1 H | 11.09 | 5.96-8 | 4.1-4.2 | 3.58-3.68 | 7.81-8.22 | |||
| M2 L | 14.89 | 11.98-12.44 | 12.9-13.4 | 11.96-12.02 | 10.33-11.13 | |||
| M2 W | 14.88 | 12.38-13.18 | 14.6-15.3 | 13.08-13.18 | 12.19-13.62 | |||
| M2 H | 13.15 | 10.36-10.42 | 8.2-9.2 | 3.72-4.84 | 8.75-9.91 | |||
| M3 L | 14.91 | 12.64-12.76 | 11.9-13.9 | 12.54-12.64 | 9.9-10.51 | |||
| M3 W | 12.81 | 11.82-12 | 12.8-13.9 | 13.06-13.5 | 11.67-12.88 | |||
| M3 H | 13.01 | 9.32-11.64 | 7.7-10.3 | 4.34-4.88 | 8.6-8.99 | |||
| M1-3 L | 41.47 | 36.18-36.2 | 33.82-34.28 | 29.88 | ||||
新窗口打开|下载CSV
The p2 is present on V 26267.3 only and simply composed of a main cusp and some minor cusps (Fig. 2B). Its trigonid basin, entoflexid and talonid basin are present and opened lingually. The p3 is present on V 26267.4 and V 26267.5 and composed of two main cusps and some minor cusps (Fig. 2C-D). Paraflexid, trigonid basin, entoflexid and talonid basin are all present and opened lingually, and the hypoflexid is also present. The p4 is also present on V 26267.4 and V 26267.5 and similar to p3, but evidently larger, and metaconid extends mesiodistally (Fig. 2C-D). The dp4 (14.16 mm×7.45 mm×5.7 mm) is present on V 26267.5 and composed of three lobes with a buccal basal pillar between every two lobes (Fig. 2D).
The m1 and m2 are present on all three mandibular fragments and they are all composed of four selenodont main cusps, the Palaeomeryx fold is evidently absent, precingulum is present but undeveloped, while ectostylid is developed (Fig. 2B-D). The m3 is present only on V 26267.4 and composed of three lobes, the anterior two lobes resemble those of m1 and m2, but the third lobe is composed of a developed hypoconulid and a small entoconulid (Fig. 2C).
The measurements of lower cheek teeth are listed in Table 3.
Table 3
Table 3Measurements of the lower cheek teeth of Muntiacus bohlini from Xinyaozi Ravine and comparison (mm)
| M. bohlini | M. lacustris | Muntiacus sp. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| V 26267.3 | V 26267.4 | V 26267.5 | Xiashagou1) | Loc. 182) | Sanhe3) | Juyuandong4) | Longgudong5) | |||
| p2 L | 8.31 | 8.5* | 7* | 8.7 | 7.7 | |||||
| p2 W | 4.91 | 4.6 | 4.0 | |||||||
| p2 H | 6.08 | 8.2 | ||||||||
| p3 L | 9.07 | 10.09 | 9.7* | 8.5* | 9.2-9.8 | |||||
| p3 W | 6.21 | 6.44 | 5.2-5.7 | |||||||
| p3 H | 8.37 | 8.44 | ||||||||
| p4 L | 9.68 | 10.43 | 9.7* | 9.9* | 9.5 | 9.7 | 10.4 | |||
| p4 W | 7.21 | 7.32 | 5.8* | 6.9 | 6.8 | 6.0 | ||||
| p4 H | 9.68 | 9.75 | 4.3 | |||||||
| m1 L | 13.82 | 13.84 | 13.14 | 13* | 12.7* | 12.4 | 11.0 | 10.1-11.5 | ||
| m1 W | 9.02 | 9.09 | 8.05 | 8.0 | 7.1 | 6.9-7.9 | ||||
| m1 H | 8.04 | 7.98 | 10.2 | 7.1 | ||||||
| m2 L | 14.00 | 14.30 | 14.48 | 15.4* | 14.2* | 13.0-13.2 | 11.5 | 11.0-13.2 | ||
| m2 W | 9.69 | 9.61 | 9.49 | 7.9-8.7 | 7.6 | 7.7-8.2 | ||||
| m2 H | 10.01 | 10.34 | 10.77 | 9.8-11.8 | ||||||
| m3 L | 19.97 | 19.5* | 18.3* | 17.3 | 13.6-17.2 | 14.2-17.5 | ||||
| m3 W | 9.73 | 9.1 | 6.8-8.6 | 7.2-8.5 | ||||||
| m3 H | 9.23 | 9.7 | ||||||||
| p2-4 L | 27.9 | 27 | 24-26 | |||||||
| m1-3 L | 47.72 | 46* | 38-40 | |||||||
新窗口打开|下载CSV
Comparison and determination The antler (V 26267.1) is small, its missing brow tine is obviously tiny, its main beam is also small and inclines medially at distal part. It is therefore similar to that from Xiashagou previously identified as Muntiacus cf. sinensis (Teilhard de Chardin and Piveteau, 1930) and later revised as M. bohlini (Teilhard de Chardin, 1940). Its morphology falls well in the variation range of those of M. bohlini from Loc. 18 near Beijing (=Peking) (Teilhard de Chardin, 1940). It is also metrically similar to that from Xiashagou and those from Loc. 18 (Table 1). The antler (V 26267.1) is also morphologically close to those of M. lacustris from Yushe Basin (Teilhard de Chardin and Trassaert, 1937) and from Juyuandong (Han, 1987). For example, the brow tine is tiny, the main beam is small and curves medially, and the dimensions are close to each other. But its bifurcation position is evidently higher. It differs from those of M. leilaoensis (Dong et al., 2004) and M. zhaotongensis (Dong et al., 2014) by its smaller burr and simpler curvature of the main beam. It differs from that of M. hengduanshanensis from Yunnan (Zong et al., 1996) by thinner and much longer main beam.The upper dentition (V 26267.2) is generally close to those of M. bohlini (RV 4002) from Loc. 18 near Beijing (Teilhard de Chardin, 1940), e.g. presence of spur and entostyle. But its dimensions are slightly larger than those from Loc. 18. Compared with M. lacustris from Sanhe in Guangxi (Dong et al., 2011), V 26267.2 is evidently larger (Table 2), and the accessory elements such as neocrista and spur are obvious in V 26267.2 but weak in Sanhe specimens. V 26267.2 is metrically larger than extant Muntiacus sp. (OV 949) and the Late Miocene muntjaks from Zhaotong (Table 2). The lower dentitions, represented by two adult and a young mandibular fragments (V 26267.3-5), are close to that of M. bohlini from Xiashagou (Teilhard de Chardin and Piveteau, 1930) and Loc. 18 near Beijing (Teilhard de Chardin, 1940) both morphologically and metrically (Table 3). For example, in the p3 and p4, the metaconid is rounded and not fused with paraconid; the paraflexid, trigonid basin, entoflexid, talonid basin and hypoflexid are all present and open in specimens from Taijiaping, Xiashagou and Loc. 18. And for molars, the precingulum is present but undeveloped while ectostylid is developed in specimens from Taijiaping, Xiashagou and Loc. 18. Compared with M. lacustris from Juyuandong and Sanhe in Guangxi and Muntiacus sp. from Longgudong in Hubei, Taijiaping lower dentitions are obviously larger (Table 3).Both described antler and dentitions are generally in accordance with the diagnosis of M. bohlini and evidently different from other muntjaks and can be included into this taxon.Muntiacus is placed in Muntiacinae under Cervidae based on morphology (e.g. Vislobokova, 1990; Wang, 2003; Azanza et al., 2013; Dong et al., 2018) while Gilbert et al. (2006) suggested to replace the genus into Cervinae based on their analyses of two mitochondrial protein-coding genes and two nuclear introns. But the replacement is not yet widely accepted and we follow the classical classification in the present work.

(Figs. 3-4; Tables 4-6)
Material A right antler fragment with two broken tines (IVPP V 26268.1); a right maxillary fragment with DP4-M2 (V 26268.2) and a right maxillary fragment with DP2-4 (V 26268.3); a right mandibular fragment with dp2-m2 (V 26268.4), a right mandibular fragment with dp3-m3 (V 26268.5) and a right mandibular fragment with dp3-m1 (V 26268.6), all from Taijiaping, Tianzhen, Shanxi.Description The specimen V 26268.1 is a right antler fragment with two broken tines (Fig. 3A-B). The pedicle emerges from the frontal just above the orbit and inclines backwards (Fig. 3B). The pedicle is thinner than antler base and with moderate length (Table 4). The cross sections of the pedicle are oval and the surface of the pedicle is smooth. The burr is not very developed based on the preserved part and is composed of a series of bony nodes. The brow tine is set a short distance above the burr. The main beam is straight as a continuation from the antler base in medial view, but curves smoothly upwards from the antler base in lateral view. The surface of the antler is ornamented with longitudinal crests and furrows.
Fig. 3
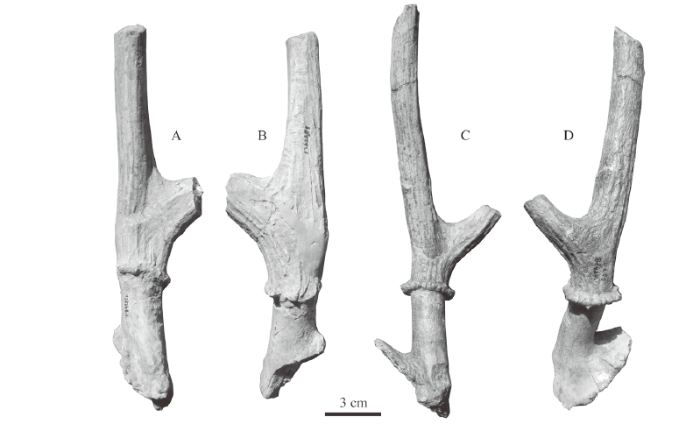 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Comparison of antler of Cervavitus cf. C. huadeensis from Xinyaozi Ravine with that of C. shanxius from Yushe
A, B. right antler of Cervavitus cf. C. huadeensis (IVPP V 26268.1) from Taijiaping in medial (A) and ventral-lateral (B) views; C, D. the antler of C. shansius (V 9378) from Hounao of Yushe in medial (C) and lateral (D) views
Table 4
Table 4Measurements of antler of Cervavitus cf. C. huadeensis from Xinyaozi Ravine and comparison (mm)
| Taxon | C. cf. C. huadeensis | C. shanxius | Taxon | C. cf. C. huadeensis | C. shanxius | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Locality | Taijiaping | Yushe | Tuchengzi1) | Locality | Taijiaping | Yushe | |||||
| APD pedicle | 28.9 | 21.7 | 27.8 | Bifurcation angle (°) | 70 | 60 | |||||
| TD pedicle | 27.6 | 20.7 | 26.6 | APD stem2 | 26.7 | 20.0 | |||||
| L pedicle | 36.3 | 36.5 | 45.5 | TD stem2 | 24.8 | 19.46 | |||||
| APD burr | 35.8 | 35.7 | 37.8 | L stem2 | >105 | >140 | |||||
| TD burr | 37.2 | 33.1 | 36.1 | C stem2 | 83.9 | 67 | |||||
| H burr | 7.6 | 7.1 | 7.6 | APD tine1 | 31.1 | 16.6 | |||||
| APD stem1 | 40.2 | 31 | 26.2 | TD tine1 | 18.7 | 13.9 | |||||
| TD stem1 | 29.1 | 21 | 22.9 | C tine1 | 82.5 | 53 | |||||
| L stem1 | 71.3 | 36 | L total(burr-tip) | >171 | >180 | ||||||
| C stem1 | 113.2 | 77 | |||||||||
新窗口打开|下载CSV
The DP2 appears as an elongated triangle and the DP3 appears also as a triangle but wider in occlusal view. Both DP2 and DP3 are composed of four selenodont main cusps. Their precingulum is very developed, entocingulum is moderately developed but entostyle (lingual basal pillar) is absent. The DP4 is molarized and composed of four selenodont main cusps (Fig. 4A-B). Its precingulum and entocingulum are moderately developed, and the entostyle and spur are developed. The M1 and M2 are all composed of four main cusps; their length and width are close to each other. The neocrista and entocingulum are visible but not developed, while mesostyle is well developed, entostyle and spur are developed. Measurements of the upper cheek teeth are listed in Table 5.
Fig. 4
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Cheek teeth of Cervavitus cf. C. huadeensis from Xinyaozi Ravine
A, B. right maxillary fragments in lingual and occlusal views: A. IVPP V 26268.2, with DP4-M2,B. V 26268.3, with DP2-4; C, D. right mandibular fragments in occlusal and buccal views: C. V 26268.4, with dp2-m2, D. V 26268.5, with dp3-m3
Table 5
Table 5Measurements of upper cheek teeth of Cervavitus cf. C. huadeensis from Xinyaozi Ravine and comparison (mm)
| C. cf. C. huadeensis | C. fenqii | C. shanxius | |||||||
|---|---|---|---|---|---|---|---|---|---|
| V 26268.2 | V 26268.3 | Juyuandong1) | Longgudong2) | Renzidong3) | Tuchengzi4) | Hounao5) | |||
| DP2 L | 16.17 | 11.05-11.80 | |||||||
| DP2 W | 11.96 | 7.75-8.05 | |||||||
| DP3 L | 18.50 | 12.3-14.7 | 11.60-14.05 | ||||||
| DP3 W | 13.75 | 12.1-15.4 | 7.55-12.10 | ||||||
| DP4 L | 17.72 | 16.68 | 6.18-13.34 | 8.10-14.40 | |||||
| DP4 W | 16.42 | 15.62 | 5.72-13.48 | 10.15-13.80 | |||||
| M1 L | 17.99 | 14.8-17.0 | 12.4-19.0 | 16.81 | 14.22-17.5 | 11.25-16.10 | |||
| M1 W | 20.18 | 17.5-19.6 | 14.4-20.4 | 16.55 | 14.38-17.8 | 13.40-16.70 | |||
| M1 H | 17.66 | 13.66 | 5.54-13.8 | 4.10-12.90 | |||||
| M2 L | 20.51 | 16.8-20.4 | 16.8-23.1 | 18.55 | 16.4-18.9 | 13.00-17.50 | |||
| M2 W | 19.13 | 16.0-20.2 | 17.4-22.4 | 18.54 | 17.5-20.8 | 15.30-18.50 | |||
| M2 H | 19.83 | 15.93 | 7.9-14.5 | 3.10-13.50 | |||||
| DP2-4 L | 33.20-39.30 | ||||||||
新窗口打开|下载CSV
The dp2 (Fig. 4C) is dominated by a main selenodont cusp (protoconid). Its crown is little worn and very low when compared with the permanent teeth. The dp3 is dominated by two main cusps (protoconid and hypoconid). Its trigonid basin is widely open but talonid basin is very narrow. The dp4 is molarized and composed of three lobes with developed buccal basal pillars. The m1 and m2 are composed of four selenodont main cusps with evident precingulid and developed ectostylid. The m3 is composed of three lobes. Its anterior two lobes resemble m1 and m2, and the third lobe is small (Fig. 4D). Measurements of the lower cheek teeth are listed in Table 6.
Table 6
Table 6Measurements of the lower cheek teeth of Cervavitus cf. C. huadeensis from Xinyaozi Ravine and comparison (mm)
| C. cf. C. huadeensis | C. huadeensis | C. fenqii | C. shanxius | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V 26268.4 | V 26268.5 | V 26268.6 | Tuchengzi1) | Juyuandong2) | Longgudong3) | Renzidong4) | Tuchengzi1) | Hounao5) | ||||
| dp2 L | 10.07 | 9.4 | 8.26-9.72 | 7.50-10.35 | ||||||||
| dp2 W | 5.08 | 5.1 | 3.96-5.18 | 3.75-5.45 | ||||||||
| dp3 L | 15.20 | 14.88 | 14.3 | 11.58-13.42 | 10.35-13.70 | |||||||
| dp3 W | 7.10 | 7.20 | 7.52 | 5.72-6.42 | 4.40-6.70 | |||||||
| dp4 L | 21.98 | 20.20 | 22.22 | 21.98 | 17.36-19.6 | 6.70-8.80 | ||||||
| dp4 W | 10.60 | 9.92 | 9.50 | 10.0 | 8.1-8.52 | 6.70-8.80 | ||||||
| m1 L | 17.15 | 18.03 | 19.17 | 19.42 | 12.9-15.7 | 14.4-15.8 | 14.58-17.38 | 12.56-16.7 | 11.80-15.70 | |||
| m1 W | 02.30 | 11.44 | 10.50 | 11.2 | 10.5-12.0 | 9.5-11.0 | 11.09-11.34 | 9.38-10.8 | 8.50-11.00 | |||
| m1 H | 14.07 | 14.61 | 13.50 | 15.3 | 6.48-11.81 | 3.4-11.4 | 4.30-11.20 | |||||
| m2 L | 21.54 | 20.19 | 23.78 | 16.8-19.5 | 16.8-18.3 | 18.27-19.21 | 15.1-18.7 | 13.90-17.20 | ||||
| m2 W | 12.47 | 12.25 | 13.6 | 12.2-13.2 | 11.3-12.2 | 10.80-12.42 | 10.78-12.36 | 9.80-12.00 | ||||
| m2 H | 21.44 | 19.06 | 16.7 | 10.92 | 4.9-15.0 | 4.90-13.00 | ||||||
| m3 L | 24.47 | 22.6-23.0 | 22.4-26.7 | 24.60-27.09 | 20.3-22.9 | 17.70-24.20 | ||||||
| m3 W | 11.08 | 11.6-14.6 | 10.5-14.1 | 12.07-12.72 | 11.12-11.78 | 9.25-11.60 | ||||||
| m3 H | 17.30 | 13.62-20.90 | 5.68-13.0 | 5.70-13.00 | ||||||||
| dp2-4 L | 44.80 | 45.34 | 36.28-40.7 | 33.70-42.10 | ||||||||
| m1-3 L | 67.76 | 48.4-56.1 | 44.90-54.60 | |||||||||
新窗口打开|下载CSV
Comparison and determination The described antler (IVPP V 26268.1) is evidently larger than that of M. bohlini from Shuichongkou, but it obviously smaller than the antlers of the other cervids from Xinyaozi Ravine. It is morphologically close to that (V 9378, Fig. 3C-D) of Cervavitus shanxius from Hounao in Yushe Basin (Dong and Hu, 1994) and that from Tuchengzi (Dong et al., 2018). For example, the pedicle is moderately long and inclines backwards, the brow tine does not emerge from the burr but from the base a short distance above the burr, the first bifurcation angle is moderate. However, Hounao specimen is evidently thinner than Taijiaping one (Fig. 3; Table 4). Compared with C. huadeensis from Tuchengzi (Qiu, 1979; Dong et al., 2018), Taijiaping specimen is similar to Tuchengzi specimen V 5634.3 (Qiu, 1979), e.g. the pedicle is not long, the antler base or stem 1 is neither long, the main beam is straight in medial view. But the brow tine is set quite high above the burr in other specimens of C. huadeensis (Qiu, 1979) and different from that of Taijiaping. Taijiaping antler differs from those of C. fenqii from Juyuandong (Han, 1987), Longgudong (Chen, 2004) and Renzidong (Dong et al., 2009) by larger dimensions. It differs from those of C. ultimus (Lin et al., 1978) by its larger angle of the first bifurcation.
The described upper cheek teeth are characterized by the presence of neocrista, spur and entostyle. The neocrista is present in C. shanxius from Hounao and Tuchengzi but more developed than in Taijiaping specimens. The spur and entostyle are similarly developed in Taijiaping as well as in Hounao and Tuchengzi specimens. The upper dentition of C. huadeensis is not yet available from Tuchengzi specimens. Compared with C. fenqii from Juyuandong (Han, 1987), Longgudong (Chen, 2004) and Renzidong (Dong et al., 2009), Taijiaping specimens also have developed entostyle, but have evident neocrista and developed spur which are absent in Juyuandong and Longgudong specimens. In addition, Taijiaping specimens are slightly larger (Table 5). The lower cheek teeth from Taijiaping are generally similar to those of C. huadeensis from Tuchengzi (Dong et al., 2018) both morphologically and metrically (Table 6). Taijiaping specimens are evidently larger than those of C. fenqii from Juyuandong, Longgudong and Renzidong, as well as those of C. shanxius from Hounao and Tuchengzi (Table 6).
The comparison between Taijiaping specimens and those of Cervavitus from other localities indicates that Taijiaping specimens are closer to those of C. huadeensis from Tuchengzi than those of C. fenqii and C. shanxius. But there are still some differences between Taijiaping specimens and those of C. huadeensis, e.g., the ornamentation of antler in Tuchengzi antlers are more developed than in Taijiaping specimens, ectostylid is less developed in Tuchengzi lower molars. In addition, C. huadeensis are previously uncovered from the Neogene deposits rather than in the Pleistocene ones, and the material from Taijiaping is limited to a broken antler and some fragmental jaws. We place them temporarily as a conformis species of C. huadeensis.

(Figs. 5-6; Tables 7-9)
Axis shansius Teilhard de Chardin and Trassaert, 1937, p. 44-50
Axis cf. shansius Chi, 1975, p. 173
Axis shansius Chia and Wang, 1978, p. 21-23
Axis shansius Dong and Fang, 2004, p. 299-302
Axis shansius Dong, 2006, p. 335-338
Axis shansius Bai et al., 2017, p. 821-823
Material A large part of a left antler with partial frontal (IVPP V 26264.1); a left maxillary fragment with DP2-M1 (V 26264.2), a left maxillary fragment with a broken DP3 and complete DP4 and M1 (V 26264.3), a left maxillary fragment with DP3-4 (V 26264.4); a right mandibular fragment with p2-m3 (V 26264.5), a left mandibular fragment with a broken p2 and completed p3-m3 (V 26264.6), a left mandibular fragment with dp2-m1 (V 26264.7). All specimens are from Taijiaping, Tianzhen, Shanxi.
Description The specimen V 26264.1 is a large part of a left antler with partial frontal (Fig. 5; see Table 7 for measurements). The pedicle is relatively short, and its cross sections are oval. The burr is composed of a series of developed bony nodules. The antler base, or stem 1, is relatively short. The distance between the first bifurcation and the burr is similar to that between the frontal and burr. The brow tine is long and curved in the middle. The main beam is very long and lyrately curved. The first bifurcation angle is very large, but the second one is relatively small. The second and third tines are broken near their base. The surface of the antler is ornamented with developed longitudinal crests and furrows.
Fig. 5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Lateral view (left) and medial view (right) of the left antler of Axis shansius (IVPP V 26264.1) from Taijiaping
Table 7
Table 7Measurements of antler of Axis shansius from Taijiaping and comparison (mm)
| V 26264.1 | Yushe Larger1) | Yushe Smaller1) | Xihoudu2) | Jinyuandong3) | |
|---|---|---|---|---|---|
| APD pedicle | 38.1 | ||||
| TD pedicle | 42.1 | 32-43 | 26 | 55 | |
| L pedicle | 53.3 | 37-50 | 40 | 44 | |
| APD burr | 69.6 | 50-58 | 37-48 | 63-78 | |
| TD burr | 62.5 | ||||
| H burr | 10.4 | ||||
| APD base | 60.3 | 35-45 | 26-35 | 55-73 | |
| TD base | 55.4 | ||||
| APD stem1 | 67.1 | ||||
| TD stem1 | 49.9 | ||||
| L stem1 | 81.1 | 66-130 | 67-74 | 62-94 | |
| C stem1 | 197.2 | ||||
| APD stem2 | 45.3 | 28-42 | 22-25 | 41-51 | 42.31 |
| TD stem2 | 42.5 | 39.80 | |||
| L stem2 | 506.2 | 430-507 | 255-315 | 360-470 | 245 |
| C stem2 | 149.6 | ||||
| APD tine1 | 40.7 | ||||
| TD tine1 | 36.1 | ||||
| L tine1 | 250.1 | 162-280 | 111-127 | 196-226 | |
| C tine1 | 137.4 | ||||
| APD tine2 | 28.3 | 14-34 | 12-15 | 36.5 | |
| TD tine2 | 28.2 | 29.1 | |||
| APD tine3 | 28.7 | 20-34 | 14-25 | 36.6 | |
| TD tine3 | 32.9 | 28.3 | |||
| L total (burr-tip) | >607 | 724-985 | 461-490 | ||
| Angle 1st bifurcation | 105° | 63-117° | 84-110° | 85-140° | |
| Angle 2nd bifurcation | 60° | 75° |
新窗口打开|下载CSV
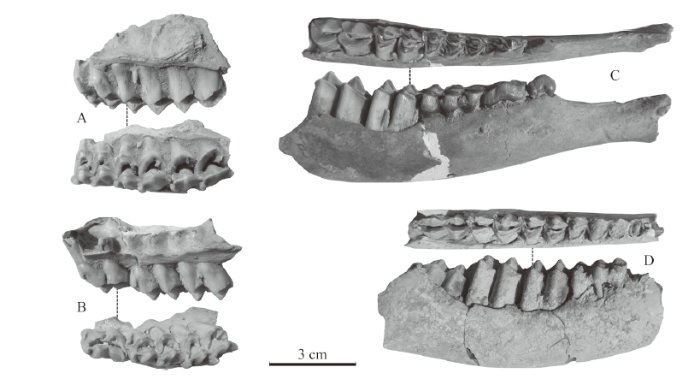
The DP2 appears as an elongated triangle (Fig. 6A) and the DP3 (Fig. 6A, C) also but wider in occlusal view. Both DP2 and DP3 are composed of four selenodont main cusps. Their precingulum is very weak, entocingulum is moderately developed and entostyle (lingual basal pillar) is present. The DP4 is molarized and composed of four selenodont main cusps (Fig. 6A-C). Its precingulum is absent, and its entocingulum is moderately developed, while the entostyle and spur are developed. The M1 are all composed of four main cusps; their length and width are close to each other (Fig. 6A-B). The precingulum, neocrista and entocingulum are visible but very weak, while mesostyle, entostyle and spur are developed. The M2 and M3 are not available but might be similar to the M1. Measurements of the upper cheek teeth are listed in Table 8.
Fig. 6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Cheek teeth of Axis shansius from Taijiaping
A. IVPP V 26264.2, a left maxillary fragment with DP2-M1 in lingual and occlusal views; B. V 26264.3, a left maxillary fragment with a broken DP3 and complete DP4 and M1 in buccal and occlusal views;C. V 26264.4, a left maxillary fragment with DP3-4 in buccal and occlusal views;D. V 26264.5, a right mandibular fragment with p2-m3 in occlusal and lingual views; E. V 26264.6, a left mandibular fragment with a broken p2 and completed p3-m3 in occlusal and buccal views;F. V 26264.7, a left mandibular fragment with dp2-m1 in occlusal and buccal views
Table 8
Table 8Measurements of upper cheek teeth of Axis shansius from Taijiaping and comparison (mm)
| V 26264.2 | V 26264.3 | V 26264.4 | Yushe1) | Tuozidong2) | Dajushan3) | |
|---|---|---|---|---|---|---|
| DP2 L | 15.28 | |||||
| DP2 W | 11.91 | |||||
| DP3 L | 18.56 | 20.15 | ||||
| DP3 W | 14.27 | 15.5 | 16.28 | |||
| DP4 L | 18.90 | 19.41 | 20.43 | |||
| DP4 W | 17.16 | 19.13 | 18.49 | |||
| M1 L | 22.83 | 22.92 | 18 | 19.74-21.99 | 18.20-19.22 | |
| M1 W | 21.25 | 22.79 | 17-19 | 19.71-23.07 | 16.90-19.20 | |
| M1 H | 21.09 | 19.45 | 8.10-14.75 | 15.34-16.00 | ||
| DP2-4 L | 52.5 |
新窗口打开|下载CSV
The dp2 (Fig. 6F) is dominated by a main selenodont cusp (protoconid). The dp3 (Fig. 6F) is dominated by two main cusps (protoconid and hypoconid). Its trigonid basin is widely open but talonid basin is very narrow. The dp4 (Fig. 6F) is molarized and composed of three lobes with developed buccal basal pillars. The p2 (Fig. 6D) is dominated by a main selenodont cusp (protoconid), its trigonid and talonid basins are weak but entoflexid is very developed. The p3 (Fig. 6D-E) is also dominated by protoconid. The trigonid basin is well developed and widely open. The entoflexid and talonid basin are very narrow. The p4 (Fig. 6D-E) is dominated by protoconid and metaconid. The metaconid does not extend forwards but backwards. The trigonid basin is moderately developed and open. The entoflexid and talonid basin are narrow but still open. The m1 (Fig. 6D-F) and m2 (Fig. 6D-E) are composed of four selenodont main cusps. The m3 (Fig. 6D-E) is composed of three lobes. Its anterior two lobes resemble the m1 and m2, and the third lobe is small. The Palaeomeryx fold and goat fold are absent in all molars, but precingulid is present and weak on the m1 and absent on the m2 and m3. The ectostylid (buccal basal pillar) is developed on the m1, evident on the m2 and fully-grown on the m3. Measurements of the lower cheek teeth are listed in Table 9.
Table 9
Table 9Measurements of lower cheek teeth of Axis shansius from Taijiaping and comparison (mm)
| V 26264.5 | V 26264.6 | V 26264.7 | Yushe1) | Tuozidong2) | Dajushan3) | |
|---|---|---|---|---|---|---|
| dp2 L | 9.78 | |||||
| dp2 W | 5.11 | |||||
| dp3 L | 14.52 | |||||
| dp3 W | 7.02 | |||||
| dp4 L | 21.90 | |||||
| dp4 W | 9.18 | |||||
| p2 L | 13.17 | 13.9 | 12.79-15.59 | 11.98 | ||
| p2 W | 8.68 | >6.0 | 6.92-7.77 | 6.96 | ||
| p2 H | 9.58 | 7.00-10.31 | 10.20 | |||
| p3 L | 14.94 | 15.98 | 15.0 | 16.00-20.95 | 17.30 | |
| p3 W | 9.81 | 8.30 | 9.2 | 8.45-11.78 | 8.70 | |
| p3 H | 11.13 | 12.73 | 9.3 | 8.05-14.31 | 12.10 | |
| p4 L | 16.58 | 16.56 | 16.8 | 16.48-22.38 | 16.90 | |
| p4 W | 11.09 | 9.32 | 9.5 | 10.12-14.09 | 9.50 | |
| p4 H | 12.22 | 12.35 | 9.4 | 7.96-15.71 | 15.30 | |
| m1 L | 16.94 | 18.23 | 18.24 | 18.80-23.82 | 16.70-19.70 | |
| m1 W | 12.56 | 12.13 | 10.64 | 11.49-15.68 | 8.70-11.30 | |
| m1 H | 10.50 | 12.61 | 12.71 | 3.82-15.69 | 12.40-13.80 | |
| m2 L | 20.47 | 19.77 | 20.0 | 19.13-26.13 | 18.32-21.30 | |
| m2 W | 14.31 | 12.20 | 14.0 | 13.53-17.05 | 11.60-14.30 | |
| m2 H | 15.53 | 13.32 | 10.0 | 5.51-15.80 | 10.60-16.80 | |
| m3 L | 27.19 | 26.57 | 28.5 | 26.09-32.70 | 24.76-28.36 | |
| m3 W | 14.17 | 12.85 | 14.1 | 13.14-15.70 | 8.60-11.38 | |
| m3 H | 15.01 | 15.63 | 13.8 | 11.60-16.00 | ||
| dp2-4 L | 46.30 | 47.50 | ||||
| p2-4 L | 44.67 | 46.8 | 45.41-52.31 | |||
| m1-3 L | 66.33 | 62.61 | 65 | 70.89-76.53 | 67.00 | |
| p2-m3 L | 112.33 | 113.6 | 110 | 121.52-126.28 |
新窗口打开|下载CSV
Comparison and determination The described antler has unbranching brow tine and bifurcated posterior beam that distinguishes it from Elaphurus although its size is similar to that of the latter. It differs from stratigraphically associated Cervavitus cf. C. huadeensis by its long and lyrated main beam. It fits well the diagnosis of Axis shansius (Teilhard de Chardin and Trassaert, 1937) by its lyrated main beam with three tines and widely opened first fork, as well as its very long first tine (brow tine). Compared with A. shansius from Yushe, Taijiaping antler is very close to the antler of THP 12387 (=No. 12.387) (Teilhard de Chardin and Trassaert, 1937), although its main beam is less curved than that of the type. The dimensions of Taijiaping antler falls well in variation range of antlers of A. shansius from Yushe (Table 7). Both upper and lower dentitions of Taijiaping specimens are also metrically similar to those of A. shansius from Yushe (Tables 8-9). Taijiaping antler is also close to that of A. shansius from Xihoudu (Chia and Wang, 1978), e.g. three-tined antler, and lyrated main beam. But the first bifurcation of Xihoudu specimens is closer to the burr, and the first bifurcation angle is larger. Taijiaping antler is similar to that of A. shansius from Tuozidong (Dong and Fang, 2004) and Jinyuandong (Bai et al., 2017), e.g. the curvature of the main beam and the form of the second bifurcation. Both upper and lower dentitions of Taijiaping specimens share the same morphological traits with those from Tuozidong and Dajushan (Dong, 2006), e.g. the accessary elements such as precingulum, entostyle, spur etc. are present. Their dimensions are also close to each other (Tables 8-9).Compared with other species of Axis, Taijiaping specimens are much larger than those of A. speciosus from Ertemte in Nei Mongol (Schlosser, 1924). For example, the main beam of Taijiaping specimen is twice longer, the burr is twice thicker, the dentitions are nearly one and half larger. Taijiaping antler is moderately longer and thicker than that of A. rugosus from Tongshan at Yuanqu in Shanxi (Chow, 1954). In addition, the brow tine is set very close to the burr (as in extant A. axis), the furrow ornamentation is very developed in Tongshan antler. Taijiaping antler is moderately longer but slightly thinner than that of A. lingjingensis from Lingjing Man site in Henan (Dong and Li, 2008, Bai et al., 2017), and its first bifurcation angle is larger than that in Lingjing specimens. Taijiaping specimens are therefore morphologically and metrically closest to A. shansius and clearly different from other species of Axis.

(Figs. 7-8; Tables 10-12)
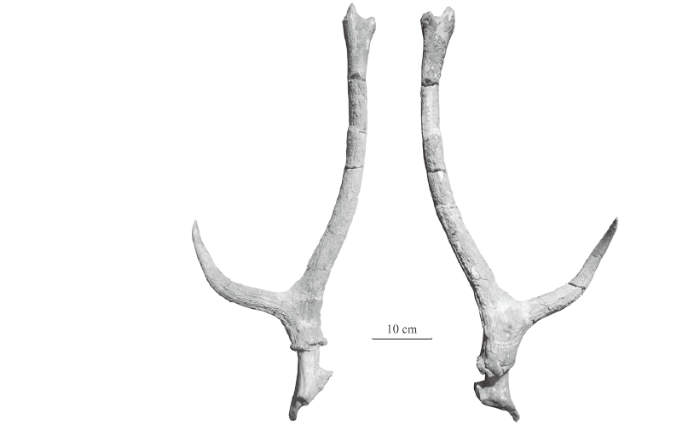
Cervus (Rusa) elegans Teilhard de Chardin and Piveteau, 1930, p. 54-63Rusa elegans Wei, 1997, p. 196Material A broken skull with a pair of broken antlers (IVPP V 26265.1), a broken right antler with pedicle and partial frontal (V 26265.2), a broken left antler with pedicle and partial frontal (V 26265.3); specimens V 26265.1-3 are from Taijiaping. A left maxillary fragment with P2-M3 (V 26265.4), a right maxillary fragment with P2-M3 (V 26265.5), a left maxillary fragment with P3-M3 (V 26265.6), a right maxillary fragment with P2-4 (V 26265.7), a right mandibular fragment with p4-m3 (V 26265.8), a left mandibular fragment with p2-m3 (V 26265.9), a left mandibular fragment with p3-m2 (V 26265.10) and a right mandibular fragment with p4-m3 (V 26265.11) from Xinyaozi Ravine, Tianzhen, Shanxi.Description Although the referred antlers are all broken, the length of broken main beams indicates the antlers bifurcate at least twice and with at least three tines. The specimen V 26265.1 is a broken skull with a pair of broken antlers (Fig. 7A-B). The frontals are flat and wide in anterior view. Two pedicles, relatively short, emerge from lateral-posterior parts of the frontals above the orbits and extend backwards and laterally. The distance between the bases of two pedicles measures 37 mm. The angle between two pedicles is about 60°. The cross sections of the pedicles are oval. The burrs are composed of a series of bony nodules and moderately developed although partially broken. The antler base or stem 1 of the pair is well preserved and moderately long. The first bifurcation angle is 50°-65°. The first tine (brow tine) is relatively long (Table 10) and slightly curves medially. Both left and right first tines are broken near their tips. Both left and right main beams are also broken. The surface of the antler is ornamented with longitudinal crests and furrows.
Fig. 7
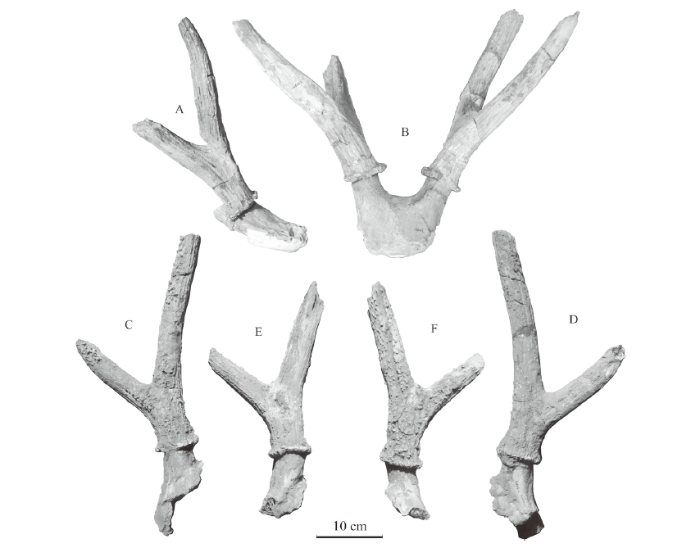 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 7Antlers of Nipponicervus elegans (IVPP V 26265.1-3) from Taijiaping
A, B. IVPP V 26265.1, a broken skull with a pair of broken antlers: A. lateral view of the right antler, B. anterior view; C, D. V 26265.2, a broken right antler with pedicle and partial frontal in medial (C) and lateral (D) views; E, F. V26265.3, a broken left antler with pedicle and partial frontal in lateral (E) and medial (F) views
Table 10
Table 10Measurements of antlers of Nipponicervus elegans from Taijiaping and comparison (mm)
| V 26265.1L | V 26265.1R | V 26265.2 | V 26265.3 | Xiashagou1) | Yushe2) | Longdan3) | |
|---|---|---|---|---|---|---|---|
| APD pedicle | 34.3 | 35.9 | 45.9 | 40.9 | 32 | 48 | |
| TD pedicle | 40.5 | 39.7 | 44.2 | 43.8 | 41 | ||
| L pedicle | 39.4 | 38.3 | 53.8 | 39.3 | 40-42 | 30 | |
| APD burr | 58.7 | 67.7 | 62.7 | 50 | 38 | ||
| TD burr | 62.3 | 65.2 | 63.2 | ||||
| H burr | 6.9 | 9.6 | 10.2 | 9.4 | |||
| APD base | 45.2 | 46.1 | 55.1 | 50.7 | |||
| TD base | 46.1 | 50.6 | 50.4 | 50.2 | |||
| APD stem1 | 44.6 | 53.1 | 58.3 | 53.1 | 55 | ||
| TD stem1 | 41.5 | 41.8 | 46.6 | 41.3 | |||
| L stem1 | 108.9 | 88.9 | 92.7 | 94.2 | 67-95 | 115 | 56 |
| C stem1 | 145.2 | 153.2 | 164.5 | 166.4 | 125-192 | ||
| APD stem2 | 33.3 | 35.7 | 37.6 | 40.4 | 27 | 34 | 48 |
| TD stem2 | 30.6 | 34.6 | 35.4 | 38.3 | 31 | ||
| L stem2 | >209.2 | >111.2 | >238.6 | >182.3 | 225 | 382 | 517 |
| C stem2 | 115.9 | 117.2 | 122.3 | 127.6 | |||
| APD tine1 | 32.6 | 33.8 | 36.4 | 34.5 | 39 | ||
| TD tine1 | 29.2 | 39.6 | 42.3 | 29.3 | |||
| L tine1 | 225.3 | >160.4 | 310.9 | >111.7 | 114 | 117 | 310 |
| C tine1 | 98.9 | 111.5 | 112.2 | 109.1 | |||
| L total(burr-tip) | >302.1 | >199.3 | >318.9 | >256.9 | 380 | 800 | |
| Angle 1st bifurcation | 65° | 50° | 65° | 65° | 60° | 45° | 80° |
新窗口打开|下载CSV
The specimen V 26265.2 is a broken right antler with pedicle and partial frontal (Fig. 7C-D). It is very similar to the right antler of the specimen V 26265.1 described above, e.g. relatively short pedicle with oval cross sections, burr formed with a ring of bony nodules, antler base and the first tine moderately long (Table 10). But its main beam continuously curves laterally and posteriorly instead of laterally and upwards. It is also ornamented with longitudinal crests and furrows and with some bony nodules or pearled on medial side.
The specimen V 26265.3 is a broken left antler with pedicle and partial frontal (Fig. 7E-F). It is also very similar to the specimen V 26265.1 (see Table 10 for measurements). But its ornamentation has many bony nodules on medial side in addition to longitudinal crests and furrows.
The P2 is semi-molarized, the lingual main cusps (protocone and metaconule) are separated by entoflexus, but the buccal main cusps (paracone and metacone) are fused end to end (Fig. 8A-B). The neocrista is developed, entocingulum is very weak, but the precingulum, entostyle and spur are absent. The P3 is similar to P2, but with larger dimensions (Fig. 8A-C). The P4 is one lobed and evidently different from P2 and P3, the anterior and posterior main cusps on both lingual and buccal sides are fused end to end; the precingulum, entocingulum, entostyle and spur are all absent, but neocrista is still present and developed (Fig. 8A-C). The M1 is composed of four main cusps; its length and width are close to each other (Fig. 8A-C). The neocrista is absent, precingulum and entocingulum are visible but not developed, while mesostyle, entostyle and spur are developed. The morphology of M2 is similar to that of M1, but its size is larger than that of M1 and its length is slightly larger than its width; there is a fold on the lingual side of postprotocrista (posterior crest of protocone) equivalent to the Palaeomeryx fold of lower molar. It differs from neocrista by its position on the lingual side, instead of buccal side, of postprotocrista. The morphology of M3 is also similar to that of the M1, but its length is evidently larger than its width. Measurements of the upper cheek teeth are listed in Table 11.
Fig. 8
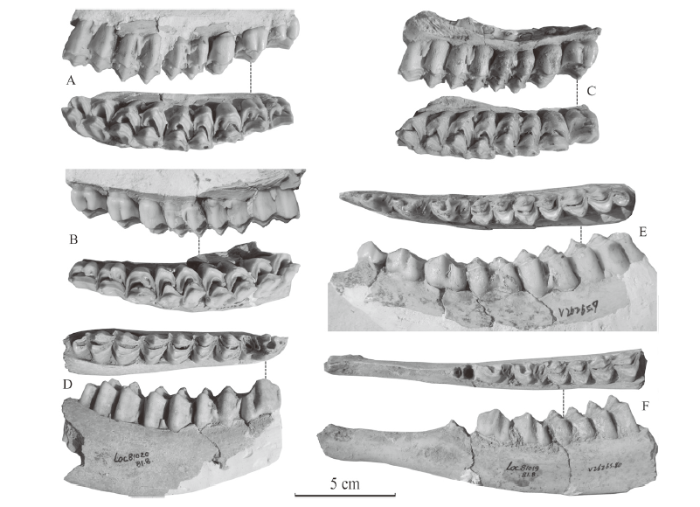 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 8Cheek teeth of Nipponicervus elegans from Xinyaozi Ravine
A. IVPP V 26265.4, a left maxillary fragment with P2-M3 in lingual and occlusal views; B. V 26265.5, a right maxillary fragment with P2-M3 in lingual and occlusal views; C. V 26265.6, a left maxillary fragment with P3-M3 in lingual and occlusal views; D. V 26265.8, a right mandibular fragment with p4-m3 in occlusal and buccal views; E. V 26265.9, a left mandibular fragment with p2-m3 in occlusal and buccal views; F. V 26265.10, a left mandibular fragment with p3-m2 in occlusal and buccal views
Table 11
Table 11Measurements of upper cheek teeth of Nipponicervus elegans from Xinyaozi Ravine and comparison (mm)
| N. elegans | N. longdanensis | Arvernoceros ardei | |||||||
|---|---|---|---|---|---|---|---|---|---|
| V 26265.4 | V 26265.5 | V 26265.6 | V 26265.7 | Xiashagou1) | Longdan2) | Etouaires3) | |||
| P2 L | 16.56 | 17.62 | 16.42 | 16.5 | 14-16 | ||||
| P2W | 16.70 | 16.39 | 15.30 | 14 | 13.5-17.5 | ||||
| P2 H | 13.64 | 14.68 | 15.96 | ||||||
| P3 L | 16.59 | 17.03 | 16.31 | 16.13 | 19 | 14-15.5 | |||
| P3 W | 18.85 | 19.08 | 18.50 | 17.82 | 17.8 | 15.5-17.5 | |||
| P3 H | 13.74 | 16.07 | 15.82 | 19.15 | |||||
| P4 L | 15.15 | 15.24 | 14.16 | 14.73 | 17 | 13.5-15.5 | |||
| P4 W | 19.38 | 18.68 | 20.20 | 19.18 | 20.2 | 17-20 | |||
| P4 H | 14.68 | 14.95 | 15.04 | 16.32 | |||||
| M1 L | 22.98 | 22.61 | 20.64 | 28 | 19-22 | ||||
| M1 W | 22.98 | 22.28 | 20.91 | 25 | 20-23.5 | ||||
| M1 H | 15.75 | 17.53 | 13.04 | ||||||
| M2 L | 24.42 | 24.62 | 22.85 | 26 | 19-25.5 | ||||
| M2 W | 23.26 | 23.10 | 23.93 | 23.5 | 21.5-26.5 | ||||
| M2 H | 20.71 | 18.86 | 16.78 | ||||||
| M3 L | 24.90 | 25.38 | 23.05 | 27.3 | 20-24.5 | ||||
| M3 W | 21.43 | 22.45 | 22.90 | 25.5 | 21.5-25.5 | ||||
| M3 H | 19.80 | 19.92 | 17.64 | ||||||
| P2-4 L | 50.60 | 52.04 | 49.95 | 43-55 | 51 | 43-46.5 | |||
| M1-3 L | 70.9 | 71.61 | 63.39 | 60-77 | 72 | 58.5-68.5 | |||
| P2-M3 | 116.97 | 117.93 | 103-136 | 120 | 97.5-104.5 | ||||
新窗口打开|下载CSV
The p2 (Fig. 8E) is dominated by a main selenodont cusp (protoconid); the paraflexid is absent, its trigonid basin is developed and widely open, its entoflexid and talonid basins are very narrow, its hypoflexid is very weak. The p3 (Fig. 8E-F) is also dominated by protoconid; its paraflexid is present but very weak, its trigonid basin is well developed and widely open, its entoflexid and talonid basins are relatively narrow, its hypoflexid is moderately developed. The p4 (Fig. 8D-E) is dominated by protoconid and metaconid; its paraflexid is weak and open, its metaconid extends moderately forwards and backwards but leaves trigonid basin and entoflexid open; entoconid inclines backward and nearly closes talonid basin. The m1 (Fig. 8D-F) and m2 (Fig. 8D-F) are composed of four selenodont main cusps; the Palaeomeryx fold and goat fold are absent, but precingulid is present and weak, ectostylid (buccal basal pillar) is developed; the dimensions of m2 are evidently larger than those of m1. The m3 (Fig. 8D-E) is composed of three lobes. Its anterior two lobes resemble the m1 and m2, and the third lobe is small. Measurements of the lower cheek teeth are listed in Table 12.
Table 12
Table 12Measurements of lower cheek teeth of Nipponicervus elegans from Xinyaozi Ravine and comparison (mm)
| N. elegans | N. longdanensis | Arvernoceros ardei | |||||||
|---|---|---|---|---|---|---|---|---|---|
| V 26265.8 | V 26265.9 | V 26265.10 | V 26265.11 | Xiashagou1) | Longdan2) | Etouaires3) | |||
| p2 L | 14.41 | 17.52 | 11.5-15 | ||||||
| p2 W | 8.04 | 7.66 | 6.5-9.5 | ||||||
| p2 H | 8.96 | ||||||||
| p3 L | 17.36 | 18.44 | 15-20.5 | ||||||
| p3 W | 10.28 | 11.48 | 9-12.5 | ||||||
| p3 H | 11.72 | 14.20 | |||||||
| p4 L | 21.96 | 18.96 | 19.18 | 19.39 | 16-22 | 15-21 | |||
| p4 W | 12.29 | 11.18 | 12.35 | 10.61 | 10-14 | 9.5-13.5 | |||
| p4 H | 18.65 | 12.93 | 15.94 | 12.19 | |||||
| m1 L | 23.66 | 19.63 | 21.29 | 20.16 | 24.41 | 16.5-24 | |||
| m1 W | 15.94 | 13.85 | 14.24 | 14.67 | 13.73 | 12.5-15 | |||
| m1 H | 17.27 | 12.81 | 11.62 | 10.51 | 26.16 | ||||
| m2 L | 27.64 | 2.87 | 25.56 | 21.34 | 20.5-25.5 | ||||
| m2 W | 18.03 | 15.52 | 15.84 | 16.14 | 14-16.5 | ||||
| m2 H | 18.22 | 14.21 | 15.99 | 16.06 | |||||
| m3 L | 34.40 | 30.39 | 29.63 | 28-35 | |||||
| m3 W | 17.75 | 14.34 | 16.34 | 13-15.5 | |||||
| m3 H | 19.90 | 14.56 | 17.05 | ||||||
| p2-4 L | 50.57 | 51.68 | 52-56 | 43-53 | |||||
| m1-3 L | 82.37 | 72.51 | 71.45 | 70-89 | 65-77.5 | ||||
| p2-m3 L | 121.85 | 112-145 | 108-129.5 | ||||||
新窗口打开|下载CSV
Comparison and determination The type and associated specimens of Nipponicervus elegans, previously named as Cervus (Rusa) elegans, were collected from Xiashagou area, i.e. type locality, in Nihewan Basin (Teilhard de Chardin and Piveteau, 1930). Compared with Xiashagou specimens, Xinyaozi specimens are very close to those from type locality. For example, pedicle relatively short, the antler base is moderately long, the first bifurcation angle is moderate, the main beam is nearly straight, the brow tine is relatively long and slightly curves medially; the entoflexus is developed on P2 and P3, entostyle is developed on upper molars; trigonid basin is well open on p3 and p4 and ectostylid is developed on lower molars. The dimensions of antlers and dentitions are also very close (Tables 10-12). In addition, Xinyaozi specimens are geographically and stratigraphically close to Xiashagou specimens and they can be regarded as the same species. Some antlers from Yushe Basin were resigned to Cervus (Rusa) cf. elegans (Teilhard de Chardin and Trassaert, 1937). They were revised as N. elegans (Qiu et al., 2004). Xinyaozi specimens are also similar to those from Yushe.
Compared with N. longdanensis from Longdan (Qiu et al., 2004), Xinyaozi antlers are similar by relatively long brow tine, nearly straight main beam, but different by longer antler base, smaller angle of the first bifurcation (Table 10). The Xinyaozi dentitions are also similar to that from Longdan (Tables 11-12).
Nipponicervus elegans from Xinyaozi and Xiashagou are comparable with Cervus rhenanus (Heintz, 1970; de Vos et al., 1995) in Europe. However, the antlers of the latter are slenderer and the dentitions are smaller. N. elegans from Xinyaozi and Xiashagou are also comparable with Arvernoceros ardei from étouaires in France (Heintz, 1970), especially for dimensions of dentitions (Tables 11-12). Nevertheless, the main beams are much longer and more curved, and the entocingulum of upper cheek teeth is very developed in French specimens.
(Fig. 9; Table 13)
Material An incomplete right antler with pedicle and partial frontal (IVPP V 26266) from Dazhuangkecun (Dazhuangke village), about 3 km southwest of Taijiaping, Tianzhen, Shanxi.
Description The specimen V 26266 is a right adult antler with pedicle and partial frontal (Fig. 9). The pedicle is relatively short and evidently thinner than the antler base; its cross sections are oval. The burr is well developed and preserved. It is composed of a series of bony nodules. The antler base, Stem 1, is very short, with elongated oval cross sections. The brow tine is broken but its base is preserved. Its preserved part indicates that it emerges from the antler base, or stem 1, anteriorly and upwards. It is set very low, almost on the burr. The bez tine is absent, i.e. not developed at all. The trez tine is nearly complete, only its tip part is missing; it is moderately long and curves slightly upwards. Stem 2, the main beam between the first and second bifurcations, or between the bases of brow and trez tines, is relatively long; its cross sections are somewhat triangular near the first bifurcation and become elongated oval towards the second bifurcation. The part about 95 mm above the second bifurcation is missing. The ornamentation is composed of moderate longitudinal furrows and without crests.
Fig. 9
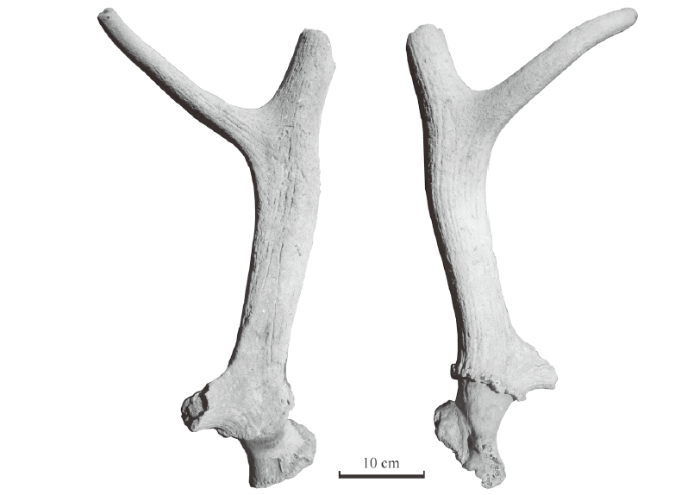 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 9Medial (left) and lateral (right) views of the right antler (IVPP V 26266) of Cervus elaphus from Dazhuangke
Comparison and determination The described antler is evidently different from the other specimens from Xinyaozi by its large size and large angle of the first bifurcation. It is very close to that of red deer, e.g. the brow tine set very low and almost on the burr, the angle between brow tine and the main beam is very large (Table 13), etc. But it has no bez tine as in some European red deer, such as Cervus elaphus aretinus from Veneto to Calabria in Italy (Abbazzi, 1995). Compared with C. canadensis mongoliae from northern China (Zdansky, 1925), Dazhuangke specimen lacks bez tine and is slightly larger (Table 13), the rest morphological characters are very similar. Compared with Cervus cf. C. canadensis from Yushu in northeastern China (Xue, 1959), besides the bez tine on the antlers from Yushu, Dazhuangke specimen appears more robust, but the burrs are more developed in Yushu specimens.
Table 13
Table 13Measurements of antler of Cervus elaphus from Dazhuangke and comparison with that of C. canadensis mongoliae (mm)
| V 26266 | C . canadensis mongoliae1) | V 26266 | C . canadensis mongoliae1) | |||
|---|---|---|---|---|---|---|
| APD pedicle | 56.7 | APD stem3 | 49.4 | |||
| TD pedicle | 54.5 | TD stem3 | 50.5 | |||
| L pedicle | 37.6 | L stem3 | >94.4 | 235 | ||
| APD burr | 90.8 | 81 | C stem3 | 164.3 | ||
| TD burr | 65.4 | APD tine1 | 39.1 | |||
| H burr | 11.1 | TD tine1 | 38.2 | |||
| APD base | 80.6 | 74 | L tine1 | >50 | ||
| TD base | 66.7 | C tine1 | 237 | |||
| APD stem1 | 85.5 | 72 | APD tine2 | 42.2 | ||
| TD stem1 | 56.2 | TD tine2 | 36.1 | |||
| L stem1 | 36.2 | L tine2 | >198.2 | |||
| C stem1 | 238 | C tine2 | 120.9 | |||
| APD stem2 | 53.1 | L total(burr-tip) | >403.5 | 1090 | ||
| TD stem2 | 55.1 | Angle 1st bifurcation | 120° | 120° | ||
| L stem2 | 282.5 | Angle 2nd bifurcation | 80° | 87° | ||
| C stem2 | 175.1 | Angle 3rd bifurcation | 80° |
新窗口打开|下载CSV
3 Discussion and conclusion
Muntiacus bohlini was established by Teilhard de Chardin (1940) for the specimens from Loc. 18, and he reassigned Muntiacus cf. M. sinensis from Xiashagou (Teilhard de Chardin and Piveteau, 1930) to this species. The present study on the material from Xinyaozi reconfirmed the presence of the taxon in Nihewan Formation. The extant muntjaks range only in China, southern and southeastern Asia (Nowak and Paradiso, 1983). The earliest muntjak, M. noringenensis, was found at Tuosu Nor in the eastern Qaidam of the Qinghai-Tibetan Plateau from the lower part of the Upper Youshashan Formation (9-11 Ma), the Late Miocene (Dong, 2007). It is a relatively large muntjak and it indicates probably the genus originated in the Qinghai-Tibetan Plateau. The second earliest muntjak, M. leilaoensis, was discovered at Leilao hominoid locality, Yuanmou in Yunnan Province (Dong et al., 2004) from lower part of Xiaohe Formation (8.2-7.2 Ma), the Late Miocene (Qi et al., 2006). It is much smaller than M. noringenensis and very close to the extant muntjaks. M. zhaotongensis was uncovered at Shuitangba hominoid locality, Zhaotong in Yunnan Province (Dong et al., 2014) from Shuitangba lignite beds (6.1-5.9 Ma), the terminal Miocene (Ji et al., 2013). It is roughly contemporary with M. lacustris and M. nanus from Yushe Basin in Shanxi Province (Teilhard de Chardin and Trassaert, 1937). The Early Pleistocene muntjaks are mainly those survived from the Neogene such as M. nanus and M. lacustris, and new members such as M. bohlini and M. fenhoensis (Chow, 1956) in northern China, as well as M. hengduanshanensis in southern China (Zong et al., 1996). The generic status of M. ?huangi from Yantouxi at Wushan (Dong and Chen, 2015) is not certain, otherwise it is another Early Pleistocene muntjak. M. lacustris ranged in northern, central and southern China, it is a widely distributed muntjak in the Early Pleistocene. On the contrary, M. bohlini ranged mostly in Palaeoarctic and transitional regions in China during the Early Pleistocene.Cervavitus ranges mostly in the Neogene (Vislobokova, 1990), with two species Cervavitus fenqii (Han, 1987) and C. ultimus (Lin et al., 1978) survived to the Early Pleistocene. The pliocervine from Xinyaozi is larger than other Pleistocene ones such as C. fenqii and C. ultimus but relatively close to the Late Miocene C. huadeensis (Tables 5-6). If the taxonomic status of Cervavitus cf. C. huadeensis of Xinyaozi specimens can be confirmed, its chronological range could extend from the Late Miocene to the Early Pleistocene, and its geographic distribution from Nei Mongol southwards to Shanxi.
Axis originated in the Late Miocene and ranges to the present (Bai et al., 2017). Axis shansius appeared first in the Early Pliocene in Yushe Basin of North China (Teilhard de Chardin and Trassaert, 1937; Li et al., 1984; Tedford et al., 2013) and dispersed to Xihoudu (Chia and Wang, 1978) and Lantian (Chi, 1975) in North China, Tuozidong in East China (Dong and Fang, 2004), Yuanmou in South China (Lin et al., 1978) and Luotuoshan in Northeast China during the Early Pleistocene (Bai et al., 2017). It is therefore a common member of the Pleistocene mammalian faunas in China.
Nipponicervus elegans was first uncovered in Xiashagou locality in Nihewan Basin and it was placed in subgenus Cervus (Rusa) (Teilhard de Chardin and Piveteau, 1930). It was reported from Yushe (Teilhard de Chardin and Trassaert, 1937), Yangguo of Lantian (Chi, 1975) and Anping of Liaoyang (Zhang et al., 1980). It was reassigned to Nipponicervus (Qiu et al., 2004). Its chronological range is limited from the Early to Middle Pleistocene and its geographic range in northern and northeastern China.
The taxonomic status of red-deer-wapiti, or elaphoid group is quite complicated. They were regarded as a subgenus Cervus (e.g. Nowak and Paradiso, 1983) or Elaphus (e.g. Wang and Wu, 1979). The group has many fossil and extant species or subspecies (e.g. Nowak and Paradiso, 1983; Sheng, 1992; Wang, 2003; Croitor and Obada, 2018; Croitor, in press). The first fossil red-deer-wapiti in China was described as Cervus canadensis fossilis, or Cervus canadensis mongoliae (Zdansky, 1925). It was revised later as Cervus (Elaphus) canadensis (Wang and Wu, 1979), and recently as Cervus canadensis mongoliae (Croitor, in press). The group in China was found mostly in northern regions such as Xujiayao (Hou et al., 2013), Salawusu (Qi, 1975; Hou et al., 2013), Shiyu (Jia et al., 1972), Upper Cave (Pei, 1940), Xiaogushan (Dong et al., 2010), Yanjiagang (Wei et al., 1986), etc. Morphology based taxonomy regarded European and Asian red deer (Cervus elaphus elaphus) and North American wapiti (Cervus elaphus canadensis) as two subspecies (e.g. Nowak and Paradiso, 1983; Sheng, 1992; Wang, 2003). It was supported by the fully fertile hybrids that can be produced under captive conditions (Moore and Littlejohn, 1989; Perez-Espona et al., 2013). But that based on molecular evidence place them in two separate species (Kuwayama and Ozawa, 2000; Pitra et al., 2004). And that based on both is very complicated (Groves and Grubb, 2011). The absence of bez tine is mostly seen in European red deer (Abbazzi, 1995; Baker et al., 2014). The Dazhuangke specimen is therefore included into the European red deer group as Cervus (Elaphus) elaphus. But the horizon of the Dazhuangke specimen is not yet certain. Because the most elaphoid specimens were uncovered from the Late Pleistocene deposits (Wang and Wu, 1979; Huang, 1991; Croitor, in press), some from the Middle Pleistocene (Abbazzi, 1995; Croitor, in press). On the other hand, Dazhuangke is about 2.8 km southwest of Taijiaping and 4.2 km southwest of Shuichongkou although it is in Xinyaozi Ravine. There is no other identified specimens can help to correlate the horizon. The described antler is the only identified specimen from Dazhuangke and it looks unlike those from the Early Pleistocene deposits at Shuichongkou and Taijiaping localities. It is therefore probably not contemporary with the Early Pleistocene deposits at Shuichongkou and Taijiaping localities, but not certain.
To conclude, total seven species of Cervidae were identified from the fossil specimens collected from Xinyaozi Ravine: Muntiacus bohlini, Cervavitus cf. C. huadeensis, Axis shansius, Nipponicervus elegans, Elaphurus davidianus predavidianus, Elaphurus bifurcatus and Cervus (Elaphus) elaphus. The previous six species with different body sizes and antler patterns are from the Early Pleistocene deposits at Taijiaping and Shuichongkou localities (with a chronological range of 2.6-1.7 Ma), and the last one from uncertain horizon at Dazhuangke. At least six species of cervids existed inside and around Sangganhe Basin during the Early Pleistocene. C. cf. C. huadeensis and A. shansius were survivors from the Late Neogene; M. bohlini, N. elegans, E. davidianus predavidianus and E. bifurcatus are new forms of the Early Pleistocene. If Dazhuangke horizon can be dated as those of Shuichongkou and Taijiaping, the appearance of elaphoid cervids could be traced back to the Early Pleistocene, and the evolution of elaphoid antler would start from the absence to the presence of bez tine. Cronin et al. (1994) indicated that eight glacial events can be inferred between 2.7 and 2.3 Ma and the sea level dropped about 50-60 m in the Sea of Japan. The presence of the Early Pleistocene Elaphurus and Nipponicervus in mainland China (Teilhard de Chardin and Piveteau, 1930; Chi, 1975; Chia and Wang, 1978; Qiu et al., 2004; Dong et al., 2019) and Japanese archipelago (Otsuka, 1968, 1972; Otsuka and Hasegawa, 1976; Otsuka and Shikama, 1978) added new evidence that the sea level was dropped down that these cervids could migrate from the mainland to the islands. The abundance of folivorous cervid specimens in the Xinyaozi Ravine area implies the existence of a certain scale of forested environment in Sangganhe Basin area during the Early Pleistocene.
Acknowledgements
The present work was supported by the Strategic Priority Research Program of Chinese Academy of Sciences, Grant No. XDB 26030304. The authors would like to acknowledge Professor Wei Qi for providing the material of the present study. They would also like to thank reviewers for comments and suggestions to improve the manuscript.参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 3]
DOIURL [本文引用: 1]

Dental and cranial appendage remains of Cervidae from the fossil site of Dorn-Dürkheim are studied in detail. The material mainly includes isolated teeth, isolated pedicles and antler pieces. Neither tooth rows nor complete appendages are recorded. Comparative morphology and statistics of morphometrics (principal component analysis and discriminant analysis) allow for the classification of small and large dentitions, small cranial appendages, two morphotypes of large pedicles and two morphotypes of large antlers. Possible combinations of the classified units document the sympatric occurrence of three species, namely, Procapreolus sp., Muntiacinae gen. and sp. indet., cf. Cervavitulus mimus, but the fragmentary condition of the material leads to ambiguity regarding their composition and, consequently, to a certain extent regarding the taxonomic identification. However, these remains indicate the contemporaneous occurrence of early Turolian members of the crown cervids Muntiacinae and Capreolinae and close a previous spatiotemporal gap in the European cervid record. In addition, their presence proves the progressive turnover from dichotomous-antlered muntiacines to early monopodial-antlered crown cervids from NE to SW Europe in the late Miocene. The taxonomical assignment challenges the recent hypothesis on the origin of crown Cervidae around the middle/late Miocene border since Dorn-Dürkheim cervids provide further evidence for the successive achievement of derived characters in cranial appendages of crown cervids (mediopostorbital position and backwards orientation of pedicles, coronet development, shaft development/elongation, beam development and increase in number of antler tines) in the lineage of crown cervids, which originated during the middle Miocene.]]>
[本文引用: 6]
DOIURL [本文引用: 1]
Oxford: Windgather Press.
[本文引用: 1]
[本文引用: 5]
[本文引用: 4]
Beijing: Cultural Relics Publishing House.
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 5]
[本文引用: 3]
[本文引用: 1]
DOIURL
[本文引用: 4]
DOIURL [本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 7]
DOIURL [本文引用: 4]
DOIURLPMID [本文引用: 1]
The family Cervidae includes 40 species of deer distributed throughout the northern hemisphere, as well as in South America and Southeast Asia. Here, we examine the phylogeny of this family by analyzing two mitochondrial protein-coding genes and two nuclear introns for 25 species of deer representing most of the taxonomic diversity of the family. Our results provide strong support for intergeneric relationships. To reconcile taxonomy and phylogeny, we propose a new classification where the family Cervidae is divided in two subfamilies and five tribes. The subfamily Cervinae is composed of two tribes: the tribe Cervini groups the genera Cervus, Axis, Dama, and Rucervus, with the Pere David's deer (Elaphurus davidianus) included in the genus Cervus, and the swamp deer (Cervus duvauceli) placed in the genus Rucervus; the tribe Muntiacini contains Muntiacus and Elaphodus. The subfamily Capreolinae consists of the tribes Capreolini (Capreolus and Hydropotes), Alceini (Alces), and Odocoileini (Rangifer + American genera). Deer endemic to the New World fall in two biogeographic lineages: the first one groups Odocoileus and Mazama americana and is distributed in North, Central, and South America, whereas the second one is composed of South American species only and includes Mazama gouazoubira. This implies that the genus Mazama is not a valid taxon. Molecular dating suggests that the family originated and radiated in central Asia during the Late Miocene, and that Odocoileini dispersed to North America during the Miocene/Pliocene boundary, and underwent an adaptive radiation in South America after their Pliocene dispersal across the Isthmus of Panama. Our phylogenetic inferences show that the evolution of secondary sexual characters (antlers, tusk-like upper canines, and body size) has been strongly influenced by changes in habitat and behaviour.
[本文引用: 1]
[本文引用: 6]
[本文引用: 4]
[本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURLPMID [本文引用: 1]
We determined the mitochondrial cytochrome b gene sequences (1140 bp) of one subspecies of the European red deer (Cervus elaphus in Europe), three subspecies of the wapiti (C. elaphus in Asia and North America), and six subspecies of the sika deer (C. nippon in Japan). Our phylogenetic analysis revealed the monophyly of the European red deer, that of the wapiti, and that of the sika deer. The wapiti, however, was shown to be more closely related to the sika deer than to the European red deer. This is in conflict with traditional morphological results, which suggest a close sister group relationship between the wapiti and the European red deer. The divergence time between the European red deer and the wapiti plus the sika deer was estimated to be approximately 0.80 Ma, and that between the wapiti and the sika deer was estimated to be 0.57 Ma. The sika deer was subdivided into two subspecies groups, and the wapiti was also found to consist of an Asian group and a North American group.
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURLPMID [本文引用: 1]
The red deer (Cervus elaphus) is an iconic species in Scotland and, due to its value as a game species, an important element of the Scottish rural economy. The native status of this species is sometimes questioned because of many recorded introductions of nonnative deer in the past that were an attempt to improve trophy size. In this study, we assessed the impact of past introductions on the genetic makeup of Scottish red deer by genotyping at 15 microsatellite loci a large number of samples (n = 1152), including mainland and island Scottish red deer and individuals from several putative external source populations used in introductions to improve trophy size. Population structure and introgression assessment analyses revealed that the impact of introductions was weak in Highland red deer populations but more prominent on the islands, especially on those where current red deer populations are mostly or entirely derived from introductions (Harris & Lewis, Arran, and Rum). Frequent imports of Central-Eastern European red deer into English deer parks were reflected in the higher genetic introgression values found in some of the individuals collected in parks.
DOIURL [本文引用: 1]
AbstractThe phylogenetic pattern and timing of the radiation of Old World deer was determined based on the complete mitochondrial cytochrome b gene from 33 Cervinae taxa. Using rooted and unrooted phylogenies derived from distinct theoretical approaches, strong support was achieved for monophyly of the Old World deer with muntjacs as sister group as well as for the divergence of at least three distinct genera: Rucervus, Dama, and Cervus. The latter clade comprises what have previously been regarded as the genera or subgenera Panolia, Rusa, Cervus, Sika, and probably Przewalskium. Our data also consistently confirmed paraphyly of nominate C. elaphus and did not support the monophyly of Axis. We used these molecular phylogenies to assess the homoplastic evolution of morphological, geographical, ecological, and selected behavioural character state differences within the Cervinae. Reliable fossil calibrations, large molecular data sets, and improved dating methods are shaping a molecular time scale for the evolutionary radiation of Old World deer that occurred at the Miocene/Pliocene transition and is largely compatible with existing palaeontological evidence. Using node ages estimated from sequence data, we estimated an average per-lineage diversification rate of 0.51 ± 0.1 species per million years (my) over roughly the last 6 mya.]]>
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 6]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 7]
[本文引用: 12]
[本文引用: 13]
[本文引用: 1]
[本文引用: 2]
[本文引用: 4]
[本文引用: 1]
Beijing: China Forestry Publishing House.
[本文引用: 3]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
Beijing: China Ocean Press.
[本文引用: 2]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}