 ,1, 王晓鸣,1,4, 李强1, 李录1, 王洪江2, 陈海峰3
,1, 王晓鸣,1,4, 李强1, 李录1, 王洪江2, 陈海峰3Late Miocene mammalian fauna of Halajin Hushu in Nei Mongol, China
QIU Zhu-Ding,1, WANG Xiao-Ming,1,4, LI Qiang1, LI Lu1, WANG Hong-Jiang2, CHEN Hai-Feng3通讯作者: qiuzhuding@ivpp.ac.cn;xwang@nhm.org
收稿日期:2020-07-10网络出版日期:2021-01-20
| 基金资助: |
Received:2020-07-10Online:2021-01-20

摘要
过去40年,在内蒙古中部地区的陆相新近纪化石地点中应用了筛选技术,采集到大量标本,这些发现迅速填补着晚新生代动物群演替和生物年代学认识的空白。在把孤立的化石组合按时代先后进行排序和建立动物群序列框架中,小哺乳动物的发现和研究发挥了至关重要的作用。报道了另一个产自哈拉津胡舒地点的晚中新世动物群,这个动物群再次展示了蒙古高原上令人“叹为观止”的化石种类和丰度。新地点位于该地区的最北部,并为玄武岩所覆盖,产出的标本计有7000余件,代表大、小哺乳动物达63种。虽然这一动物群混杂了少量下部层位的属种,但它不失为内蒙古中部地区一个较为真实地反映晚中新世小哺乳动物组成和生态环境的动物群。初步分析表明,哈拉津胡舒动物群属中国陆生哺乳动物年代的晚中新世早期的灞河期,可能比阿木乌苏动物群稍晚,但比沙拉动物群略早。
关键词:
Abstract
Discoveries during the past 40 years of continental Neogene fossil sites in central Nei Mongol, through the adoption of screen washing techniques, have rapidly filled previous gaps in our knowledge of late Cenozoic faunal evolution and biochronology. Small mammals have become the backbone of our efforts in serializing individual fossil assemblages and in establishing a detailed framework of faunal sequence. We present another Late Miocene fauna from a new Halajin Hushu locality that, once again, shows the “embarrassing riches” of the Mongolian Plateau. The new locality is the northern-most fossil site in this region and is capped by a sheet of basalt. The new fossil assemblage is represented by 62 taxa of small mammals and one large mammal based on more than 7000 specimens. Our collections thus captured a remarkably complete small mammal fauna, possibly including 6 reworked species. Despite this mixture of reworked taxa from strata of older age, the Halajin hushu Fauna closely represents a coherent Late Miocene small mammal assemblage and its environments. Our preliminary analysis suggests a Bahean Chinese Land Mammal age in the early Late Miocene for the Halajin Hushu Fauna, probably slightly younger than Amuwusu Fauna but older than Shala Fauna.
Keywords:
PDF (4248KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
邱铸鼎, 王晓鸣, 李强, 李录, 王洪江, 陈海峰. 内蒙古哈拉津胡舒晚中新世动物群. 古脊椎动物学报[J], 2021, 59(1): 19-26 DOI:10.19615/j.cnki.1000-3118.201126
QIU Zhu-Ding, WANG Xiao-Ming, LI Qiang, LI Lu, WANG Hong-Jiang, CHEN Hai-Feng.
1 Introduction
Neogene deposits with dense fossil records are widely distributed in the middle part of the Nei Mongol Autonomous Region (Inner Mongolia), China. Substantial advances in the study of biostratigraphy in this area have been made in the last 40 years. Since 1980s investigations and excavations have been carried out frequently at 15 localities with fossiliferous exposures, Aoerban, Balunhalagen, Bilutu, Gashunyinadege, Moergen in Sonid Zuoqi, Loc. 346, Tairum Nur, Amuwusu, Shala in Sonid Youqi, Ulan Hushuyin, Baogeda Ula, Huitenghe in Abag Qi, Bilike in Huade County, as well as the classic localities Tunggur, Ertemte and Gaotege (Fig. 1), and 18 faunas have been recovered from the sediments ranging from Early Miocene through Early Pliocene (Fahlbusch et al., 1983; Qiu et al., 1988; Qiu, 1996; Meng et al., 1996; Qiu and Storch, 2000; Li et al., 2003; Wang et al., 2009). Based on our studies of mammals, largely by abundant small mammals, a Neogene mammal succession in this area has been developed by faunal seriation. Growth of our knowledge in mammalian biostratigraphy resulted in a sequence of finely-spaced faunal units that span from Early Miocene to much of the Pliocene. At present, a preliminary framework of Neogene biostratigraphy and biochronology has been established for this area (Qiu and Wang, 1999; Qiu et al., 2006, 2013; Wang et al., 2009; Qiu and Li, 2016).The Halajin Hushu (Mongolian for “bold head”) locality (44o27′18.4″N, 114o33′06.8″E; IVPP IM1103 loc) is another Neogene site endowed with rich mammal fossils, which is a small basalt-covered hill situated about 41 km north of Baogeda Ula Sumu, and 130 km northwest of Xilinhot (Fig. 1). In 2011, while we were prospecting in Genghis Baogedu area, Mr. Chen Haifeng, director of the Abag Museum, Nei Mongol guided us to the locality. Short surveys in the site immediately demonstrated that the grayish white sandy mudstone near the top beneath the basalt is rather fossiliferous, especially suitable for small mammal screening. In that field season, we excavated and screen-washed about 2.5 tons of the sediments, with the aim of collecting fossil mammals. As a result, the sample yielded more than 7000 specimens representing at least 62 taxa, dominated by isolated teeth of small mammals. Besides the small mammals, there are a few remains of large mammals and fossil fishes, amphibians and reptiles. It is clear that Halajin Hushu is one of the most important Neogene localities producing abundant and diverse assemblage known from central Nei Mongol. In the present paper, we briefly report the preliminary results of our field campaign and discuss related items of the vertebrate microfauna. Detailed descriptions of the material will be presented in the future.
2 Geologic settings
The Central Asian Orogenic Belt (CAOB), which encompasses the study area, is an accreted complex of Paleozoic and early Mesozoic age wedged between the Siberia Craton and North China Craton (e.g., Kr?ner et al., 2014; Windley et al., 2007). The late Cenozoic uplift of the Mongolian Plateau, which partly coincides with the CAOB, is associated with mantle upwelling, lithospheric extension and thinning, and alkalic magmatism (Chen et al., 2015; He and Wu, 2020; He et al., 2016; Windley and Allen, 1993). Continental intraplate basalts are widespread in central Nei Mongol and named Abag-Dalai Nor Volcanic Field (Chen et al., 2013; Gong et al., 2016; Ho et al., 2008; Zhang and Guo, 2016), which extends north of the Chinese border in the Dariganga Plateau of Mongolia (Deng and Macdougall, 1992; Devyatkin and Smelov, 1980; Whitford-Stark, 1987). Within the Abag Volcanic Field, the ages of the basalts are as old as 14.57±0.36 Ma in the Middle Miocene at Baiyinxile (sample B47 of Luo and Chen, 1990), through Late Miocene (5-7 Ma) in the Baogeda Ula area plus areas south of Abag (samples 54, 55 of Ho et al., 2008; Shao et al., 2008) and Late Pliocene (2.80±0.4 Ma) at Haernudenghua near Abag (Shao et al., 2008), to as young as Pleistocene (less than 1 Ma) further south (Liu et al., 1992; Luo and Chen, 1990; Zhang and Guo, 2016).Except the above volcanism, a relatively stable tectonic setting in the study region prevailed with terrestrial sedimentations being typically thin-bedded with low rate of sedimentation. All deposits are flat-lying, overbank and flood plain sediments, and show no sign of tectonic deformation. As elsewhere in the Mongolian Plateau, vertebrate fossils are enriched in the low-rate sedimentation environment. Most of the late Cenozoic sediments in the study region are closely associated with capping basalts of different ages, which protect underlying sediments from erosions and form steep exposures along their edges, free of vegetation coverage (Fig. 2). Within the Abag Volcanic Field, there are at least three discrete exposures containing faunas of different ages. The oldest is the Bahean Halajin Hushu fauna in the early Late Miocene (this study). The age for IM0708 loc is less certain because it includes mostly reworked specimens of Paleogene taxa (Wang and Li, 2011), but presumably the sedimentation age is somewhere in the Late Miocene. The Baogeda Ula Formation, as represented by IM0702 in Fig. 2, has produced a Baodean fauna (Deng et al., 2016; Qiu and Wang, 1999; Qiu et al., 2006; Tseng and Wang, 2007; Wang et al., 2012). Paleomagnetic dating of the Baogeda Ula Formation (Sun et al., 2018), constrained by both radioisotopic dates and fossil mammals, yielded an age range for the strata that suggest immediate burial of the sediments by basalt flows, i.e., no significant hiatus is observable between capping basalts and underlying sediments. In a section east of the Baogeda Ula Sumu, two layers of basalts are in superpositional relationship with sediments below and in between, but, unfortunately, repeated attempts at dating the weathered basalts have failed to yield a consistent result.
Fig. 1
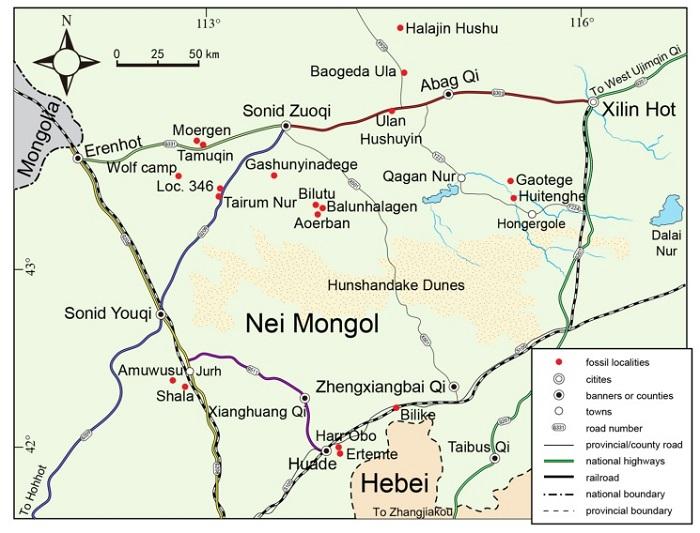 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 1Geographic location of the fossil site Halajin Hushu in Nei Mongol
The Halajin Hushu locality is the northern-most exposure with abundant fossils capped by a thin, weathered basalt. The basalt is weathered to a narrow, north-south oriented band, and sediments, about 200 m in lateral extent, are mostly exposed toward the northern end of the basalt band (inset in Fig. 2). We measured 36 m for a total thickness of Halajin Hushu bed, although the lower 18 m are covered by soil and vegetation, i.e., the observable section is only about 18 m in thickness (Fig. 3). Sediments are predominantly siltstones and mudstones with a few layers of carbonate nodules and one finely laminated lacustrine bed containing plant fragments (at 28 m). The fine-grained sediments are mostly reddish brown (mudstones) and light yellowish grey (nodular zones) in color, the latter giving a light gray tone where carbonate nodules (thin-bedded ones likely of in situ precipitation but thicker beds possibly secondarily enriched) are abundant (Fig. 3B). We interpret the sediments as floodplain deposits with several horizons of paleosols containing pedogenic mottling and carbonate nodules, plus one thin layer of ponded, indurated bed at 28 m. A semiarid climate is indicated by the precipitation of soil carbonates. The capping basalt is thin, about 1 m or less, and is extensively weathered. The basalt sheet dips southward, presumably following an ancient topographic surface of similar dip (assuming no post-depositional disturbance). As is the case in IM0708 loc (Wang and Li, 2011) and Aoerban (Wang et al., 2009), the Halajin Hushu bed also contains reworked taxa, presumably by transportation from older strata nearby (see Mammal assemblage below).
Fig. 2
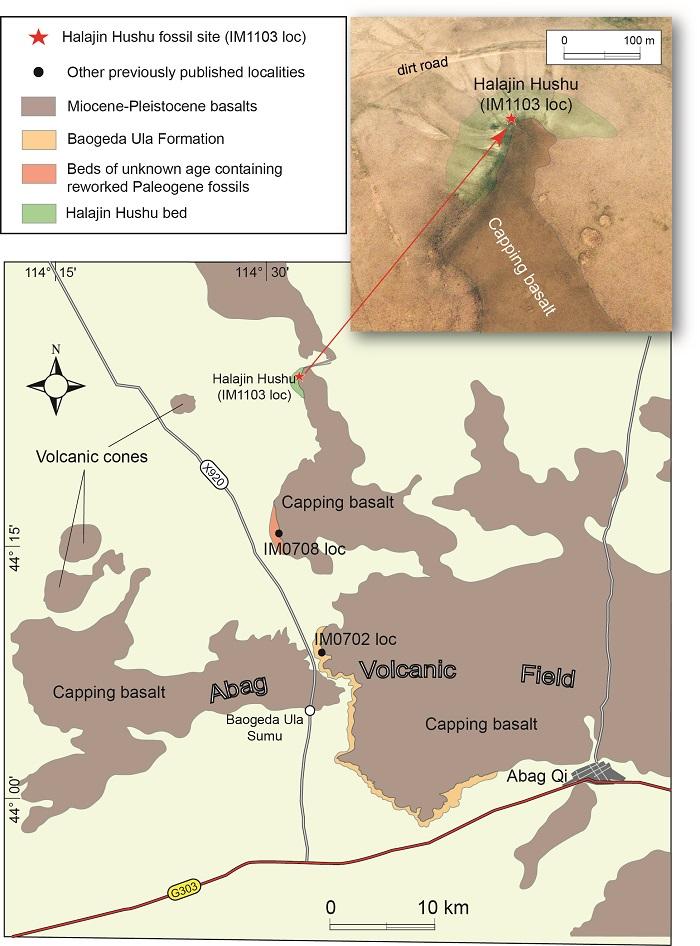 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 2Map of Abag Volcanic Field showing contact relationships of Neogene sediments and capping basalts The Halajin Hushu locality is the northern-most fossil site associated with basalts. Only one fossil site is shown for each formations or beds that contain a discrete fauna. Cenozoic exposures and basalts are drawn from satellite images in Google Earth Pro (Version 7.3.3.7699) (2020), as is the inset (upper right) with semitransparent color enhancements of basal and Cenozoic exposures
3 Mammal assemblage
The Halajin Hushu assemblage includes at least 63 forms in 17 families and 5 orders of mammals. Table 1 is a nominal list of the mammals recognized in the collection and numbers of the specimens yielded from the locality.Table 1
Table 1Taxa and specimen number (N) of the Halajin Hushu assemblage
| Taxa | N | Taxa | N | |
|---|---|---|---|---|
| Eulipotyphla | Zapodidae | |||
| Erinaceidae | Parasminthus tangingoli* | 11 | ||
| Mioechinus? gobiensis | 1142 | P. parvulus* | 5 | |
| Erinaceus sp. | 2 | Eozapus major | 13 | |
| Talpidae | Sinozapus parvus | 264 | ||
| Yanshuella cf. Y. primaeva | 117 | Sicista sp. | 257 | |
| Quyania cf. Q. chowi | 52 | Heterosminthus orientalis | 931 | |
| Desmanella storchi | 155 | Lophocricetus xianensis | 1119 | |
| Talpinae indet. | 41 | Zapodidae indet. 1 | 17 | |
| Soricidae | Zapodidae indet. 2 | 7 | ||
| Mongolosorex sp. | 2 | Dipodidae | ||
| Sorex cf. S. minutoides | 12 | Protalactaga lantianensis | 4 | |
| Sorex ertemteensis | 128 | Paralactaga shalaensis | 4 | |
| Petenyia cf. P. katrinae | 28 | Cricetidae | ||
| Cokia kowalskae | 19 | Alloeumyarion sp.* | 3 | |
| Paranourosorex cf. P. inexspectatus | 171 | Democricetodon lindsayi | 8 | |
| Paenelimnoecus obtusus | 2 | Plesiodipus leei* | 10 | |
| Soricidae indet. | 2 | P. progressus | 7 | |
| Chiroptera | P. robustus | 4 | ||
| Chiroptera indet. | 1 | Gobicricetodon robustus | 5 | |
| Rodentia | Gobicricetodon aff. G. flynni | 3 | ||
| Ctenodactylidae | Khanomys baii | 72 | ||
| Tataromys sp.* | 2 | Colloides xiaomingi | 8 | |
| Aplodontidae | Colloides sp. nov. | 35 | ||
| Ansomys borealis | 9 | Kowalskia shalaensis | 233 | |
| A. lophodens | 21 | Microtoscoptes sp. | 1 | |
| Pseudaplodon amuwusuensis | 1 | Ischymomys sp. | 2 | |
| Sciuridae | Epimeriones sp. nov. | 116 | ||
| Tamias ertemtensis | 339 | Baranomyinae indet. | 98 | |
| Spermophilinus sp. | 13 | Spalacidae | ||
| Atlantoxerus sp. | 5 | Pararhizomys qinensis | 3 | |
| Prospermophilus orientalis | 76 | Gerbillidae | ||
| Hylopetes sp. | 3 | Myocricetodon sp.* | 3 | |
| Gliridae | Myospalacidae | |||
| Orientiglis wuae | 227 | Prosiphneus qiui | 199 | |
| Eomyidae | Lagomorpha | |||
| Leptodontomys gansus | 405 | Ochotonidae | ||
| L. lii | 52 | Desmatolagus sp. | 23 | |
| Keramidomys fahlbuschi | 41 | Ochotona sp. | 364 | |
| K. magnus | 254 | Artiodactyla | ||
| Castoridae | Moschidae | |||
| Monosaulax tungurensis | 10 | Moschidae indet. | 15 | |
| Castor sp. | 1 |
新窗口打开|下载CSV
Eulipotyphla Three families of the order, Erinaceidae, Talpidae, and Soricidae are represented in the sample. It is one of the richest insectivore assemblages in the Neogene of central Nei Mongol, both in taxonomic diversity and abundance of materials.
Erinaceidae are the most abundant insectivores in the collection despite their low diversity. Eleven hundred and forty-two isolated teeth, showing distinct variation in size, are identical to the hedgehogs reported as Mioechinus? gobiensis ranging from Aoerban of Early Miocene to Shala of early Late Miocene. An M2 and an M3 are referred to an indeterminate species of Erinaceus. The two molars are evidently larger than those of M.? gobiensis with a short crest connecting the strong protocone and the conical hypocone on M2, which are comparable to the fragments assigned to an indeterminate Erinaceinae from Moergen (Qiu, 1996). Further work may prove that records of Erinaceus should be earlier than expected.
The Talpidae include four forms belonging to two subfamilies, Talpinae and Desmaninae. One hundred and seventeen teeth are referred to Yanshuella, and 52 teeth to Quyania of Talpinae from Ertemte and Harr Obo of Huade (Storch and Qiu, 1983). In morphology, the Halajin Hushu Yanshuella is similar to Y. primaeva from Huade, but differs in having stronger cusps and crests, and the oblique crista extending more lingually on m1 (Fig. 4); the Quyania is similar to Q. chowi, but different in having more lingual extension of the oblique crista on m1-2. The development of cusps and crests, and the direction of oblique crista deserve further attention because of a lack knowledge of these characters. One hundred fifty-five isolated teeth can be identical as Desmaninae and referred to Desmanella storchi known from Moergen (Qiu, 1996). Forty-one teeth represent another talpine in this assemblage. On the basis of the relatively small size, presence of prominent conules on P4 and M1-2, and a lingual furrow between the protocone and protoconule on upper molars, and the oblique crista extending towards or connecting with the metastylid on lower molars, these teeth probably represent an undescribed talpine, but no generic assignment can be made at this time.
Shrews are the most diverse insectivores in the assemblage, and eight forms belonging to at least six genera of Soricidae are recognized. A large M1/2 with quadrate occlusal outline and wide hypoconal flange, and an m3 with low buccal reentrant valley and pronounced cingulum demonstrate some similarities to the corresponding teeth of Dinosorex from Europe (Engesser, 1972), and to the m3 of Mongolosorex qiui from Moergen, which was named based on only lower dentition (Qiu, 1996). The two teeth are assigned to Heterosoricinae and referred to an indeterminate species of Mongolosorex due to slightly smaller size and scarce material. Soricinae, including five genera, are more common than the Heterosoricinae in the sample. Sorex are identified by 140 specimens based on the pigmentary teeth with distinct posterior emargination on P4 and M1, the moderate development of entocristid (entoconid crests) on lower molars and the less reduced talonid on m3. Two species can be easily distinguished by size, the smaller one (n=12) is close to S. minutoides from Ertemte, except for the less cuspules on lower incisor; the larger one (n=128) to S. ertemteensis. Twenty eight specimens, including fragmentary maxilla and mandibles, are referred to Petenyia based on the P4 with anteriorly projecting paracone, strong longitudinal posterior crest of protocone, rectangular molars with narrow cingulum around the bases on M1-2, high entoconid closely positioned to metaconid on m1-2, and the m3 with rather reduced talonid. They are similar to those of P. katrinae from Bilike, but differ in smaller size and having wider postentoconid valley (Qiu and Storch, 2000). Nineteen specimens with the parastyle of P4 not particularly protruding anteriorly, rectangular M1-2 with very slight emargination, strong protocone and distinct metaloph, quadrate m1-2 with broad and inflated labial cingulid, prominent entocristid, and protoconid rather closed to metaconid indicate similarity to Cokia kowalskae from Ertemte and Harr Obo. One hundred and seventy-one specimens are identical as Paranourosorex cf. P. inexspectatus. They resemble those of P. inexspectatus from Ertemte, but having more distinct posterior emargination on M1, and four roots on M2, characters that may be indicative of more primitive status. Paenelimnoecus obtusus is represented by two mandibular fragments that possess the following characters: smaller teeth, lower molars lacking entocristid, protoconid closely situated to metaconid, and obvious reduction of talonid on m3. A mandibular fragment with m1-2 and an isolated m2 are referred to an indeterminate Soricinae. Lower molars in the two specimens are distinguished from others by the lack of entoconid and entocristid, which exhibit some Allosorex-like characters, but it cannot be identified below the family level.
Chiroptera Only an M2 indicates the presence of bats; at present it is only identified to an indeterminate Chiroptera.
Rodentia Rodents are the most diverse and abundant small mammals in the sample. Thirty-three genera belonging to 12 families (Ctenodactylidae, Aplodontidae, Sciuridae, Gliridae, Eomyidae, Castoridae, Zapodidae, Dipodidae, Cricetidae, Spalacidae, Gerbillidae and Myospalacidae) are recognized.
A P4 and an M3 represent the existence of Ctenodactylidae in the collection. In size and in having a complete protoloph and metaloph on P4, a V-shaped protocone, complete metaloph joining the posterocone, straight posterior arm of protocone (also known as entoloph), and shallow sinus on M3, the two specimens are similar to the corresponding teeth of Tataromys parvus commonly known from the Oligocene in Central Asia. They are assigned to an indeterminate species of Tataromys because of the scarce material, and are considered to be a mixed one, presumably due to the reworking of deposits of Oligocene or Early Miocene age.
Thirty isolated teeth are referred to the genus Ansomys of Aplodontidae on the basis of the brachydont cheek teeth with high cusps and crests, distinct mesostyle forming a handle-shaped flexure of ectoloph and single metaconule on upper molars, and the anteroposteriorly compressed main cusps and crest-like metaconid on lower molars. Based on the poorly developed or absent mesoconid on lower cheek teeth, most specimens can be identified as A. lophodens, a more derived species in the genus mainly known from Amuwusu, Balunhalagen and Huitenghe of early Late Miocene, and a few as A. borealis, a relatively primitive species known from the Early and Middle Miocene (Qiu and Li, 2016). An m1/2 represents a large-sized mountain beaver in the assemblage, which is comparable in size and morphology to that of Pseudaplodon amuwusuensis from Amuwusu.
Five taxa of squirrels, including sciurids of different adaptive types, are recognized by size and morphology. Three hundred and thirty-nine isolated cheek teeth are referred to Tamias ertemtensis, the most frequent chipmunks known from the Early Miocene to Early Pliocene in central Nei Mongol. Thirteen specimens can be assigned to an indeterminate species of Spermophilinus, another larger chipmunk with heavier built, less convergence of protoloph and metaloph towards the protocone on P4-M2. Spermophilinus are never very frequent in the fossil record of central Nei Mongol, with their occurrence of S. mongolicus at Balunhalagen, Amuwusu and Bilutu of Late Miocene age. The added material seems to show somewhat heavier built in dentition than that of S. mongolicus. Five cheek teeth can be identified as Atlantoxerus, a genus of ground squirrels widely occurring in the early and middle Neogene of Palearctic Region. The taxon assigned to Atlantoxerus sp. in the sample can be distinguished from A. exilis and A. major, but similar to A. orientalis known from Moergen and Balunhalagen in smaller size, and having metaconule close or connected with the posteroloph (Qiu and Li, 2016). One maxillary fragment with M2-3 and 75 teeth are identical to Prospermophilus orientalis in size and in having prominent metaconule, protoloph and metaloph converging towards protocone on P4-M2, and ectolophid curving posterolingually on lower molars. P. orientalis is a relatively small ground squirrel known from several localities of Late Miocene and Early Pliocene in central Nei Mongol. Three molars are referred to an indeterminate species of Hylopetes. The only flying squirrel in the collection is characterized by its distinct entoconid, angular entoconid corner, anterolingually extending buccal valley, and crenulated enamel on surface. It resembles H. bellus from Balunhalagen in size and morphology.
Fig. 3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 3Stratigraphic section of Halajin Hushu
A. photograph of Halajin Hushu section (photo looking southward and taken on August 4, 2015 by X. Wang); B. closeup of exposures at fossiliferous horizon and a nodular zone immediately above (photo looking eastward and taken on June 18, 2018 by X. Wang); C. measured section of Halajin Hushu strata
Two hundred and twenty-seven isolated cheek teeth are identified as Gliridae. They can be all referred to Orientiglis wuae by small-sized cheek teeth with concave occlusal surface, relatively equal main ridges and accessory ridges in thickness, less frequent anterotropes and posterotropes, longer precentroloph than postcentroloph on M1-2, lingually free anteroloph in majority of M1, and presence of anterotropids and posterotropids on m1-2 (Fig. 4). The only species of dormouse in the sample is also known from Middle Miocene of Gansu, as well as from Early Miocene through Late Miocene in central Nei Mongol (Qiu, 2001b; Qiu and Li, 2016).
More than seven hundred specimens are referred to bunodont Leptodontomys and lophodont Keramidomys of Eomyidae. The two genera are common in several localities of early Neogene in central Nei Mongol, and as in European Miocene microfaunas they usually occur in association. Leptodontomys is more frequent and can be assigned to a smaller species, L. gansus (n=405), with relatively weaker cusps and crests (Fig. 4) and a larger species, L. lii (n=52), with stronger cusps and crests. Keramidomys also can be distinguished by size and morphology. The smaller form is identical with K. fahlbuschi (n=41) and the larger with K. magnus (n=254).
Eleven isolated teeth represent two genera and two species of Castoridae, of which 10 are assigned to Monosaulax tungurensis and one to an indeterminate species of Castor. M. tungurensis frequently occurs in the Middle Miocene and the early Late Miocene in the Tunggur tableland. The added material is identical with those known from Moergen and Amuwusu in size and morphology, but with poor development of cement fill in the striae (ids). A P4 corresponds morphologically to the diagnosis of Castor, based on its large size, four flexi filled with thick cement, closure of flexi in distinctly different stages of wear. The premolar is close to that of Hystricops mengensis in size and structure, but differs in the hypostria and mesostria more extending to the bases of crown. In comparison with that of C. anderssoni from Ertemte, Bilike and Gaotege, the two striae show less extension, may be indicative of a more primitive status.
Zapodidae are the most abundant rodent family in the collection, and six genera and nine species are recognized. Sixteen specimens are referred to the genus Parasminthus, two species of which can be readily distinguished by size and morphology. The larger species (n=11) is identical with P. tangingoli. A mandible fragment with m1 and m2, and 4 isolated teeth can be comparable to that of P. parvulus, which show some similarities to Plesiosminthus vegrandis in size, but differ in being less lophodont. Both P. tangingoli and P. parvulus are known from Oligocene localities such as Taben-buluk, Gansu and Ulantatal, western Nei Mongol (Bohlin, 1946; Huang, 1992). It is noteworthy that Parasminthus is considered to survive in Central Asia into Early Miocene before becoming extinct, and no remain of the genus has been found in the Neogene of central Nei Mongol until now. Therefore, we consider the occurrence of Parasminthus in Halajin Hushu as another case of reworked fossils from older strata. Thirteen molars can be referred to Eozapus major based on their relatively simple molar structure and developed lophs, lack of mesocone(id), presence of strong mesoloph(id) and posteroloph(id)s, single protoloph on upper molars and metalophid on lower molars. They are similar to those of E. similis from Ertemte and Harr Obo, but larger in size with less strong lophs. The M2s in the assemblage correspond in all features with the type specimen of the species from Balunhalagen (Fig. 4). These materials afford a good understanding of the species E. major, which was named based on limited specimens (Qiu and Li, 2016). A mandible fragment with m1, and 263 isolated teeth are referred to Sinozapus parvus. In size and morphology, they are close to those from Balunhalagen, Amuwusu, Shala and Huitenghe, but smaller than those of S. volkeri from Bilike. Two hundred and fifty-seven molars are referred to Sicista sp. The indeterminate species of birch mice is larger than S. prima from Aoerban and Gashunyinadege, but smaller than S. bilikeensis from Bilike. It appears to represent a population morphologically intermediate between the Early Miocene and the Pliocene taxa with moderately developed accessory spurs on molars, moderate amount of protoloph joining protocone or anterocone on M1, and transitional characters in the direction of hypolophid on m1 and the development of secondary ridges in posterosinusid on m1-2. Heterosminthus orientalis and Lophocricetus xianensis are recognized in the sample, by 931 and 1119 specimens, respectively. These two frequently co-occurring taxa are similar in general dental morphology. Those molars possessing the following features are here assigned to H. orientalis: smaller dimension, relatively long and narrow occlusal outline, weakly developed cusps, protostyle lacking or present as a posterolingual rib of protocone and long mesoloph on M1, frequent double anteroloph on M2, long-triangular M3, metaconid usually connecting to mesoconid on m1, double rooted m2 with poorly developed ectocingulid and distinct posterior arm of protoconid, otherwise to L. xianensis (Fig. 4). Twenty-four molars are referred to two indeterminate genus and species of Zapodidae by size and morphology. The smaller one, represented by 17 specimens, possesses features highly similar to Parasminthus: buno-lophodont molars with well-developed mesoloph(id)s, asymmetric sinus(id)s, single protoloph in M1, and single metolophid in m2-3, but can be easily distinguished from all known species of the genus by its small size. The larger one shares some characters with Eozapus major, but larger in size and relatively shorter and wider in occlusal outline. Further study may show these specimens to represent new taxa of Zapodidae.
Fig. 4
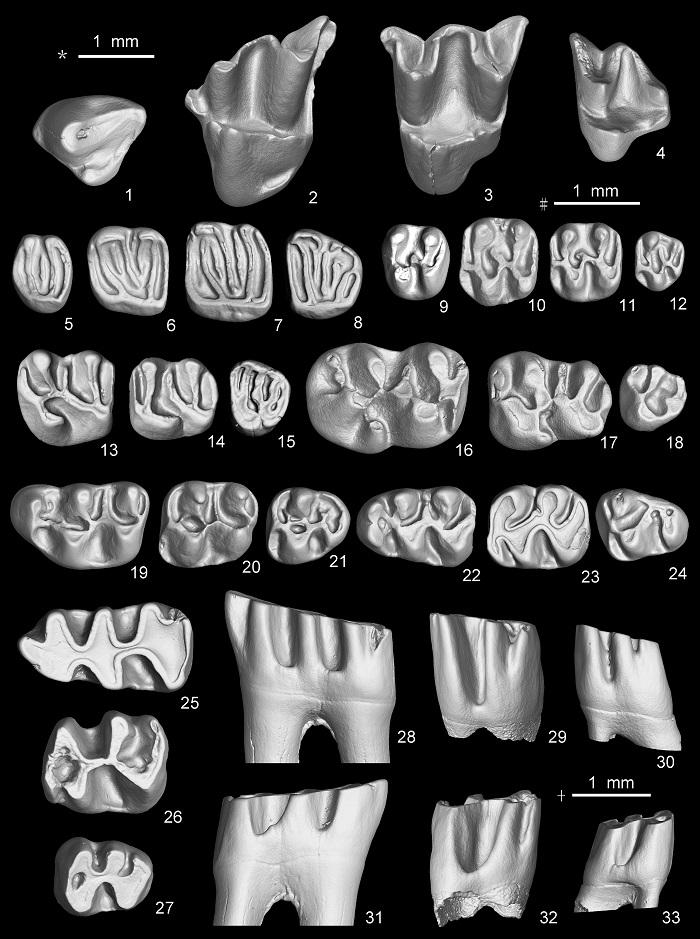 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 4Selected specimens of small mammals from Halajin Hushu
Yanshuella cf. Y. primaeva (1-4), left P4-M3; Orientiglis wuae (5-8), left P4-M3;
Leptodontomys gansus (9-12), left P4-M3; Eozapus major (13-15), right M1-M2 (reversed) and left M3;
Lophocricetus xianensis (16-18), left M1-M3; Kowalskia shalaensis (19-21), left M1-M3;
Colloides sp. nov. (22-24), left m1-m3; Epimeriones sp. nov. (25-33), left m1-m3;
1-27. occlusal view, 28-30. lingual view, 31-33. buccal view; scales: * for 1-4, # for 5-24, + for 25-33
Remains of Dipodidae are rather scarce, and only two genera and species, representing Allactaginae, are represented in the assemblage. Four cheek teeth are identified as Protalactaga lantianensis known from the early Late Miocene of Bahe in Shaanxi and Balunhalagen in Nei Mongol, based on its smaller size, uncompressed cusps anteroposteriorly, protoloph and metaloph joining respectively entoloph and posteroloph on M2, and hypolophid connecting with the middle of mesolophid on m2. Three P4s and one M1 are referred to Paralactaga shalaensis from Shala, representing another jerboa with large size, high crown, strong lophs and protoloph connected to the middle of mesoloph on M1.
Cricetidae are the most diverse rodent family in the sample, and at least 10 genera and 15 species including cricetids with buno-lophodont type and lophodont (microtoid) type cheek-teeth are represented in the assemblage. Three M2s are referred to an indeterminate species of Alloeumyarion, which are similar in morphology to the corresponding tooth of A. sihongensis from the Early Miocene Xiacaowan Formation of Jiangsu in having low crown and square occlusal outline, more prominent cusps than crests, slightly anterolingually-directed protoloph and transverse metaloph, short mesoloph, and straight entoloph connected with protoloph and hypocone, but smaller in size (Qiu, 2010). Eight molars are identical with Democricetodon lindsayi, a commonly known taxon from Early Miocene to early Late Miocene in Nei Mongol, in M1 and m1 having wide and simple anterocone(id) and medium or long mesoloph(id), M1-2 having metaloph connected with posteroloph, m1-2 lacking metalophid II, hypolophid joining with ectolophid. They fall within the range exhibited by the early Late Miocene populations from Amuwusu, Balunhalagen and Huitenghe as to size and morphology. Twenty-one molars are referred to the genus Plesiodipus based on the absence of mesoloph(id), cusps and crests aligned in three rows, and the presence of pronounced “medial diagonal ridge”. Three taxa of the genus can be readily distinguished by size and morphology. The smaller species represented by 10 molars is identified as P. leei, which are frequent in the Middle Miocene of central Nei Mongol, Xinjiang and Gansu (Qiu, 2001a; Wu et al., 2009; Qiu and Li, 2016). All 10 teeth in the collection distinctly show signs of water erosions and we suspect that they have been reworked. The medium-sized one (n=7) is similar to P. progressus from Tamuqin, Balunhalagen and Bilutu (Qiu, 1996; Qiu and Li, 2016), in having more elongated cheek teeth with more compressed cusps, more reduced protosinus of M1, narrower and shallower buccal valleys of lower molars. In respect of reduction of valleys, it is closer to those populations from Late Miocene than to the Middle Miocene. The largest one (n=4) is referred to P. robustus based on the large size, heavily built and lophodont molars with high crown and strong ridges, M1 with completely reduced protosinus and roughly “?”-like ridges, m1 with single and thick anterolophulid. P. robustus is also known from the Late Miocene localities, Balunhalagen, Huitenghe and Bilutu. Eight molars are assigned to Gobicricetodon by their large size and buno-lophodonty, M1 and M2 with oppositely arranged main cusps, single protoloph and metaloph, and slightly anteriorly-directed sinus, M1 and m1 with simple anterocone(id) and short mesoloph(id). Two species of the genus are recognized, with the larger species G. robustus by five molars, and the smaller one Gobicricetodon aff. G. flynni by three m1s with weakly or undeveloped metalophid and mesolophid. Both are also known from Amuwusu, Balunhalagen and Bilutu. Seventy-two specimens, including two maxillary fragments with M1 and M2, represent the genus Khanomys. In morphology, they are roughly similar to K. baii from Amuwusu and Balunhalagen, and K. cheni from Shala, Huitenghe and Bilutu, but more similar to the former in less frequent presence of a circular lake-like posterosinus on M1-2, in having curved or smooth anterior wall on M1 and absence of protosinus on M2. Forty-three isolated molars are characterized by their relatively hypsodont cheek teeth with deep and narrow valleys, short or absent mesoloph(id)s, and undivided anterocone on M1, single and posteriorly-directed protolophs and metalophs on M1-2, slightly split anteroconid, single anterolophulid, metalophid and hypolophid on m1, lack of lingual anterolophid on m2-3, which correspond to the diagnosis of Colloides given by Qiu and Li in 2016. Eight molars can be referred to C. xiaomingi, the larger species from Amuwusu, Balunhalagen, Huitenghe and Shala by its more distinctly split anteroconid on m1, and the other thirty-five molars to a smaller form, probably representing a new species of Colloides (Fig. 4). Kowalskia is represented by 233 specimens, including 4 maxillary and mandibular fragments, on the basis of their smaller-sized cheek teeth with relatively long mesoloph(id)s, M1 having narrow and weakly divided anterocone, protoloph I and metaloph II present in the majority of the three-rooted M1-2, m1 having nearly undivided anteroconid and occasionally the ectomesolophid. The dental features in the specimens completely correspond to the diagnosis of K. shalaensis from Shala, except for the presence of a single anterolophulid in most of m1s (Fig. 4). Microtoscoptes is a genus possessing hypsodont molars with prismatic structure, which occurs in the Late Miocene of Eurasia. A lower m3 identified as an indeterminate species of the genus is comparable in size and in reduction of the posterior lobe to that of M. fahlbuschi from Balunhalagen, Bilutu and Baogeda Ula or the indeterminate species from Shala. On the basis of relatively large size, hypsodont prismatic and cementless, undifferentiated enamel walls, slightly alternating synclines/anticlines with tendency to form a rhombic dentine area in the middle of the occlusal surface, a mandibular fragment with m1-2, and an m3 are identified as an indeterminate species of Ischymomys. This genus is a rare microtoid rodent distributed in the Late Miocene of Kazakhstan and Ukraine, as well as in Nei Mongol (Topachevsky et al., 1978; Qiu and Li, 2016). Two hundred and fourteen specimens represent two taxa of microtoid cricetids with semihypsodont molars and trilophodont M1. The first form, based on 116 molars, are referred to a new species of Epimeriones, representing the first record of the genus in Asia (Fig. 4). The taxon is similar to the type species E. austriacus from Eichkogel, Austria (MN 11) in possessing two lobes on M2-3 and m2-3, additional lingual distal sincline on m1, but differs in having narrower longitudinal enamel connected dentine areas on molars and more distinct additional lingual sinclines on lower molars, characters that may be indicative of a more primitive status. Epimeriones was originally treated as a gerbil and later as a “Muridae” incertae sedis (Daxner-H?ck, 1972; Daxner-H?ck and H?ck, 2015). The possible assignment to Gerbillinae or “Muridae” incertae sedis is still under discussion (Fejfar, 1999; McKenna and Bell, 1997). It is likely that Epimeriones shows relatively hypsodont prismatic molars lacking cement in synclines, with flat chewing surface, dentine areas connected by longitudinal enamel or very narrow dentine bands, which demonstrates some Microtoscoptes-like characters. Further study may lead to the recognition of affinities of the microtoid cricetids to Microtoscoptinae. The second form, based on 98 molars, is assigned to Baranomyinae gen. et sp. indet. The taxon is somewhat larger than the Epimeriones, slightly lower crowned molars with two lingual and buccal synclines on M1, and three lingual synclines and two buccal synclines on m1. Morphologically, it is more or less similar to Microtodon from Asia, and “Celadensis” from Europe, but much larger in size, and in having distinctly narrower dentine bands connected dentine areas, and shallower posterosinusids on m1-2. All of these specimens probably represent a new species of Microtodon or new genus of microtoid cricetids, but generic assignment must await a further study.
Spalacidae are represented by only three isolated molars, belonging to the genus Pararhizomys. The taxon is characterized by its small size, lower crown, uprightly W-shaped occlusal surface of M1 and two-lobed M3 and m3. It is smaller than P. hipparionum from Baogeda Ula (Li, 2010), and all the species of Pararhizomys and Pseudorhizomys from Linxia Basin, Gansu (Wang and Qiu, 2018), but closely identical to P. qinensis from the Late Miocene Bahe Formation, in Lantian, Shaanxi (Zhang et al., 2005).
Three M1s showing wear by water are referred to an indeterminate species of Myocricetodon, which is more similar to M. plebius from the Quantougou Formation of Middle Miocene in Gansu than to M. lantianensis from the Bahe Formation of early Late Miocene in having single anterocone, less closer cusp-pairs and presence of an initial “new” longitudinal crest (Qiu, 2001b). These specimens represent the first record of Myocricetodon in central Nei Mongol.
Myospalacidae are recognized based on 199 specimens. These molars are similar to those of Khanomys, but larger in size, having very short or remnant mesoloph(id)s in early wear, and less developed protosinusid on m2-3. They are identical in size and morphology to Prosiphneus qiu, a primitive zokor commonly known from Amuwusu and Balunhalagen.
Lagomorpha Lagomorphs are abundant small mammals in this sample, but only two genera, Desmatolagus and Ochotona of the family Ochotonidae can be recognized. Desmatolagus are mainly distributed in the Oligocene and Early Miocene of Central Asia. Twenty-three rooted cheek teeth may be represented the last record of the genus in central Nei Mongol, characterized by the unilateral upper P3-M3, P3 having anteroloph crossing about two-thirds width of the tooth and crescentic valley connecting to anteroexternal wall, P4 having developed anterobuccal and middle buccal elements, and P4-M2 having hypostria crossing more than one-third occlusal width. Three hundred and sixty-four cheek teeth are referred to an indeterminate species of Ochotona on the basis of their smaller size, rootless teeth, deep hypostria on upper molariform teeth, presence of a posteriorly directed process from the posteroloph on M2, nearly equal width of trigonid and talonid on lower molariform teeth, having two external and one internal cement-filled folds on p3. These specimens seem to be quite homogenous, but there is distinct variation in size and morphology. Ochotona are frequent in the Late Miocene faunas in central Nei Mongol. The taxon from Halajin Hushu, as well as those from Shala and Huitenghe of early Late Miocene age appears to be smaller than O. lagreli from the Ertemte Fauna of late Late Miocene in size.
Artiodactyla Because of the limited exposures, the Halajin Hushu site produced only one identifiable large mammal, mostly through screening efforts. A species of musk deer (Moschidae) is represented by two complete and one partial upper molar, one complete and one partial lower molar, and 10 complete or partial lower premolars (Fig. 5). The upper molars show some resemblance to Hispanomeryx, but its short and compact p4 seems consistent with Micromeryx. Halajin Hushu moschid may represent an undescribed species (Israel M. Sánchez pers. comm.) and until a detailed analysis is performed, we conservatively treat it as Moschidae indet. The Halajin Hushu materials also appear to be different from that in Amuwusu (see below).
Fig. 5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 5Selected specimens of moschids, Moschidae indet. from Halajin Hushu
Left upper molar (1-3), right upper molar (4-6), left m1/2 (7-9), right p4 (10-12), partial right p3 (13-15), and right p2 (16-18); 1, 4, 7, 12, 15, 18. buccal views; 2, 5, 8, 11, 14, 17. occlusal views; 3, 6, 9, 10, 13, 16. lingual views
Three genera of moschids are commonly recognized: the extinct Micromeryx and Hispanomeryx, and the extant Moschus. All three have been recorded in the Neogene of Nei Mongol, although generic status of these records is evolving. Schlosser (1924) first named a species of Moschus, M. grandaevus from Ertemte, Harr Obo, and Olan Chorea, which is perhaps closest to the living musk deer clade (Sánchez et al., 2010). Shortly after, Teilhard de Chardin (1926) erected another species, Moschus primaevus (he also spelled it M. primigenius in the captions of his figures in plate V), from Huitenghe (Chiton-gol), which was reassigned to Micromeryx (Mennecart et al., 2019). Recently, Sánchez et al. (2011) established a third species, Hispanomeryx andrewsi, from Wolf Camp. Other, largely undescribed, moschids from Nei Mongol include Micromeryx throughout the Damiao section (Kaakinen et al., 2015; Sukselainen et al., 2017; Zhang et al., 2011) and Micromeryx sp. from Amuwusu (Wang et al., 2015), the latter being first mentioned in an unpublished master’s thesis (Zhou, 1984). Although knowledge about Chinese moschids generally lacks behind its European counterparts, known records seem to indicate a comparable diversity and age range across Eurasia (Aiglstorfer et al., 2018; Mennecart et al., 2019; Sánchez and Morales, 2006; Wang et al., 2015), but Europe lacks the latest records of “Moschus” in the latest Miocene to Pliocene of East Asia (Vislobokova and Lavrov, 2009).
4 Composition of micromammalian fauna
The Halajin Hushu sample is one of the most diverse and abundant Neogene fauna in China. Compared to well-documented faunas in central Nei Mongol and other part of northern China, the assemblage seems to consist of a relatively complete fauna of small mammals, and may be considered as a rather representative association for Nei Mongol and the time of its origin. Nevertheless, it is reasonable to suspect that the sample is a mixed assemblage containing elements that appears to “not belong” to Late Miocene. There seem to be two grounds for this conjecture. The first is because of dominant genera commonly known from the Late Miocene localities of central Nei Mongol (Amuwusu, Balunhalagen, Huitenghe, Shala, Baogeda Ula, Bilutu and Ertemte), and quite a number of genera recorded for the first time in Late Miocene, such as Castor, Eozapus, Sinozapus, Lophocricetus, Paralactaga, Khanomys, Kowalskia, Microtoscoptes, Epimeriones, Pararhizomys, Prosiphneus and Ochotona, etc. (Daxner-H?ck, 1972; Qiu and Li, 2016). This seems to suggest that the real association (fauna) should be younger than Middle Miocene in age. However, the assemblage contains a few specimens of archaic genera which are considered to have become extinct by Early Miocene, for example, Tataromys and Parasminthus. It is unlikely that the two genera lived through much of the Middle Miocene and survived into the Late Miocene without being detected in those strata in Nei Mongol. The second reason is that distinct traces of erosion or transportation are seen in some specimens (e.g. Plesiodipus leei and Myocricetodon sp.) recovered from the channel deposits, which are frequently known genera or species in the Middle Miocene localities (Tairum Nur, 346 RM, Moergen, Tamuqin), but unknown in the Late Miocene. This seems to indicate that the assemblage contains specimens reworked by water from underlying strata. Out of 62 taxa in the Halajin Hushu small mammal assemblage, we recognize the following 6 species of mixed age (reworked form older strata): Tataromys sp., Parasminthus tangingoli, P. parvulus, Alloeumyarion sp., Plesiodipus leei, and Myocricetodon sp., or about 10% of the total. Most of these taxa are represented by less than 10 specimens.Reworked elements in an assemblage are easy to disturb consideration and age assessment of a fauna. Only those newcomers and prosperous taxa should serve as the true indicators of the depositional age. Thus, it is necessary to recognize contemporaries and tease out reworked elements. We prefer to consider taxa in this assemblage with the following characters as contemporaries: 1) forms only occurred or commonly known in the Late Miocene localities of central Nei Mongol, such as Yanshuella, Quyania, Paranourosorex, Ansomys, Tamias ertemtensis, Orientiglis, Leptodontomys, Castor, Eozapus, Sinozapus, Sicista, Lophocricetus, Paralactaga, Plesiodipus progressus, Gobicricetodon robustus, Khanomys, Kowalskia, Ochotona, etc; 2) new comers of Late Miocene, Zapodidae indet. 1 and 2, Colloides sp. nov., Epimeriones, Baranomyinae indet., for example. They are presumed to be true elements of the Halajin Hushu Fauna. However, those thriving in Oligocene and becoming extinct by Middle Miocene in the Asian Palearctic, i.e. Tataromys, Parasminthus and Alloeumyarion, and those predominant in the Middle Miocene faunas, but disappeared in the Late Miocene, such as Plesiodipus leei and Myocricetodon (specimens usually scarce and with traces of erosion or transportation) are here arbitrated to be elements from reworking of earlier strata, which should be rejected from the fauna (Table 1).
5 Biochronology
Among the 63 forms of mammals in the Halajin Hushu assemblage, 57 are considered as contemporary association derived from central Nei Mongol (Table 1). Although the list is a preliminary determination of the fauna, some tentative ideas on the aspects of biochronology, paleoecology and biogeography may be given.The Halajin Hushu Fauna contains 43 genera, 56 species of contemporary small mammals, belonging to 15 families. The majority of the families and quite a number of genera are already present in the Middle Miocene and are distributed over Eurasia, but the age of the fauna cannot be Middle Miocene because of the appearance of the family Myospalacidae, as well as some genera first appeared in the Late Miocene, such as Sorex and Paranourosorex in Insectivora, Castor, Colloides, and Kowalskia in Rodentia, and Ochotona in Lagomorpha. All the genera of the fauna, except Epimeriones are recorded in the Neogene of central Nei Mongol. Fig. 6 shows the generic occurrence of rodents of the fauna in the Neogene of this area. Among the 30 genera of rodents, half made their first appearance in Late Miocene, i.e. Pseudaplodon, Prospermophilus, Hylopetes, Eozapus, Sinozapus, Lophocricetus, Khanomys, Colloides, Microtoscoptes, Pararhizomys etc. The presence of these genera indicates an age of later than Middle Miocene for the fauna. Nevertheless, it is unlikely to be later than Late Miocene, because the families Arvicolidae, Muridae and Leporidae commonly known in Pliocene are absent from the fauna, and the Myospalacidae in this fauna are less diverse than those in Pliocene. This suggests that the age of the Halajin Hushu Fauna is younger than Middle Miocene but older than Pliocene.
Fig. 6
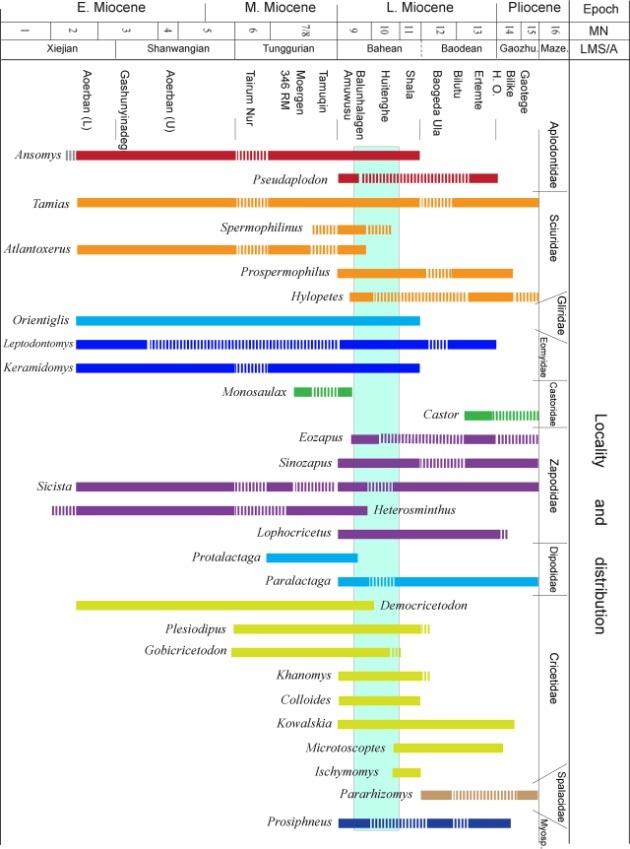 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPTFig. 6Biostratigraphic range of rodent genera of the Halajin Hushu Fauna in the Neogene of central Nei Mongol
Generally, the Halajin Hushu Fauna is characterized by the frequent retention of Middle Miocene holdovers, i.e. Ansomys lophodens, Orientiglis wuae, Leptodontomys lii, Keramidomys fahlbuschi, Monosaulax tungurensis, Heterosminthus orientalis, Democricetodon lindsayi, Gobicricetodon robustus, and highly specialized Middle Miocene survivors, e.g. Plesiodipus robustus, by the high diversity of Zapodidae, and the absence of Muridae, Leporidae, and some modern genera in Cricetidae, such as Nannocricetus, Sinocricetus, Microtodon, etc. The more retention of Middle Miocene holdovers, the high diversity of zapodids and the lacking of murids, leporids and modern hamsters in this fauna are indicative of an earlier age of the fauna in the Late Miocene.
According to the Chinese Neogene Land Mammal Stage/Age (LMS/A) proposed, the Late Miocene includes Bahean age and Baodean age (Qiu et al., 2013). In central Nei Mongol, the localities Amuwusu, Balunhalagen, Huitenghe and Shala are subordinated to the Bahean, while Baogeda Ula, Bilutu, Ertemte and probably Harr Obo to the Baodean (Qiu and Li, 2016). The following comparison of faunas may further demonstrate that the Halajin Hushu Fauna is closer to the Bahean faunas rather than the Baodean faunas. Although it shares quite a number of taxa with the Baodean Ertemte and Harr Obo faunas, such as Yanshuella, Quyania, Sorex, Paranourosorex, Pseudaplodon, Tamias ertemtensis, Prospermophilus orientalis, Hylopetes, Leptodontomys gansus, Castor, Eozapus, Sinozapus, Sicista, Lophocricetus, Paralactaga, Kowalskia, Microtoscoptes, Prosiphneus, and Ochotona, the Halajin Hushu Fauna lacks many Baodean newcomers that are very prosperous in the two faunas, such as Myomimus, Dipoides, Paralophocricetus, Brachyscirtetes, Dipus, Nannocricetus, Sinocricetus, Microtodon, Anatolomys, Pseudomeriones, and Alilepus. In addition, both the Ertemte Fauna and Harr Obo Fauna possess Muridae and Leporidae, and some genera, Paralophocricetus, Microtodon and Micromys for example, are rather dominant in the assemblages. This indicates that the Halajin Hushu Fauna is much older than the two faunas of Late Baodean age. The assemblage of Bilutu obviously contains elements reworked from earlier deposits. The presence of a relatively higher diversity of murids, and of more advanced genera and species, such as Pliopetaurista, Lophocricetus grabaui, Dipus, Nannocricetus, Sinocricetus, Microtodon, Anatolomys and Pseudomeriones, distinctly indicates that it is younger than the Halajin Hushu Fauna. Although the Halajin Hushu Fauna has Sicista, Lophocricetus xianensis, Paralactaga, Kowalskia shalaensis, Microtoscoptes, Prosiphneus, Pararhizomys, and Ochotona in common with the Baogeda Ula Fauna, the absence of Muridae and Leporidae, and the lack of some forms that also occur at Ertemte, i.e., Myomimus, Dipoides, Brachyscirtetes, Dipus, Nannocricetus, Sinocricetus, Microtodon, etc., imply that the Halajin Hushu fauna is still older than these Baodean faunas. In general, the Halajin Hushu Fauna shows a close community structure and taxonomic composition with the Bahean faunas, containing relatively more genera frequently seen in Middle Miocene and fewer taxa in Late Miocene. Among the small mammals, 17 genera and 15 species occur in the Amuwusu Fauna, 21 genera and 21 species in the Balunhalagen Fauna, 14 genera and 11 species in the Huitenghe Fauna, and 15 genera and 11 species in the Shala Fauna, respectively (Qiu and Li, 2016). This seems to present a substantial evidence for the Bahean age of the Halajin Hushu Fauna. As far as the lacking of Muridae and the advanced genera of Cricetidae is concerned, the fauna is closely similar to the Amuwusu Fauna, but the presence of more derived species, i.e., Protalactaga lantianensis and Paralactaga shalaensis, and the absence of Pentabuneomys and Hystricops appear to suggest that the Halajin Hushu Fauna may be slightly younger than the latter. On the other hand, the occurrence of Sinotamias gravis, Paralactaga parvidens, Salpingothus, Cardiocranius, Khanomys cheni, Microtocricetus, and Progonomys in the Shala Fauna may be indicative of a younger age than the Halajin Hushu Fauna. Thus, the fauna is considered to represent a stage of faunal evolution intermediate between the Shala Fauna and the Amuwusu Fauna in central Nei Mongol.
Outside Nei Mongol, the Halajin Hushu Fauna has in common with the early Late Miocene Bahe Fauna in Shaanxi Province the genera Pseudaplodon, Tamias, Protalactaga, Paralactaga, Lophocricetus, Kowalskia and Pararhizomys, but contains more survivors from the Middle Miocene and lacks Muridae and some genera frequently occurred in higher horizons, e.g. Sciurotamias in Sciuridae, Salpingothus and Cardiocranius in Dipodidae, Nannocricetus in Cricetidae, and Abudhabia in Gerbillidae (Qiu et al., 2004a, b). This testifies to an older age of the Halajin Hushu Fauna than the Bahe Fauna. The Shengou Fauna from Qinghai Province is considered to be another early Late Miocene fauna in China, which was thought to be close to but slightly younger than the Bahe Fauna in age due to the content of co-genera and the presence of more derived murid Huerzelerimys (Qiu and Li, 2008). Thus, the Nei Mongol fauna is even earlier in age than the Qinghai fauna. Other Late Miocene faunas in China, such as the Lingtai Fauna in Gansu, and the Lufeng and Yuanmou faunas in Yunnan, share few congeners with the Halajin Hushu Fauna (Ni and Qiu, 2002; Zheng and Zhang, 2000). These faunas contain abundant murids and leporids, which should be younger than the Nei Mongol fauna. The Halajin Hushu Fauna has at least 11 genera, Tamias, Spermophilinus, Keramidomys, Leptodontomys, Castor, Eozapus, Democricetodon, Kowalskia, Ischymomys, Epimeriones, and Ochotona, in common with European Late Miocene faunas, among which Democricetodon made its last occurrence in the MN 9 in Europe.
In summary, the age of the Halajin Hushu Fauna is likely of early Late Miocene, and of Bahean Chinese Land Mammal Stage/Age, or roughly equivalent to MN 9-10 of European land mammal zonation (Fig. 6).
6 Biogeography and paleoecology
Among the 15 families of small mammals known from the Halajin Hushu Fauna, most are eurytopic, spreading throughout the Old World and the New World. These are Erinaceidae, Talpidae, Soricidae, Aplodontidae, Sciuridae, Eomyidae, Castoridae, Zapodidae, Cricetidae, and Ochotonidae. Gliridae and Spalacidae are distributed over Eurasia and North Africa. Only Myospalacidae and Dipodidae are endemic to northern Asian Palearctic Region. At the genus level, more than half are confined to the Mongolian highland and its adjacent regions. Few are recorded from the Late Miocene of other areas of Asia (only Kowalskia in Yunnan), or from North America (Monosaulax and Castor) or Africa, but quite a number of genera, Tamias, Spermophilinus, Hylopetes, Leptodontomys, Keramidomys, Castor, Eozapus, Sicista, Democricetodon, Kowalskia and Epimeriones, occur in Europe, accounting for about one fifth of the total. This suggests dispersal of small mammals to some extent between the two contiguous continents during the Late Miocene, as shown by the other contemporary faunas in central Nei Mongol.In the mammals known from the Halajin Hushu locality, all the families except Eomyidae are extant. Among the extant families, Erinaceidae, Talpidae, Soricidae, Sciuridae and Cricetidae are eurytopic, others occur today either throughout the Holarctic Region (Castoridae and Zapodidae), or the Palearctic Region (Gliridae, Dipodidae and Myospalacidae), or the Nearctic Region (Aplodontidae), but none of the family is confined to the Oriental Region. The diversity and abundance of zapodids and cricetids, the presence of dipodids, and the flourishing of myospalacids and ochotonids in the fauna are indicative of a predominance of dry and open habitats in central Nei Mongol during the Early Miocene. It is evident that faunal composition of Halajin Hushu shows a temperate, relatively arid steppe environment, similar to the present northern Holarctic Region, as the other Bahean faunas in central Nei Mongol. Nevertheless, remains in this assemblage show that the present-day environment is not completely analogous to the Late Miocene. Relatively permanent water bodies are indicated by fossil fishes and frogs. The presence of beavers fit into such an environment. Judging from their living relatives, Ansomys, Pseudaplodon, Hylopetes and Eozapus would require dense thickets of shrubs, even forests. Modern moles also prefer moist shrub habitats and friable soils. The area of deposition of the fossiliferous sediments must have been more diversified in early Late Miocene time than it is today. The climate might be less dry, and the vegetation more dense than today.
It is worth mentioning that the high diversity of Zapodidae and Cricetidae, and the thriving and increasingly high-crowned Cricetidae (e.g. Epimeriones sp. nov., Baranomyinae indet.) in the assemblage are distinctive in the faunas of Bahean age. The faunal turnover seems to bear out again an environmental change towards drier and more grassland condition from Middle Miocene to Late Miocene. It is reasonable to infer that with the ecological and environment deterioration happened in early Late Miocene of central Nei Mongol, plant-eating animals rapidly increased and herbivores increased their crown height for adaptation to abrasive diets.
Acknowledgments
The authors would like to express their gratitude to Dr. Z. J. Tseng from University of Southern California Los Angeles, USA, Dr. Y. Tomida from National Science Museum Tokyo, Japan, Mr. W. Q. Feng from the IVPP for their hard works in the collections under study. We are indebted to Gertrud R?ssner and Israel Sánchez for discussions about fossil moschids. Many thanks are also due to Mr. Deliger from the Xilinhot Cultural Relic Station for his enthusiastic participation in providing logistical help, Song Yanping for help with Photoshop manipulations of images.参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
URL [本文引用: 1]

2, consists of three levels of lava platforms at different elevations, and the age of each lava platform is 2.31~2.41Ma、1.56~1.61Ma、0.51~0.61Ma. The basalt which can be divided into quartz tholeiite and olivine tholeiite has an excessive nature of the characteristics, all being derived from the garnet peridotite source region. Based on the geochemistry characteristics, there does not exist the evolutionary relationship between the olive tholeiite and quartz tholeiite. They are interpreted to be resulted probably from the varying degrees of the garnet peridotite source region and the depth of partial melting. Both olive tholeiite and quartz tholeiite were affected by crustal contamination, but the latter is more obviously. Tectonically, both Beilike and Chifeng lie on the the southern margin of the Xing'an-Mongolia Orogen Belt, compared with the northern margin of North China Craton, they have a similar magma source and the lithospheric mantle thermal state, but different enrichment lithospheric mantle types, namely, the former displays a DMM-EMⅡ array different from the latter, it may be related to different ages and tectonic settings of the lithospheric mantle. Based on the asthenosphere-lithospheric mantle interaction model, lithospheric thinning phenomenon in North China is not confined to the craton, instead in northwest craton, and even the south margins of Xing'an-Mongolia Orogen Belt experienced lithospheric thinning too. They just have different time and degree of the lithospheric thinning process.]]>
[本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
We argue that the production of mantle-derived or juvenile continental crust during the accretionary history of the Central Asian Orogenic Belt (CAOB) has been grossly overestimated. This is because previous assessments only considered the Palaeozoic evolution of the belt, whereas its accretionary history already began in the latest Mesoproterozoic. Furthermore, much of the juvenile growth in Central Asia occurred in late Permian and Mesozoic times, after completion of CAOB evolution, and perhaps related to major plume activity. We demonstrate from zircon ages and Nd-Hf isotopic systematics from selected terranes within the CAOB that many Neoproterozoic to Palaeozoic granitoids in the accreted terranes of the belt are derived from melting of heterogeneous Precambrian crust or through mixing of old continental crust with juvenile or short-lived material, most likely in continental arc settings. At the same time, juvenile growth in the CAOB occurred during the latest Neoproterozoic to Palaeozoic in oceanic island arc settings and during accretion of oceanic, island arc, and Precambrian terranes. However, taking together, our data do not support unusually high crust-production rates during evolution of the CAOB. Significant variations in zircon epsilon(Hf) values at a given magmatic age suggest that granitoid magmas were assembled from small batches of melt that seem to mirror the isotopic characteristics of compositionally and chronologically heterogeneous crustal sources. We reiterate that the chemical characteristics of crustally-derived granitoids are inherited from their source(s) and cannot be used to reconstruct tectonic settings, and thus many tectonic models solely based on chemical data may need re-evaluation. Crustal evolution in the CAOB involved both juvenile material and abundant reworking of older crust with varying proportions throughout its accretionary history, and we see many similarities with the evolution of the SW Pacific and the Tasmanides of eastern Australia. (C) 2013 International Association for Gondwana Research. Published by Elsevier B.V.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
DOIURL [本文引用: 1]
AbstractThe discovery of Lufengpithecus at Yuanmou, in Yunnan Province, China, provides important clues to understanding the evolution of early hominoids in eastern Asia. However, the age and paleoecology of the fauna remain to be established. An assemblage of micromammalian fossils, collected recently by screenwashing at Leilao, one of the hominoid-bearing localities at Yuanmou, contains 41 species belonging to 16 families and 4 orders, and represents the most diverse Neogene micromammalian fauna known from southwestern China. This paper presents an analysis of the taxonomy, age, and paleoecology of the micromammalian fauna. The Yuanmou hominoid fauna is now considered older than the Lufeng fauna, and is currently thought to correspond to an age of about 9 Ma. The micromammalian fauna indicates a predominantly forested habitat, with marginal environments consisting of bush-grassland.]]>
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 11]
DOIURL [本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOIURLPMID [本文引用: 1]
Damiao, Inner Mongolia, has three main fossil horizons representing the early, middle, and late Miocene. The middle Miocene locality DM01 is the only primate locality from the region and also represents the latest occurrence of pliopithecoids in northern China. The presence of pliopithecoid primates in central Asia after the middle Miocene climatic optimum seems to contradict the general trend of strengthening climatic zonality and increasing aridity. To investigate this enigma, we employ faunal similarity, ecometrics, and stable isotope analysis. Our results support previous inferences concerning the presence of locally humid environments within the increasingly arid surroundings that characterized central Asia. Hypsodonty, estimated mean annual precipitation (MAP), local sedimentology, and large mammal fossils suggest more humid and possibly more forested and wooded environments for the DM01 locality. We compared our results with the adjacent fossil-rich middle Miocene Tunggur localities. However, the small mammal fauna and isotope data are consistent with a mosaic of forest and grassland environment for all Damiao localities. Based on our results, Tunggur may have been too seasonal or not sufficiently humid for pliopithecids. This is supported by the higher mean hypsodonty and lower estimated MAP estimates, as well as slightly higher delta(13)C values. We suggest that DM01, the driest known Asian pliopithecid locality, may have been a more humid refugium within a generally drier regional context.
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 2]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 1]
URL [本文引用: 1]
e.g. Lagomeryx sp. ,Stephanocemas sp.nov,and Micromeryx sp.Small mammals are dominated by Metexallerix gaolanshanensis,Sinolagomys ulunguensis,Tachyoryctoides sp.nov,Prodistylomys wangae,Distylomys cf.D.tedfordi, Sinodonomys sp. ,Plesiosminthus sp. ,Heterosminthus sp. Pseudotheridomys sp.nov. ,and Eomyidae gen.et sp.nov.The late Middle Miocene fauna includes more than 30 species,sharing with the classical Tunggur fauna Mioechinus?gobiensis,Desmanella storchi,Bellatona forsythmajori,Atlantoxerus orientalis,Heterosminthus orientalis,Protalactaga grabaui,Gobicricetodon flynni,and Stenofiber hesperus,however, with more diversified cervoids and a species of pliopithecid.The early Late Miocene fauna show great similarity with the previous fauna,except with the earliest record of Late Miocene elements Lophocricetus,Eozapus, Nannocricetus etc,and disappearance of the typical Middle Miocene Atlantoxerus,Democricetodon,and Heterosminthus.All three faunas have rich small mammals(insectivores,rodents and lagomorphs)and diversified cervoids.The similar faunal community structure and continuity of main lineages suggest a relatively stable moderate humid and warm forest-grassland environment from the Early Miocene to early Late Miocene.The significant environment change seems to have happened after the earliest Late Miocene.]]>
[本文引用: 1]
[本文引用: 1]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}