 ,2)中国科学院大学生物运动力学实验室, 北京 100049
,2)中国科学院大学生物运动力学实验室, 北京 100049PROPAGATION CHARACTERISTICS OF CURVATURE WAVE ALONG THE BODY IN ANGUILLIFORM FISH SWIMMING 1)
Shen Haoyan, Zhu Bowen, Wang Zhihui, Yu Yongliang,2)The Lab of Biomechanics for Animal Locomotion, University of Chinese Academy of Sciences, Beijing 100049, China通讯作者: 2)余永亮,教授,主要研究方向:飞行和游动的生物运动力学、空气动力学. E-mail:ylyu@ucas.ac.cn
收稿日期:2019-04-9接受日期:2019-05-9网络出版日期:2019-07-18
| 基金资助: |
Received:2019-04-9Accepted:2019-05-9Online:2019-07-18
作者简介 About authors

摘要
将鳗鲡模式游动的七鳃鳗简化成材料性质均匀的变截面黏 弹性梁,通过数值方法求解鱼体在主动弯矩波(作为激励的驱动波)的驱动下匀速游动时身体变形曲率波的传播特性. 结果表明,当主动弯矩 的驱动频率高于鱼体结构基频时,可以观察到曲率波相对于驱动波存在相位滞后,且越靠近尾部滞后现象越明显,这意味着曲率波的波速 小于驱动波的波速,也间接地验证了前人的实验结果. 通过参数研究发现,鱼体变形曲率波与驱动波的波速比与表征流体黏性作用的雷诺数无关,而与表征驱动波和鱼体材料属性的 无量纲激励频率、激励波长及鱼体黏性系数有关. 对于鳗鲡模式游动的鱼类,曲率波与驱动波的波速比随着无量纲激励频率和波长的增大而降低,随着鱼体黏性系数的增大而增大. 进一步研究发现,通过小扰动分析得到的组合相似性参数$\varPi$可以统一描述波速比与激励参数、材料参数之间的关系.
关键词:
Abstract
In this study, the homogeneous viscoelastic beam with a variable cross-section is used to model the lamprey, an anguilliform (eel-like) swimmer, and to research the propagation characteristics of the curvature wave driven by an active bending moment wave along the fish body. The results show that as long as the excitation frequency is higher than the structure fundamental frequency of the fish body, there will be a phase lag between the two waves, which increases from the head to the tail of the fish body. The increasing phase lag indicates that there exists a speed difference between propagations of the active bending moment and the body curvature, that is, the speed of the later is lower than that of the former. This features are consistent with the experimental results published. The dimensional analysis indicates the ratio of the speed of bending curvature wave to that of the active bending moment wave is associated with the dimensionless excitation frequency, wavelength and damping coefficient, but independent of the swimming Reynolds number. For anguilliform (eel-like) swimmers, the wave speed ratio decreases with increasing frequency or wave length of active bending moment, and it rises if the damping coefficient becomes larger. In addition, we also carried out a small perturbation analysis to linearize the equations, and found an integrated similarity parameter which includes the dimensionless frequency, wave length and damping coefficient. This parameter can uniformly describe the dependence of the speed ratio on the excitation and material parameters.
Keywords:
PDF (9071KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
沈昊嫣, 朱博闻, 王智慧, 余永亮. 鳗鲡模式游动的鱼体变形曲率波的传播特性研究 1). 力学学报[J], 2019, 51(4): 1022-1030 DOI:10.6052/0459-1879-19-087
Shen Haoyan, Zhu Bowen, Wang Zhihui, Yu Yongliang.
引言
鳗鲡模式游动的鱼类(如鳗鱼)在匀速游动中都会呈现出明显的波动特征,与鲹科模式游动的鱼类(如鲤鱼)相比,鱼体弯曲变形的波 长显著小于体长. 然而,生物学家在测量了前游的鱼体两侧肌电信号后发现,肌电信号并不与鱼体弯曲变形完全对 应[1-2]. 鱼体发生变形的这一过程是在神经信号刺 激、肌肉收缩、鱼体与周围流体介质的相互作用下实现的. 在这过程中,鱼体左右两侧的肌肉交替收缩产生主动弯矩,且肌电信号显示鱼体上的肌肉是从头到尾依次收缩的[3-4]. Williams等[5]对七鳃鳗的定量实验表明(如图1所示),鱼体上各点均存在着弯曲变形滞后于肌电激励信号的现象,在前游时尾 部滞后更加明显,肌电信号的传播波长约为1.05倍的体长,且从头到尾的传播速度比鱼体弯曲变形的曲率行波传播速度快.图1
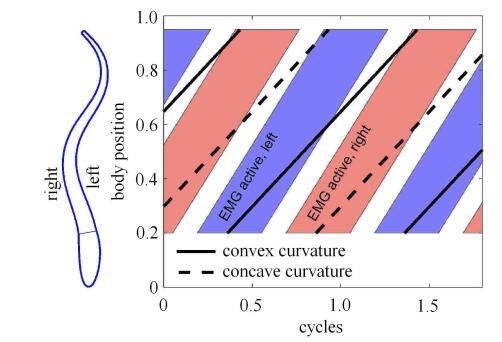 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1鱼体游动时肌电信号序列和鱼体弯曲变形示意图
Fig. 1Sketch of passage of the electromyograms (EMG) signal and maximum convex (solid lines) and concave (dashed lines) curvature of bending body during swimming cycles
从神经信号刺激到鱼体变形的整个过程中可以划分为两个阶段,一个是从神经信号到肌肉输出主动弯矩的动力学阶段,另一个是鱼体 在主动弯矩和流体力作用下的变形. 这两个阶段均会存在相位滞后和传播速度的差异,主动弯矩的相位滞后于神经信号,鱼体弯曲变形的相位滞后于主动弯矩. 前者是生理学研究的重点,后者可归结为一个力学问题. Wardle等[6]实验测量了多种模式游动的鱼体,总结归纳出相位滞后及波速差异的现象与肌肉功能和激活周期有关. McMillen等[7-8]从钙离子肌肉力模 型[9]出发分析此问题,认为鱼体材料的黏弹性和鱼体形状是影响相位滞后的主要原因. 由于涉及复杂的生理学过程,对相位滞后及传播速度差异的定量研究存在困难,故本文暂不考虑神经信号的传递过程,主要关注鱼体 变形与主动弯矩的相位滞后和传播速度的差异.
鱼体的变形是在主动弯矩和流体力的共同作用下发生的,这可被看作是一个流固耦合过程[10]. 在先前对鱼类游动的研究中,大 多采用给定鱼体变形规律求解鱼体受力和运动[11-14]. 而本文基于Williams等[5]为研究七鳃鳗而建立的鱼体黏弹性连续梁模型,通过量纲分析和小扰动假设下的量级分析,研 究鱼体在主动弯矩和流体力共同作用下发生弯曲变形的力学响应.
1 研究模型与研究方法
1.1 黏弹性梁的鱼体模型
通常,成年七鳃鳗的体长约30 cm,身体截面近似椭圆形(如图2(a)所示),鱼体高度随体长的变化小,而厚度随体长逐渐减小;鱼体的头 部较硬,变形量很小;除头部外的身体较柔软,在游动时可呈现出波状变形. 因此,根据七鳃鳗的体态特征,建立了如图2(b)所示的简化几何模型,设定模型长度为$L=30$ cm,头部长度为$L_{ h}=0.2L$;截面为椭圆, 椭圆在鱼体高度方向为长轴,在宽度方向为短轴,半长轴$a$和半短轴$b$沿着体长分布规律可描述为其中,$a_0$和$b_0$分别是半长轴和半短轴的特征尺寸,两者皆取为1 cm; $\alpha$是鱼体宽度的线性收缩比,在此取为0.8.
图2
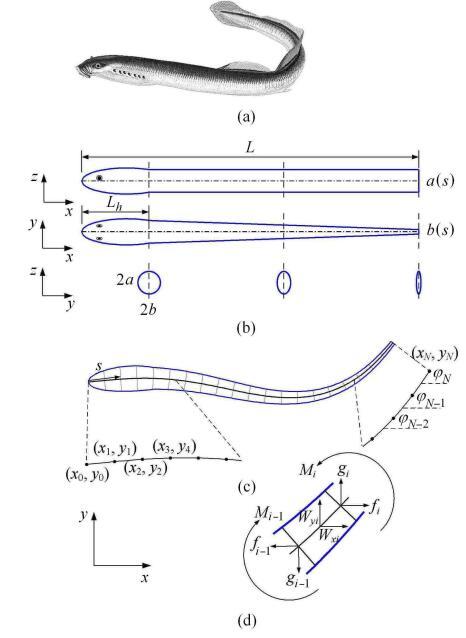 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2七鳃鳗简化模型和鱼体单元的受力分析图. (a)七鳃鳗Lampetra fluviatilis(图片来源于公共网络) (b) 鱼体模型侧视图、俯视图和截面,鱼体长度为$L$,鱼头长度为$L_{h}$,鱼体半高度为$a(s)$,半宽度为 $b(s)$;(c) 鱼体离散单元和(d)离散单元的受力图
Fig.2(a) Lamprey (Lampetra fluviatilis). (b)Side view, top view and sectional view of lamprey model. $L$: length of lamprey; $L_{ h}$: length of fish head; $a(s), b(s)$: semi-axes of elliptical cross-section. (c) discrete segments of lamprey model and (d) Force and moments acting on a discrete element
进一步地,把鱼体看作是变截面的黏弹性梁模型,为了简化分析,不妨忽略鱼体的内部结构,把鱼体骨骼、肌肉、皮肤,甚至 内脏等都看作具有统一力学行为的材料,故而在该黏弹性梁的力学模型中,材料的等效力学参数仅包括了密度$\rho$、杨氏模 量$E$和黏性系数$\mu$. 那么,用该模型就可以模拟细长体鱼类在流体中受主动弯矩驱动下发生变形、与周围流体相互作用进而产生整体游动的过程.
在惯性参考系中建立直角坐标系,用$(x,y)$表示鱼体中线上的坐标. 同时,沿着鱼体中线建立曲线坐标$(s,\varphi)$, $s$表示从鱼 头开始的曲线长度,$\varphi$表示鱼体中线的切线与$x$轴夹角(简称偏转角). 如图2(d)所示,选择鱼体上微元$ds$,根据动量和动量矩定理,可建立鱼体运动和变形的方程
其中,$A=\pi ab$和$I=\pi ab^3/4$分别是鱼体截面的面积和面积矩, $W_x$和$W_y$是流体力沿$x$轴和$y$轴的分量, $f$和$g$是鱼体内部接触力沿$x$轴和$y$轴的分量, $M$是作用于梁的弯矩. 鱼体中轴线不可拉伸,满足几何关系
在该动力学问题中,方程(4)右端的弯矩$M$包含了由肌肉产生的主动弯矩、由内应力(弹性应力和黏性应力)导致的等效力矩,即
上式仅肌肉产生的主动弯矩是鱼游动力学过程的输入项.
当鱼体在定速游动时,通常靠体侧的红肌伸缩产生主动弯矩,McMillen等[7-8]认为主动弯矩的大小与鱼体宽度的平 方成正比;Cheng等[15]和张薇 等[16-17]从鱼体变形反推了鱼体内肌肉产生的主动弯矩强度,认为该强度具有两端 小中间大的钟形分布特征. 因此,不妨设鱼体内主动弯矩为如下形式
其中,$\omega$和$\lambda_{act}$ 分别是主动弯矩波的角频率和波长,则主动弯矩波的传播速度为$v_{ act}=\omega \lambda_{act}/(2\pi)$;$\beta$为主动 弯矩的强度因子,通过调节该因子可实现调控鱼体摆尾的幅值$A_{tail}$.
对于流体力${\pmb W}=(W_x, W_y)$,本文采用Taylor抗力模型[18]来计算. 在曲线坐标系里
在法向${\pmb n}$和切向${\pmb \tau}$的分量分别为
其中,$v_n$和$v_{\tau}$分别表示鱼体表面法向和切向运动速度;$\rho_f$ 为流体密度,$\mu_{ f}$为流体的黏性系数. $C_n$为法向力系数(在该问题中取为1). Taylor抗力模型适用的雷诺数范围为 $20<Re<10^5$,此处雷诺数 $Re=2a\rho_{f} A_{tail}\omega/\mu_{f}$.
1.2 参数的无量纲化
鱼体在主动弯矩的作用下发生动态的弯曲变形可视为曲率波的传播. 但由于问题的复杂性,鱼体波动并不是简谐波,故而在此给 出鱼体平均曲率波传播速度的计算方法. 当鱼体处于稳定游动状态时,在一个周期$T$内对曲率$\kappa (s,t)=\partial \varphi (s,t)/\partial s$ 进行如下运算可获得鱼体中线各点弯曲变形的初始相位$\psi_0(s)$,即则鱼体的相位为$\psi(s,t)=\psi_0(s)-\omega t$,波数$k$的分布为
由于鱼体上各点的曲率波传播速度并不相等,可采用平均波数 $\bar k$和频率$\omega$来定义鱼体曲率波波速$v_{cur}$,故曲率波与主动弯矩波的传播速度比(简称波速比)可表达为
对于1.1节给出的简化鱼体动力学模型,存在着三类参数. 第一类是主动的输入参数,如主动弯矩的频率$\omega$、 波长$\lambda_{act}$和强度因子$\beta$等. 第二类是鱼体的属性参数,包括鱼体的几何参数和材料参数. 几何参数如鱼体长度$L$、截面参数$a_0$, $b_0$和$\alpha$等;材料参数包括密度$\rho$、等效的杨氏模量$E$和黏性系数$\mu$ 等. 第三类为流体的属性参数,包括流体密度$\rho_{f}$和黏性系数$\mu_{f}$ 等. 在本文研究的问题中,设鱼体的密度和流体密度相等,即$\rho=\rho_{f}$. 此外,本文还引入了一个约束条件,即鱼尾摆幅$A_{tail}$ 要与实验观察一致,这就使得强度因子$\beta$ 成为了计算中的调节量而不再作为激励的输入参量. 因此,形式上,对于我们关心的鱼体曲率波传播速度$v_{ cur}$,可以表达为
根据相似性定理,本文选择$L, E$和$\rho$ 作为参考物理量对其他物理量进行无量纲化. 在此,引入变截面梁的基频
基频表达式中的参数$\gamma$与梁的几何参数有关,对于本文给定的鱼体模型,$\gamma=0.27$. 因此,波速比可以写作4个无量纲量的函数
1.3 小变形下的线性化分析
当鱼体小幅波动达到匀速前游时(小扰动假设),可忽略身体轴向内力和鱼体局部的转动加速度,于是方程(2)~(5)可以简 化成一个方程,即鱼体中线相对于平衡位置的偏离量$h(s,t)$满足的连续梁方程上式即为在流体力和主动弯矩作用下的变截面Euler-Bernoulli梁的方程. 方程左边的三项为响应,分别是:鱼体弹性力项、 黏性力项以及鱼体波动时惯性力项,依次记为$F_E, F_{\mu}$ 和$F_i$. 其中,鱼体弹性力和黏性力与鱼体的材料属性有关,而惯性力项与鱼体的质量分布有关.
作为简化分析,不妨设鱼体中线法向偏移量$h(s,t)$具有简谐特性,即$h(s,t)=h_0 {e}^{{i}(2\pi s/\lambda -\omega t)}$, 故而,运动方程左侧三项的量级大小分别为
其中,$I_0$和$A_0$为特征截面参数,而$\omega$和$\lambda$分别为激励频率和波长. 与材料属性有关的弹性力和黏性力可以表示为
由此可知,$\omega \mu/E $将带来相位的差异. 进一步地,考虑到惯性力项的相对大小,可以得到
为了方便后续分析,此处引入组合无量纲量
它包含了3个无量纲参量${\omega}/{\omega_0}$, $\lambda_{act}/L $ 和${\omega_0 \mu}/E $,这表明该动力学过程中3个可变的无量纲参量之间具有一定的联系,将在第3节中进行讨论.
2 计算结果
2.1 参数的选取和计算方法
七鳃鳗通常生活在近海区域或淡水中,在本文的定量研究中需要确定水的物性参数,在此设水的密度和黏性系数分别 为$\rho_{f}=1.0\times 10^3$ kg/m$^3$ 和$\mu_{f}=1.0$ mPa$\cdot$s.在第1.1节给出的模型中,鱼体的材料参数包括鱼体的密度$\rho$、杨氏模量$E$ 和黏性系数$\mu$,其中鱼体密度与水的密度相等,但是鱼体等效的杨氏模量和黏性系数难以确定. 在不同的实验中,获得的$E$和 $\mu$取值范围不同. 例如,Long[19]测量了鳗鱼在摆尾频率3 Hz情况下等效杨氏模量和黏性系数的平均值分别是$E=0.39$ MPa 和$\mu=11$ kPa;Long等[20]测量出盲鳗的等效杨氏模量$E=0.30$ MPa,但在摆尾频率3 Hz和1 Hz的情况下,黏性系 数分别取值为$\mu=2$ kPa$\cdot$s 和$\mu=5.7$ kPa$\cdot$s;Tytell等[21-22]研究的另外一种七鳃鳗(ichthyomyzon unicuspis)在摆尾频率1 Hz的情况下,等效的材料参数的平均值$E=73$ kPa和$\mu=0.22$ kPa$\cdot$s. 在本文的研究中,不妨先采用Tytell等[21-22]给出的七鳃鳗鱼体材料参数,即$E=73$ kPa 和$\mu=0.22$ kPa$\cdot$s,然后在第3节中讨论黏性系数对曲率波传播速率的影响.
对于输入参数的取值,在Williams等[5]的实验测量结果中,肌电信号的波长是鱼体体长的1.05倍,鱼体摆尾频率$f=5$ Hz. 因此,在本文中不妨假设肌肉产生的主动弯矩波的波长与肌电信号的波长一致,即$\lambda_{ act}=1.05L$,且主动弯矩波的频率与鱼体响应频率一致,即$\omega=2\pi f$. 根据Williams等[5]实验拍摄的图像(文献[5]中的图1),单侧摆尾的幅值大约为0.10$L$,故本文中不妨设定摆尾幅值为$A_{ tail}=0.1L$,此时肌肉的强度因子$\beta=4.6\times 10^5$ N/m$^2$.
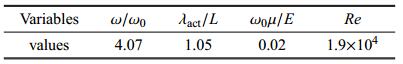
由此,可以得到模型的结构基频$\omega_0=7.72$ s$^{-1}$,则式(15)中4个无量纲参数的取值由表1给出.
Table 1
表1
表1无量纲参数表
Table 1
 |
新窗口打开|下载CSV
在本文对方程(2)$\sim $(4)的数值求解中,采用McMillen等[7]附录中提供的空间和时间离散格式,即空间 的二阶中心差分格式和时间的二阶向后差分格式,对离散后得到的隐式非线性方程组采用Newton迭代法求解. 空间上把鱼体沿中轴线均分为200份,时间上把一个摆尾周期均分为500份,计算结果均验证了空间和时间步长的无关 性(与体长均分1000份和周期均分2000份相比较,鱼体上偏转角度$\varphi$的峰值误差小于0.3%).
2.2 结果分析
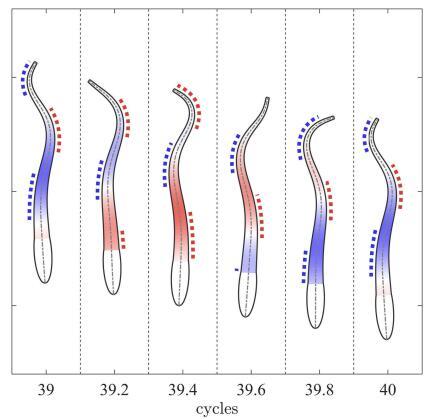
基于表1给定的无量纲参数,通过数值方法计算出鱼体在主动弯矩作用下的摆动(如图3所示)和身体中线曲率的时 空分布(如图4(a)所示). 虽然鱼头和鱼尾的主动弯矩都小,但由于鱼头质量分布大而变形较小,鱼尾因质量分布小而变形较大. 图3显示,鱼体在一个周期中,曲率波从头部传向尾部,这与观测到的鱼体运动相符. 从图中可以看到,体内主动弯矩为正的区域,曲率也对应为正,反之亦然;但是,在尾部区域,这种对应关系减弱,曲率的波峰明显 滞后于主动弯矩的波峰.鱼体中线曲率$\kappa (s,t)$的分布如图4(a)所示,正曲率的等值线用细实线表示,负曲率的等值线用细虚线表示,两者分别表示正 向弯曲和反向弯曲;红色粗虚线显示的是主动弯矩波峰的传播曲线,黑色粗实线显示的是正曲率波峰的传播曲线. 结果显示,正向弯曲和反向弯曲交替从头部传播到尾部;曲率波的波峰滞后于主动弯矩的波峰,且相位滞后程度从头至尾逐渐增加, 这表明曲率波的传播速度慢于主动弯矩的波速. 此现象与Williams等[5]观测到的曲率波滞后于肌电信号类似.
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3一个周期内鱼体的姿态变化图. 鱼体内色彩深度表示主动弯矩绝对值的大小,鱼体两侧的虚线分别表示 正曲率(红色为 $0 \pm 0.3\pi$ 相位区)和负曲率(蓝色为$\pi\pm 0.3\pi$相位区)
Fig. 3Locomotion of fish body during one swimming cycle. Color intensity on the body indicates the absolute value of active bending moment. The dashed lines near the two sides of body indicate positive and negative curvature regions
图4
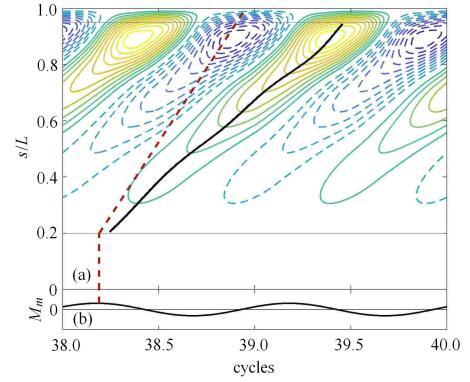 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4(a)鱼体中线各点曲率的时空分布图 和(b)鱼体上距鱼头0.2倍体长处主动弯矩随时间的变化图. 图(a)中细实线和细 虚线分别表示正曲率和负曲率的等值线,黑色粗实线(鱼体中线各点曲率达到最大值的连线)和红色虚线(鱼体中线各点主动弯矩达到最大值连线)的斜率分别表示曲率波和主动弯矩波的传播速度
Fig. 4(a) Spatio-temporal patterns of body curvature, and (b) active bending moment wave at the location of 0.2 times of body length. Thin solid and dashed lines indicate positive and negative curvature respectively. Slopes of thick solid line (each body point reaches maximum curvature) and red thick dashed line (each body point reaches maximum active bending moment) indicate the wave speeds of curvature and active bending moment respectively
进一步地,分别根据式(10)和式(12)计算出鱼体中线各点的初始相位$\psi_0(s)$ 和曲率波的平均传播速度. 计算中忽略变形较小的鱼头和变形较大的鱼尾, 只对 $0.2L\sim 0.95L$区间求平均波数,求得曲率波的平均波长为$0.59L$,波速比 $v_{cur}/v_{act}=0.56$,该值略低于根据Williams等[5]的实验结果计算的值$0.69\pm 0.09$. 这表明,尽管驱 动鱼体变形的弯矩波的波长接近一倍体长,但由于鱼体黏性和流体力的影响,鱼体变形的曲率波长小于体长,故而可以观察到鱼体 上存在一个以上的完整曲率波. 为此,在下面一节中将讨论不同的无量纲参数取值对波速比的影响.
3 分析与讨论
3.1 雷诺数${Re}$对波速比的影响
根据1.1节中介绍Taylor抗力模型时定义的雷诺数进行估算,采用鳗鲡模式游动的生物雷诺数范围大致为$10^3\sim 10^5$,在抗力理 论适用范围之内. 在 式(9)中,法向流体力$W_n$ 由两项组成,第二项和第一项的比值量级为$1/\sqrt{Re}$,这表明在本文给定的雷诺数范围内第二项(黏性项)是个小量. 由于流体 法向力对连续梁振动波的影响远大于切向力,因此,可以估计波速比受雷诺数的影响较小.图5给出了不同雷诺数条件下波速比随无量纲频率变化的曲线,结果显示,在$Re=1.0\times 10^3 \sim 1.0\times 10^5$范围内,曲线 基本重合,这说明雷诺数对波速比的影响的确较小,可以忽略.
图5
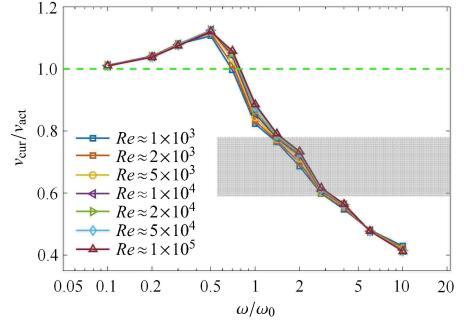 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5不同雷诺数条件下的波速比$v_{cur}/v_{act}$ 随无量纲参数 $\omega/\omega_0$的变化关系. 阴影区域表示根据Williams等[5]的实验结果计算的波速比的区域($0.69\pm 0.09$)
Fig. 5The speed ratio $v_{cur}/v_{act}$ in relation to dimensionless frequency $\omega/\omega_0$ under different $Re$. Shaded area represents the speed ratio ($0.69\pm 0.09$) calculated from experiment (Williams et al[5])
图5显示,随着$\omega/\omega_0$的增大,波速比$ v_{cur}/v_{act}$ 先增大后减小,大体上,在$\omega/\omega_0<1$ 的区域,会出现波速比大于1的情形;在$\omega/\omega_0>1$ 的区域,波速比小于1. 在对鳗鲡模式游动的鱼类的实验观测中,波速比通常小于1,比如Williams等[5]的波速比实验结果 为 $0.69\pm 0.09$,该部分区域在图5中用阴影标记. 这表明,对于鳗鲡模式游动的鱼类,在游动时激励频率$\omega$大于鱼体结构基频$\omega_0$.
当激励频率$\omega$小于鱼体结构基频$\omega_0$时,其波速比 $ v_{cur}/v_{ act}$大于1,这意味着曲率波的波速大于主动弯矩的传播速度. 这是由于激励频率较低时,鱼体以一阶振动 响应为主,扰动波的传播由鱼体的基频决定,因此,最终形成的曲率波传播速度超过了肌肉产生弯矩波速度. 在本文研究的鳗鲡模式鱼类的游动中,这属于没有观测到的反常现象,故而在下文 我们将不再讨论$\omega/\omega_0<1$的情形.
3.2 鱼体黏性$\omega_0\mu/E$对波速比的影响
在无量纲 参数中,影响波速比的材料参数仅剩下 黏性系数$\omega_0\mu/E$. 然而, 根据Long等[19-20]和Tytell等[21-22]对鳗鲡类鱼体材料参数的测量结果,无量纲黏性系数的取值 在0.02$\sim $0.5之间. 作为模型研究,本文 讨论$\omega_0\mu/E$ 的变化范围为0$\sim $5.图6(a)给出了在不同无量纲黏性系数下的波速比随无量纲频率变化的曲线图. 从图中可看出,在$\omega/\omega_0>1$的区域,黏 性不改变波速比随频率增大而减小的趋势;但是,当黏性增大时,曲线抬升,这意味着黏性将增大波速比,使得曲率波的波速越接近 主动弯矩波的波速. 定性上,根据介质中的振动理论,黏性相当于增大了等效刚度,提高了扰动波的速度,故而使得鱼体曲率波增大,这是黏性改变波速 比的主要原因.
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6不同无量纲黏性系数$\omega_0\mu/E$下波速比与(a)驱动波频率和(b)组合无量纲量$\varPi$的关系. 阴影区域为实验观 察到的波速比区域
Fig. 6The speed ratio in relation to (a) the dimensionless frequency $\omega/\omega_0$ and (b) the dimensionless variable $\varPi$ under different damping coefficients $\omega_0\mu/E$
进一步,采用1.2节中由小扰动梁导出的组合无量纲参数$\varPi$作为自变量,重新对数据进行处理,结果如图6 (b)所示. 从图 中可以看出,在$\omega/\omega_0>1$ 的区域,不同黏性系数下的波速比随$\varPi$变化的曲线有重合的趋势,特别是在阴影区域中曲线几乎重合(根 据Williams等[5]的实验结果标记). 由于组合无量纲量$\varPi$ 中黏性部分与$\omega \mu /E$有关,即黏性对波速比的影响大小与无量纲频率$\omega/\omega_0$和无量纲黏性$\omega_0\mu /E$的乘积有关.
3.3 激励波长$\lambda_{act}/L$对波速比的影响
在2.1节中,由于未考虑肌电信号与肌肉输出主动弯矩之间的差异,假定了主动弯矩波的波长与肌电信号的波长一致,不失一般性,在 此讨论主动弯矩波的波长对波速比的影响. 不妨假设波长变化范围为$\lambda_{act}/L =0.6\sim 1.2$.图7(a)给出了不同主动弯矩波长条件下波速比随$\omega/\omega_0$ 的变化规律,结果表明,在$\omega/\omega_0>1$的区域,当波长增大时,曲线下降,这意味着波速比随着主动弯矩的波长增大而减小. 定性上,由于波长增大,对于给定激励频率$\omega$的弯矩波,相当于增大了弯矩波的波速$v_{ act}$,故使得波速比降低.
进一步地,采用无量纲参数$\varPi$作为自变量,重新对数据进行处理,结果如图7 (b)所示. 在$\omega/\omega_0>1$的区域,不同 的弯矩波波长条件下的波速比$v_{cur}/v_{ act}$随$\varPi$变化的曲线几乎重合. 这表明弯矩波的波长对波速比的影响强弱与$(\lambda_{act}/L)^2$相关.
图7
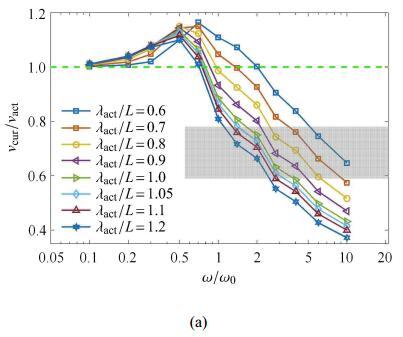 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同主动弯矩波长$\lambda_{act}/L$ 下波速比与(a)驱动波频率和(b)组合无量纲量$\varPi$的关系图
Fig. 7The speed ratio in relation to (a) the dimensionless frequency $\omega/\omega_0$ and (b) the dimensionless variable $\varPi$ under different wavelengths of active bending moment
图7
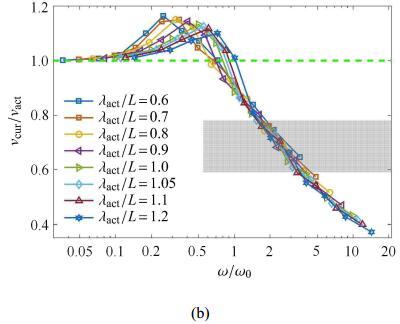 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7不同主动弯矩波长$\lambda_{act}/L$ 下波速比与(a)驱动波频率和(b)组合无量纲量$\varPi$的关系图(续)
Fig. 7The speed ratio in relation to (a) the dimensionless frequency $\omega/\omega_0$ and (b) the dimensionless variable $\varPi$ under different wavelengths of active bending moment (continued)
综上所述,对于鳗鲡模式游动的鱼类,影响曲率波波速和主动弯矩传播速度比值$v_{cur}/v_{act}$的主要因素 有主动弯矩 波的无量纲频率$\omega/\omega_0$、波长 $\lambda_{act}/L $和表征鱼体材料属性的无量纲黏性$\omega_0\mu/L$,而与游动的雷诺数$Re$无关. 对于本文研究对象七鳃鳗而言,由于无量纲 黏性较小,对参数$\varPi$ 的值影响小,因此即使黏性测量结果有误差,也对波速比的结果影响不大.
4 结论
本文对七鳃鳗进行了几何和物理建模,通过小扰动假设进行了定性分析,然后采用数值计算验证了鳗鲡模式游动的鱼类在定速前游时 鱼体变形的曲率波和驱动变形的主动弯矩波之间存在相位滞后和传播速度减慢的现象. 通过对结果的分析和讨论,可以得到如下两点结论.(1) 当驱动鱼体变形的主动弯矩频率大于鱼体结构基频时,可以观察到鱼体变形的曲率波相对于驱动波具有相位滞后的现象,且曲率波 的波长$\lambda_{cur}$ 小于驱动波的波长$\lambda_{act}$,曲率波的波速$v_{ cur}$低于主动弯矩波的速度$v_{act}$;波速比$v_{cur}/v_{act}$ 与无量纲的驱动频率$\omega/\omega_0$、波长$\lambda_{ act}/L$和鱼体黏性$\omega_0\mu$有关,而与游动的雷诺数$Re$无关.
(2) 通过小扰动分析,给出了无量纲量$\varPi=\dfrac{\omega}{\omega_0}\Big ( \dfrac{\lambda_{act}}L \Big )^2 \Big [ 1+\Big ( \dfrac{\omega \mu}E\Big )^2 \Big ]^{-\tfrac 14}$,它是一个重要的组合控制参数. 对本文研究的鳗鲡模式游动的鱼 类($\omega>\omega_0$),鱼体变形的曲率波与主动弯矩波的波速比近似地仅依赖于该组合物理量的取值.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 11]
[本文引用: 1]
DOIURL [本文引用: 3]
DOIURL [本文引用: 2]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
DOIURL
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 1]
DOIMagsci [本文引用: 1]

在综合游动力学的框架下采用整体模化分析方法,对比研究了鳗鲡鱼类和鲹科鱼类巡游中红肌(驱动鱼类巡游的骨骼肌)的力学性能异同. 该方法将观测总结的鱼体动态变形规律作为已知条件,首先确定鱼游中体外作用(包括流体动力响应和惯性力),然后间接预测活体的体内作用(包括肌肉主动力和生物组织被动应力). 研究结果显示,鳗鲡鱼类尾部的肌肉强度明显低于其躯干部分,而鲹科鱼类尾部和躯干部分肌肉强度相当,这与各自的体内作用主导机制相适应. 且体外作用的区别也导致了鲹科鱼类的能效更高. 同时发现,两种模式游动下都存在鱼体躯干内部从头至尾的能量传递,其肌肉输出净功沿鱼体轴向都呈“钟形” 分布. 总体上沿轴向各点的肌肉都输出正的净功.
DOIMagsci [本文引用: 1]
在综合游动力学的框架下采用整体模化分析方法,对比研究了鳗鲡鱼类和鲹科鱼类巡游中红肌(驱动鱼类巡游的骨骼肌)的力学性能异同. 该方法将观测总结的鱼体动态变形规律作为已知条件,首先确定鱼游中体外作用(包括流体动力响应和惯性力),然后间接预测活体的体内作用(包括肌肉主动力和生物组织被动应力). 研究结果显示,鳗鲡鱼类尾部的肌肉强度明显低于其躯干部分,而鲹科鱼类尾部和躯干部分肌肉强度相当,这与各自的体内作用主导机制相适应. 且体外作用的区别也导致了鲹科鱼类的能效更高. 同时发现,两种模式游动下都存在鱼体躯干内部从头至尾的能量传递,其肌肉输出净功沿鱼体轴向都呈“钟形” 分布. 总体上沿轴向各点的肌肉都输出正的净功.
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
[本文引用: 2]
[本文引用: 3]
DOIURL [本文引用: 3]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}