 , 戴尔阜, 葛全胜
, 戴尔阜, 葛全胜Modelling the integrated effects of land use and climate change scenarios on forest aboveground biomass: A case study in Taihe County of China
WUZhuo, DAIErfu, GEQuansheng通讯作者:
收稿日期:2016-12-10
修回日期:2017-06-6
网络出版日期:2017-09-30
版权声明:2017《地理学报》编辑部本文是开放获取期刊文献,在以下情况下可以自由使用:学术研究、学术交流、科研教学等,但不允许用于商业目的.
基金资助:
作者简介:
-->
展开
摘要
关键词:
Abstract
Keywords:
-->0
PDF (5717KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
1 引言
土地利用和气候变化是土地科学、全球变化科学和全球生态学关注的关键问题[1-2],以土地利用和气候变化为特征的全球和区域环境变化已经给自然和社会系统带来了深刻影响[3]。作为陆地生态系统中重要的组成部分,森林不仅能够为人类提供大量的木材以及健康舒适的生存环境,而且在维持全球气候稳定、保护生物多样性等方面发挥着重要的作用。由于土地系统、气候系统和森林生态系统之间存在着复杂的相互影响、相互作用的反馈关系,因此,在这种多系统、多学科相互交叉的背景下,土地利用和气候变化将不可避免地对森林分布、结构、功能及演替过程产生重要影响[4]。传统的土地利用、气候变化以及森林的生态响应研究都是相对独立的,即研究不同时间或空间尺度上土地利用变化对森林的单独影响或气候变化对森林的单独影响[5-7]。然而随着研究的不断深入,这种将多要素分离的研究已经不能满足人类-自然系统耦合和多学科综合发展的需求[8],同时也无法满足政策制定者们进行自然资源与环境规划管理的总体需求[9]。因此,为了更加适应和减缓未来全球变化带来的影响,有必要探索和预测土地利用和气候变化给森林生态系统带来的综合影响。全球气候变化已经并将持续影响森林生态系统的多个方面,主要包括树种丰度、森林生产力、森林组成以及森林生物量等[10-13]。而其中森林生物量的变化是评估森林结构优劣和功能高低的最直接表现,并被广泛应用于森林与气候变化的影响与响应研究中[14-15]。从时间尺度上来看,气候变化对森林生态系统的影响是一个相对长期的过程,并存在一定的滞后性[16-17]。而土地利用变化,如建设用地扩张、森林采伐、农用地和林地之间的转换都将给森林生态系统带来更直接和更剧烈的影响[18]。由于气候系统、土地系统和森林生态系统之间存在复杂的相互作用关系,因此准确模拟和预测多系统中的综合影响难度很大[19]。近年来多模型耦合及综合利用为模拟和评估这种综合影响提供了一个新的思路。例如,全球土地计划(Global Land Project, GLP)中指出将土地利用和气候系统的变化与人类活动相结合,在不同空间尺度上(全球、区域/景观、地区)应用多主体土地利用模型和植被动态模型,定量模拟分析陆地生态系统的动态变化[7]。而欧洲陆地生态系统分析与模拟(ATEAM)计划则提出利用多种全球变化情景(气候、社会经济和土地利用)来评估和模拟不同区域、不同部门由全球变化导致的生态系统脆弱性,以实现土地利用和气候变化影响的综合评价[20]。目前,土地利用和气候变化对森林生态系统的影响研究在区域和景观尺度上取得了较好的进展。例如,Nepstad等利用气候模型、植被动态模型和经济预测模型模拟研究亚马逊地区森林、土地利用和气候之间的相互关系,通过定量化的模拟方法探求在未来不同干扰情景下森林衰退的临界点[21]。Thompson等利用森林景观模型LANDIS-II模拟了未来50年土地利用和气候变化对美国马萨诸塞州森林组成及森林生物量的影响,结果表明尽管未来气候变化会使森林生物量有所增加,但是土地利用变化尤其是林地转变为建设用地对森林的影响将更加明显,最终将使森林生物量显著下降[22]。以上这些研究主要集中在欧美地区,国内的研究更关注土地利用和气候变化对区域碳储量、生产力以及水文过程等方面的单独影响[23-25],对综合模拟的研究相对较少,因此有必要开展土地利用和气候变化对森林生态系统影响的综合研究。
中国南方红壤丘陵区是人工林分布的主要区域,其面积和蓄积量分别占全国人工林的63%和62%[26]。过去几十年,尽管南方人工林的快速增长已经为人类提供了大量的木材,但仍存在树种结构单一、生产力衰退、森林蓄积量低下等问题[27]。同时,随着中国城镇化进程的加快,人们对于土地资源的需求也日益增加,这些变化都将给区域森林管理和森林的可持续发展带来巨大压力[18]。本文选择中国南方人工林典型分布区江西省泰和县,开展区域土地利用和气候变化对森林生物量影响的综合研究,通过综合利用多主体土地利用模型(ABM/LUCC)、生态系统过程模型(PnET-II)以及森林景观动态模型(LANDIS-II)构建了一个综合模拟研究框架来模拟未来研究区不同干扰下森林地上总生物量的变化。由于土地利用变化受政策等影响较大,而土地政策又随着经济、人口、环境等多方面变化而变化,因此选择2010-2050年这一相对近期的时间段来模拟研究区土地利用变化,而该时间段也能够反映未来近期气候变化的一个整体趋势。此外,本文通过设计土地利用和气候变化(RCP 2.6、RCP 4.5和RCP 8.5)情景,并将这些情景进行组合来模拟土地利用和气候变化对森林生态系统的单独及综合影响。
2 数据与方法
2.1 研究区概况
江西省泰和县(26.45°~26.98°N, 114.95°~115.33°E)位于江西省中南部,是中国南方红壤丘陵区吉泰盆地的主要组成部分(图1)。全县总面积为2667 km2,属中亚热带季风气候,水热资源丰富,年均气温18.6 °C,一月平均气温为6.5 °C,七月平均气温为29.7 °C,年平均降水量为1370 mm,降水主要集中在夏季[28]。研究区主要土地利用类型包括林地、耕地、城镇用地、水域和其他用地。其中林地面积为1632 km2,占研究区总面积的61.2%,人工林的面积为733 km2,占森林总面积的44.9%,是中国南方人工林分布的典型区。研究区地带性植被为亚热带常绿阔叶林带,主要植被类型有常绿阔叶林、亚热带针叶林、亚热带针阔混交林、亚热带竹林等。主要的树种包括:马尾松(Pinus massoniana),湿地松(Pinus elliottii),杉木(Cunninghamia lanceolata),柏木(Cupressus funebris),樟树(Cinnamomum camphora),楠木(Phoebe zhennan),木荷(Schima superba),枫香(Liquidambar formosana),檫木(Sassafras tzumu),甜槠(Castanopsis eyrei),青冈(Cyclobalanopsis gracilis),拟赤扬(Alniphyllum fortunei),栲树(Castanopsis fargesii),桤木(Alnus cremastogyne),白栎(Quercus fabri),光皮桦(Betula luminifera),苦楝(Melia azedarach)和杨树(Populus deltoids)。此外,中国科学院生态系统研究网络(CERN)的千烟洲红壤丘陵综合开发试验站也位于泰和县灌溪镇(图1),生态站多年的观测数据为模型参数化和模型验证提供了科学的数据支持。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1研究区位置及土地利用类型
-->Fig. 1Location and land use types of Taihe County, Jiangxi Province, China
-->
2.2 气候变化数据
未来气候变化数据来自国家气候中心提供的“WCRP的耦合模式比较计划—阶段5的多模式数据(CMIP5数据)”(http://www.climatechange-data.cn)。该数据将21个CMIP5全球气候模式的模拟结果,经过插值计算将其统一降尺度到同一分辨率下(1°×1°),利用简单平均方法进行多模式耦合,得到2006-2100年的月平均资料。主要气候要素包括:月平均气温,月最高气温、月最低气温以及月降水量,受气候数据分辨率的限制将研究区的气候条件视为均质,反映的是区域未来气候变化的一个变化趋势。提取并处理了数据集中泰和县RCP 2.6、RCP 4.5以及RCP 8.5典型浓度路径排放情景下2010-2050年的未来气候数据。可以看出,到2050年泰和县年均温在不同排放情景下将增加1.09~1.73 °C(图2)。由于受亚热带季风气候的影响,预估的降水数据显示出明显的年际波动。此外,本文设计了一个控制情景,即假设未来气候不发生变化而保持当前的气候状态,该状态数据来自千烟洲生态试验站2001-2011年观测的气温、降水的平均值。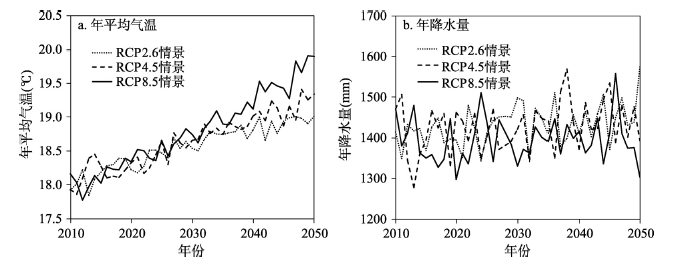 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2不同气候情景下江西省泰和县2010-2050年年均气温和年降水量变化趋势
-->Fig. 2Variation trend of annual mean temperature and precipitation in the study area under RCPs scenarios from 2010 to 2050
-->
2.3 土地利用变化模拟
为了明晰研究区过去土地利用变化特征,更好地模拟未来土地利用变化趋势,首先对研究区1990年、1995年、2000年、2005年以及2010年的土地利用数据进行分析,发现建设用地扩张、森林采伐和林地类型的转变是该区域土地利用变化对森林影响的3个关键过程。因此,土地利用变化的模拟主要关注与人类活动密切相关的建设用地和林地的变化。建设用地的模拟是基于Agent Analyst软件构建的多主体的土地利用模型—ABM/LUCC,该模型利用Not Quite Python(NQPy)编程语言在Recursive Porous Agent Simulation Toolkit(Repast)平台中进行程序编写,通过构建居民主体的行为规则及空间属性,实现“自下而上”的区域空间格局模拟。Agent Analyst是将多主体模型开发平台Repast和GIS有效结合的开源软件,在编程语言和模拟环境方面具有一定的优势,模型模拟流程如图3所示。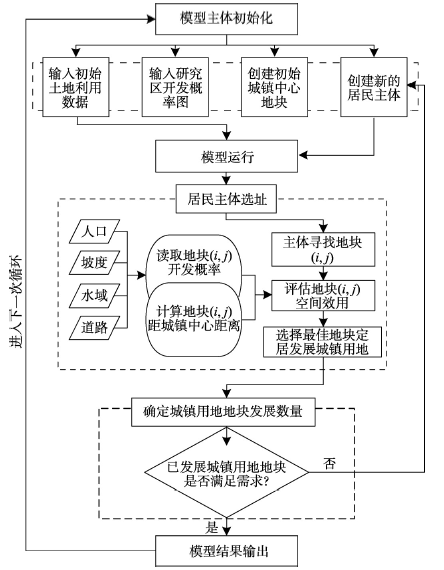 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图3基于主体的土地利用模型(ABM/LUCC)模拟流程
-->Fig. 3Flow chart of agent based model/land use and land cover change (ABM/LUCC)
-->
模型模拟初始化包括模型输入及主体的初始化。对于模型输入,土地利用图和地块开发概率图均采用100 m×100 m栅格数据,初始的输入数据选择2010年的土地利用数据,地块开发概率图由人口、坡度、水域和道路等要素综合计算得到。在输入所需的栅格数据后,居民主体开始进行选址。居民主体通过评估待开发地块的最大效用值来判断待选地块是否进行开发,在模型进行多次循环后最终形成建设用地的扩张格局。其中,最大效用值反映的是居民主体选择地块的主观意愿,是由待开发地块距城镇中心的距离、待开发地块的开发概率以及居民主体对距离和开发概率的偏好权重计算得到。而最大效应值估算指标的选择依据包括几个方面:① 澄江镇是泰和县的经济、政治、医疗和人口中心,具有较高的社会服务效应,居民主体在选择开发地块时会考虑距城镇中心区较近的地块,因此选择待开发地块距城镇中心距离(tcdist)作为估算指标来反映主体的选择意愿。② 通过评估研究区自然条件和人口数量来赋予每个待开发地块一个开发概率(devpro)。在本文中,建设用地的开发受一些自然要素的限制,如建设用地开发一般限制在坡度小于15°且用地类型为非水域的区域。同时受人类活动的影响,对主要道路以30 m做缓冲区,将该区域作为开发概率较高的区域。③ 人口的分布对开发概率也有重要影响,将不同镇级行政区划的人口总数进行标准化及空间化处理,将上述变量以相同权重叠加得到地块开发概率图。最后,假设主体具有相同的属性并对距离和开发概率具有同样的偏好,即居民主体对距离和开发概率的偏好权重相同,这种做法可以使模型代码更加简化并能够达到模型模拟的目的。最大效用值计算公式如下:
式中:devpro表示待开发地块的开发概率;tcdist表示待开发地块距城镇中心的距离;βd表示居民主体对城镇中心的最大偏好程度;βp表示待开发地块对开发概率的最大偏好程度,在本模型中,βd =βp =1;αp表示居民主体对待开发地块的偏好权重;αd表示主体对待开发地块距城镇中心的偏好权重。其中地块开发概率(devpro)计算公式如下:
式中:devpro代表每个待开发地块的开发概率;αp表示泰和县各镇人口数量;βs表示坡度。本文将待开发地块坡度限制为低于15°,当坡度大于15°时,待开发地块开发概率为0。γw表示水域,同样开发概率设为0;λr表示道路的缓冲区,该区域受人类活动的影响较大,开发概率设为0.8。开发概率取值区间为0~1,基于计算公式(2)得到的江西省泰和县开发概率如图4所示。
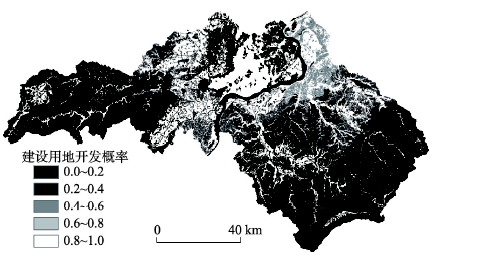 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图4江西省泰和县开发概率
-->Fig. 4Development probability maps in Taihe County, Jiangxi Province
-->
未来模拟中建设用地待开发的地块数量根据调研中收集到的《泰和县土地利用总体规划(2006-2020)》和《泰和县城市总体规划(2014-2030)》来确定。土地利用总体规划一定程度上反映了当地政府在近期内的土地利用变化政策,代表了政府对不同区域、不同土地利用类型变化的选择意愿,在未来土地利用变化的实施过程中具有指导与约束作用。尤其在中国,土地利用受政策的影响更加显著,政府在土地利用变化的过程中起主导作用,因此使用规划数据作为模拟基础数据能够较为真实地反映未来近期该地区土地利用变化的情况,具有一定的可靠性。2010年泰和县建设用地约为147.73 km2,根据研究区土地利用规划,到2020年建设用地预计增加到171.73 km2。因此,每年大约有240个地块被居民主体选择并转变为建设用地。在本文中,假设泰和县土地利用政策和建设用地发展速率保持当前情况不变,模拟2010-2050年泰和县建设用地的扩张格局。
对于森林采伐和林地类型转变,利用LANDIS-II模型中的Biomass Harvest模块和Land Use模块来模拟森林景观在采伐和土地利用干扰下的变化。Biomass Harvest模块能够模拟森林采伐所导致的林地变化。Land Use模块通过动态输入每十年的土地利用数据,能够模拟林地和非林地之间的地类的转变[29],输入的土地利用数据则来自于ABM/LUCC模型的模拟结果。通过将模型的两个模块整合可以得到每十年的林地变化以及相应的森林地上生物量的变化。LANDIS-II模拟中林地的采伐方式保持当前森林管理政策,即每十年采伐林地面积的10%。
2.4 森林地上生物量模拟
本文利用LANDIS-II和PnET-II模型来模拟森林地上生物量的变化。LANDIS-II模型是一个基于栅格的空间动态森林景观模型,它可以模拟森林干扰、演替以及森林管理过程[30]。模型通过输入物种生活史参数、物种建立系数(species establishment probability, SEP)、地表净初级生产力(aboveground net primary production, ANPP)以及生态分区数据来模拟森林景观动态变化,模拟的空间尺度较为灵活,可模拟104~107 ha 的森林景观变化[31]。同时,模型可以用来处理气候变化对森林的影响并广泛应用于森林与气候变化关系的研究中[32]。本文利用Biomass Succession模块、Biomass Harvest模块和Land Use模块来模拟土地利用和气候变化对森林地上生物量的影响。模型的输入主要包括空间数据和树种属性数据,空间数据包括初始群落图、生态分区图和土地利用图,树种属性数据主要包括物种生活史参数、ANPP和SEP。初始群落图来自江西省森林资源二类调查数据。生态分区图则根据研究区地形与海拔高度分为5个生态区,其中生态区1为非林地,生态区2~5分别表示低丘(0~100 m)、中丘(100~250 m)、高丘(250~500 m)和低山(> 500 m)(图5)。树种的物种生活史参数主要来自文献、样地调查以及咨询当地林业部门专家得到[33-34](表1)。Tab. 1
表1
表1研究区物种生活史参数
Tab. 1Species life history attributes in the study area
| 树种 | 拉丁名 | 寿命(年) | 成熟年龄(年) | 耐阴性 | 种子传播距离 | |
|---|---|---|---|---|---|---|
| 有效距离(m) | 最大距离(m) | |||||
| 杉木 | Cunninghamia lanceolata | 200 | 10 | 1 | 200 | 500 |
| 柏木 | Cupressus funebris | 500 | 35 | 2 | 70 | 200 |
| 马尾松 | Pinus massoniana | 200 | 10 | 1 | 200 | 500 |
| 湿地松 | Pinus elliottii | 200 | 10 | 1 | 200 | 500 |
| 木荷 | Schima superba | 300 | 20 | 5 | 20 | 200 |
| 樟树 | Cinnamomum camphora | 1000 | 15 | 4 | 50 | 120 |
| 楠木 | Phoebe zhennan | 1000 | 50 | 5 | 40 | 120 |
| 甜槠 | Castanopsis eyrei | 200 | 20 | 5 | 50 | 120 |
| 栲树 | Castanopsis fargesii | 150 | 30 | 5 | 60 | 250 |
| 白栎 | Quercus fabri | 120 | 15 | 4 | 20 | 200 |
| 青冈 | Cyclobalanopsis multinervis | 200 | 7 | 4 | 20 | 50 |
| 枫香 | Liquidambar formosana | 130 | 8 | 3 | 100 | 375 |
| 光皮桦 | Betula luminifera | 100 | 15 | 2 | 150 | 400 |
| 桤木 | Alnus cremastogyne | 125 | 5 | 3 | 15 | 60 |
| 拟赤杨 | Alniphyllum fortunei | 120 | 15 | 2 | 250 | 500 |
| 檫木 | Sassafras tzumu | 120 | 20 | 3 | 50 | 150 |
| 苦楝 | Melia azedarach | 80 | 5 | 2 | 200 | 400 |
| 杨树 | Populus deltoids | 90 | 10 | 2 | 150 | 500 |
新窗口打开
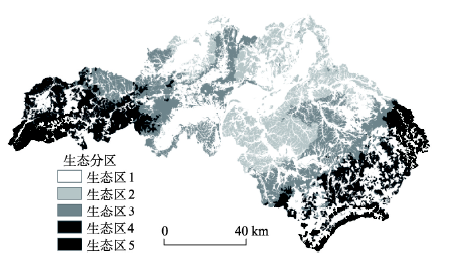 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图5江西省泰和县生态分区图
-->Fig. 5Ecoregions of Taihe County, Jiangxi Province
-->
每个树种的ANPP和SEP来自PnET-II模型的计算结果。PnET-II模型是一个模拟森林碳水平衡的生态系统过程模 型[35]。它通过输入气温、降水、光合有效辐射以及CO2浓度数据来模拟气候变化对树种及森林生态系统的影响,反映的是树种在一定立地条件下的生长状态,可适用于区域尺度的森林生态系统研究[36]。本文应用PnET-II模型来计算不同典型浓度路径(representative concentration pathways, RCPs)排放情景下泰和县18个主要树种的ANPP和SEP。模型输入重要参数包括田间持水能力(water holding capacity, WHC)、光合有效辐射(photosynthetically active radiation, PAR),数据来源于千烟洲生态试验站的观测数据。其他参数如叶氮浓度、光合作用最低温度及最适温度以及水分利用效率等来自于文献资料[37-40]。
2.5 综合模拟框架
为了研究土地利用和气候变化对森林地上生物量的综合影响,本文通过综合利用ABM/LUCC模型、PnET-II模型以及LANDIS-II模型,建立一个综合研究框架来模拟人类系统和气候系统的动态变化(图6)。首先,利用ABM/LUCC模型来模拟当前土地利用政策下的建设用地扩张,模型输出每十年的建设用地扩张图,同时该数据作为LANDIS-II模型Land Use模块的输入参数。其次,利用PnET-II模型来分别计算不同气候情景RCP 2.6、RCP 4.5、RCP 8.5以及当前气候情景下树种的ANPP及SEP,模型得到的结果用于LANIDS-II模型Biomass Succession模块的输入参数。最后,通过将土地利用和气候变化情景组合,利用LANDIS-II作为研究框架中的核心模型来模拟土地利用和气候变化综合影响下森林地上生物量的变化。该研究框架提供了一个多模型方法来帮助人们更好地理解复杂过程中的相互作用,同时,能够为人工林的结构和功能优化提供科学支持。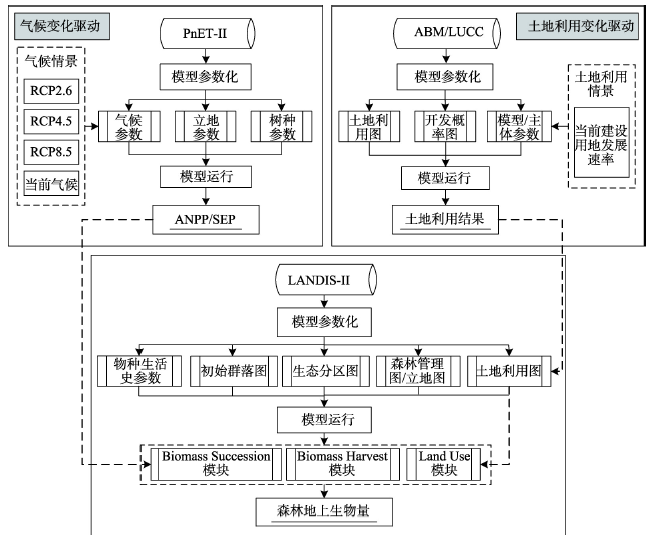 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图6土地利用和气候变化对森林地上生物量的综合模拟框架
-->Fig. 6The framework for the integrated effects of land use and climate change on forest AGB
-->
3 结果与分析
3.1 建设用地扩张模拟分析
本文土地利用变化模拟主要包括建设用地和林地的变化,而建设用地扩张的结果作为林地转变的模拟输入直接影响着林地类型的转变,进而影响整个景观的土地利用格局,因此重点分析建设用地扩张的模拟结果。研究区2010-2050年建设用地扩张的模拟结果如图7所示,结果表明2020年、2030年、2040年和2050年研究区建设用地的面积逐渐增加,分别占研究区总面积的6.4%、7.3%、8.2%和9.0%,到2050年建设用地达到240.55 km2。研究区的建设用地扩张格局表明,城镇发展是以当前城镇为基础逐渐向北部地区扩张,形成了以澄江镇为中心,以河流两岸为两翼的空间发展格局。建设用地土地利用变化的空间格局反映了居民主体的决策规则和主体行为,表明居民主体更偏好居住在距离城镇中心更近的区域,以享受更好的社会生活服务。2020年建设用地扩张的模拟结果与泰和县2020年土地利用规划图格局基本一致。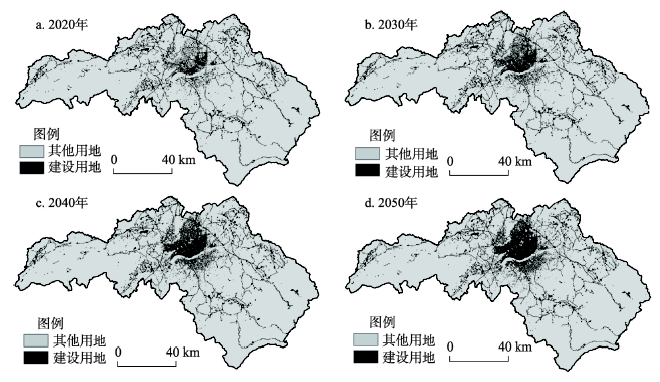 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图7ABM/LUCC模型模拟2010-2050年江西省泰和县建设用地变化结果
-->Fig. 7The results of town construction land change from the ABM/LUCC model during 2010-2050
-->
3.2 ANPP模拟分析
森林地上生物量的变化受树种ANPP、树种空间分布以及物种建群系数等多种要素的影响,其中ANPP的变化是影响森林生物量的最主要因子,直接反映了不同树种生物量的累积速率。在未来不同气候变化情景下,研究区18个树种的ANPP模拟结果表明2010-2050年所有树种的ANPP都呈现显著增加趋势,说明适当的升温对该区域树种的ANPP有积极的作用,其中常绿阔叶树种的ANPP显著高于针叶树种(图8)。在不同气候变化情景下,不同树种的ANPP结果显示出明显的差异,尤其对于一些阔叶树种,如木荷、甜槠及栲树随着模拟时间的增加ANPP的增幅越大。在RCP 2.6情景下,柏木的ANPP值最低为489 g m-2·a-1,甜槠的最高为1043 g m-2·a-1。到2050年,所有树种ANPP增加5.7%~12.5%。在RCP 4.5情景下,不同树种ANPP的范围为483~1097 g m-2·a-1,到2050年,ANPP增加了7.7%~23.4%。在RCP 8.5情景下,不同树种ANPP的范围为483~1143 g m-2·a-1,到2050年,ANPP增加了6.9%~26.5%。对比分析研究区的18个树种2010-2050年的ANPP变化量,结果表明ANPP增加较大的树种主要包括甜槠、苦楝、桤木、光皮桦、白栎及樟树,在RCP 2.6、RCP 4.5以及RCP 8.5情景下这些树种ANPP的增加程度也随着气温的增加而逐渐增大(图8)。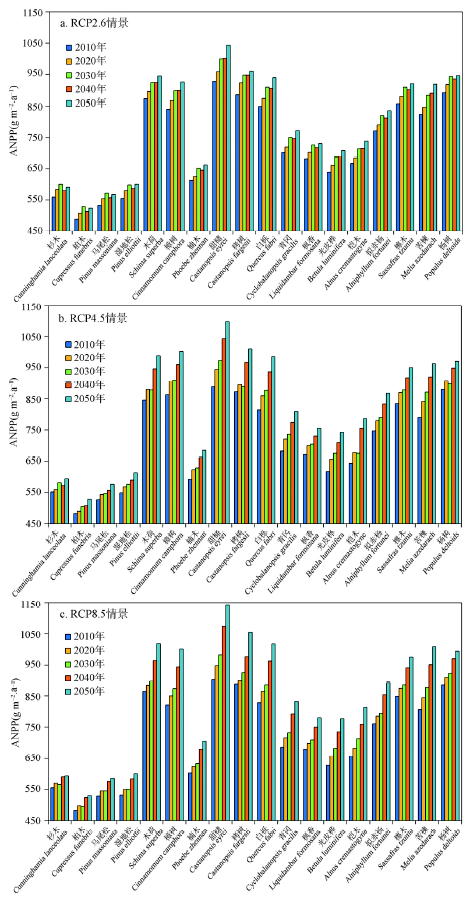 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图8PnET-II模型模拟的18个树种在不同气候情景下的ANPP结果
-->Fig. 8Aboveground net primary production (ANPP) for eighteen species simulated by PnET-II under RCPs scenarios
-->
3.3 森林地上生物量模拟分析
为了保证模型模拟结果的准确性,将LANDIS-II初始年的森林地上生物量模拟结果与野外观测数据进行比较。野外观测数据采用2010年研究区森林二类调查数据中林业小班活立木蓄积量数据,选择方精云等的生物量换算因子方法对森林地上生物量与蓄积量进行换算[41],计算研究区主要森林类型的地上生物量。在15707个森林小班中剔除非林地,经济林、竹林等非模拟小班后,随机选取500个小班的生物量的实测值与模型模拟结果进行比较(图9a),结果显示模拟值与实测值之间呈线性相关关系,且相关关系显著(R2 = 0.6463,p < 0.001)(图9b),说明该模型能够较好地模拟泰和县森林地上生物量,模拟结果可信。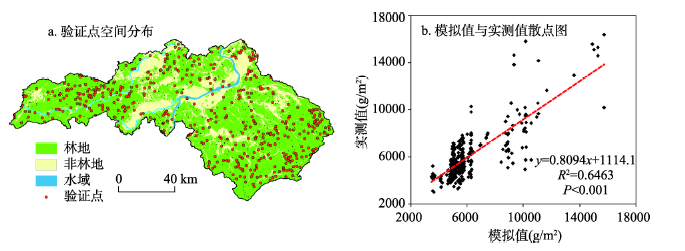 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图9江西省泰和县2010年森林地上生物量模拟值与实测值比较及验证点空间分布
-->Fig. 9Spatial distribution of the points for model validation and the comparison between simulated and investigated values of forest aboveground biomass
-->
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图10不同土地利用和气候变化情景组合下的森林地上生物量
-->Fig. 10Forest AGB of the total forest area under various scenarios
-->
模型模拟结果表明2010-2050年间,在不同气候情景下森林地上总生物量都有显著提升,而在有土地利用干扰加入的情景下森林地上总生物量结果显示先增高后保持相对稳定的趋势(图10)。在无干扰情景下,森林地上总生物量持续增加,从2010年的1055.17 t增加到2050年的1699.41 t,增加了61.06%;在气候变化的单独影响下,森林地上总生物量在不同气候变化情景下存在一定差异。到2050年研究区森林地上总生物量在RCP 2.6情景、RCP 4.5情景和RCP 8.5情景下分别增加了688.21 t、706.19 t和731.66 t,总生物量百分比分别增加了65.82%、68.91%和71.58%。在土地利用变化的单独影响下,研究区森林地上总生物量在2030年达到峰值,增加到1176.65 t,而到2050年则下降到1115.04 t。对于土地利用和气候变化的综合影响,模拟结果表明其变化趋势与单独的土地利用变化影响较为一致。不同气候变化情景与土地利用情景组合下,研究区森林地上总生物量的变化也显示出一定差异。到2050年,与单独的气候变化影响相比,加入土地利用变化干扰的森林地上总生物量在RCP 2.6情景、RCP 4.5情景、RCP 8.5情景下分别减少了33.13%、32.92%和32.42%。以上的结果表明土地利用变化在综合影响中占据主导作用,与气候变化相比土地利用变化对研究区森林生物量的影响更加显著。
4 结论与讨论
4.1 讨论
土地利用和气候变化对研究区森林地上生物量有显著影响。其中,土地利用变化,包括建设用地扩张、林地采伐在综合影响中占据主导作用,是综合模拟过程中的主要影响因子。这些结果与Thompson等和Gustafson等在全球其他地区的研究结果相一致[19, 22]。Thompson等发现尽管气候变化有助于提升美国马萨诸塞州森林的生长,但是土地利用变化的影响将远远抵消这种正效应[22]。Gustafson等同样发现西伯利亚中南部地区的原始森林受气候变化的影响远远不及受木材砍伐及其他人类活动干扰的影响[19]。本文利用ABM/LUCC模型来模拟建设用地扩张,该研究方法可以更加直观地反映主体选择地块的行为,帮助理解土地利用变化的人为驱动力。由ANPP的结果发现森林生产力随着气温、降水的增加而增加,该结果与其他人在亚热带区域的结果相一致[42]。当然在现实的生态系统中其他环境要素的变化也可能导致ANPP的变化,例如大气组成中CO2、O3以及N含量的变化都可能影响森林生产力[43-45],而这些会在未来的研究中逐渐补充完善。在森林地上生物量的模拟中,生物量的累积是以生态系统过程速率为基础,通过计算不同干扰下不同树种年龄级的地上生物量得到[46-47]。研究结果发现,在不考虑土地利用变化的影响下,生物量的累积与气温存在一个正相关的关系,该研究也与吴丹等人在泰和县的研究相一致[48]。而在土地利用变化的干扰下,森林地上生物量结果呈现先增加然后迅速下降的变化趋势。森林地上生物量下降的最主要原因是建设用地扩张和森林采伐导致的林地利用类型转换和树木移除。与气候变化影响的相对滞后性相比,这些与人类紧密相关的土地利用变化将给森林地上生物量带来更直接更快速的影响。土地、气候和森林之间存在着相互影响、相互制约的复杂关系,这种关系涉及到土地系统、气候系统以及森林生态系统这种多系统相互交叉的信息、能量与物质传递。由人类活动引起的土地系统的变化会通过林地迁移、森林采伐以及林地保护与恢复等方式改变森林景观结构[18]。气候系统的变化也与人类活动密切相关,同时气候和森林之间也存在着相互影响、相互反馈的作用机制[3]。受土地利用和气候变化的影响,森林生态系统的空间相互作用、森林生态系统演替、干扰、迁移和转换过程同时作用于个体、种群、群落及生态系统的多个尺度中[1]。因此研究土地利用和气候变化对森林景观尺度上综合影响及长期响应必然存在着一些不确定性。首先,数据及参数化过程的不确定。对于气候数据,RCPs气候数据集是来自多个全球气候模式平均后的结果,尽管比选用单独的气候模型更具有说服力,但仍然具有很大的不确定性[49],而气候数据的空间分辨率也是限制本研究的重要因素,将气候数据进行降尺度处理也是下一步研究需要关注的重点。对于PnET-II和LANDIS-II模型参数化,虽然数据采用的是基于森林调查和样地观测数据,模型也通过了随机数据验证,但参数化过程仍存在着不确定性。其次,模拟的限制性来自模型本身。本文应用较为简化的ABM/LUCC模型,ABM是基于复杂性理论通过模拟微观主体的行为过程来表达系统宏观层面的作用结果,反映多系统间的相互作用过程[50]。在模拟过程中,主体受环境要素的影响来改变其行为规则,而人作为土地利用等活动的主要行动者则是模拟结果产生不确定性的主要来源。受环境及其他主体行为的影响,人类活动对土地利用方式选择机制会随自然条件和不同主体之间关系而发生变化。而在本文中并未考虑自然要素和主体属性在未来模拟中动态变化,这必然也会给模拟结果带来一定的不确定性。当然,这种模型设计的简化为计算机建模也带来了一定的便利,能够更加快速的实现研究目标。对于PnET-II和LANDIS-II模型也是复杂生态系统过程的简化,它们更加关注景观尺度上的关键过程。如LANDIS-II模型模拟树种年龄级的动态变化,而不是模拟每一棵树的变化,反映的是大尺度长时间的森林景观动态[47]。最后,综合利用多模型模拟土地利用和气候变化的综合影响是基于一些合理的假设。例如,假设在模拟过程中物种的生活史参数和地形、土壤以及植物物候特征不随时间的变化而变化;假设不同模型在模拟过程中不受模型空间尺度效应的影响;同时,假设未来的土地利用变化不受气候变化的影响,这些假设都是为了更好地实现本文的目标。综上所述,尽管模拟综合影响存在一些不确定性和限制性,但是这些都是计算机建模与复杂系统之间的权衡,本文的研究目的不是为了准确地预测森林地上生物量,而是力求模拟在多因素干扰下森林景观的变化趋势,是综合模拟研究的一个新的探索。
未来的土地利用变化和气候变化可能将阻碍森林管理目标的实现。因此,将土地利用利用变化和气候变化的适应性理论与森林管理实践相结合将变得十分必要。本文基于模拟结果提出一些适应性建议,以保证区域的森林可持续经营与管理。首先,在土地利用规划中,应明确规划生态红线,确保林地面积不发生明显减少;其次,在保证木材砍伐量的前提下选择适当的树种进行人工林的种植,提高森林质量。例如,一些速生阔叶树种,如木荷、桤木可以适当加入到特定区域的人工纯林中来提升应对气候变化的脆弱性。最后,林业政策的制定应与当地的土地利用需求相结合,做到不断更新、不断发展。在中国尤其是南方的森林分布区,公有林是森林资源的主要组成部分,它们受林业政策的影响强烈。中央和地方政府应因地制宜地鼓励并引导人工林的管理,充分发挥森林生态系统服务效益,最终实现森林的可持续发展。
4.2 结论
本文综合利用基于主体的土地利用模型(ABM/LUCC)、生态系统过程模型(PnET-II)以及森林景观动态模型(LANDIS-II)并构建了综合模拟研究框架。利用该框架模拟了土地利用和气候变化情景下江西省泰和县2010-2050年森林地上生物量的变化,对比分析了不同组合情景下森林地上生物量的变化差异。主要结论如下:(1)2010-2050年,泰和县的建设用地发展逐渐由城镇中心向北部地区扩张,形成了以澄江镇为中心,以河流两岸为两翼的空间发展格局。多主体土地利用模型能够很好地反映居民主体的选择意愿,更好地阐释土地利用格局变化的内部驱动力,模拟结果对未来的城镇扩张模式具有一定的借鉴意义。
(2)在未来不同气候情景下,研究区所有树种的ANPP都呈现显著增加趋势,说明适当的升温对该区域树种的ANPP有积极的作用,但不同树种对不同气候情景的响应程度有所差异。
(3)研究区未来土地利用变化(建设用地扩张、森林采伐和林地转换)对泰和县森林地上总生物量的影响比气候变化所带来的影响更加显著。在综合模拟中,研究区森林地上总生物量在有土地利用变化干扰的情景下与RCP 2.6、RCP 4.5和RCP 8.5单独的气候情景下相比分别减少33.13%、32.92%和32.42%。尽管未来气候变化将有利于森林地上总生物量的增加,但是土地利用变化将使森林地上总生物量显著减少,其影响将抵消气候变化带来的正效应。
(4)本文提出的综合模拟研究框架可以很好地模拟土地利用和气候变化对森林生态系统的综合影响,为提升和优化人工林结构和功能、开展可持续森林管理提供科学建议。
The authors have declared that no competing interests exist.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . |
| [2] | . Global change will alter the supply of ecosystem services that are vital for human well-being. To investigate ecosystem service supply during the 21st century, we used a range of ecosystem models and scenarios of climate and land-use change to conduct a Europe-wide assessment. Large changes in climate and land use typically resulted in large changes in ecosystem service supply. Some of these trends may be positive (for example, increases in forest area and productivity) or offer opportunities (for example, "surplus land" for agricultural extensification and bioenergy production). However, many changes increase vulnerability as a result of a decreasing supply of ecosystem services (for example, declining soil fertility, declining water availability, increasing risk of forest fires), especially in the Mediterranean and mountain regions. |
| [3] | . |
| [4] | . |
| [5] | . |
| [6] | . Contemporary land-use pressures have a significant impact on the extent and condition of forests in the eastern United States, causing a regional-scale decline in forest cover. Earlier in the 20th century, land cover was on a trajectory of forest expansion that followed agricultural abandonment. However, the potential for forest regeneration has slowed, and the extent of regional forest cover has declined by more than 4.0%. Using remote-sensing data, statistical sampling, and change-detection methods, this research shows how land conversion varies spatially and temporally across the East from 1973鈥2000, and how those changes affect regional land-change dynamics. The analysis shows that agricultural land use has continued to decline, and that this enables forest recovery; however, an important land-cover transition has occurred, from a mode of regional forest-cover gain to one of forest-cover loss caused by timber cutting cycles, urbanization, and other land-use demands. |
| [7] | . |
| [8] | . |
| [9] | . |
| [10] | . |
| [11] | . Abstract Climate change is driving latitudinal and altitudinal shifts in species distribution worldwide, leading to novel species assemblages. Lags between these biotic responses and contemporary climate changes have been reported for plants and animals. Theoretically, the magnitude of these lags should be greatest in lowland areas, where the velocity of climate change is expected to be much greater than that in highland areas. We compared temperature trends to temperatures reconstructed from plant assemblages (observed in 76,634 surveys) over a 44-year period in France (1965-2008). Here we report that forest plant communities had responded to 0.54 C of the effective increase of 1.07 C in highland areas (500-2,600 m above sea level), while they had responded to only 0.02 C of the 1.11 C warming trend in lowland areas. There was a larger temperature lag (by 3.1 times) between the climate and plant community composition in lowland forests than in highland forests. The explanation of such disparity lies in the following properties of lowland, as compared to highland, forests: the higher proportion of species with greater ability for local persistence as the climate warms, the reduced opportunity for short-distance escapes, and the greater habitat fragmentation. Although mountains are currently considered to be among the ecosystems most threatened by climate change (owing to mountaintop extinction), the current inertia of plant communities in lowland forests should also be noted, as it could lead to lowland biotic attrition. 漏2011 Macmillan Publishers Limited. All rights reserved |
| [12] | . Abstract Variation in terrestrial net primary production (NPP) with climate is thought to originate from a direct influence of temperature and precipitation on plant metabolism. However, variation in NPP may also result from an indirect influence of climate by means of plant age, stand biomass, growing season length and local adaptation. To identify the relative importance of direct and indirect climate effects, we extend metabolic scaling theory to link hypothesized climate influences with NPP, and assess hypothesized relationships using a global compilation of ecosystem woody plant biomass and production data. Notably, age and biomass explained most of the variation in production whereas temperature and precipitation explained almost none, suggesting that climate indirectly (not directly) influences production. Furthermore, our theory shows that variation in NPP is characterized by a common scaling relationship, suggesting that global change models can incorporate the mechanisms governing this relationship to improve predictions of future ecosystem function. |
| [13] | . Abstract In the past three decades, our global climate has been experiencing unprecedented warming. This warming has and will continue to significantly influence the structure and function of forest ecosystems. While studies have been conducted to explore the possible responses of forest landscapes to future climate change, the representative concentration pathways (RCPs) scenarios under the framework of the Coupled Model Intercomparison Project Phase 5 (CMIP5) have not been widely used in quantitative modeling research of forest landscapes. We used LANDIS-II, a forest dynamic landscape model, coupled with a forest ecosystem process model (PnET-II), to simulate spatial interactions and ecological succession processes under RCP scenarios, RCP2.6, RCP4.5 and RCP8.5, respectively. We also modeled a control scenario of extrapolating current climate conditions to examine changes in distribution and aboveground biomass (AGB) among five different forest types for the period of 2010 2100 in Taihe County in southern China, where subtropical coniferous plantations dominate. The results of the simulation show that climate change will significantly influence forest distribution and AGB. (i) Evergreen broad-leaved forests will expand into Chinese fir and Chinese weeping cypress forests. The area percentages of evergreen broad-leaved forests under RCP2.6, RCP4.5, RCP8.5 and the control scenarios account for 18.25%, 18.71%, 18.85% and 17.46% of total forest area, respectively. (ii) The total AGB under RCP4.5 will reach its highest level by the year 2100. Compared with the control scenarios, the total AGB under RCP2.6, RCP4.5 and RCP8.5 increases by 24.1%, 64.2% and 29.8%, respectively. (iii) The forest total AGB increases rapidly at first and then decreases slowly on the temporal dimension. (iv) Even though the fluctuation patterns of total AGB will remain consistent under various future climatic scenarios, there will be certain responsive differences among various forest types. |
| [14] | . Tropical forest conversion, shiftingcultivation and clearing of secondary vegetation makesignificant contributions to global emissions ofgreenhouse gases today, and have the potential forlarge additional emissions in future decades. Globally, an estimated 3.1×10 9 t of biomasscarbon of these types is exposed to burning annually,of which 1.1×10 9 t is emitted to the atmospherethrough combustion and 49×10 6 t is converted tocharcoal (including 26–31×10 6 t C of blackcarbon). The amount of biomass exposed to burningincludes aboveground remains that failed to burn ordecompose from clearing in previous years, andtherefore exceeds the 1.9×10 9 t of abovegroundbiomass carbon cleared on average each year. Above-and belowground carbon emitted annually throughdecomposition processes totals 2.1×10 9 t C. Atotal gross emission (including decomposition ofunburned aboveground biomass and of belowgroundbiomass) of 3.41×10 9 t C year -1 resultsfrom clearing primary (nonfallow) and secondary(fallow) vegetation in the tropics. Adjustment fortrace gas emissions using IPCC Second AssessmentReport 100-year integration global warming potentialsmakes this equivalent to 3.39×10 9 t ofCO 2 -equivalent carbon under a low trace gasscenario and 3.83×10 9 t under a high trace gasscenario. Of these totals, 1.06×10 9 t (31%)is the result of biomass burning under the low tracegas scenario and 1.50×10 9 t (39%) under thehigh trace gas scenario. The net emissions from allclearing of natural vegetation and of secondaryforests (including both biomass and soil fluxes) is2.0×10 9 t C, equivalent to 2.0–2.4×10 9 t of CO 2 -equivalent carbon. Adding emissions of0.4×10 9 t C from land-use category changesother than deforestation brings the total for land-usechange (not considering uptake of intact forest,recurrent burning of savannas or fires in intactforests) to 2.4×10 9 t C, equivalent to 2.4–2.9×10 9 t of CO 2 -equivalent carbon. The totalnet emission of carbon from the tropical land usesconsidered here (2.4×10 9 t C year -1 )calculated for the 1981–1990 period is 50% higherthan the 1.6×10 9 t C year -1 value used by the Intergovernmental Panel on Climate Change. The inferred (= `missing') sink in the global carbonbudget is larger than previously thought. However,about half of the additional source suggested here maybe offset by a possible sink in uptake by Amazonianforests. Both alterations indicate that continueddeforestation would produce greater impact on globalcarbon emissions. The total net emission of carboncalculated here indicates a major global warmingimpact from tropical land uses, equivalent toapproximately 29% of the total anthropogenic emissionfrom fossil fuels and land-use change. |
| [15] | . The 90,674 wildland fires that burned 2.9 million ha at an estimated suppression cost of $1.6 billion in the United States during the 2000 fire season demonstrated that forest fuel loading has become a hazard to life, property, and ecosystem health as a result of past fire exclusion policies and practices. The fire regime at any given location in these regions is a result of complex interactions between forest biomass, topography, ignitions, and weather. Forest structure and biomass are important aspects in determining current and future fire regimes. Efforts to quantify live and dead forest biomass at the local to regional scale has been hindered by the uncertainty surrounding the measurement and modeling of forest ecosystem processes and fluxes. The interaction of elevated CO 2 with climate, soil nutrients, and other forest management factors that affect forest growth and fuel loading will play a major role in determining future forest stand growth and the distribution of species across the southern United States. The use of satellite image analysis has been tested for timely and accurate measurement of spatially explicit land use change and is well suited for use in inventory and monitoring of forest carbon. The incorporation of Landsat Thematic Mapper data coupled with a physiologically based productivity model (PnET), soil water holding capacity, and historic and projected climatic data provides an opportunity to enhance field plot based forest inventory and monitoring methodologies. We use periodic forest inventory data from the USDA Forest Service's Forest Inventory and Analysis (FIA) project to obtain estimates of forest area and type to generate estimates of carbon storage for evergreen, deciduous, and mixed forest classes for use in an assessment of remotely sensed forest cover at the regional scale for the southern United States. The displays of net primary productivity (NPP) generated from the PnET model show areas of high and low forest carbon storage potential and their spatial relationship to other landscape features for the southern United States. At the regional scale, predicted annual NPP in 1992 ranged from 836 to 2181 g/m 2 /year for evergreen forests and 769 2634 g/m 2 /year for deciduous forests with a regional mean for all forest land of 1448 g/m 2 /year. Prediction of annual NPP in 2050 ranged from 913 to 2076 g/m 2 /year for evergreen forest types to 1214 2376 g/m 2 /year for deciduous forest types with a regional mean for all forest land of 1659 g/m 2 /year. The changes in forest productivity from 1992 to 2050 are shown to display potential areas of increased or decreased forest biomass. This methodology addresses the need for spatially quantifying forest carbon in the terrestrial biosphere to assess forest productivity and wildland fire fuels. |
| [16] | . KNOWLEDGE of the vegetation response to climate change is necessary to assess and predict realistic ecosystem development in the anticipated, CO 2 -induced warmer world, particularly at high latitudes where greater warming is expected 1–3 . Reconstruction of vegetation development over the past 1,000 years may be helpful in this respect, because this period was characterized by contrasting climatic conditions 4–9 . Here we report the reconstruction of wind-exposed, tree-line vegetation associated with long-term climate change in northern Canada, using tree-ring and growth-form analyses of spruce subfossils. Three major types of growth form within the exposed, but stable, lichen–spruce community successively predominated in response to climate forcing: high krummholz (dwarf spruce, 2–3 m high) and high krummholz (AD 1435–1570, warm period) and low krummholz ( 6750 cm) (little ice age to present: AD 1570 onwards, cold period and present climate, respectively). Whereas the expansion of a marginal lichen–spruce woodland climaxed during the late Middle Ages (AD 1435–1570), present development of a low-krummholz vegetation at these sites seems to be out of phase with the twentieth century warming. This suggests that ecosystem recovery to global warming is not straightforward, depending on the nature of vegetation structure present at the time climate change occurred. The implications of such ecosystem resilience for the detection and monitoring of the expected CO 2 -induced warming is discussed, particularly for the climate-sensitive arctic and subarctic regions. |
| [17] | . Abstract 6168 Data from 13 long-term (>1 yr), field-based studies of the effects of elevated CO 2 concentration ([CO 2 ]) on European forest tree species were analysed using meta-analysis and modelling. Meta-analysis was used to determine mean responses across the data sets, and data were fitted to two commonly used models of stomatal conductance in order to explore response to environmental conditions and the relationship with assimilation. 6168 Meta-analysis indicated a significant decrease (21%) in stomatal conductance in response to growth in elevated [CO 2 ] across all studies. The response to [CO 2 ] was significantly stronger in young trees than old trees, in deciduous compared to coniferous trees, and in water stressed compared to nutrient stressed trees. No evidence of acclimation of stomatal conductance to elevated [CO 2 ] was found. 6168 Fits of data to the first model showed that growth in elevated [CO 2 ] did not alter the response of stomatal conductance to vapour pressure deficit, soil water content or atmospheric [CO 2 ]. Fits of data to the second model indicated that conductance and assimilation responded in parallel to elevated [CO 2 ] except when water was limiting. 6168 Data were compared to a previous meta-analysis and it was found that the response of g s to elevated [CO 2 ] was much more consistent in long-term (>1yr) studies, emphasising the need for long-term elevated [CO 2 ] studies. By interpreting data in terms of models, the synthesis will aid future modelling studies of responses of forest trees to elevated [CO 2 ]. |
| [18] | . Places experience forest transitions when declines in forest cover cease and recoveries in forest cover begin. Forest transitions have occurred in two, sometimes overlapping circumstances. In some places economic development has created enough non-farm jobs to pull farmers off of the land, thereby inducing the spontaneous regeneration of forests in old fields. In other places a scarcity of forest products has prompted governments and landowners to plant trees in some fields. The transitions do little to conserve biodiversity, but they do sequester carbon and conserve soil, so governments should place a high priority on promoting them. |
| [19] | . Multiple global changes such as timber harvesting in areas not previously disturbed by cutting and climate change will undoubtedly affect the composition and spatial distribution of boreal forests, which will, in turn, affect the ability of these forests to retain carbon and maintain biodiversity. To predict future states of the boreal forest reliably, it is necessary to understand the complex interactions among forest regenerative processes (succession), natural disturbances (e.g., fire, wind, and insects), and anthropogenic disturbances (e.g., timber harvest). We used a landscape succession and disturbance model (LANDIS-II) to study the relative effects of climate change, timber harvesting, and insect outbreaks on forest composition, biomass (carbon), and landscape pattern in south-central Siberia. We found that most response variables were more strongly influenced by timber harvest and insect outbreaks than by the direct effects of climate change. Direct climate effects generally increased tree productivity and modified probability of establishment, but indirect effects on the fire regime generally counteracted the direct effects of climate on forest composition. Harvest and insects significantly changed forest composition, reduced living aboveground biomass, and increased forest fragmentation. We concluded that: (1) Global change is likely to significantly change forest composition of south-central Siberian landscapes, with some changes taking ecosystems outside the historic range of variability. (2) The direct effects of climate change in the study area are not as significant as the exploitation of virgin forest by timber harvest and the potential increased outbreaks of the Siberian silk moth. (3) Novel disturbance by timber harvest and insect outbreaks may greatly reduce the aboveground living biomass of Siberian forests and may significantly alter ecosystem dynamics and wildlife populations by increasing forest fragmentation. |
| [20] | . |
| [21] | . Some model experiments predict a large-scale substitution of Amazon forest by savannah-like vegetation by the end of the twenty-first century. Expanding global demands for biofuels and grains, positive feedbacks in the Amazon forest fire regime and drought may drive a faster process of forest degradation that could lead to a near-term forest dieback. Rising worldwide demands for biofuel and meat are creating powerful new incentives for agro-industrial expansion into Amazon forest regions. Forest fires, drought and logging increase susceptibility to further burning while deforestation and smoke can inhibit rainfall, exacerbating fire risk. If sea surface temperature anomalies (such as El Ni o episodes) and associated Amazon droughts of the last decade continue into the future, approximately 55% of the forests of the Amazon will be cleared, logged, damaged by drought or burned over the next 20 years, emitting 15-26Pg of carbon to the atmosphere. Several important trends could prevent a near-term dieback. As fire-sensitive investments accumulate in the landscape, property holders use less fire and invest more in fire control. Commodity markets are demanding higher environmental performance from farmers and cattle ranchers. Protected areas have been established in the pathway of expanding agricultural frontiers. Finally, emerging carbon market incentives for reductions in deforestation could support these trends. |
| [22] | . Land use and climate change have complex and interacting effects on naturally dynamic forest landscapes. To anticipate and adapt to these changes, it is necessary to understand their individual and aggregate impacts on forest growth and composition. We conducted a simulation experiment to evaluate regional forest change in Massachusetts, USA over the next 50 years (2010-2060). Our objective was to estimate, assuming a linear continuation of recent trends, the relative and interactive influence of continued growth and succession, climate change, forest conversion to developed uses, and timber harvest on live aboveground biomass (AGB) and tree species composition. We examined 20 years of land use records in relation to social and biophysical explanatory variables and used regression trees to create "probability-of-conversion" and "probability-of-harvest" zones. We incorporated this information into a spatially interactive forest landscape simulator to examine forest dynamics as they were affected by land use and climate change. We conducted simulations in a full-factorial design and found that continued forest growth and succession had the largest effect on AGB, increasing stores from 181.83 Tg to 309.56 Tg over 50 years. The increase varied from 49% to 112% depending on the ecoregion within the state. Compared to simulations with no climate or land use, forest conversion reduced gains in AGB by 23.18 Tg (or 18%) over 50 years. Timber harvests reduced gains in AGB by 5.23 Tg (4%). Climate change (temperature and precipitation) increased gains in AGB by 17.3 Tg (13.5%). Pinus strobus and Acer rubrum were ranked first and second, respectively, in terms of total AGB throughout all simulations. Climate change reinforced the dominance of those two species. Timber harvest reduced Quercus rubra from 10.8% to 9.4% of total AGB, but otherwise had little effect on composition. Forest conversion was generally indiscriminate in terms of species removal. Under the naive assumption that future land use patterns will resemble the recent past, we conclude that continued forest growth and recovery will be the dominant mechanism driving forest dynamics over the next 50 years, and that while climate change may enhance growth rates, this will be more than offset by land use, primarily forest conversion to developed uses. |
| [23] | . . |
| [24] | . . |
| [25] | . 为定量分析潮河流域土地利用和气候变化对流域径流变化的影响,应用SWAT模型对流域上游至下游的大阁、戴营和下会3个水文站径流进行模拟,采用情景法分析径流对土地利用和气候变化的响应。在模型校准期和验证期采用两个参数:<em>p</em>因子和<em>r</em>因子来评价模拟的拟合度及不确定性。结果表明,3个水文站在校准期和验证期的<em>p</em>因子值分别为:0.70和0.77,0.87和0.82,0.92和0.78,<em>r</em>因子值分别为0.63和0.90,0.97和0.79,0.88和0.92,评价整个流域模拟有效性的模型目标函数<em>g</em>最佳值为0.66,说明该模型对潮河流域的产水量模拟具有很好的适用性。以1981-1990年为基准期,1991-2000年流域土地利用变化造成年径流量减少了4.10 mm,而气候变化导致年径流增加了29.68 mm;2001-2009年土地利用变化造成年径流量减少2.98 mm,气候变化造成年径流量减少了14.30 mm。与1999年土地利用条件模拟径流值相比,几种极端情景法模拟分析结果表明: 灌木林地情景下年径流增加了158.2%,草地情景下年径流增加了4.1%,林地和耕地情景下年径流分别减少23.7% 和41.7%;不同气候变异情景模拟结果显示,径流对降水的变化敏感性高于对温度变化的敏感性,降水每增加10%,径流平均增加23.9%。温度每增加12%,径流平均减少6%。 因此,在气候变化背景下,优化土地利用结构与方式是实现流域水资源科学管理的途径之一。 . 为定量分析潮河流域土地利用和气候变化对流域径流变化的影响,应用SWAT模型对流域上游至下游的大阁、戴营和下会3个水文站径流进行模拟,采用情景法分析径流对土地利用和气候变化的响应。在模型校准期和验证期采用两个参数:<em>p</em>因子和<em>r</em>因子来评价模拟的拟合度及不确定性。结果表明,3个水文站在校准期和验证期的<em>p</em>因子值分别为:0.70和0.77,0.87和0.82,0.92和0.78,<em>r</em>因子值分别为0.63和0.90,0.97和0.79,0.88和0.92,评价整个流域模拟有效性的模型目标函数<em>g</em>最佳值为0.66,说明该模型对潮河流域的产水量模拟具有很好的适用性。以1981-1990年为基准期,1991-2000年流域土地利用变化造成年径流量减少了4.10 mm,而气候变化导致年径流增加了29.68 mm;2001-2009年土地利用变化造成年径流量减少2.98 mm,气候变化造成年径流量减少了14.30 mm。与1999年土地利用条件模拟径流值相比,几种极端情景法模拟分析结果表明: 灌木林地情景下年径流增加了158.2%,草地情景下年径流增加了4.1%,林地和耕地情景下年径流分别减少23.7% 和41.7%;不同气候变异情景模拟结果显示,径流对降水的变化敏感性高于对温度变化的敏感性,降水每增加10%,径流平均增加23.9%。温度每增加12%,径流平均减少6%。 因此,在气候变化背景下,优化土地利用结构与方式是实现流域水资源科学管理的途径之一。 |
| [26] | . Abstract Background Planted forests are expanding throughout the world, and now account for 7% of global forest cover and provide more than 60% of global industrial round wood. Negative ecological and social impacts of the establishment of planted forests and the challenges of their multi-purpose management have also given rise to concern. China has been playing an important role in global expansion of planted forests while reducing emission from deforestation and forest degradation. Methods This article attempts to conduct an overall analysis and review of the current status, challenges and future perspectives of planted forests in China to obtain a better understanding on how to manage planted forests for multiple uses under a changing environment. Data from several national forest inventories and other sources, as well as new empirical data, were used for a statistical analysis on the dynamics of planted forests in China. Results Planted forests in China have undergone a continuous expansion in the past 20 years, which has significantly contributed to an increase in total forest cover and timber supply as well as other ecosystem services like carbon sequestration. The three key driving forces for this expansion were government programmes, and market and technology development. However, the predominance of very few tree species in the plantations, uneven spatial distribution, skewed age-class distribution, and low volumes in growing stock, coupled with increasing complexity of multiple purpose forestry management under a changing environment, have generated several major challenges confronting planted forests in China. Conclusions A strategic transition in the management of Chinese planted forests is needed, with a shifting emphasis from area expansion to stand productivity and quality enhancement, from traditional timber production to multi-purpose management for forest goods and services, and from monoculture plantations to biodiversity rich mixed forests. A landscape-design approach and adaptive management practices should be put in place to meet the diversified demands of stakeholders for different goods and ecosystem services while enhancing forest resilience under the changing climate. |
| [27] | |
| [28] | . Despite the significance of interannual variation of soil respiration (R(S)) for understanding long-term soil carbon dynamics, factors that control the interannual variation of R(S) have not been sufficiently investigated. Interannual variation of R(S) was studied using a 6-year data set collected in a subtropical plantation dominated by an exotic species, slash pine (Pinus elliottii Engelm.), in China. The results showed that seasonal variation of R(S) was significantly affected by soil temperature and soil water content (SWC). R(S) in the dry season (July October) was constrained by seasonal drought. Mean annual R(S) was estimated to be 736 +/- 30 g C.m(-2).year(-1), with a range of 706-790 g C.m(-2).year(-1). Although this forest was characterized by a humid climate with high precipitation (1469 mm.year(-1)), the interannual variation of R(S) was attributed to the changes of annual mean SWC (R(2) = 0.66, P = 0.03), which was affected by annual rainfall frequency (R(2) = 0.80, P < 0.01) and not rainfall amount (P = 0.84). Consequently, precipitation pattern indirectly controlled the interannual variation of R(S) by affecting soil moisture in this subtropical forest. In the context of climate change, interannual variation of R(S) in subtropical ecosystems is expected to increase because of the predicted changes of precipitation regime. |
| [29] | . Forest landscape models (FLMs) are widely used to examine the influence of disturbances on long-term and broad-scale forest ecosystem dynamics. However, FLMs are not well-suited to simulating some types of management or disturbance regimes, including land-use change. Consequently, there are situations in which a researcher may wish to estimate the timing and location of events externally, either from a different model, empirical observations, or some other source, and then incorporate them into an FLM. We present Land Use Plus (LU+), an extension for the LANDIS-II FLM that allows users to integrate externally-developed, spatially and temporally explicit representations of land use or other disturbance into simulations. LU+ allows users to model the proximate effects of these events on forest composition and biomass, as well as subsequent dynamics, including tree establishment and the potential for future management. LU+ will significantly increase the breadth of research questions for which LANDIS-II may be appropriately used. |
| [30] | . We introduce LANDIS-II, a landscape model designed to simulate forest succession and disturbances. LANDIS-II builds upon and preserves the functionality of previous LANDIS forest landscape simulation models. LANDIS-II is distinguished by the inclusion of variable time steps for different ecological processes; our use of a rigorous development and testing process used by software engineers; and an emphasis on collaborative features including a flexible, open architecture. We detail the variable time step logic and provide an overview of the system architecture. Finally, we demonstrate model behavior and sensitivity to variable time steps through application to a large boreal forest landscape. We simulated pre-industrial forest fire regimes in order to establish base-line conditions for future management. Differing model time steps substantially altered our estimates of pre-industrial forest conditions. Where disturbance frequency is relatively high or successional processes long, the variable time steps may be a critical element for successful forest landscape modeling. |
| [31] | |
| [32] | . |
| [33] | . 1991~1992年在贵州黎平、锦屏、天柱杉木种子园对园内花粉密度、树冠各部位胚珠接受花粉的数量和花粉飞散距离作了观测,结果表明,在树冠范围内,上部的花粉接收量大于下部,不同高度和方位花粉接收量的差异还受林分密度和地形的影响;在同一林分中,疏林地段的花粉云密度大于密林地段;杉木花粉传播的有效距离因年份和方位而异,顺主风方向超过600m,逆主风方向在200~400m间。 . 1991~1992年在贵州黎平、锦屏、天柱杉木种子园对园内花粉密度、树冠各部位胚珠接受花粉的数量和花粉飞散距离作了观测,结果表明,在树冠范围内,上部的花粉接收量大于下部,不同高度和方位花粉接收量的差异还受林分密度和地形的影响;在同一林分中,疏林地段的花粉云密度大于密林地段;杉木花粉传播的有效距离因年份和方位而异,顺主风方向超过600m,逆主风方向在200~400m间。 |
| [34] | |
| [35] | . PnET is a simple, lumped-parameter, monthlytime-step model of carbon and water balances of forests built on two principal relationships: 1) maximum photosynthetic rate is a function of foliar nitrogen concentration, and 2) stomatal conductance is a function of realized photosynthetic rate. Monthyly leaf area display and carbon and water balances are predicted by combining these with standard equations describing light attenuation in canopies and photosynthetic response to diminishing radiation intensity, along with effects of soil water stress and vapor pressure deficit (VPD). PnET has been validated against field data from 10 well-studied temperate and boreal forest ecosystems, supporting our central hypothesis that aggregation of climatic data to the monthly scale and biological data such as foliar characteristics to the ecosystem level does not cause a significant loss of information relative to long-term, mean ecosystem responses. Sensitivity analyses reveal a diversity of responses among systems to identical alterations in climatic drivers. This suggests that great care should be used in developing generalizations as to how forests will respond to a changing climate. Also critical is the degree to which the temperature responses of photosynthesis and respiration might acclimate to changes in mean temperatures at decadal time scales. An extreme climate change simulation (+3 C maximum temperature, 25% precipitation with no change in minimum temperature or radiation, direct effects of increased atmospheric CO 2 ignored) suggests that major increases in water stress, and reductions in biomass production (net carbon gain) and water yield would follow such a change. |
| [36] | . Abstract Many studies have been conducted to quantify the possible ecosystem/landscape response to the anticipated global warming. However, there is a large amount of uncertainty in the future climate predictions used for these studies. Specifically, the climate predictions can be very different based on a variety of global climate models and alternative greenhouse emission scenarios. In this study, we coupled a forest landscape model, LANDIS-II, and a forest process model, PnET-II, to examine the uncertainty (that results from the uncertainty in the future climate predictions) in the forest-type composition prediction for a transitional forest landscape [the Boundary Water Canoe Area]. Using an improved global-sensitivity analysis technique [Fourier amplitude sensitivity test], we also quantified the amount of uncertainty in the forest-type composition prediction contributed by different climate variables including temperature, CO 2 , precipitation and photosynthetic active radiation (PAR). The forest landscape response was simulated for the period 2000 2400 ad based on the differential responses of 13 tree species under an ensemble of 27 possible climate prediction profiles (monthly time series of climate variables). Our simulations indicate that the uncertainty in the forest-type composition becomes very high after 2200 ad , which is close to the time when the current forests are largely removed by windthrow disturbances and natural mortality. The most important source of uncertainty in the forest-type composition prediction is from the uncertainty in temperature predictions. The second most important source is PAR, the third is CO 2 and the least important is precipitation. Our results also show that if the optimum photosynthetic temperature rises due to CO 2 enrichment, the forest landscape response to climatic change measured by forest-type composition may be substantially reduced. |
| [37] | . Canopy foliar Nitrogen Concentration(CNC) is one of the most important parameters influencing vegetation productivity in forest ecosystems. In this study, we explored the potential of imaging spectrometry(hyperspectral) remote sensing of CNC in conifer plantations in China's subtropical red soil hilly region. Our analysis included data from 57 field plots scattered across two transects covered by Hyperion images. Single regression and partial least squares regression(PLSR) were used to explore the relationships between CNC and hyperspectral data. The correlations between CNC and nearinfrared reflectance(NIR) were consistent in three data subsets(subsets A鈥揅). For all subsets, CNC was significantly positively correlated with NIR in the two transects(R2=0.29, 0.33 and 0.36, P0.05 or P0.01, respectively). It suggested that the NIR-CNC relationship exist despite a weak one, and the relationship may be weakened by the single canopy structure. Besides, we also applied a shortwave infrared(SWIR) index Normalized Difference Nitrogen Index(NDNI) to estimate CNC variation. NDNI presented a significant positive correlation with CNC in different subsets, but like NIR, it was also with low coefficient of determination(R2=0.38, 0.20 and 0.17, P0.01, respectively). Also, the correlations between CNC and the entire spectrum reflectance(or its derivative and logarithmic transformation) by PLSR owned different significance in various subsets. We did not find the very robust relationship like previous literatures, so the data we used were checked again. The paired T-test was applied to estimate the influence of inter-annual variability of FNC on the relationships between CNC and Hyperion data. The inter-annual mismatch between period of fieldwork and Hyperion acquisition had no influence on the correlations of CNC-Hyperion data. Meanwhile, we pointed out that the lack of the canopy structure variation in conifer plantation area may lead to these weak relationships. |
| [38] | |
| [39] | |
| [40] | . From July 2008 to August 2008, 72 leaf samples from 22 species and 81 soil samples in the nine natural forest ecosystems were collected, from north to south along the North-South Transect of Eastern China (NSTEC). Based on these samples, we studied the geographical distribution patterns of vegetable water use efficiency (WUE) and nitrogen use efficiency (NUE), and analyzed their relationship with environmental factors. The vegetable WUE and NUE were calculated through the measurement of foliar 13C and C/N of predominant species, respectively. The results showed: (1) vegetable WUE, ranging from 2.13 to 28.67 mg C g-1 H2O, increased linearly from south to north in the representative forest ecosystems along the NSTEC, while vegetable NUE showed an opposite trend, increasing from north to south, ranging from 12.92 to 29.60 g C g-1 N. (2) Vegetable WUE and NUE were dominantly driven by climate and significantly affected by soil nutrient factors. Based on multiple stepwise regression analysis, mean annual temperature, soil phosphorus concentration, and soil nitrogen concentration were responding for 75.5% of the variations of WUE (p0.001). While, mean annual precipitation and soil phosphorus concentration could explain 65.7% of the change in vegetable NUE (p0.001). Moreover, vegetable WUE and NUE would also be seriously influenced by atmospheric nitrogen deposition in nitrogen saturated ecosystems. (3) There was a significant trade-off relationship between vegetable WUE and NUE in the typical forest ecosystems along the NSTEC (p0.001), indicating a balanced strategy for vegetation in resource utilization in natural forest ecosystems along the NSTEC. This study suggests that global change would impact the resource use efficiency of forest ecosystems. However, vegetation could adapt to those changes by increasing the use efficiency of shortage resource while decreasing the relatively ample one. But extreme impacts, such as heavy nitrogen deposition, would break this trade-off mechanism and give a dramatic disturbance to the ecosystem biogeochemical cycle. |
| [41] | . The location and mechanisms responsible for the carbon sink in northern mid-latitude lands are uncertain. Here, we used an improved estimation method of forest biomass and a 50-year national forest resource inventory in China to estimate changes in the storage of living biomass between 1949 and 1998. Our results suggest that Chinese forests released about 0.68 petagram of carbon between 1949 and 1980, for an annual emission rate of 0.022 petagram of carbon. Carbon storage increased significantly after the late 1970s from 4.38 to 4.75 petagram of carbon by 1998, for a mean accumulation rate of 0.021 petagram of carbon per year, mainly due to forest expansion and regrowth. Since the mid-1970s, planted forests (afforestation and reforestation) have sequestered 0.45 petagram of carbon, and their average carbon density increased from 15.3 to 31.1 megagrams per hectare, while natural forests have lost an additional 0.14 petagram of carbon, suggesting that carbon sequestration through forest management practices addressed in the Kyoto Protocol could help offset industrial carbon dioxide emissions. |
| [42] | . <FONT face=Verdana>利用基于生理生态学过程的EALCO模型,探讨了千烟洲中亚热带人工针叶林生态系统对未来气候变化的响应.结果表明:CO<SUB>2</SUB>浓度、温度和降水的变化对该人工林生态系统碳水通量<BR>影响的程度不同,其中CO<SUB>2</SUB>浓度>温度>降水.CO<SUB>2</SUB>浓度是生态系统总光合生产力(GPP)<BR>的主要驱动因子,温度与CO<SUB>2</SUB>浓度均是控制生态系统呼吸的主要环境因子,温度的升高使<BR>植物地上部分呼吸明显增加,而CO<SUB>2</SUB>浓度升高则对土壤呼吸影响较大.温度升高使蒸散(ET)<BR>增加,而CO<SUB>2</SUB>浓度升高则使ET减少.在未来气候变化情景(2100年)下,该人工林生态系统的<BR>净初级生产力将增加22%,说明其仍具有较强的固碳潜力.<BR></FONT> . <FONT face=Verdana>利用基于生理生态学过程的EALCO模型,探讨了千烟洲中亚热带人工针叶林生态系统对未来气候变化的响应.结果表明:CO<SUB>2</SUB>浓度、温度和降水的变化对该人工林生态系统碳水通量<BR>影响的程度不同,其中CO<SUB>2</SUB>浓度>温度>降水.CO<SUB>2</SUB>浓度是生态系统总光合生产力(GPP)<BR>的主要驱动因子,温度与CO<SUB>2</SUB>浓度均是控制生态系统呼吸的主要环境因子,温度的升高使<BR>植物地上部分呼吸明显增加,而CO<SUB>2</SUB>浓度升高则对土壤呼吸影响较大.温度升高使蒸散(ET)<BR>增加,而CO<SUB>2</SUB>浓度升高则使ET减少.在未来气候变化情景(2100年)下,该人工林生态系统的<BR>净初级生产力将增加22%,说明其仍具有较强的固碳潜力.<BR></FONT> |
| [43] | . How forests will respond to rising [CO2] in the long term is uncertain, most studies having involved juvenile trees in chambers prior to canopy closure. Poplar free-air CO2 enrichment (Viterbo, Italy) is one of the first experiments to grow a forest from planting through canopy closure to coppice, entirely under open-air conditions using free-air CO2 enrichment technology. Three Populus species: P. alba, P. nigra and P. x euramericana, were grown in three blocks, each containing one control and one treatment plot in which CO2 was elevated to the expected 2050 concentration of 550 ppm. The objective of this study was to estimate gross primary production (GPP) from recorded leaf photosynthetic properties, leaf area index (LAI) and meteorological conditions over the complete 3-year rotation cycle. From the meteorological conditions recorded at 30 min intervals and biweekly measurements of LAI, the microclimate of leaves within the plots was estimated with a radiation transfer and energy balance model. This information was in turn used as input into a canopy microclimate model to determine light and temperature of different leaf classes at 30 min intervals which in turn was used with the steady-state biochemical model of leaf photosynthesis to compute CO2 uptake by the different leaf classes. The parameters of these models were derived from measurements made at regular intervals throughout the coppice cycle. The photosynthetic rates for different leaf classes were summed to obtain canopy photosynthesis, i.e. GPP. The model was run for each species in each plot, so that differences in GPP between species and treatments could be tested statistically. Significant stimulation of GPP driven by elevated [CO2] occurred in all 3 years, and was greatest in the first year (223-251%), but markedly lower in the second (19-24%) and third years (5-19%). Increase in GPP in elevated relative to control plots was highest for P. nigra in 1999 and for P. x euramericana in 2000 and 2001, a |
| [44] | Under the site conditions of a temperate forest, the exchange of short-chained oxygenated carbonyls (aldehydes, ketones) was assessed from leaves of adult European beech trees. The crowns of the trees were either exposed to an elevated O3 regime as released by a free-air fumigation system (2 x O3) or to the unchanged O3 regime at the site (1 x O3, 'control'). Daily fluctuations of oxygenated carbonyls were quantified in relation to environmental and physiological factors. In particular, the effect of O3 on carbonyl exchange was studied. Measurements of leaf gas exchange were performed with a dynamic cuvette system, and carbonyl fluxes were determined using 2,4-dinitrophenylhydrazine (DNPH)-coated silica gel cartridges. Leaves mainly emitted acetaldehyde, formaldehyde and acetone. Acetaldehyde dominated the emissions, amounting up to 100 nmol m-2 min-1, followed by formaldehyde (approximately 80 nmol m-2 min-1) and acetone (approximately 60 nmol m-2 min-1). Carbonyl emissions were highest during midday and significantly lowered at night, irrespective of the O3 exposure regime. Trees exposed to 2 x O3 emitted acetaldehyde and acetone at enhanced rates. The findings are of particular significance for future climate change scenarios that assume increased O3 levels. |
| [45] | . Phosphorus (P) often limits plant growth in subtropical forests. Many studies have focused on plant P responses to rising atmospheric carbon dioxide (CO 2 ) and increasing nitrogen (N) deposition; however, the response of soil P availability to these anthropogenic changes remains unclear. We used open-top chambers to study the changes in soil P fractions in model subtropical forests after 5 years of exposure to elevated CO 2 and N addition. Soils studied here were highly weathered and P-deficient. Elevated CO 2 increased plant available P in soils as well as P input from litter. The increase in soil P availability was related to the decreased recalcitrant P under elevated CO 2 . Soil P availability was not reduced by N addition due to the replenishment of P during litter decomposition. N addition led to a small increase in the most readily desorbed inorganic P. N addition with elevated CO 2 depleted recalcitrant P. Our results suggest that elevated CO 2 and N addition could maintain or even increase soil P availability. |
| [46] | . Predicting the long-term dynamics of forest systems depends on understanding multiple processes that often operate at vastly different scales. Disturbance and seed dispersal are landscape-scale phenomena and are spatially linked across the landscape. Ecosystem processes (e.g., growth and decomposition) have high annual and inter-specific variation and are generally quantified at the scale of a forest stand. To link these widely scaled processes, we used biomass (living and dead) as an integrating variable that provides feedbacks between disturbance and ecosystem processes and feedbacks among multiple disturbances. We integrated a simple model of biomass growth, mortality, and decay into LANDIS, a spatially dynamic landscape simulation model. The new biomass module was statically linked to PnET-II, a generalized ecosystem process model. The combined model simulates disturbances (fire, wind, harvesting), dispersal, forest biomass growth and mortality, and inter- and intra-specific competition. We used the model to quantify how fire and windthrow alter forest succession, living biomass and dead biomass across an artificial landscape representative of northern Wisconsin, USA. In addition, model validation and a sensitivity analysis were conducted. |
| [47] | . We introduce LANDIS-II, a landscape model designed to simulate forest succession and disturbances. LANDIS-II builds upon and preserves the functionality of previous LANDIS forest landscape simulation models. LANDIS-II is distinguished by the inclusion of variable time steps for different ecological processes; our use of a rigorous development and testing process used by software engineers; and an emphasis on collaborative features including a flexible, open architecture. We detail the variable time step logic and provide an overview of the system architecture. Finally, we demonstrate model behavior and sensitivity to variable time steps through application to a large boreal forest landscape. We simulated pre-industrial forest fire regimes in order to establish base-line conditions for future management. Differing model time steps substantially altered our estimates of pre-industrial forest conditions. Where disturbance frequency is relatively high or successional processes long, the variable time steps may be a critical element for successful forest landscape modeling. |
| [48] | . <p>根据第6次森林清查小班数据,运用生物量转换因子法和平均生物量法估算了2003年江西省泰和县森林植被的生物量和碳储量,采用空间替代时间的方法,利用Logistic方程拟合了泰和主要森林类型年龄与碳密度的曲线关系,并结合小班轮伐信息,估算了全县1985—2003年的植被生物量和碳储量,分析了期间的时空动态特征,并以2003年为基准年,假定到2020、2030年泰和县森林植被面积保持稳定、且不考虑轮伐期,推算了此情景下2020、2030年泰和县植被碳储量.结果表明:2003年,泰和县森林林分总面积15.74×10<sup>4</sup> hm<sup>2</sup>,总生物量6.71 Tg,植被碳储量4.14 Tg C,平均碳密度26.31 t C·hm<sup>- 2</sup>. 1985、1994、2003、2020、2030年泰和县森林植被碳储量分别为1.06、2.83、4.14、5.65和6.35Tg C,森林植被碳密度的空间分布由东西部向中部递减.人工造林使泰和县林分面积大幅增加,全县森林植被的固碳能力明显增强.</p> . <p>根据第6次森林清查小班数据,运用生物量转换因子法和平均生物量法估算了2003年江西省泰和县森林植被的生物量和碳储量,采用空间替代时间的方法,利用Logistic方程拟合了泰和主要森林类型年龄与碳密度的曲线关系,并结合小班轮伐信息,估算了全县1985—2003年的植被生物量和碳储量,分析了期间的时空动态特征,并以2003年为基准年,假定到2020、2030年泰和县森林植被面积保持稳定、且不考虑轮伐期,推算了此情景下2020、2030年泰和县植被碳储量.结果表明:2003年,泰和县森林林分总面积15.74×10<sup>4</sup> hm<sup>2</sup>,总生物量6.71 Tg,植被碳储量4.14 Tg C,平均碳密度26.31 t C·hm<sup>- 2</sup>. 1985、1994、2003、2020、2030年泰和县森林植被碳储量分别为1.06、2.83、4.14、5.65和6.35Tg C,森林植被碳密度的空间分布由东西部向中部递减.人工造林使泰和县林分面积大幅增加,全县森林植被的固碳能力明显增强.</p> |
| [49] | |
| [50] | . 基于复杂系统理论与Agent模型,分别从理论与方法的角度梳理并总结了当前土地变化科学的研究进展。复杂系统理论为研究土地变化所处的"耦合的人类—自然系统"及其复杂性问题提供理论支撑;而基于Agent的模拟作为研究复杂系统的重要工具,为传统土地变化模拟提供新的方法支持。当前,ABMs与土地变化模型相结合(ABM/LUCC),无论是模型构建还是模型应用均取得了显著进展。然而,其也存在诸多问题,尤其是理论与应用脱节,使很多建模者容易陷入"为建模而建模"的误区。此外,现有的ABM模型设计较为随意,模型间差异太大,不利于跨区域比较研究的开展。因此,虽然ABM是认识复杂系统的有效方法,但是建模之前离不开对系统复杂性的深入研究;基于Agent的土地变化模拟的意义在于充分表达"人类—自然"系统的综合复杂关系,从而合理的预测土地变化(结构与功能)的动态过程,解释并评价土地变化的可能影响。 . 基于复杂系统理论与Agent模型,分别从理论与方法的角度梳理并总结了当前土地变化科学的研究进展。复杂系统理论为研究土地变化所处的"耦合的人类—自然系统"及其复杂性问题提供理论支撑;而基于Agent的模拟作为研究复杂系统的重要工具,为传统土地变化模拟提供新的方法支持。当前,ABMs与土地变化模型相结合(ABM/LUCC),无论是模型构建还是模型应用均取得了显著进展。然而,其也存在诸多问题,尤其是理论与应用脱节,使很多建模者容易陷入"为建模而建模"的误区。此外,现有的ABM模型设计较为随意,模型间差异太大,不利于跨区域比较研究的开展。因此,虽然ABM是认识复杂系统的有效方法,但是建模之前离不开对系统复杂性的深入研究;基于Agent的土地变化模拟的意义在于充分表达"人类—自然"系统的综合复杂关系,从而合理的预测土地变化(结构与功能)的动态过程,解释并评价土地变化的可能影响。 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}