Behavioral analyses in mouse models of autism spectrum disorders
Jianfei Wang, Junhai Han, Zichao Zhang,The Key Laboratory of Developmental Genes and Human Disease, School of Life Science and Technology, Southeast University, Nanjing 210096, China
Supported by the National Natural Science Foundation of China Nos.31871374 Supported by the National Natural Science Foundation of China Nos.81730034 the Natural Science Foundation of Jiangsu Province No.BK20170080
作者简介 About authors 王剑飞,在读硕士研究生,专业方向:孤独症致病机制。E-mail:w13912971049@163.com。
Abstract Autism spectrum disorder is a group of genetically-related developmental disorders of the nervous system. Patients mainly present with core symptoms such as social behavior defects, repetitive stereotyped behaviors, and learning and memory disorders. The mouse models are critical for the studies of the pathogenic mechanisms and potential therapeutic strategies of autism spectrum disorder. The assessments of mouse behaviors provide understandings of the effects of different genetic manipulations as well as pathogenic mechanisms of these diseases. This article describes various mouse behavioral assays corresponding to the core symptoms of ASD patients and provides a detailed description of protocols, cautions, and data analysis for those assays, thereby helping researchers to establish their own experimental designs. In addition, behavioral phenotypes of currently known ASD mouse models are summarized to provide a reference for researchers in the field. Keywords:autism spectrum disorder;mouse behavioral analysis;social disorder;repetitive behavior;learning and memory;anxiety
PDF (954KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 王剑飞, 韩俊海, 张子超. 孤独症谱系障碍小鼠模型行为学检测方法. 遗传[J], 2021, 43(5): 501-519 doi:10.16288/j.yczz.20-391 Jianfei Wang. Behavioral analyses in mouse models of autism spectrum disorders. Hereditas(Beijing)[J], 2021, 43(5): 501-519 doi:10.16288/j.yczz.20-391
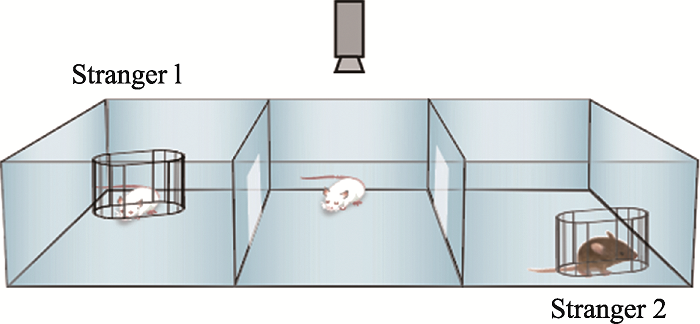
三厢社交实验(three-chamber social test):2004年Crawley等[7]根据小鼠的群居性和社会性以及形成一定社交记忆的原理,开发出自动化的三厢社交实验,可以直接自动化的测量小鼠社交行为,并对实验结果进行标准化和高通量的分析。三厢社交实验的基础是正常小鼠具有社交能力,当面对同类时,会更愿意和同类小鼠接触,而不是单处一室;当出现陌生的同类时,正常小鼠能把它和熟悉的同伴区分开来,更多的和陌生个体接触,表现出社交新颖性。
旷场实验(open field test):将动物置于一个被周围墙壁阻止逃跑的未知环境,是一种被广泛应用的类焦虑行为模型,起初为了评估动物情绪而开发,随后也被Archer等[26,27]用于检测动物的行为运动能力。旷场实验中,小鼠具有畏惧空旷场地而进行躲避的天性,而另一方面又会对空旷中心产生探究的好奇心理,两者冲突从而引发焦虑。
A:行为箱;B:行为箱俯视观测图(中间蓝色圆圈是人为划定的中央区,红色曲线为小鼠运动轨迹)。 Fig. 6Open field test
1.11 高架十字迷宫实验
高架十字迷宫实验(the elevated plus maze test):小鼠具有恐高的天性,对高处天然趋避,但对新环境又有探索的好奇心。1958年Montgomery等发现暴露在高架的迷宫通道中所引发的这种探究-趋避冲突,比暴露在封闭的旷场中所引发的焦虑冲突要强烈得多,基于此设计出开放和封闭区域不均一的高架Y-迷宫实验;1985年Pellow等[28]利用等长的开放和封闭臂优化成高架十字迷宫检测小鼠的焦虑行为。
水迷宫实验(Morris water maze test):小鼠天生厌水,在水中会选择最佳的路线逃离水环境。根据此特性1981年Morris通过强迫实验动物游泳设计出水迷宫实验,并迅速成为评估空间记忆和空间导航的标准任务。通过观测小鼠在逃亡过程中是否能利用周围环境进行定位寻找隐藏在水中的逃生平台来检验小鼠的空间参考记忆能力。小鼠搜寻并登上平台所用的时间越短,小鼠的学习和记忆能力也就越强[31]。
MattilaML, KielinenM, LinnaSL, JussilaK, EbelingH, BloiguR, JosephRM, MoilanenI . Autism spectrum disorders according to DSM-IV-TR and comparison with DSM-5 draft criteria: an epidemiological study J Am Acad Child Adolesc Psychiatry, 2011,50(6):583-592. [本文引用: 1]
SatterstromFK, KosmickiJA, WangJ, BreenMS, De RubeisS, AnJY, PengM, CollinsR, GroveJ, KleiL, StevensC, ReichertJ, MulhernMS, ArtomovM, GergesS, SheppardB, XuX, BhaduriA, NormanU, BrandH, SchwartzG, NguyenR, GuerreroEE, DiasC, BetancurC, CookEH, GallagherL, GillM, SutcliffeJS, ThurmA, ZwickME, BorglumAD, StateMW, CicekAE, TalkowskiME, CutlerDJ, DevlinB, SandersSJ, RoederK, DalyMJ, BuxbaumJD . Large-scale exome sequencing study implicates both developmental and functional changes in the neurobiology of autism Cell, 2020,180(3):568-584. [本文引用: 1]
BergR . Autism--an environmental health issue after all? J Environ Health, 2009,71(10):14-18. [本文引用: 1]
ZhouRY, DangWL, ZhouZ, LiHW, ZhangX . Advances in research of animal models of autism spectrum disorders Acta Lab Anim Sci Sin, 2019,27(3):380-386 [本文引用: 1]
CrawleyJN . Mouse behavioral assays relevant to the symptoms of autism Brain Pathol, 2007,17(4):448-459. [本文引用: 1]
HollisF, KabbajM . Social defeat as an animal model for depression Ilar J, 2014,55(2):221-232. [本文引用: 1]
OreficeLL, ZimmermanAL, ChirilaAM, SlebodaSJ, HeadJP, GintyDD . Peripheral mechanosensory neuron dysfunction underlies tactile and behavioral deficits in mouse models of ASDs Cell, 2016,166(2):299-313. [本文引用: 2]
DeaconRM . Assessing nest building in mice Nat Protoc, 2006,1(3):1117-1119. [本文引用: 1]
RavizzaSM, SolomonM, IvryRB, CarterCS . Restricted and repetitive behaviors in autism spectrum disorders: the relationship of attention and motor deficits Dev Psychopathol, 2013,25(3):773-784. [本文引用: 1]
PremoliM, MemoM, BoniniSA . Ultrasonic vocalizations in mice: relevance for ethologic and neurodevelopmental disorders studies Neural Regen Res, 2021,16(6):1158-1167. [本文引用: 10]
PankseppJB, LahvisGP . Social reward among juvenile mice Genes Brain Behav, 2007,6(7):661-671. [本文引用: 1]
EgnorSR, SeagravesKM . The contribution of ultrasonic vocalizations to mouse courtship Curr Opin Neurobiol, 2016,38:1-5. [本文引用: 1]
Pe?agarikanoO, LázaroMT, LuXH, GordonA, DongH, LamHA, PelesE, MaidmentNT, MurphyNP, YangXW, GolshaniP, GeschwindDH . Exogenous and evoked oxytocin restores social behavior in the Cntnap2 mouse model of autism Sci Transl Med, 2015,7(271):271r-278r. [本文引用: 1]
ZalaSM, ReitschmidtD, NollA, BalazsP, PennDJ . Sex-dependent modulation of ultrasonic vocalizations in house mice (Mus musculus musculus) PLoS One, 2017,12(12):e188647. [本文引用: 1]
YanT, HeB, WanS, XuM, YangH, XiaoF, BiK, JiaY . Antidepressant-like effects and cognitive enhancement of Schisandra chinensis in chronic unpredictable mild stress mice and its related mechanism Sci Rep, 2017,7(1):6903. [本文引用: 1]
RuM, LiuH . Association between Y-maze acquisition learning and major histocompatibility complex class ii polymorphisms in mice Biomed Res Int, 2018,2018:6381932. [本文引用: 1]
BerridgeKC, AldridgeJW, HouchardKR, ZhuangXX . Sequential super-stereotypy of an instinctive fixed action pattern in hyper-dopaminergic mutant mice: a model of obsessive compulsive disorder and Tourette's Bmc Biol, 2005,3:4. [本文引用: 1]
BurstenSN, BerridgeKC, OwingsDH . Do California ground squirrels (Spermophilus beecheyi) use ritualized syntactic cephalocaudal grooming as an agonistic signal? J Comp Psychol, 2000,114(3):281-290. [本文引用: 1]
KannerL . The conception of wholes and parts in early infantile autism Am J Psychiatry, 1951,108(1):23-26. [本文引用: 1]
EllegoodJ, CrawleyJN . Behavioral and neuroanatomical phenotypes in mouse models of autism Neurotherapeutics, 2015,12(3):521-533. [本文引用: 1]
SimmonsDH, TitleyHK, HanselC, MasonP. Behavioral tests for mouse models of autism: an argument for the inclusion of cerebellum-controlled motor behaviors Neuroscience, 2020, S0306-4522(20):30304. [本文引用: 1]
ThomasA, BurantA, BuiN, GrahamD, Yuva-PaylorLA, PaylorR . Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety Psychopharmacology (Berl), 2009,204(2):361-373. [本文引用: 1]
RodgersRJ . Animal models of 'anxiety': where next? Behav Pharmacol, 1997, 8(6-7): 477-496, 497-504. [本文引用: 1]
PellowS, ChopinP, FileSE, BrileyM . Validation of open: closed arm entries in an elevated plus-maze as a measure of anxiety in the rat J Neurosci Methods, 1985,14(3):149-167. [本文引用: 1]
TakaoK, MiyakawaT . Light/dark transition test for mice J Vis Exp, 2006, ( 1):104. [本文引用: 1]
EnnaceurA, DelacourJ . A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data Behav Brain Res, 1988,31(1):47-59. [本文引用: 1]
TomasPI, BurwellRD . Using the spatial learning index to evaluate performance on the water maze Behav Neurosci, 2015,129(4):533-539. [本文引用: 1]
WangSS, YanY, ZhaoYS . Interventions for social skills in autism spectrum disorders: a meta-analysis of effects and influencing factors Chin J Spec Edu, 2019, ( 10):43-51. [本文引用: 1]
NaviauxJC, WangL, LiK, BrightAT, AlaynickWA, WilliamsKR, PowellSB, NaviauxRK . Antipurinergic therapy corrects the autism-like features in the Fragile X (Fmr1 knockout) mouse model Mol Autism, 2015,6:1. [本文引用: 1]
HanS, TaiC, WestenbroekRE, YuFH, CheahCS, PotterGB, RubensteinJL, ScheuerT, dela Iglesia HO, CatterallWA. Autistic-like behaviour in Scn1a+/- mice and rescue by enhanced GABA-mediated neurotransmission Nature, 2012,489(7416):385-390. [本文引用: 1]
ZhuJW . TRIM32 deficiency leads to autistic behaviors in mice[Dissertation] Southern Medical University, 2016. [本文引用: 5]
BurrowsEL, LaskarisL, KoyamaL, ChurilovL, BornsteinJC, Hill-YardinEL, HannanAJ . A neuroligin-3 mutation implicated in autism causes abnormal aggression and increases repetitive behavior in mice Mol Autism, 2015,6:62. [本文引用: 1]
HessSE, RohrS, DufourBD, GaskillBN, PajorEA, GarnerJP . Home improvement: C57BL/6J mice given more naturalistic nesting materials build better nests J Am Assoc Lab Anim Sci, 2008,47(6):25-31. [本文引用: 1]
ChaboutJ, SerreauP, EyE, BellierL, AubinT, BourgeronT, GranonS . Adult male mice emit context- specific ultrasonic vocalizations that are modulated by prior isolation or group rearing environment PLoS One, 2012,7(1):e29401. [本文引用: 1]
BrunnerD, KabitzkeP, HeD, CoxK, ThiedeL, HananiaT, SabathE, AlexandrovV, SaxeM, PelesE, MillsA, SpoorenW, GhoshA, FelicianoP, BenedettiM, LuoCA, BiemansB . Comprehensive analysis of the 16p11. 2 deletion and null cntnap2 mouse models of autism spectrum disorder PLoS One.2015,10(8):e134572. [本文引用: 1]
NingN, ZhangYS, YangGX . A review of the researches into restricted and repetitive behaviors of children with autism spectrum disorder Chin J Spec Edu, 2015, ( 2):46-52. [本文引用: 1]
TakayanagiY, FujitaE, YuZ, YamagataT, MomoiMY, MomoiT, OnakaT . Impairment of social and emotional behaviors in Cadm1-knockout mice Biochem Biophys Res Commun, 2010,396(3):703-708. [本文引用: 1]
WilliamsDL, GoldsteinG, MinshewNJ . The profile of memory function in children with autism Neuropsychology, 2006,20(1):21-29. [本文引用: 1]
ShojiH, MiyakawaT . Age-related behavioral changes from young to old age in male mice of a C57BL/6J strain maintained under a genetic stability program Neuropsychopharmacol Rep, 2019,39(2):100-118. [本文引用: 1]
HanRT, KimYB, ParkEH, KimJY, RyuC, KimHY, LeeJ, PahkK, ShanyuC, KimH, BackSK, KimHJ, KimYI, NaHS . Long-term isolation elicits depression and anxiety-related behaviors by reducing oxytocin- induced GABAergic transmission in central amygdala Front Mol Neurosci, 2018,11:246. [本文引用: 1]
ZhaoH, JiangYH, ZhangYQ . Modeling autism in non-human primates: opportunities and challenges Autism Res, 2018,11(5):686-694. [本文引用: 1]
ChenQ, DeisterCA, GaoX, GuoBL, Lynn-JonesT, ChenNY, WellsMF, LiuRP, GoardMJ, DimidschsteinJ, FengSJ, ShiYW, LiaoWP, LuZH, FishellG, MooreCI, FengGP . Dysfunction of cortical GABAergic neurons leads to sensory hyper-reactivity in a Shank3 mouse model of ASD Nat Neurosci, 2020,23(4):520-532. [本文引用: 1]
BurketJA, YoungCM, GreenTL, BensonAD, DeutschSI . Characterization of gait and olfactory behaviors in the Balb/c mouse model of autism spectrum disorders Brain Res Bull, 2016,122:29-34. [本文引用: 3]
RoemersP, HulstY, vanHeijningen S, vanDijk G, vanHeuvelen MJG, DeDeyn PP, vander Zee EA. Inducing physical inactivity in mice: preventing climbing and reducing cage size negatively affect physical fitness and body composition Front Behav Neurosci, 2019,13:221. [本文引用: 1]
GargSK, LioyDT, ChevalH, McgannJC, BissonnetteJM, MurthaMJ, FoustKD, KasparBK, BirdA, MandelG . Systemic delivery of MeCP2 rescues behavioral and cellular deficits in female mouse models of Rett syndrome J Neurosci.2013,33(34):13612-13620. [本文引用: 2]
OreficeLL, ZimmermanAL, ChirilaAM, SlebodaSJ, HeadJP, GintyDD . Peripheral mechanosensory neuron dysfunction underlies tactile and behavioral deficits in mouse models of ASDs Cell, 2016,166(2):299-313. [本文引用: 4]
YooJ, BakesJ, BradleyC, CollingridgeGL, KaangBK . Shank mutant mice as an animal model of autism Philos Trans R Soc Lond B Biol Sci, 2013, 369(1633):20130143. [本文引用: 3]
WonH, LeeHR, GeeHY, MahW, KimJI, LeeJ, HaS, ChungC, JungES, ChoYS, ParkSG, LeeJS, LeeK, KimD, BaeYC, KaangBK, LeeMG, KimE . Autistic-like social behaviour in Shank2-mutant mice improved by restoring NMDA receptor function Nature, 2012,486(7402):261-265. [本文引用: 3]
WangX, MccoyPA, RodriguizRM, PanY, JeHS, RobertsAC, KimCJ, BerriosJ, ColvinJS, Bousquet-MooreD, LorenzoI, WuG, WeinbergRJ, EhlersMD, PhilpotBD, BeaudetAL, WetselWC, JiangYH . Synaptic dysfunction and abnormal behaviors in mice lacking major isoforms of Shank3 Hum Mol Genet, 2011,20(15):3093-3108. [本文引用: 3]
JaramilloTC, SpeedHE, XuanZ, ReimersJM, EscamillaCO, WeaverTP, LiuS, FilonovaI, PowellCM . Novel shank3 mutant exhibits behaviors with face validity for autism and altered striatal and hippocampal function Autism Res, 2017,10(1):42-65. [本文引用: 3]
JaramilloTC, XuanZ, ReimersJM, EscamillaCO, LiuS, PowellCM . Early restoration of Shank3 Expression in shank3 knock-out mice prevents core ASD-like behavioral phenotypes eNeuro, 2020,7(3). [本文引用: 3]
ReynoldsCD, NolanSO, JeffersonT, LugoJN . Sex-specific and genotype-specific differences in vocalization development in FMR1 knockout mice Neuroreport, 2016,27(18):1331-1335. [本文引用: 2]
RotschaferSE, TrujilloMS, DansieLE, EthellIM, RazakKA . Minocycline treatment reverses ultrasonic vocalization production deficit in a mouse model of Fragile X Syndrome Brain Res, 2012,1439:7-14. [本文引用: 2]
SpencerCM, AlekseyenkoO, SeryshevaE, Yuva- PaylorLA, PaylorR . Altered anxiety-related and social behaviors in the Fmr1 knockout mouse model of fragile X syndrome Genes Brain Behav, 2005,4(7):420-430. [本文引用: 2]
DeloreyTM, SahbaieP, HashemiE, HomanicsGE, ClarkJD . Gabrb3 gene deficient mice exhibit impaired social and exploratory behaviors, deficits in non-selective attention and hypoplasia of cerebellar vermal lobules: a potential model of autism spectrum disorder Behav Brain Res, 2008,187(2):207-220. [本文引用: 2]
ShihPY, HsiehBY, LinMH, HuangTN, TsaiCY, PongWL, LeeSP, HsuehYP . CTTNBP2 controls synaptic expression of zinc-related autism-associated proteins and regulates synapse formation and autism-like behaviors Cell Rep, 2020,31(9):107700. [本文引用: 3]
YooT, KimSG, YangSH, KimH, KimE, KimSY . A DLG2 deficiency in mice leads to reduced sociability and increased repetitive behavior accompanied by aberrant synaptic transmission in the dorsal striatum Mol Autism, 2020,11(1):19. [本文引用: 3]
GilbertJ, O'ConnorM, TempletS, MoghaddamM, DiVia IA, SinclairA, ZhuLQ, XuW, ManHY. Nexmif/ Kidlia knock-out mouse demonstrates autism-like behaviors, memory deficits, and impairments in synapse formation and function J Neurosci, 2020,40(1):237-254. [本文引用: 3]
ArmstrongEC, CarusoA, ServadioM, AndreaeLC, TrezzaV, ScattoniML, FernandesC . Assessing the developmental trajectory of mouse models of neurodevelopmental disorders: Social and communication deficits in mice with Neurexin 1alpha deletion Genes Brain Behav, 2020,19(4):e12630. [本文引用: 3]
EthertonMR, BlaissCA, PowellCM, SudhofTC . Mouse neurexin-1alpha deletion causes correlated electrophysiological and behavioral changes consistent with cognitive impairments Proc Natl Acad Sci USA, 2009,106(42):17998-18003. [本文引用: 3]
BurrowsEL, LaskarisL, KoyamaL, ChurilovL, BornsteinJC, Hill-YardinEL, HannanAJ . A neuroligin-3 mutation implicated in autism causes abnormal aggression and increases repetitive behavior in mice Mol Autism, 2015,6:62. [本文引用: 2]
CaoW, LinS, XiaQQ, DuYL, YangQ, ZhangMY, LuYQ, XuJ, DuanSM, XiaJ, FengG, XuJ, LuoJH . Gamma oscillation dysfunction in mpfc leads to social deficits in neuroligin 3 R451C knockin mice Neuron, 2018,97(6):1253-1260. [本文引用: 2]
El-KordiA, WinklerD, HammerschmidtK, K?stnerA, KruegerD, RonnenbergA, RitterC, JathoJ, RadyushkinK, BourgeronT, FischerJ, BroseN, EhrenreichH . Development of an autism severity score for mice using Nlgn4 null mutants as a construct-valid model of heritable monogenic autism Behav Brain Res, 2013,251:41-49. [本文引用: 3]
NakanishiM, NomuraJ, JiX, TamadaK, AraiT, TakahashiE, Bu?anM, TakumiT . Functional significance of rare neuroligin 1 variants found in autism PLoS Genet, 2017,13(8):e1006940. [本文引用: 3]
Clipperton-AllenAE, PageDT . Pten haploinsufficient mice show broad brain overgrowth but selective impairments in autism-relevant behavioral tests Hum Mol Genet, 2014,23(13):3490-3505. [本文引用: 4]
CupolilloD, HoxhaE, FaralliA, De LucaA, RossiF, TempiaF, CarulliD . Autistic-Like Traits and Cerebellar Dysfunction in Purkinje Cell PTEN Knock-Out Mice Neuropsychopharmacol, 2016,41(6):1457-1466. [本文引用: 2]
EhningerD, HanS, ShilyanskyC, ZhouY, LiW, KwiatkowskiDJ, RameshV, SilvaAJ . Reversal of learning deficits in a Tsc2+/- mouse model of tuberous sclerosis Nat Med, 2008,14(8):843-848. [本文引用: 3]
TsaiPT, RudolphS, GuoC, EllegoodJ, GibsonJM, SchaefferSM, MogaveroJ, LerchJP, RegehrW, SahinM . Sensitive Periods for cerebellar-mediated autistic- like behaviors Cell Rep, 2018,25(2):357-367. [本文引用: 3]
ZhengF, KasperLH, BedfordDC, LerachS, TeubnerBJ, BrindlePK . Mutation of the CH1 domain in the histone acetyltransferase crebbp results in autism-relevant behaviors in mice PLoS One, 2016,11(1):e146366. [本文引用: 3]
SilvermanJL, YangM, LordC, CrawleyJN . Behavioural phenotyping assays for mouse models of autism Nat Rev Neurosci, 2010,11(7):490-502. [本文引用: 1]
BowersJM, KonopkaG. ASD-relevant animal models of the foxp family of transcription factors Autism Open Access, 2012, (Suppl.) 1(10):10082. [本文引用: 1]
MedvedevaVP, RiegerMA, ViethB, MombereauC, ZiegenhainC, GhoshT, CressantA, EnardW, GranonS, DoughertyJD, GroszerM . Altered social behavior in mice carrying a cortical Foxp2 deletion Hum Mol Genet, 2019,28(5):701-717. [本文引用: 1]
NakataniJ, TamadaK, HatanakaF, IseS, OhtaH, InoueK, TomonagaS, WatanabeY, ChungYJ, BanerjeeR, IwamotoK, KatoT, OkazawaM, YamauchiK, TandaK, TakaoK, MiyakawaT, BradleyA, TakumiT . Abnormal behavior in a chromosome-engineered mouse model for human 15q11-13 duplication seen in autism Cell, 2009,137(7):1235-1246. [本文引用: 2]
LacariaM, SpencerC, GuW, PaylorR, LupskiJR . Enriched rearing improves behavioral responses of an animal model for CNV-based autistic-like traits Hum Mol Genet, 2012,21(14):3083-3096. [本文引用: 2]
MolinaJ, Carmona-MoraP, ChrastJ, KrallPM, CanalesCP, LupskiJR, ReymondA, WalzK . Abnormal social behaviors and altered gene expression rates in a mouse model for Potocki-Lupski syndrome Hum Mol Genet, 2008,17(16):2486-2495. [本文引用: 2]
MeyzaKZ, DefensorEB, JensenAL, CorleyMJ, PearsonBL, PobbeRL, BolivarVJ, BlanchardDC, BlanchardRJ . The BTBR T+ tf/J mouse model for autism spectrum disorders-in search of biomarkers Behav Brain Res, 2013,251:25-34. [本文引用: 3]
BrodkinES . BALB/c mice: low sociability and other phenotypes that may be relevant to autism Behav Brain Res, 2007,176(1):53-65. [本文引用: 2]
RyanBC, YoungNB, CrawleyJN, BodfishJW, MoySS . Social deficits, stereotypy and early emergence of repetitive behavior in the C58/J inbred mouse strain Behav Brain Res, 2010,208(1):178-188. [本文引用: 3]
SunXP, WangQ, ShiZ, ChenSG, LiuX . Review and prospect of experiment methodology on animal behavior Chin J Comp Med, 2018,28(3):1-7. [本文引用: 1]
 ,东南大学生命科学与技术学院,发育与疾病相关基因教育部重点实验室,南京 210096
,东南大学生命科学与技术学院,发育与疾病相关基因教育部重点实验室,南京 210096
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT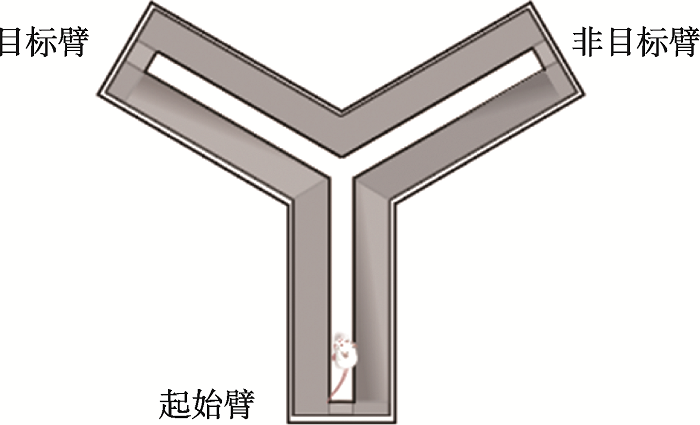 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT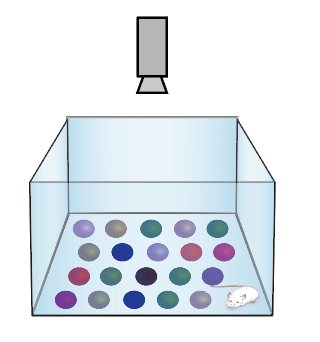 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT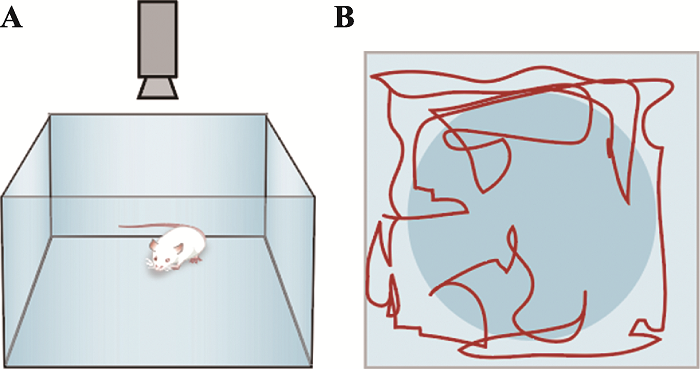 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT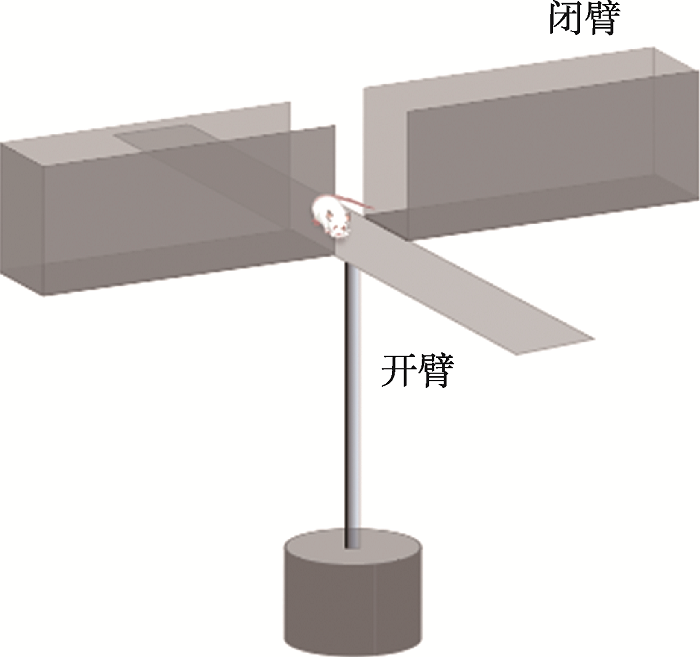 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT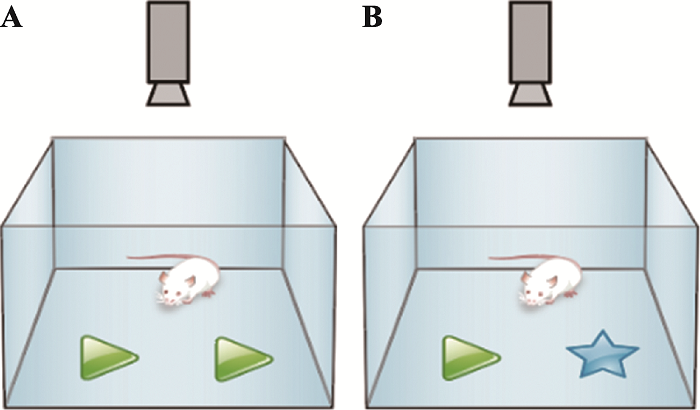 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT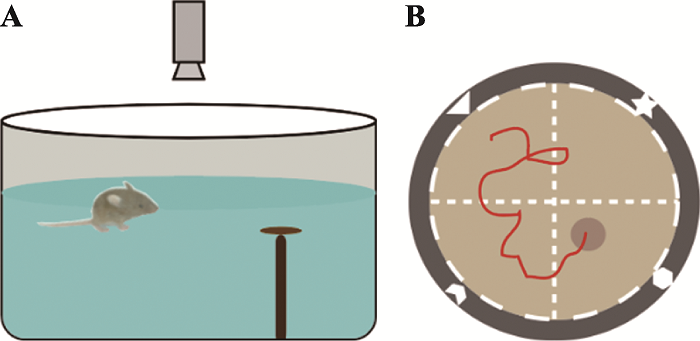 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}