 ,1
,1Genetics of fertility restoration in the isocytoplasm allonuclear C-group of cytoplasmic male sterility in maize
Zhuofan Zhao1, Ling Huang1,2, Yongming Liu1, Peng Zhang1, Gui Wei1, Moju Cao,1通讯作者:
编委: 严建兵
收稿日期:2017-12-7修回日期:2018-02-7网络出版日期:2018-05-20
| 基金资助: |
Editorial board:
Received:2017-12-7Revised:2018-02-7Online:2018-05-20
| Fund supported: |
作者简介 About authors
赵卓凡,硕士研究生,专业方向:玉米生物技术育种E-mail:

摘要
关键词:
Abstract
Keywords:
PDF (5570KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
赵卓凡, 黄玲, 刘永明, 张鹏, 魏桂, 曹墨菊. 玉米CMS-C同质异核不育系育性恢复的遗传研究. 遗传[J], 2018, 40(5): 402-414 doi:10.16288/j.yczz.17-401
Zhuofan Zhao, Ling Huang, Yongming Liu, Peng Zhang, Gui Wei, Moju Cao.
植物在有性繁殖过程中不能产生正常的花药、花粉或雄配子的遗传现象称为雄性不育(male sterility)。根据遗传特点,植物雄性不育可以分为两类:细胞质雄性不育(cytoplasmic male sterility, CMS)和细胞核雄性不育(genic male sterility, GMS),CMS由核基因与线粒体基因互作不协调引起,而GMS由核基因单独控制[1]。玉米(Zea mays L.)是最早利用细胞质雄性不育生产杂交种的作物,根据恢复专效性,玉米CMS分为T、S和C共3种类型[2, 3]。由于玉米T小种的爆发流行致使玉米T型雄性不育的利用受到限制[4];S型属于配子体不育,败育不彻底且受环境影响较大,在生产上难以大面积推广[5];C型不育系属于孢子体不育,花粉败育彻底,育性表现相对稳定,在生产上可能具有较大的应用前景[6, 7]。
早期基于线粒体DNA内切酶消化图谱的差异,人们推测植物细胞质雄性不育可能与线粒体有 关[8,9,10]。经进一步的研究表明,大多数植物细胞质雄性不育的形成与嵌合的线粒体开放阅读框(open reading frame, ORF)有关[11,12]。细胞质雄性不育系的育性恢复机理复杂多样,恢复基因一方面可通过抑制线粒体嵌合不育基因的转录或者翻译而使育性恢 复[9,13~16],另一方面也可通过改变线粒体基因组DNA的结构使育性恢复[17,18,19]。关于玉米CMS-C的育性恢复,不同研究者利用不同材料,从不同角度进行了大量研究,结果不尽相同。陈伟程等[20]认为玉米CMS-C育性恢复受两对独立的显性重复基因控制,其后来研究表明玉米CMS-C育性恢复受Rf4、Rf5强恢复基因及Rf6弱恢复基因控制[21];Khey-Pour等[22]证明玉米CMS-C亚组中的C、RB、ES、Bb共4种胞质的育性恢复均只受Rf4单基因控制;Sisco[23]以玉米CMS-C中的CⅠ亚组不育胞质为材料,用RFLP分子标记技术将恢复系A619携带的恢复基因定位在第8号染色体短臂末端,但同时指出在第3号染色体上也可能存在Rf4的部分序列;Hu等[24]认为玉米CMS-C育性恢复基因Rf5存在一个显性抑制基因Rf-I,该基因对恢复基因Rf4不具有抑制作用。
玉米CMS-C育性恢复除了受恢复基因影响,特殊环境也可能对其表现有影响。陈伟程等[25]指出凉爽高湿更有利于玉米CMS-C育性恢复,并且植株的育性表现与环境有关。Christophe等[26]将玉米7个CMS-T、8个CMS-C和7个CMS-S 3种CMS胞质的杂交群体在17种环境中种植,发现CMS-T的稳定性最高,其次是CMS-C,CMS-S的稳定性最差。Bueckmann等[27]对玉米的2个CMS-S(Torres和Zidane)和1个CMS-T(DSP2)进行研究,结果表明在高温条件下DSP2和Torres几乎都完全不育,而在低温条件下有部分植株花药外露且能少量散粉。Tracy等[28]对玉米几个恢复程度不同的A632亚系进行研究,发现亚系A632-Pr-MS对不育性几乎完全保持,亚系A632-Pr-LB-1和A632-Pr-LB -2对不育性表现不同程度的育性恢复。将A632-Pr-LB-1和A632-Pr-LB -2与A632-Pr-MS分别进行杂交,连续两年种植其杂交后代并观察育性表现,发现不同年份间植株雄花颖壳表现迟开裂并部分可育(late-breaking partial fertility, LBPMF)的比例差异较大,表明环境对植株雄花颖壳的开裂有明显影响。陈伟等[29]对新鉴定出的爆裂玉米细胞质雄性不育系CMS-P进行研究,发现不同测验系对相同不育系的育性恢复能力存在差别,同一测验系与CMS-P的测交F1在不同年份间花粉可染率也不同。关于环境对植物细胞质雄性不育系育性恢复表现的影响,在其他植物上已有研究报道。如杨光圣等[30]对甘蓝型油菜(Brassica napus L.) 4种主要不育胞质类型Ogu、Pol、nap、陕2A和白菜型油菜(Brassica campestris L.)细胞质雄性不育系阿油A,通过在不同光照条件和不同地区种植观察,发现油菜细胞质雄性不育的育性表现受温度影响,而与光照无关。
此外,玉米CMS的育性恢复受多个QTL位点控制。如Kohls等[31]研究表明,玉米CMS-C的育性恢复受多基因控制,并将其3个主要的QTL位点定位在染色体bin 2.09、bin 3.06和bin 7.03,同时检测到6个微效QTL位点。Feng等[5]通过对玉米CMS-S进行育性恢复的QTL定位研究,除定位到Rf3外,还定位到19个控制花粉育性的主效QTL位点、3个控制花药开裂的主效QTL位点以及8个控制散粉的主效QTL位点。
本课题组前期研究发现,测验系18白能够恢复C48-2的育性,但对C478为保持;自330能恢复C478,但对C48-2为保持;而自交系A619则对C48-2和C478均为恢复。为了进一步探究玉米CMS-C育性恢复机理,本研究一方面将相同胞质不同核背景(以下简称同质异核)的不育系C48-2、C黄早四、C478与测验系18白、自330、5022以及恢复系A619杂交进行恢保关系测定,同时将不育系C48-2、C黄早四、C478分别与保持系48-2、黄早四、478组配不完全双列杂交F1;另一方面将育性恢复F1:(C黄早四×自330) F1、(C478×自330) F1、(C48-2×18白) F1、(C48-2×A619) F1自交获得F2,同时将育性恢复F1:(C48-2×18白) F1F、(C48-2×A619) F1F和(C478×自330) F1F的花粉分别授予育性保持F1:(C478×18白) F1S、(C48-2×自330) F1S,以获得双交群体。通过在不同年份不同地点种植观察杂交后代的育性表现,探索玉米CMS-C育性恢复的影响因素,为玉米CMS-C不育系的生产利用提供参考。
1 材料和方法
1.1 材料
不育系:C48-2,C黄早四,C478;保持系:48-2、黄早四、478;测验系:18白、自330、5022;恢复系A619。不育系与测验系杂交获得F1,用于恢保关系鉴定。根据恢保关系鉴定结果,将育性恢复F1:(C黄早四×自330) F1、(C478×自330) F1、(C48-2×18白) F1、(C48-2×A619) F1自交获得4个F2群体;将育性恢复F1:(C48-2×18白) F1F、(C48-2×A619) F1F和(C478×自330) F1F分别与不育性保持F1:(C478×18白) F1S、(C48-2×自330) F1S 杂交组配6个双交群体。4个F2群体及6个双交群体具体信息见表1。其中A619由河南农业大学汤继华老师馈赠,其余供试材料均由四川农业大学玉米研究所提供。Table 1
表1
表1 供试F2及双交群体的信息
Table 1
| 材料名称 | 群体大小(株) | 田间区号 | 种植时间 | 种植地点 |
|---|---|---|---|---|
| (C黄早四×自330) F2 | 134 | 2008年4月 | 四川雅安 | |
| (C478×自330) F2 | 142 | |||
| (C48-2×18白) F2 | 127 | 154 | 2014年10月 | 云南西双版纳 |
| (C48-2×A619) F2 (2013年10月云南组配) | 170 | 155 | ||
| (C478×自330) F2 | 126 | 156 | ||
| (C478×18白) F1S×(C48-2×18白) F1F | 94 | 157 | ||
| (C478×18白) F1S×(C48-2×A619) F1F | 129 | 158 | ||
| (C478×18白) F1S×(C478×自330) F1F | 91 | 159 | ||
| (C48-2×自330) F1S×(C48-2×18白) F1F | 110 | 160 | ||
| (C48-2×自330) F1S×(C48-2×A619) F1F | 131 | 161 | ||
| (C48-2×自330) F1S×(C478×自330) F1F | 133 | 162 | ||
| (C48-2×A619) F2 (2013年10月云南组配) | 156 | Z1 | 2015年4月 | 四川温江 |
| (C48-2×A619) F2 (2014年10月云南组配) | 95 | Z2 |
新窗口打开|下载CSV
1.2 材料种植
2006年4月在四川雅安分别播种不育系:C48-2、C黄早四、C478;保持系:48-2、黄早四、478;测验系:18白、自330、5022和恢复系A619,并以不育系为母本,保持系、测验系及恢复系为父本,配制杂交F1。2007年4月在四川雅安种植所有杂交Fl,并进行育性鉴定,将育性恢复F1进行自交获得F2,同时用育性恢复F1给不育性保持F1授粉获得双交群体。2008年4月在四川雅安播种(C黄早四×自330) F2、(C478×自330) F2群体。2014年10月在云南西双版纳种植3个F2群体(田间区号为154、155、156)和6个双交群体(田间区号为157~162)。2015年4月在四川温江种植2个(C48-2×A619) F2群体(田间区号为Z1、Z2)。所有材料均采用常规大田管理,单株挂牌,在抽雄后进行田间育性调查和花粉室内染色观察。1.3 田间育性鉴定
在成株期,采用Duvick[32]提出的5级育性鉴定标准(表2),对所有供试材料进行单株育性调查。每两天调查一次,直到雄穗散粉完毕。Table 2
表2
表2 玉米雄花育性5级分级标准
Table 2
| 级别 | 类型 | 表型 |
|---|---|---|
| Ⅰ级 | 完全不育 | 雄花没有花粉或花粉发生败育,花药干瘪,不露出颍壳 |
| Ⅱ级 | 高度不育 | 雄花没有花粉或花粉发生败育,少数花药露出颍壳,花药干瘪 |
| Ⅲ级 | 半可育 | 花药半饱满,露出颍壳的花药常占半数,不同植株差异较大 |
| Ⅳ级 | 高度可育 | 雄花小穗大部分开花散粉,花粉正常,大部分花药露出颍壳,花药饱满 |
| Ⅴ级 | 完全可育 | 雄花小穗都能正常开花,并大量散粉,花粉正常 |
新窗口打开|下载CSV
1.4 室内花粉镜检
当可育植株雄穗有少量花药外露时,在每株雄穗主轴上、中、下3个部位分别取1对小穗,用FAA固定液(100 mL甲醛、100 mL乙酸、1800 mL70%乙醇)将其固定于1.5 mL EP管中。室内镜检时,将上、中、下3对小穗的花药混合,用1~2滴1% I2-IK染色,观察3个视野,分别对可染、半染和不染的花粉粒数目进行统计,分类标准见表3,计算花粉的可染率。Table 3
表3
表3 花粉育性鉴定标准
Table 3
| 类型 | 表型 |
|---|---|
| 可染 | 花粉与I2-IK染液反应后成黑色或蓝黑色,花粉的形状规则,表现为椭球形或圆球形,且充满整个花粉粒 |
| 不染 | 花粉与I2-IK染液反应后不变色,仍为染液的颜色,花粉形状为透明的空壳 |
| 半染 | 花粉与I2-IK染液反应后成黄褐色,花粉形状规则,表现为圆球形或椭球形 |
新窗口打开|下载CSV
1.5 统计分析
通过χ2法检验各群体育性分离比并对小穗开裂与花粉可染性进行独立性检验。2 结果与分析
2.1 恢保关系测定
以测验系18白、自330、5022和恢复系A619为父本,对玉米CMS-C同质异核不育系C48-2、C黄早四、C478进行授粉杂交,获得杂交F1。对F1进行种植观察。在成株期,按照Duvick[32]的5级育性标准进行田间育性鉴定,结果见表4;对F1进行花粉室内染色镜检,结果见表5。由表4及表5可以发现,18白能完全恢复C48-2,但不能恢复C黄早四和C478;自330能恢复C黄早四和C478,但不能恢复C48-2;而5022对C48-2、C黄早四和C478均不能恢复其育性;A619则完全恢复C48-2、C黄早四和C478。Table 4
表4
表4 不育系与测验系杂交F1田间育性表现
Table 4
| 不育系 | 测验系 | |||
|---|---|---|---|---|
| 18白 | 自330 | 5022 | A619 | |
| C48-2 | F | S | S | F |
| C黄早四 | S | F | S | F |
| C478 | S | F | S | F |
新窗口打开|下载CSV
Table 5
表5
表5 不育系与测验系杂交F1花粉镜检结果
Table 5
| 杂交组合 | 可育株 | 不育株 | 花粉镜检 | ||
|---|---|---|---|---|---|
| 可染(%) | 半染(%) | 不染(%) | |||
| (C48-2×18白) F1 | 42 | 0 | 89.5 | 1.4 | 9 |
| (C黄早四×18白) F1 | 0 | 42 | 27 | 0 | 73 |
| (C478×18白) F1 | 0 | 40 | 1.3 | 0.2 | 89.5 |
| (C48-2×自330) F1 | 0 | 42 | 5.2 | 0 | 94.4 |
| (C黄早四×自330) F1 | 42 | 0 | 97.5 | 0 | 2.5 |
| (C478×自330) F1 | 42 | 0 | 54.2 | 0.7 | 45.1 |
| (C48-2×5022) F1 | 0 | 41 | 3.2 | 0 | 96.8 |
| (C黄早四×5022) F1 | 0 | 42 | 0 | 0 | 100 |
| (C478×5022) F1 | 0 | 42 | 13.3 | 0 | 86.7 |
新窗口打开|下载CSV
将不育系C48-2、C黄早四、C478与保持系48-2、黄早四、478组配不完全双列杂交,并对杂交F1的田间育性表现进行调查,结果见表6。由表6可以发现,48-2对C48-2和C478不育性保持,对C黄早四恢复育性;黄早四对C黄早四和C478不育性保持,对C48-2恢复育性;478对C48-2、C黄早四和C478均保持不育性。
Table 6
表6
表6 同质异核不育系与其相应保持系组配杂交F1的育性表现
Table 6
| 不育系 | 保持系 | ||
|---|---|---|---|
| 48-2 | 黄早四 | 478 | |
| C48-2 | S | F | S |
| C黄早四 | F | S | S |
| C478 | S | S | S |
新窗口打开|下载CSV
可见,同一测验系对相同胞质不同核背景下的不育系具有不同的恢复能力,同一不育系对不同测验系的育性恢复反应不同。
2.2 育性恢复组合F2群体的育性表现
将育性恢复组合(C48-2×18白) F1、(C黄早四×自330) F1、(C478×自330) F1和(C48-2×A619) F1自交获得F2群体。2008年4月,在四川雅安种植(C黄早四×自330) F2和(C478×自330) F2,2014年10月在云南西双版纳种植(C48-2×18白) F2、(C48-2×A619) F2和(C478×自330) F2,成株期进行田间育性调查和室内花粉镜检。结果表明,2008年4月在四川雅安种植的(C黄早四×自330) F2群体,其可育株(F)∶不育株(S)符合3∶1;而(C478×自330) F2群体可育株(F)∶不育株(S)符合9∶7,然而2014年10月在云南种植的(C478×自330) F2群体可育株(F)与不育株(S)的分离比为39∶87,与2008年4月在四川雅安种植的(C478×自330) F2群体育性分离结果存在明显的偏差(表7)。可见,自330对C478和C黄早四的恢复效应不同,且在不同环境条件下自330对C478恢复后代的育性表现不同。
Table 7
表7
表7 F2群体的育性表现及卡平方检验
Table 7
| 材料名称 | 总株数 | F∶S | 期望比例 | χ2 | 种植时间 | 种植地点 | 田间区号 |
|---|---|---|---|---|---|---|---|
| (C黄早四×自330)F2 | 134 | 92∶42 | 3∶1 | 2.55 | 2008年4月 | 四川雅安 | |
| (C478×自330) F2 | 142 | 80∶62 | 9∶7 | 0.00 | |||
| (C48-2×18白) F2 | 127 | 74∶53 | 9∶7 | 0.14 | 2014年10月 | 云南西双版纳 | 154 |
| (C48-2×A619) F2 (2013年10月云南组配) | 170 | 156∶14 | 15∶1 | 0.83 | 155 | ||
| (C478×自330) F2 | 126 | 39∶87 | 156 | ||||
| (C48-2×A619) F2 (2013云南组配) | 156 | 143∶13 | 15∶1 | 2.47 | 2015年4月 | 四川温江 | Z1 |
| (C48-2×A619) F2 (2014云南组配) | 95 | 88∶7 | 15∶1 | 0.06 | 2015年4月 | Z2 |
新窗口打开|下载CSV
2014年10月在云南西双版纳种植(C48-2×18白) F2群体,可育株(F)∶不育株(S)分离比符合9∶7;在不同地点(云南、四川)种植(C48-2×A619) F2群体,可育株(F)∶不育株(S)分离比均为15∶1,表明A619对C48-2表现出两对恢复基因。
2014年10月,在云南西双版纳种植(C48-2× A619) F2 (2013年10月云南组配,田间区号为155),可育株(Ⅲ+Ⅳ+Ⅴ)占整个群体的91.76%,其中完全可育株(Ⅴ级)只占整个群体的18.24% (图1)。2015年4月,在四川温江种植不同年份组配的(C48-2× A619) F2,其中2013年10月云南组配材料田间区号记为Z1,2014年10月云南组配材料田间区号记为Z2。结果表明,Z1、Z2两组材料的可育株(Ⅲ+Ⅳ+Ⅴ)分别占整个群体的91.03%和92.63%,其中完全可育株(Ⅴ级)分别占整个群体的73.08%和68.42%;同一年份组配的(C48-2×A619) F2,分别于2014年10月在云南(田间区号为155)和2015年4月在四川(田间区号为Z1)种植,可育株在育性表现级别上存在差异,2014年10月在云南种植的可育株多为Ⅲ、Ⅳ级,而2015年4月在四川种植的可育株多为Ⅴ级;不同年份组配的(C48-2× A619) F2 (Z1与Z2),2015年4月在四川温江种植,可育株在育性表现级别上差异不大,均为Ⅴ级可育株占多数(图1)。可见,不同年份配制的组合,在同一环境下种植,其育性表现较为一致,而同一年份配制的组合在不同环境条件下种植,其花药的外露程度或育性级别有较大差异。
图1
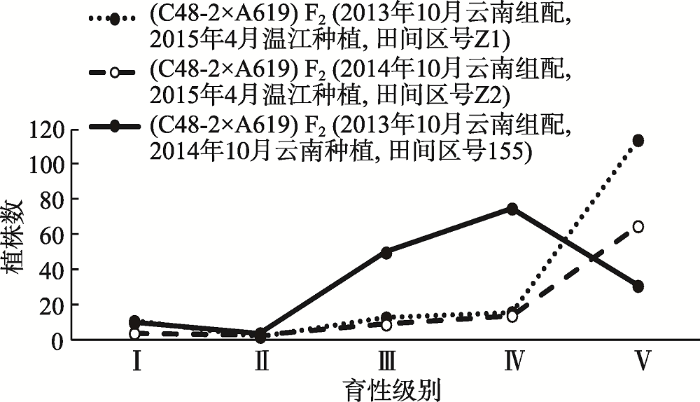 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1(C48-2×A619) F2在不同年份、不同地点种植的田间育性表现
Fig. 1Fertility-restoration reactions of (C48-2×A619) F2 planted at distinct locations in different years
对2014年10月在云南种植的3个F2群体的所有单株编号,进行单株调查和镜检。结果发现,在(C48-2×18白)F2群体小穗颖壳不开裂和半开裂的植株共有109株,占整个群体的85.8%;其花粉表现不染的植株有52株,占整个群体的40.9%;半染的植株有42株,占整个群体的33.1%;花粉可染植株有15株,占11.8%。2014年10月在云南种植的(C48-2× A619) F2群体小穗颖壳不开裂和半开裂的植株有82株,占整个群体的48.2%;其花粉表现不染的植株有12株,占整个群体的7.1%;半染的植株有23株,占整个群体的13.5%;花粉可染植株有47株,占27.6% (图2)。可见,(C48-2×18白) F2群体小穗颖壳不开裂和半开裂的植株比例以及花粉不染和半染的比例均高于(C48-2×A619) F2群体。然而,2015年4月在温江种植的两个(C48-2×A619) F2群体,出现两个极端现象,即不开裂的小穗其花粉不染,全开裂的小穗其花粉全部可染,没有半开裂的小穗,也没有半染的花粉。虽然在云南和四川种植(C48-2×A619) F2群体的育性分离比均符合15∶1,但植株的小穗开裂程度和花粉中淀粉的累积量有一定差异,表明环境对强恢复系所配组合的育性恢复表现也有影响。(C478×自330) F2群体小穗颖壳不开裂和半开裂的植株有112株,占整个群体的88.9%;其花粉表现不染的植株有87株,占整个群体的69%;整个群体花粉半染只有6株,与(C48-2×18白) F2相比,恢复度更低(图2)。
图2
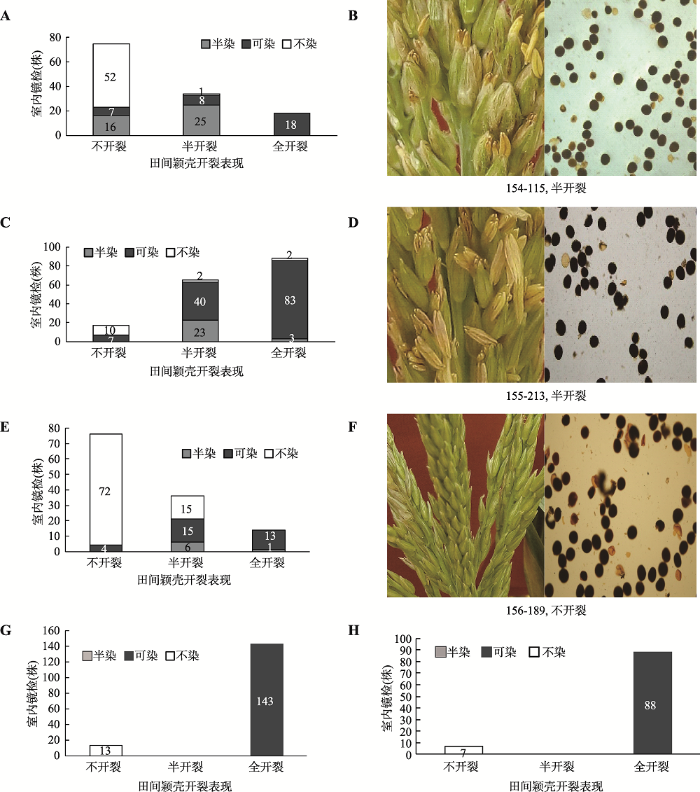 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2F2群体雄穗颖壳开裂程度与花粉可染性
A:(C48-2×18白) F2群体雄穗颖壳开裂程度与花粉可染性统计结果(田间区号154,共127株);B:(C48-2×18白) F2群体中株号为115的植株雄穗颖壳开裂田间表现及花粉 I2-IK染色;C:(C48-2×A619) F2群体雄穗颖壳开裂程度与花粉可染性统计结果(田间区号155,170株);D:(C48-2×A619) F2群体中株号为213的植株雄穗颖壳开裂表现及花粉 I2-IK染色;E:(C478×自330) F2群体雄穗颖壳开裂程度与花粉可染性统计结果(田间区号156,共126株);F:(C478×自330)F2群体中株号为189的植株雄穗颖壳开裂田间表现及花粉I2-IK染色;G:(C48-2×A619) F2群体雄穗颖壳开裂程度与花粉可染性统计结果(田间区号Z1,156株);H:(C48-2×A619) F2群体雄穗颖壳开裂程度与花粉可染性统计结果(田间区号Z2,95株)。柱状图中的数字代表植株数。
Fig. 2Dehiscence degree of tassel glumes and pollen staining of F2 populations
通过对2014年10月在云南种植的3个F2群体的小穗颖壳开裂性状与花粉可染率进行χ2独立性检验,发现χ2=367.94>χ20.01(4)=13.28,表明花粉可染性与小穗颖壳的开裂程度具有相关性。
2.3 双交群体的育性表现
将育性恢复组合(C48-2×18白) F1F、(C48-2× A619) F1F和(C478×自330) F1F的花粉分别授予育性保持组合(C478×18白) F1S和(C48-2×自330) F1S,获得6个双交群体,于2014年10月在云南西双版纳基地种植观察,调查结果见表8。由表8可以看出,[(C478×18白) F1S×(C48-2×18白) F1F]中可育株与不育株育性分离比符合1∶1。由前面的分析可知,18白不能恢复C478,48-2也不能恢复C478,但(C48-2×18白) F1F却能恢复C478育性,推测C48-2核背景中存在微效恢复基因,这些微效基因与18白中的微效恢复基因通过杂交聚合后能使C478的育性恢复;同样,[(C48-2×自330) F1S×(C478×自330) F1F]群体中也出现可育株与不育株的育性分离,表明C478核背景中的微效恢复基因与自330中的微效恢复基因通过杂交聚合后也能使C48-2的育性恢复。Table 8
表8
表8 双交群体育性表现及χ2检验
Table 8
| 材料名称 | 育性级别 | 总株数 | F∶S | 期望比例 | χ2 | 田间区号 | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | ||||||
| (C478×18白) F1S×(C48-2×18白) F1F | 47 | 0 | 18 | 26 | 3 | 94 | 47∶47 | 1∶1 | 0 | 157 |
| (C478×18白) F1S×(C48-2×A619) F1F | 12 | 2 | 42 | 53 | 20 | 129 | 115∶14 | - | - | 158 |
| (C478×18白) F1S×(C478×自330) F1F | 41 | 2 | 12 | 22 | 14 | 91 | 48∶43 | 1∶1 | 0.18 | 159 |
| (C48-2×自330) F1S×(C48-2×18白) F1F | 50 | 9 | 35 | 16 | 0 | 110 | 51∶59 | 1∶1 | 0.45 | 160 |
| (C48-2×自330) F1S×(C48-2×A619) F1F | 35 | 5 | 59 | 29 | 3 | 131 | 91∶40 | - | - | 161 |
| (C48-2×自330) F1S×(C478×自330) F1F | 78 | 5 | 39 | 11 | 0 | 133 | 50∶83 | - | - | 162 |
新窗口打开|下载CSV
[(C478×18白) F1S×(C478×自330) F1F]和[(C48-2×自330) F1S×(C48-2×18白) F1F] 这2个双交群体中可育株(F)∶不育株(S)分离比均符合1∶1,说明自330与18白的主效恢复基因为非等位基因。而对于[(C478×18白) F1S×(C48-2×A619) F1F]群体,129株中可育株为115株,不育株仅为14株;[(C48-2×自330) F1S×(C48-2×A619) F1F]群体有131株,其中可育株91株,不育株仅为40株,表明18白、自330的主效恢复基因均与A619的恢复基因非等位。
2014年10月在云南种植6个双交群体,也均出现小穗颖壳开裂不完全的现象(图3)。对所有单株进行花粉镜检,发现6个群体在田间都有小穗颖壳不开裂,但花粉有可染和半染的情况;田间表现半开裂的小穗几乎全部可染或者半染。有趣的是,以(C478×18白) F1S做母本时,小穗颖壳不开裂的植株其花粉可染或半染的很少;相反,以(C48-2×自330) F1S做母本时,小穗颖壳不开裂的植株其花粉可染或半染的较多(图4)。这些结果表明,C48-2与C478的恢复机理可能存在差异。
图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3双交群体植株雄穗颖壳开裂田间表现和花粉镜检
A~E:分别为双交群体[(C478×18白) F1S×(C48-2×18白) F1F](田间区号157)、[(C478×18白) F1S×(C48-2×A619) F1F] (田间区号158)、[(C478×18白) F1S×(C478×自330) F1F] (田间区号159)、[(C48-2×自330) F1S×(C48-2×18白) F1F] (田间区号160)和[(C48-2×自330) F1S×(C48-2×A619) F1F] (田间区号161)中某单株雄穗颖壳田间开裂表现和花粉I2-IK染色。每张图片下面的数字代表田间区号及相应的株号,如157-44、157-102分别表示第157区的第44株和157区的第102株。
Fig. 3Field characterization of the tassel glumes dehiscence performance and pollen staining of double-cross populations
图4
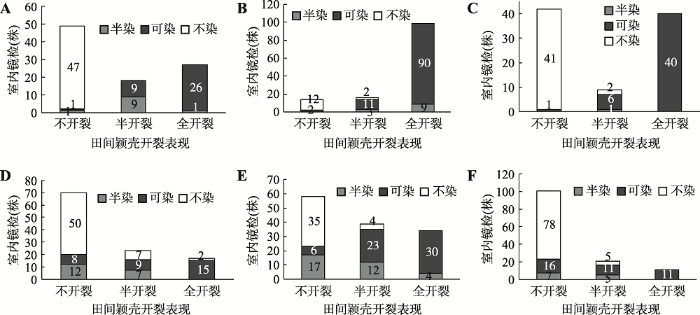 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4双交群体雄穗颖壳开裂程度与花粉可染性
A~F:分别为双交群体[(C478×18白) F1S×(C48-2×18白) F1F] (田间区号157)、[(C478×18白) F1S×(C48-2×A619) F1F] (田间区号158)、[(C478×18白) F1S×(C478×自330) F1F] (田间区号159)、[(C48-2×自330) F1S×(C48-2×18白) F1F] (田间区号160)、[(C48-2×自330) F1S×(C48-2×A619) F1F] (田间区号161)和[(C48-2×自330) F1S×(C478×自330) F1F] (田间区号162)的雄穗颖壳开裂程度与花粉可染性的统计结果。柱状图中的数字代表植株数,不同颜色代表不同的花粉染色表现。
Fig. 4Dehiscence degree of tassel glumes and pollen staining of double-cross populations
同样将6个双交群体小穗颖壳开裂性状及花粉染色调查结果合并,通过χ2独立性检验发现,χ2=529.45>χ20.01(4)=13.28,再次表明小穗颖壳开裂程度与花粉可染率具有一定的相关性。
3 讨 论
3.1 同质异核不育系对相同测验系的育性恢复反应不同,暗示不育系核背景可能参与调控育性恢复表现
本研究基于恢保关系的测定结果,发现同一个测验系对玉米CMS-C同质异核不育系的恢保关系不同。同样也发现同一个保持系对玉米CMS-C同型不育系保持,但对相同不育胞质不同核背景的不育系即异型不育系可能表现为保持,也可能表现为恢复。类似现象在前人的研究中也有发现,如郑用琏等[33]观察到恢313与CMO17的测交后代育性表现为Ⅰ级的保持,与C77的测交后代育性表现为Ⅴ级恢复,MO17与CMO17的测交后代育性表现为Ⅱ级保持,与C77的测交后代育性表现为Ⅳ级恢复。由此不难看出,所谓的恢复系应该是对特定核质组合下的不育系而言,而所谓的保持系也是针对相同核背景下的不育系即同型不育系而言。特定的核质组合不仅决定了其本身的雄花育性表现,也决定了哪些材料能恢复其不育性,哪些材料不能恢复其不育性。可见,玉米CMS-C同质异核不育系的育性恢复不仅取决于不育系本身的核质互作,也依赖于特定的不育系与父本系的核核互作。陈伟程等[34]对玉米CMS-C花粉败育的细胞学观察发现,不同核背景的不育系在败育方式上表现出多样性。Hu等[24]在研究玉米CMS-C的恢复基因Rf5时,也发现同一个材料对某些不育系恢复而对另外一些不育系则不能恢复,并且最终被解释为在不能被恢复的不育系中含有Rf5的抑制基因,所以表现为育性不被恢复。综上可知,玉米CMS-C型不育系的核背景不仅影响其花粉败育方式,同样也影响其育性恢复方式。3.2 微效基因通过累加达到某一个阈值也可以实现对玉米CMS-C不育系的育性恢复
在对本研究所配制的双交群体进行种植观察时发现,双交组合(C478×18白) F1S×(C48-2×18白) F1F和(C48-2×自330) F1S×(C478×自330) F1F都出现了育性分离。前期的研究表明,18白不能恢复C478的育性,48-2也不能恢复C478的育性,但以育性恢复组合(C48-2×18白) F1F给(C478×18白) F1S授粉,双交组合(C478×18白) F1S×(C48-2×18白) F1F却表现出育性分离;同样自330不能恢复C48-2,478也不能恢复C48-2,但以育性恢复组合(C478×自330) F1F给不育性保持组合(C48-2×自330)F1S授粉,双交组合[(C48-2×自330) F1S×(C478×自330) F1F]后代也出现了育性分离。由此我们推测,不育系本身也可能会带有某种育性恢复基因。Kohls等[31]对玉米CMS-C育性部分恢复基因进行QTL,发现母本不育系中也存在恢复基因的QTL位点。由此,本研究认为不育系的核背景很可能存在微效恢复基因,这些微效基因单独作用并不能使不育系的育性恢复,但与同样带有弱恢复基因的测验系杂交时,由于恢复基因的累加作用,可能会对原来单独不能恢复的不育系产生恢复作用。早期曾有报道指出,不育系的育性恢复有时表现为由微效基因控制的数量性状遗传方式,恢复基因对不育系的育性恢复呈现剂量效应,对特定的不育系其恢复基因数量需要达到某一阈值T1,在该阈值之下植株表现为不育,在另一阈值T2之上植株表现为完全恢复,在T1~T2之间时表现为部分可育且恢复基因有剂量效应[35]。最近在普通小麦CMS的研究中也发现微效恢复基因通过杂交聚合,当恢复基因达到一定数量后可以使不育系的育性得以恢复[36]。3.3 环境不仅对玉米CMS-C弱恢复系的恢复作用产生影响,有时对强恢复系的恢复作用也会产生影响
本研究通过对不同年份不同地点配制的杂交组合进行种植观察,发现强恢复系杂交F2代的育性分离比不会因不同年份不同点配制的杂交组合而不同,但却发现同一个强恢复系杂交F2代,在不同年份不同地点种植,其可育株的育性级别构成却存在差异;而对于弱恢复系的杂交F2,在不同年份不同地点种植,其育性分离比会明显发生偏离。如(C478×自330) F2,2008年4月在四川种植时,可育株(F)∶不育株(S)分离比符合9∶7,2014年10月在云南种植时,不育株83,可育株只有43,可育株(F)∶不育株(S)分离比明显偏离9:7。此外,2014年10月在云南及2015年4月在温江分别种植(C48-2×A619) F2,可育株与不育株的分离比均符合15∶1,但田间调查显示云南可育株中Ⅲ+Ⅳ级可育株较多,而Ⅴ级可育较少,四川的田间调查结果则显示可育株主要为V级完全可育。这些结果暗示玉米CMS-C的育性恢复表现受环境影响,并且环境对弱恢复系的恢复作用有较大影响,这与前人对玉米CMS-C、水稻细胞质雄性不育的研究结果相一致[20, 37],并且Nico等[38]详细综述了环境对植物雄性不育的影响过程和机理。而本研究的A619是一个强恢复系,虽然其F2的育性分离比例在四川和云南基本一致,但其可育株的育性级别构成在四川和云南种植有明显的差别。翟虎渠[35]曾在其报道中提出,不育系的育性恢复需要恢复基因达到某一阈值,该阈值可能会因环境条件不同而上下浮动。这更加印证了不育系的育性恢复需要不育系与恢复系的相互作用,无论是强恢复系,还是弱恢复系,其杂交后代的育性都有可能会受到环境的影响。这也暗示在分析环境对育性恢复的影响时,不仅要考虑强恢复系与弱恢复系差别,有时也需要关注杂交种父母本的种质类型或对光温敏反应的敏感性。综上所述,玉米CMS-C育性恢复的复杂性在一定程度上反映了其败育方式的多样性。因此,对玉米CMS-C育性恢复及败育机理的探究仍然是一个重要的研究方向。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URLPMID:24313845 [本文引用: 1]

Abstract In plants, male sterility can be caused either by mitochondrial genes with coupled nuclear genes or by nuclear genes alone; the resulting conditions are known as cytoplasmic male sterility (CMS) and genic male sterility (GMS), respectively. CMS and GMS facilitate hybrid seed production for many crops and thus allow breeders to harness yield gains associated with hybrid vigor (heterosis). In CMS, layers of interaction between mitochondrial and nuclear genes control its male specificity, occurrence, and restoration of fertility. Environment-sensitive GMS (EGMS) mutants may involve epigenetic control by noncoding RNAs and can revert to fertility under different growth conditions, making them useful breeding materials in the hybrid seed industry. Here, we review recent research on CMS and EGMS systems in crops, summarize general models of male sterility and fertility restoration, and discuss the evolutionary significance of these reproductive systems.
URL [本文引用: 1]
Thirty lines with male-sterile cytoplasm were backcrossed to a series of maize (Zea mays L.) inbreds. Each cytoplasm gave male-sterile plants in certain inbred backgrounds and partially or completely male-fertile plants in other backgrounds. The cytoplasms were divided into groups according to the pattern of fertility restoration observed. Each group appears to be composed of similar if not identical cytoplasms. The major groups are designated the Texas Group, the S Group, and the C Group. A fourth group (P) probably belongs to the Texas Group but may be distinct. A few sources of cytoplasm require further testing before they can be assigned to a group. C cytoplasm is assigned the permanent genetic symbol cms3.
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
Cytoplasmic control of susceptibility in maize to yellow leaf blight caused by Phyllostica zeae has been hypothesized in light of: (i) increased susceptibility to the disease of all inbreds with cytoplasm of the Texas male-sterile type, in contrast to the resistance of their normal counterparts; and (ii) uniformity in susceptibility of plants within the population of any inbred line with the Texas male-sterility type cytoplasm. Susceptibility largely governed by a cytoplasmic factor(s) conferring malesterility would insure uniform susceptibility to yellow leaf blight of maize hybrids, even though many inbred lines with a high degree of resistance are available.
URLPMID:26220224 [本文引用: 2]
Key message Thirty loci for fertility restoration of pollen fertility, anther exsertion and pollen shedding to maize CMS-S were identified by GWAS.
URL [本文引用: 1]
玉米雄性不育系利用不仅是玉米育种技术的一项重大变革,也是种子 产业发展的必然趋势.通过分析细胞质雄性不育在玉米育种及生产中的应用现状,得出采用雄性不育技术,创造出具有国际竞争力的玉米新种质和新品种,提高国内 玉米种子生产质量,增强我国种子产业在国际市场上的竞争力,对于我国的种子产业的发展将会产生深远的影响.
[本文引用: 1]
[本文引用: 1]
URLMagsci [本文引用: 1]
URLMagsci [本文引用: 2]
本研究以水稻BT型细胞质雄性不育系秋光和相应的保持系秋光为材料,提取线粒体DNA,用限制性内切酶完全酶解,以玉米线粒体atpA基因和波菜叶绿体atpA基因作为探针,进行分子杂交,将保持系线粒体atpA基因定位在3.5kb的Bam HI酶切片段上,并且以pBR322为载体,克隆了这一片段,另外,在Bam HI完全酶解普带的杂交结果中,不育系线粒体基因组中有两条阳性杂交带,分别是3.5kb和2.9kb,而保持系线粒体基因组中只有3.5kb一条阳性杂交带,因而认为水稻不育系线粒体基因组中可能有两个atpA基因拷贝,而相应的保持系线粒体基因组中只有一个atpA基因拷贝。
URLMagsci [本文引用: 2]
本研究以水稻BT型细胞质雄性不育系秋光和相应的保持系秋光为材料,提取线粒体DNA,用限制性内切酶完全酶解,以玉米线粒体atpA基因和波菜叶绿体atpA基因作为探针,进行分子杂交,将保持系线粒体atpA基因定位在3.5kb的Bam HI酶切片段上,并且以pBR322为载体,克隆了这一片段,另外,在Bam HI完全酶解普带的杂交结果中,不育系线粒体基因组中有两条阳性杂交带,分别是3.5kb和2.9kb,而保持系线粒体基因组中只有3.5kb一条阳性杂交带,因而认为水稻不育系线粒体基因组中可能有两个atpA基因拷贝,而相应的保持系线粒体基因组中只有一个atpA基因拷贝。
URLMagsci [本文引用: 1]
应用RFLP技术,研究了红莲型不育系、保持系、杂种一代及野败型、马协型不育系的线粒体基因。结果表明,红莲型育系与保持系线粒体基因组之间在多外基因区域存在明显差异,为红莲型细胞质雄性不育分子机理研究提供了线索;红莲型不育系与野败型不育系的线粒体基因组之间存在显著差异,马协型不育系与野败型不育系的线粒体基因组之间存在一定差异,在分子水平揭示了不育胞质的多样性。
URLMagsci [本文引用: 1]
应用RFLP技术,研究了红莲型不育系、保持系、杂种一代及野败型、马协型不育系的线粒体基因。结果表明,红莲型育系与保持系线粒体基因组之间在多外基因区域存在明显差异,为红莲型细胞质雄性不育分子机理研究提供了线索;红莲型不育系与野败型不育系的线粒体基因组之间存在显著差异,马协型不育系与野败型不育系的线粒体基因组之间存在一定差异,在分子水平揭示了不育胞质的多样性。
URLPMID:1664299 [本文引用: 1]
Aberrant recombinations involving the mitochondrial atp9, atp6 and coxII genes have created unique chimeric sequences in the C male0sterile cytoplasm ( cms-C ) of maize. An apparent consequence of the rearrangements is the interchanging of transcriptional and/or translational regulatory signals for these genes, and alterations in the reading frames encoding the atp6 and coxII genes in the C cytoplasm. Particularly unusual is the organization of the atp6 gene in cms-C mitochondria, designated atp6-C . The atp6-C sequence is a triple gene fusion product comprised of DNAs derived from atp9, atp6 and an open reading frame of unknown origin. Although there is no direct evidence indicating that these chimeric genes are responsible for the cytoplasmic male sterility (cms) trait, their novel arrangements and the strong correlation between these genes and the C type of male sterility suggest such a role.
URLPMID:23502780 [本文引用: 1]
Plant cytoplasmic male sterility (CMS) results from incompatibilities between the organellar and nuclear genomes and prevents self pollination, enabling hybrid crop breeding to increase yields. The Wild Abortive CMS (CMS-WA) has been exploited in the majority of 'three-line' hybrid rice production since the 1970s, but the molecular basis of this trait remains unknown. Here we report that a new mitochondrial gene, WA352, which originated recently in wild rice, confers CMS-WA because the protein it encodes interacts with the nuclear-encoded mitochondrial protein COX11. In CMS-WA lines, WA352 accumulates preferentially in the anther tapetum, thereby inhibiting COX11 function in peroxide metabolism and triggering premature tapetal programmed cell death and consequent pollen abortion. WA352-induced sterility can be suppressed by two restorer-of-fertility (Rf) genes, suggesting the existence of different mechanisms to counteract deleterious cytoplasmic factors. Thus, CMS-related cytoplasmic-nuclear incompatibility is driven by a detrimental interaction between a newly evolved mitochondrial gene and a conserved, essential nuclear gene.
URLPMID:12782297 [本文引用: 1]
Abstract A fertility restorer gene (Rf-1) of [ms-bo] cytoplasmic male sterility (CMS) in rice has been reported to be responsible for the processing of RNA of aberrant atp6 of mitochondria. We have carried out map-based cloning of the Rf-1 gene and found that a 4.7-kb genomic fragment of a restorer line promoted the processing of aberrant atp6 RNA when introduced into a CMS line. The genomic fragment contained a single open reading frame encoding 18 repeats of the 35 amino acid pentatricopeptide repeat (PPR) motif. The cloned PPR gene is a possible candidate of Rf-1. A non-restoring genotype was identified to have deletions within the coding region.
URLPMID:8000006
Abstract The mitochondrial DNA of plants containing the male sterility-causing Ogura cytoplasm of radish contain a novel gene, orf138, that is transcribed as part of a bicistronic mRNA. Genetic studies have previously linked male sterility with the orf138 locus. To determine if orf138 is expressed at the protein level, and investigate the effect of fertility restoration on ORF138 levels, we have raised antibodies to an ORF138-glutathione S-transferase fusion protein. Anti-ORF138 antibodies detect a 20 kDa protein that is associated with the mitochondrial membrane of sterile Ogura radish plants. Nuclear restoration is accompanied by a dramatic reduction in the amount of this protein in mitochondria of flowers and leaves, but not roots of fertile Ogura radish plants. The presence or absence of fertility restoration genes has no detectable effect on the size, abundance, or RNA editing patterns of orf138 transcripts. These results support genetic studies that have implicated orf138 in Ogura cytoplasmic male sterility and suggest that the restorer genes may be affecting either the translation or stability of ORF138.
URLPMID:28922764
react-text: 98 to idenify major QTls for important yield trais to construct genetic maps for these QTLs /react-text react-text: 99 /react-text
URLMagsci [本文引用: 1]
植物细胞质雄性不育性与育性恢复的分子机理一直是分子生物学的研究热点。文章综述了近十年来的主要研究进展。包括:1、线粒体不育相关区域的确定及其特点;2、不育相关区域的表达谱;3、产生细胞质雄性不育的可能机理;4、恢复基因的可能调控方式;5、恢复基因的遗传、定位及其特征等。拟兰芥、水稻等模式植物线粒体基因组测序工作已经完成,其有关生物学信息及后续研究将极大地推动植物细胞质雄性不育研究取得更快进展。
URLMagsci [本文引用: 1]
植物细胞质雄性不育性与育性恢复的分子机理一直是分子生物学的研究热点。文章综述了近十年来的主要研究进展。包括:1、线粒体不育相关区域的确定及其特点;2、不育相关区域的表达谱;3、产生细胞质雄性不育的可能机理;4、恢复基因的可能调控方式;5、恢复基因的遗传、定位及其特征等。拟兰芥、水稻等模式植物线粒体基因组测序工作已经完成,其有关生物学信息及后续研究将极大地推动植物细胞质雄性不育研究取得更快进展。
URL [本文引用: 1]
URLPMID:12354971 [本文引用: 1]
Abstract Restoration of pollen fertility to cytoplasmic male-sterile common bean by nuclear gene Fr is accompanied by mitochondrial (mt) DNA rearrangements within restored plants. These rearrangements are also observed upon spontaneous cytoplasmic reversion to fertility. An mtDNA fragment of at least 25 kilobases was lost from the genome upon restoration or reversion. This fragment contained DNA segments that were not repeated elsewhere in the genome and, therefore, were not detected within the genome upon fertility restoration. This result suggested that the particular mtDNA configuration absent from restored plants could not be maintained by a constant process of recombination but rather by autonomous replication. No evidence of excision of this region from the mt genome, in the form of a junction fragment associating flanking DNA regions, was detected in fertile restored plants. DNA gel blot hybridization of this mtDNA region, compared with hybridization to related regions of the mitochondrial genome that shared sequence homology, indicated that the mtDNA region associated with sterility was present in lower copy number. These observations, as well as the occurrence of similar or identical rearrangements upon spontaneous cytoplasmic reversion, indicate that the restoration of pollen fertility may be accompanied by loss of an independently replicating subgenomic DNA molecule from the mitochondrial genome.
URLPMID:7713444 [本文引用: 1]
Abstract Two nuclear genes, Fr and Fr2, have been identified that restore pollen fertility to cytoplasmic male sterile (CMS) common bean (Phaseolus vulgaris L.) by apparently distinct mechanisms. Whereas Fr2 appears to suppress the expression of a male sterility associated mitochondrial sequence (designated pvs), Fr restores pollen fertility by causing the elimination of this unusual mitochondrial DNA-segment. To further investigate the mechanism of Fr action, Fr and Fr2 were cointroduced into the nucleus of a bean line containing the sterility inducing cytoplasm. When the effect of pvs was suppressed by Fr2, the presence of Fr no longer directed the elimination of the mitochondrial pvs sequence. This result suggests that the Fr function is dependent on proper expression of the pvs sequence. To evaluate the temporal and spatial patterns of Fr action, we undertook a polymerase chain reaction-based approach to trace the fate of the pvs sequence in different tissues of F2 and F3 fertile-restored plants derived from a genetic cross between a cytoplasmic male sterile line of common bean, CMS-Sprite (frfr), and fertility restorer line R351 (FrFr). We demonstrate that the Fr-directed disappearance of pvs sequence occurs during flower development. Elimination of the pvs sequence from developing megaspores results in permanent fertility restoration in the following generations. Genetic analysis demonstrated that permanent fertility restoration, that is, the complete elimination of pvs from reproductive tissues requires two doses of the Fr allele or the absence of fr in F2 individuals.(ABSTRACT TRUNCATED AT 250 WORDS)
[本文引用: 2]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
URLPMID:17249089 [本文引用: 1]
The genetics of fertility restoration of cms-C group cytoplasm of maize was studied using crosses involving stable maintainer lines and lines that restored full pollen fertility. Pollen fertility in the sources of cms-C sterile cytoplasms studied was restored by a single dominant restorer (Rf4) gene. The fertility restoration was sporophytic. Allelism tests among five restorer lines showed that they all apparently carried the same alleles (Rf4 Rf4). Similar tests also demonstrated that seven nonrestoring maintainer lines had apparently the same genotype (rf4 rf4), although a partial "late break" of fertility was observed at low levels in some maintainer crosses. Comparative studies among different cms-C sources (C, Bb, ES, PR and RB) indicated that similar inheritance of fertility restoration was involved. The data indicated that a single, dominant Rf gene is involved in the restoration of several C-group cytoplasms, at least in the lines studied here. This is the first single-gene, sporophytic restorer system described in maize to date.
URL [本文引用: 1]
Previous studies of the genetics of fertility restoration for C-type cytoplasmic male sterility (cms-C) in corn (Zea mays L.) have produced conflicting results. The objective of this study was to localize the restorer gene(s) for cms-C using restriction fragment length polymorphism (RFLP) analysis. Rf4, a single dominant restorer gene for cms-C found in inbred A619, was localized to Chromosome 8, almost-equal-to 2 centimorgans from an RFLP marker locus, NPI114A. The presence in corn of extensive nucleotide duplication complicated the analysis. Because the region of Chromosome 8 near Rf4 is duplicated on Chromosome 3, an additional restorer gene for cms-C may eventually be found on Chromosome 3.
URLPMID:16791701 [本文引用: 2]
The restoration of the C-type cytoplasmic male sterility (Cms) has been a common agriculture practice in the production of hybrid seed for many years. In this study, a series of crosses between select sterile and restorer lines, as well as a backcross population of (Cms-C7702×026233)02×026233, were used to investigate the restoration of C-type Cms. Our results demonstrated that there was an inhibitor of the Rf5 restorer gene. This inhibitor gene, Rf-I , maps to chromosome 7 and is tightly linked with SSR markers, umc2326 and umc2327, at a genetic distance 4.7 and 3.402cM, respectively. After analyzing our data combined with previous studies, we propose that the restoration of C-type Cms has two dominant genes, Rf4 and Rf5 . Rf4 has the ability to restore all genotypes of Cms-C lines; however, there exists an inhibitor for the other restorer gene, Rf5 ; thus, it can restore only those genotypes of Cms-C lines lacking the Rf-I inhibitor.
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
Cytoplasmic male sterility (CMS) is a maternally transmitted trait, whereby a plant is unable to produce viable pollen. Studies have revealed that this trait is a tool for enabling efficient and reliable coexistence between genetically modified (GM) and non-GM cultivation by biocontainment of GM maize (Zea mays L.) pollen. Maize has three types of male-sterile cytoplasm (T, S, and C), the fertility of which can be restored by nuclear rf genes or by interactions with the environment. Twenty-two CMS versions of modern European maize hybrids were evaluated in 17 environments in Switzerland, France, and Bulgaria, with two or three sowing dates, in 2005 and 2006. Stable and unstable male sterility occurred in all three CMS types. T-cytoplasm hybrids were the most stable, while S-cytoplasm hybrids often showed partial restoration of fertility. C-cytoplasm was similar to T-cytoplasm with regard to maintaining male sterility. Climatic factors, especially air temperature, evapotranspiration, and water vapor, during the 10 d before anthesis as well as during anthesis, were correlated positively or negatively with the partial reversion to male fertility of CMS hybrids, indicating an interaction between genetic and climatic factors. This study illustrates that T- and C-cytoplasm in particular open up viable prospects for containing transgenic pollen, especially for Bt-maize.
URL [本文引用: 1]
Cytoplasmic male sterility (CMS) is a maternally inherited trait that suppresses the production of viable pollen. CMS is a useful biological tool for confinement strategies to facilitate coexistence of genetically modified (GM) and non-GM crops in case where it is required. The trait is reversible and can be restored to fertility in the presence of nuclear restorer genes (Rf genes) and by environmental impacts. The aim of this study was to investigate the influence of the level of irrigation on the stability of CMS maize hybrids under defined greenhouse conditions. Additionally the combination of irrigation and air temperature was studied. Three CMS maize hybrids were grown with different levels of irrigation and in different temperature regimes. Tassel characteristics, pollen production and fertility were assessed. The CMS stability was high in hot air temperatures and decreased in lower temperatures. The level of irrigation had no major effect on the level of sterility. The extent of these phenomena was depending on the genotype of CMS maize and should be known before using CMS for coexistence purposes.
URL [本文引用: 1]
ABSTRACT While late-breaking partial fertility restoration of cytoplasmic male sterility is important both economically and theoretically, its inheritance is not well understood. Sublines of the maize inbred A632 differing in the level of restoration of C-cytoplasmic male sterility were identified. One line was nearly completely male sterile, whereas the others had varying levels of partial restoration; full fertility restoration was not observed. We crossed the fertile selections to the male sterile one and evaluated the progeny of the crosses to demonstrate the genetic nature of these differences. In two different experiments the percentage of plants that exhibited partial fertility was significantly different ( P =.01). In experiment I, the progeny of the cross A632-Pr-LB-1 A632-Pr-MS were 55% partially fertile while the progeny of A632-Pr-LB-2 A632-Pr-MS were 75% partially fertile. In experiment II, 3% of the plants exhibited partial fertility in one cross while 9% did so in the other cross. All differences were significant. Progeny tests were performed over years, and the effect of years on fertility restoration was highly significant. The number of fertile individuals was strongly affected by the environment with only 1.4% of the plants shedding pollen in 1984 and 13.0% shedding pollen in 1985. However, there was no environment by inbred interaction for fertility restoration. Due to low penetrance and strong environmental effects it was not possible to estimate the number of genes involved. However, it appears that this trait is under polygenic control.
Magsci [本文引用: 1]
<P>采用育性恢复专效性测定、PCR分类鉴定、类质粒鉴定3种玉米不育细胞质分类方法,对本室从爆裂玉米(<EM>Zea mays everta</EM>)种质中发现的玉米细胞质雄性不育材料“CMS-P”进行了胞质分类研究,一致表明CMS-P属于S型不育胞质。这为CMS-P的有效利用提供了理论依据。卡方测验表明,S组田间育性表现恢复的各组合花粉可染率并不完全符合1∶1的分离比例,CMS-P向可育方向偏移的程度更大。花粉可染率在不同测验系、不同胞质、不同环境间都表现出较大差异,胞质与核微效基因间的互作可能是影响育性稳定性的主要因素。</P>
Magsci [本文引用: 1]
<P>采用育性恢复专效性测定、PCR分类鉴定、类质粒鉴定3种玉米不育细胞质分类方法,对本室从爆裂玉米(<EM>Zea mays everta</EM>)种质中发现的玉米细胞质雄性不育材料“CMS-P”进行了胞质分类研究,一致表明CMS-P属于S型不育胞质。这为CMS-P的有效利用提供了理论依据。卡方测验表明,S组田间育性表现恢复的各组合花粉可染率并不完全符合1∶1的分离比例,CMS-P向可育方向偏移的程度更大。花粉可染率在不同测验系、不同胞质、不同环境间都表现出较大差异,胞质与核微效基因间的互作可能是影响育性稳定性的主要因素。</P>
[本文引用: 1]
[本文引用: 1]
URLPMID:21479555 [本文引用: 2]
Partial restoration of male fertility limits the use of C-type cytoplasmic male sterility (C-CMS) for the production of hybrid seeds in maize. Nevertheless, the genetic basis of the trait is still unknown. Therefore, the aim to this study was to identify genomic regions that govern partial restoration by means of a QTL analysis carried out in an F 2 population ( n = 180). This population was derived from the Corn Belt inbred lines B37C and K55. F 2 BC 1 progenies were phenotyped at three locations in Switzerland. Male fertility was rated according to the quality and number of anthers as well as the anthesis-silking interval. A weak effect of environment on the expression of partial restoration was reflected by high heritabilities of all fertility-related traits. Partial restoration was inherited like an oligogenic trait. Three major QTL regions were found consistently across environments in the chromosomal bins 2.09, 3.06 and 7.03. Therefore, a marker-assisted counter-selection of partial restoration is promising. Minor QTL regions were found on chromosomes 3, 4, 5, 6 and 8. A combination of partial restorer alleles at different QTL can lead to full restoration of fertility. The maternal parent was clearly involved in the partial restoration, because the restorer alleles at QTL in bins 2.09, 6.04 and 7.03 originated from B37. The three major QTL regions collocated with other restorer genes of maize, a phenomenon, which seems to be typical for restorer genes. Therefore, a study of the clusters of restorer genes in maize could lead to a better understanding of their evolution and function. In this respect, the long arm of chromosome 2 is particularly interesting, because it harbors restorer genes for the three major CMS systems (C, T and S) of maize.
URL [本文引用: 2]
This chapter discusses the general field of cytoplasmic inheritance of pollen sterility. In five discoveries, the expression of the cytoplasmically induced sterility was influenced by nuclear genes. Investigation of the genetics of fertility restoration of both the United States Department of Agriculture (USDA) and the Texas types of cytoplasmic sterility has shown that in either case a single dominant gene is sufficient to restore pollen fertility to most cytoplasmic male-sterile inbreds although there are complications with certain genetic backgrounds and in certain environments. Specific tester genotypes enable one to differentiate USDA from Texas cytoplasms because some genotypes are sterile in one cytoplasm but not the other, some are sterile in both, and some are pollen fertile in both. This type of test can be applied to all discoveries of cytoplasmic male sterility to see if they resemble either the USDA or the Texas source or if they have a different spectrum of interactions with genotype. Cytoplasmic male sterility in corn is characterized by having two distinct forms, the USDA and the Texas. Both types interact with the nucleus to the extent that their sterility-inducing potential is revealed only by certain genes, and the number of the genes is relatively few for each cytoplasm. The Texas type has been shown to affect, to a small degree, the rate of growth and final size of the corn plant, independently of its effect on pollen sterility.
[本文引用: 1]
[本文引用: 1]
Magsci [本文引用: 1]
对3个 C 型不育系及其同型保持系和4个恢复系与这3个不育系组配的12个 F_1的花药发育过程进行了观察,以了解 C 型不育系雄性败育的细胞学机理。观察结果表明,不同核背景的不育系,其雄性败育的细胞学特点各异。Cms-C Mo17的花药绒毡层细胞为双核型。该层细胞在减数分裂的四分体时期开始液泡化,并呈辐射状膨大,至小孢子的单核前
Magsci [本文引用: 1]
对3个 C 型不育系及其同型保持系和4个恢复系与这3个不育系组配的12个 F_1的花药发育过程进行了观察,以了解 C 型不育系雄性败育的细胞学机理。观察结果表明,不同核背景的不育系,其雄性败育的细胞学特点各异。Cms-C Mo17的花药绒毡层细胞为双核型。该层细胞在减数分裂的四分体时期开始液泡化,并呈辐射状膨大,至小孢子的单核前
URLMagsci [本文引用: 2]
杂交水稻不育一恢复基因的遗传行为比较复杂,现有的关于该方面研究的报道不足以解释杂交水稻不育一恢复基因的遗传机理。本文提出杂交水稻不育一恢复基因的阑性状和剂量效应假说,其主要论点有二:(1)恢复基因只有剂量效应,没有显隐性之分,一个品种恢复力的强弱依赖于该品种携带恢复基因的剂量;(2)不育一恢复这一性状属于闻性状,是核质互作的结果,当合子中恢复基因的剂量达到某rA值时,激活了不育系细胞质的育性,因此对于某一不育系来说,其相应的恢保关系可以分为三大类:没有恢复能力的L型品种,可作为保持系;具有完全恢复能力的H型品种,可以作为恢复系;具有部分恢复能力的M型品种。根据此假说得到的推论能够解释杂交水稻不育一恢复基因的遗传行为。
URLMagsci [本文引用: 2]
杂交水稻不育一恢复基因的遗传行为比较复杂,现有的关于该方面研究的报道不足以解释杂交水稻不育一恢复基因的遗传机理。本文提出杂交水稻不育一恢复基因的阑性状和剂量效应假说,其主要论点有二:(1)恢复基因只有剂量效应,没有显隐性之分,一个品种恢复力的强弱依赖于该品种携带恢复基因的剂量;(2)不育一恢复这一性状属于闻性状,是核质互作的结果,当合子中恢复基因的剂量达到某rA值时,激活了不育系细胞质的育性,因此对于某一不育系来说,其相应的恢保关系可以分为三大类:没有恢复能力的L型品种,可作为保持系;具有完全恢复能力的H型品种,可以作为恢复系;具有部分恢复能力的M型品种。根据此假说得到的推论能够解释杂交水稻不育一恢复基因的遗传行为。
URLPMID:29177892 [本文引用: 1]
Abstract Hybrid wheat breeding has the potential to significantly increase wheat productivity compared to line breeding. The induction of male sterility by the cytoplasm of Triticum timopheevii Zhuk. is a widely discussed approach to ensure cross-pollination between parental inbred lines in hybrid wheat seed production. As fertility restoration in hybrids with this cytoplasm is often incomplete, understanding the underlying genetics is a prerequisite to apply this technology. A promising component for fertility restoration is the restorer locus Rf1, which was first detected on chromosome 1A of the restorer accession R3. In the present study, we performed quantitative trait locus (QTL) analyses to locate Rf1 and estimate its effect in populations involving the restorer lines R3, R113 and L19. Molecular markers linked to Rf1 in these populations were used to analyse the genomic target region in T. timopheevii accessions and common wheat breeding lines. The QTL analyses revealed that Rf1 interacted with a modifier locus on chromosome 1BS and the restorer locus Rf4 on chromosome 6B. The modifier locus significantly influenced both the penetrance and expressivity of Rf1. Whereas Rf1 exhibited expressivity higher than that of Rf4, the effects of these loci were not additive. Evaluating the marker haplotype for the Rf1 region, we propose that the restoring Rf1 allele may be derived exclusively from T. timopheevii. The present study demonstrates that interactions between restorer and modifier loci play a critical role in fertility restoration of common wheat with the cytoplasm of T. timopheevii.
[本文引用: 1]
[本文引用: 1]
URLPMID:4280902 [本文引用: 1]
Abstract In plants, male reproductive development is extremely sensitive to adverse climatic environments and (a)biotic stress. Upon exposure to stress, male gametophytic organs often show morphological, structural and metabolic alterations that typically lead to meiotic defects or premature spore abortion and male reproductive sterility. Depending on the type of stress involved (e.g. heat, cold, drought) and the duration of stress exposure, the underlying cellular defect is highly variable and either involves cytoskeletal alterations, tapetal irregularities, altered sugar utilization, aberrations in auxin metabolism, accumulation of reactive oxygen species (ROS; oxidative stress) or the ectopic induction of programmed cell death (PCD). In this review, we present the critically stress-sensitive stages of male sporogenesis (meiosis) and male gametogenesis (microspore development), and discuss the corresponding biological processes involved and the resulting alterations in male reproduction. In addition, this review also provides insights into the molecular and/or hormonal regulation of the environmental stress sensitivity of male reproduction and outlines putative interaction(s) between the different processes involved. 2013 The Authors. Plant, Cell & Environment published by John Wiley & Sons Ltd.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}