哈斯木其尔1,2, 张学耀1,2, 牛国祥1,2, 王银柳1,2, 黄建辉1,*, 1中国科学院植物研究所, 植被与环境变化国家重点实验室, 北京 1000932中国科学院大学生命科学学院, 北京 100049 HasiMuqier1,2, ZhangXueyao1,2, NiuGuoxiang1,2, WangYinliu1,2, HuangJianhui1,*, 1State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China2College of Life Sciences, University of Chinese Academy of Sciences, Beijing 100049, China 引用本文 哈斯木其尔, 张学耀, 牛国祥, 王银柳, 黄建辉. 氮素添加对内蒙古草甸草原生态系统CO交换的影响. 植物学报, 2018, 53(1): 27-41
Abstract: Increasing nitrogen deposition influences carbon sequestration in grassland ecosystems, but we have no consistent results on how it impacts the exchange of CO2 at the ecosystem level. As well, the impact of different types of N fertilizers and rates of N increase are not clear yet. This study aimed to evaluate the effect of N addition on ecosystem CO2 exchange and was performed in a meadow steppe in Erguna, Inner Mongolia. A field experiment compared two types of N fertilizers (urea and slow-release urea) with five N addition rates (0, 5.0, 10.0, 20.0 and 50.0 g N·m-2·a-1). At the start and middle stage of the growing season, N addition had weak and inhibitive effects on ecosystem CO2 exchange when precipitation was low, and significantly increased ecosystem CO2 exchange in the late growing season when precipitation was high. Both net ecosystem CO2 exchange and gross ecosystem photosynthesis increased significantly with N addition rate but showed a tendency of saturation when the N addition rate reached 10 g N·m-2·a-1. The two types of N fertilizers resulted in slightly different responses of ecosystem CO2 exchange: slow-release urea had a stronger positive effect at 5 g N·m-2·a-1 with no significant difference at other addition rates. Our study suggests that increasing N deposition has significant effects on carbon assimilation in this semi-arid grassland, but the direction and magnitude of effects are strongly affected by seasonal pattern and amount of precipitation. The effects of different types of N fertilizers (i.e., urea and slow-release urea) on ecosystem CO2 exchange may differ.
Key words:nitrogen addition ; net ecosystem CO2 exchange ; ecosystem respiration ; gross ecosystem photosynthesis ; meadow steppe
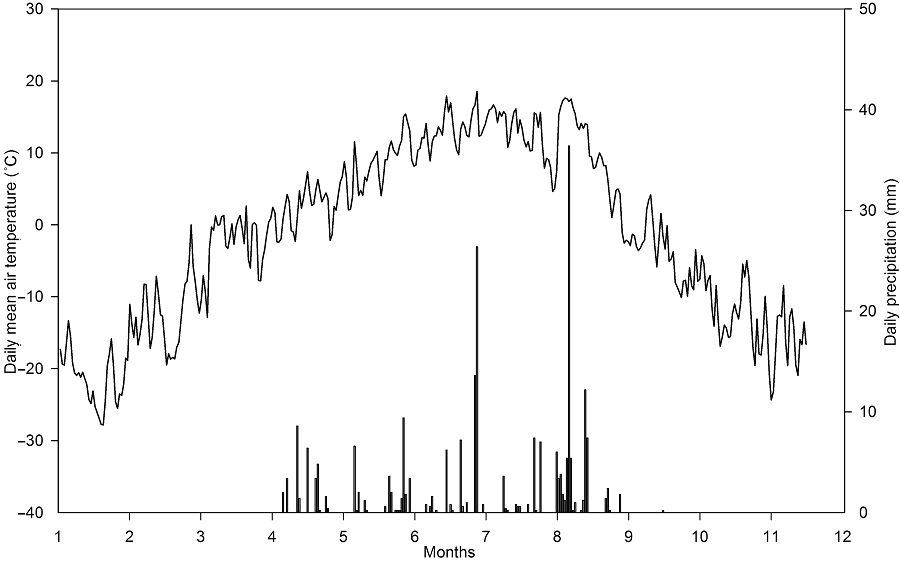
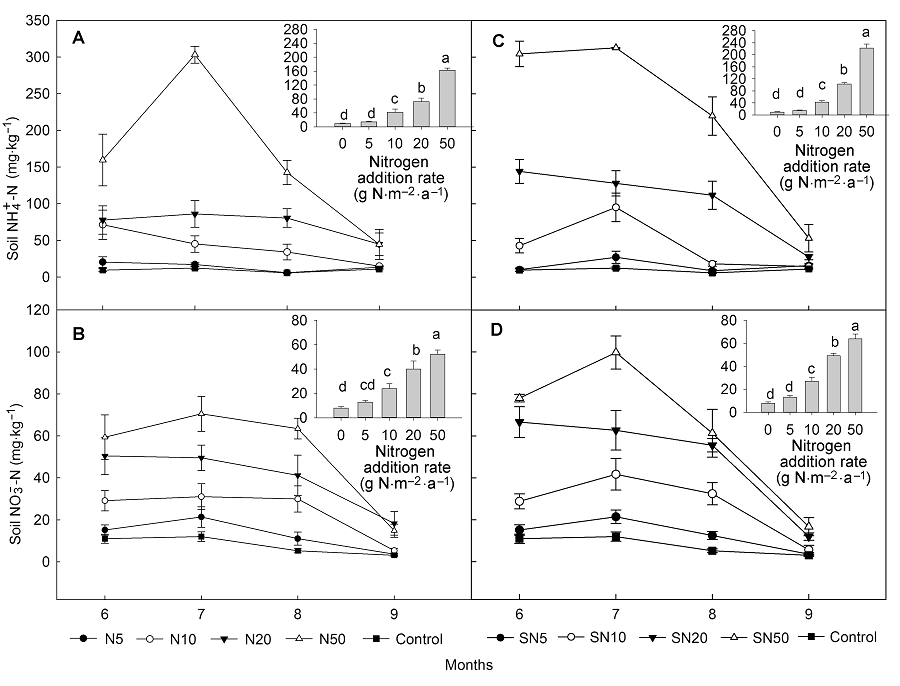
2 结果与分析2.1 非生物与生物因子研究样地2016年总降雨量为238.4 mm, 生长季(6-9月)降雨量为194 mm, 日降雨量季节动态表现为双峰曲线, 峰值分别出现在7月和9月(图1), 且降水较多出现在生长季末(9月降雨为83.6 mm)而使得生长季前期较为干旱(6-8月降雨量为110.4 mm)。日平均温度的变化表现为单峰曲线, 最大值出现在7月和8月(图1)。 与对照相比, 尿素和缓释尿素处理对土壤含水量及土壤温度影响均不显著(表1, 表2)。土壤无机氮含量在施肥量(尿素和缓释尿素)为5 g N·m-2·a-1时与对照相比没有显著差异, 但是在10、20和50 g N·m-2·a-1处理时显著增加。具体表现为尿素处理时铵态氮分别增加332.15%、654.29%和1596.23%, 硝态氮分别增加204.45%、408.74%和564.06%; 缓释尿素处理时铵态氮分别增加344.55%、971.70%和2221.26%, 硝态氮分别增加246.51%、527.67%和715.85% (图2)。随着施氮(尿素和缓释尿素)水平的增加, 土壤pH值出现降低趋势, 当尿素处理达到50 g N·m-2·a-1水平、缓释尿素达到20 g N·m-2·a-1水平时, 土壤pH值降低达显著水平(图3)。 与对照相比, 尿素处理时地上总生物量均表现出增加的趋势, 但在统计学上不显著。缓释尿素处理时地上总生物量显著增加, 表现为当施肥量达到10、20和50 g N·m-2·a-1时, 地上总生物量分别显著增加了67.23%、83.17%和78.50%。将地上生物量分为禾草生物量与杂类草生物量时, 尿素对禾草生物量的影响表现出增加的趋势, 对杂类草生物量的影响表现为降低趋势, 但在统计学上均不显著; 缓释尿素显著增加了禾草生物量, 表现为当施肥量达到10、20和50 g N·m-2·a-1时, 禾草生物量分别显著增加了94.50%、128.03%和134.44%, 但杂类草生物量没有显著变化(表3)。在20 g N·m-2·a-1尿素处理时, 地下总生物量增加了35.06% (P<0.05), 而在10 g N·m-2·a-1缓释尿素处理时, 地下生物量增加了23.80% (P<0.05), 其 它施氮浓度均表现为地下生物量增加趋势, 但在统计学上不显著 (表3)。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_2.png<b>图1</b> 2016年研究样地日降雨量(柱形)与日均气温(线形)动态<br/><b>Figure 1</b> Daily precipitation (bars) and daily mean air temperature (line) of the research plots in 2016 Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_2.png<b>图1</b> 2016年研究样地日降雨量(柱形)与日均气温(线形)动态<br/><b>Figure 1</b> Daily precipitation (bars) and daily mean air temperature (line) of the research plots in 2016
图1 2016年研究样地日降雨量(柱形)与日均气温(线形)动态 Figure 1 Daily precipitation (bars) and daily mean air temperature (line) of the research plots in 2016
表1 Table 1 表1 表1 生长季不同氮素添加水平下土壤温度、土壤含水量、生态系统净CO2交换、生态系统呼吸和生态系统总光合的季节均值 Table 1 Means of growing season soil temperature, soil moisture, net ecosystem CO2 exchange, ecosystem respiration and gross ecosystem photosynthesis under different nitrogen addition rates
N addition rates (g N·m-2·a-1)
θν
Ts
NEE
ER
GEP
Urea
0
11.48a±0.74
17.94a±0.66
-2.98a±0.38
3.85ab±0.12
6.81ab±0.48
5
12.79a±0.97
17.69a±0.76
-3.13a±0.49
3.35a±0.22
6.48a±0.31
10
12.29a±0.61
17.66a±0.32
-3.77a±0.32
4.10ab±0.42
7.88b±0.29
20
13.79a±0.47
17.71a±0.49
-3.82a±0.27
4.23ab±0.23
8.05b±0.19
50
13.41a±0.71
17.22a±0.22
-2.88a±0.44
4.59b±0.33
7.48ab±0.61
Slow-release urea
0
11.48a±0.74
17.94a±0.66
-2.98b±0.38
3.85a±0.12
6.81a±0.48
5
12.01a±0.74
18.07a±0.35
-3.77ab±0.32
3.81a±0.25
7.58a±0.19
10
12.51a±0.55
17.72a±0.35
-4.80a±0.39
4.29a±0.32
9.10b±0.39
20
14.49a±1.28
17.06a±0.27
-3.15b±0.46
4.19a±0.37
7.34a±0.23
50
14.26a±0.82
17.44a±0.42
-2.93b±0.54
4.55a±0.27
7.47a±0.40
Ts: Soil temperature (°C); θν: Soil moisture (%); NEE: Net ecosystem CO2 exchange; ER: Ecosystem respiration; GEP: Gross ecosystem photosynthesis. Data were means±SE (n=4); Different lowercase letters in a column indicate significant differences among different N addition rates (P<0.05). Ts: 土壤温度(°C); θν: 土壤含水量(%); NEE: 生态系统净CO2交换; ER: 生态系统呼吸; GEP: 生态系统总光合。数据为平均值±标准误(n=4); 同列不同小写字母表示处理间差异显著(P<0.05)。
表1 生长季不同氮素添加水平下土壤温度、土壤含水量、生态系统净CO2交换、生态系统呼吸和生态系统总光合的季节均值 Table 1 Means of growing season soil temperature, soil moisture, net ecosystem CO2 exchange, ecosystem respiration and gross ecosystem photosynthesis under different nitrogen addition rates
表2 Table 2 表2 表2 生长季测定时间和氮肥(尿素和缓释尿素) 2个因素对土壤温度、土壤含水量、生态系统净CO2交换、生态系统呼吸和生态系统总光合的主效应及其交互效应的重复测定方差分析(P值) Table 2 Results (P-values) of repeated-measures ANOVA on the effects of sampling time, N addition (urea and slow-release urea), and their interactions on soil temperature, soil moisture, net ecosystem CO2 exchange, ecosystem respiration and gross ecosystem photosynthesis during the growing season
θν
Ts
NEE
ER
GEP
Urea
Time
<0.001
<0.001
<0.001
<0.001
<0.001
N
0.22
0.91
0.29
0.08
0.07
Time×N
0.66
0.56
0.01
0.01
0.03
Slow-release urea
Time
<0.001
<0.001
<0.001
<0.001
<0.001
SN
0.38
0.49
0.04
0.34
0.01
Time×SN
0.63
0.74
0.003
0.15
0.002
Ts、θν、NEE、ER和GEP同表1。Ts, θν, NEE, ER and GEP see Table 1.
表2 生长季测定时间和氮肥(尿素和缓释尿素) 2个因素对土壤温度、土壤含水量、生态系统净CO2交换、生态系统呼吸和生态系统总光合的主效应及其交互效应的重复测定方差分析(P值) Table 2 Results (P-values) of repeated-measures ANOVA on the effects of sampling time, N addition (urea and slow-release urea), and their interactions on soil temperature, soil moisture, net ecosystem CO2 exchange, ecosystem respiration and gross ecosystem photosynthesis during the growing season
图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_3.png<b>图2</b> 土壤表层(0-10 cm)铵态氮和硝态氮的季节动态<br/>(A), (B) 添加尿素; (C), (D) 添加缓释尿素。小图为添加不同剂量氮素后土壤氮素含量的季节均值(平均值±标准误, <i>n</i>=4)。不同小写字母表示处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 2</b> Seasonal dynamics of soil NH<sub>4</sub><sup>+</sup>-N and NO<sub>3</sub><sup>-</sup>-N at 0-10 cm depth<br/>(A), (B) Addition with urea; (C), (D) Addition with slow-release urea. In-set figures showed mean soil inorganic N contents with addition of N (means±SE, <i>n</i>=4). Different lowercase letters among columns indicate significant differences among treatments (<i>P</i><0.05). Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_3.png<b>图2</b> 土壤表层(0-10 cm)铵态氮和硝态氮的季节动态<br/>(A), (B) 添加尿素; (C), (D) 添加缓释尿素。小图为添加不同剂量氮素后土壤氮素含量的季节均值(平均值±标准误, <i>n</i>=4)。不同小写字母表示处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 2</b> Seasonal dynamics of soil NH<sub>4</sub><sup>+</sup>-N and NO<sub>3</sub><sup>-</sup>-N at 0-10 cm depth<br/>(A), (B) Addition with urea; (C), (D) Addition with slow-release urea. In-set figures showed mean soil inorganic N contents with addition of N (means±SE, <i>n</i>=4). Different lowercase letters among columns indicate significant differences among treatments (<i>P</i><0.05).
图2 土壤表层(0-10 cm)铵态氮和硝态氮的季节动态 (A), (B) 添加尿素; (C), (D) 添加缓释尿素。小图为添加不同剂量氮素后土壤氮素含量的季节均值(平均值±标准误, n=4)。不同小写字母表示处理间差异显著(P<0.05)。 Figure 2 Seasonal dynamics of soil NH4+-N and NO3--N at 0-10 cm depth (A), (B) Addition with urea; (C), (D) Addition with slow-release urea. In-set figures showed mean soil inorganic N contents with addition of N (means±SE, n=4). Different lowercase letters among columns indicate significant differences among treatments (P<0.05).
图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_4.png<b>图3</b> 表层(0-10 cm)土壤pH值季节均值(平均值±标准误, <i>n</i>=4)<br/>(A) 添加尿素; (B) 添加缓释尿素。不同小写字母表示处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 3</b> Seasonal means of pH at 0-10 cm soil (means±SE, <i>n</i>=4)<br/>(A) Addition with urea; (B) Addition with slow-release urea. Different lowercase letters among columns indicate significant differences among treatments (<i>P</i><0.05). Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_4.png<b>图3</b> 表层(0-10 cm)土壤pH值季节均值(平均值±标准误, <i>n</i>=4)<br/>(A) 添加尿素; (B) 添加缓释尿素。不同小写字母表示处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 3</b> Seasonal means of pH at 0-10 cm soil (means±SE, <i>n</i>=4)<br/>(A) Addition with urea; (B) Addition with slow-release urea. Different lowercase letters among columns indicate significant differences among treatments (<i>P</i><0.05).
图3 表层(0-10 cm)土壤pH值季节均值(平均值±标准误, n=4) (A) 添加尿素; (B) 添加缓释尿素。不同小写字母表示处理间差异显著(P<0.05)。 Figure 3 Seasonal means of pH at 0-10 cm soil (means±SE, n=4) (A) Addition with urea; (B) Addition with slow-release urea. Different lowercase letters among columns indicate significant differences among treatments (P<0.05).
表3 Table 3 表3 表3 氮素添加水平分别对地上净初级生产力、禾草类生物量、杂类草生物量和地下总生物量影响的单因素方差分析结果(平均值±标准误, n=4) Table 3 Aboveground net primary productivity, grasses aboveground biomass, forb aboveground biomass and total belowground biomass under different nitrogen addition rates (means±SE, n=4)
N addition rates (g N·m-2·a-1)
ANPP
GB
FB
BGB
Urea
0
142.52a±16.85
96.86a±19.68
42.37a±13.05
1718.44a±172.36
5
157.74a±5.51
129.37a±11.10
24.33a±8.48
1575.31a±64.48
10
216.29a±23.24
190.59a±19.71
24.07a±5.09
1941.26ab±226.12
20
167.54a±21.06
125.55a±38.81
37.01a±16.07
2320.85b±99.68
50
191.38a±40.72
182.04a±42.34
10.26a±3.16
1915.27ab±74.55
Slow-release urea
0
142.52a±16.85
96.86a±19.68
42.37a±13.05
1718.44a±172.36
5
166.24ab±11.86
129.16ab±10.48
28.49a±3.39
1632.48a±67.81
10
238.34bc±35.71
188.39bc±45.45
50.16a±25.57
2127.48b±111.51
20
261.06c±26.01
220.86c±19.61
40.48a±7.82
1942.56ab±123.40
50
254.41c±24.63
227.08c±16.94
31.36a±16.22
1750.71ab±105.22
ANPP: Aboveground net primary productivity; GB: Grasses aboveground biomass; FB: Forb aboveground biomass; BGB: Total belowground biomass. Different lowercase letters in a column indicate significant differences among different N addition rates (P<0.05). ANPP: 净初级生产力; GB: 禾草类生物量; FB: 杂类草生物量; BGB: 地下总生物量。每列不同小写字母表示处理间差异显著(P<0.05)。
表3 氮素添加水平分别对地上净初级生产力、禾草类生物量、杂类草生物量和地下总生物量影响的单因素方差分析结果(平均值±标准误, n=4) Table 3 Aboveground net primary productivity, grasses aboveground biomass, forb aboveground biomass and total belowground biomass under different nitrogen addition rates (means±SE, n=4)
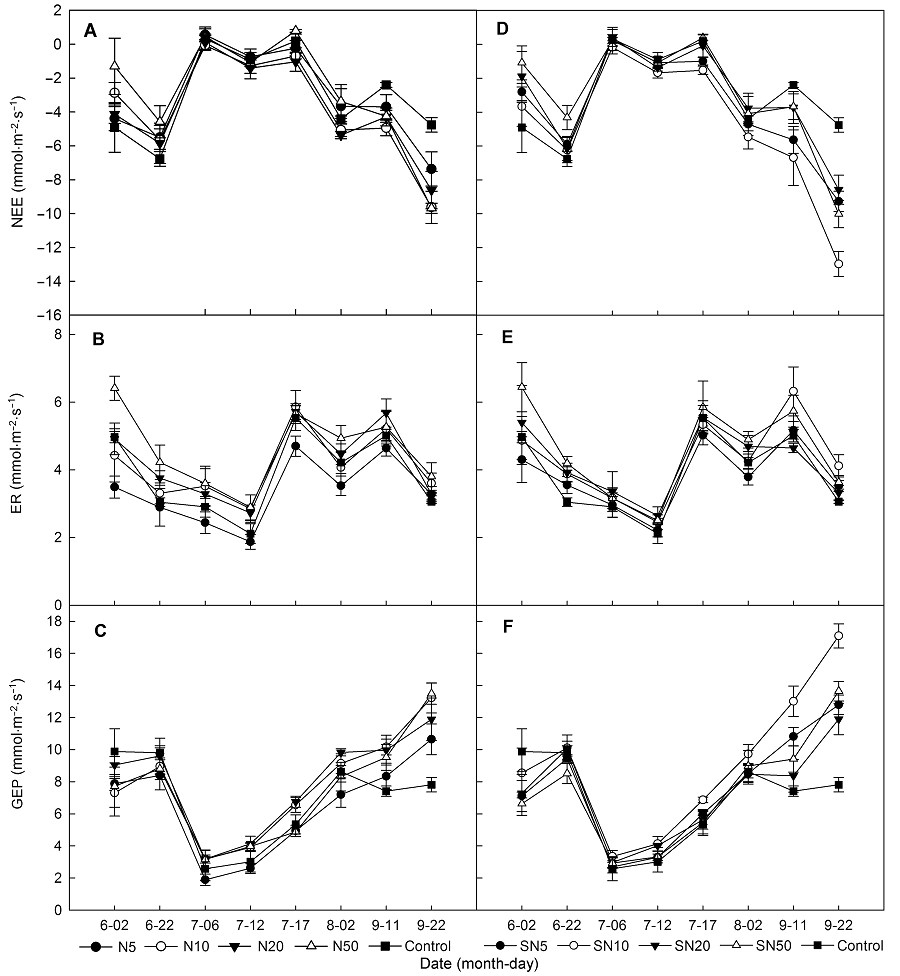
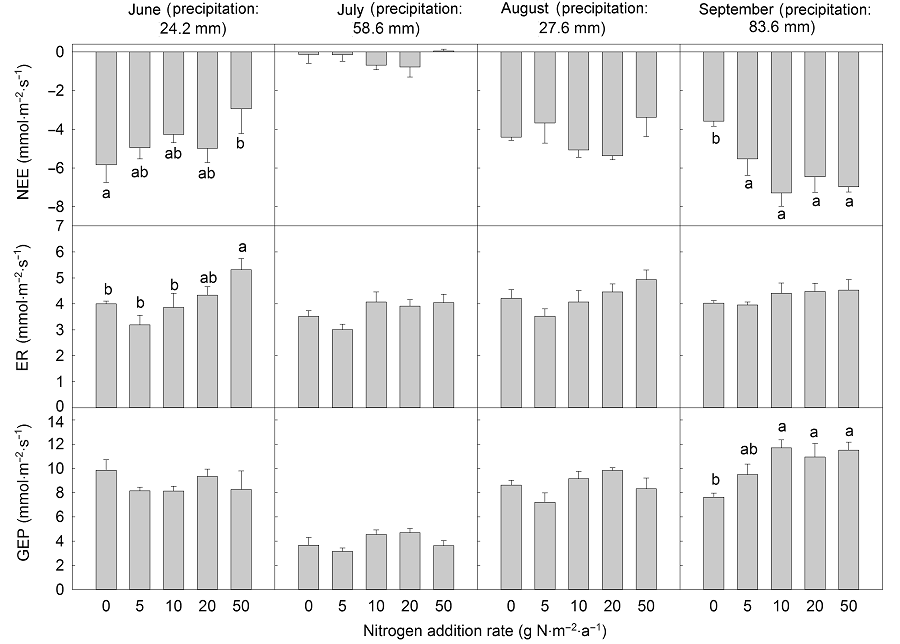
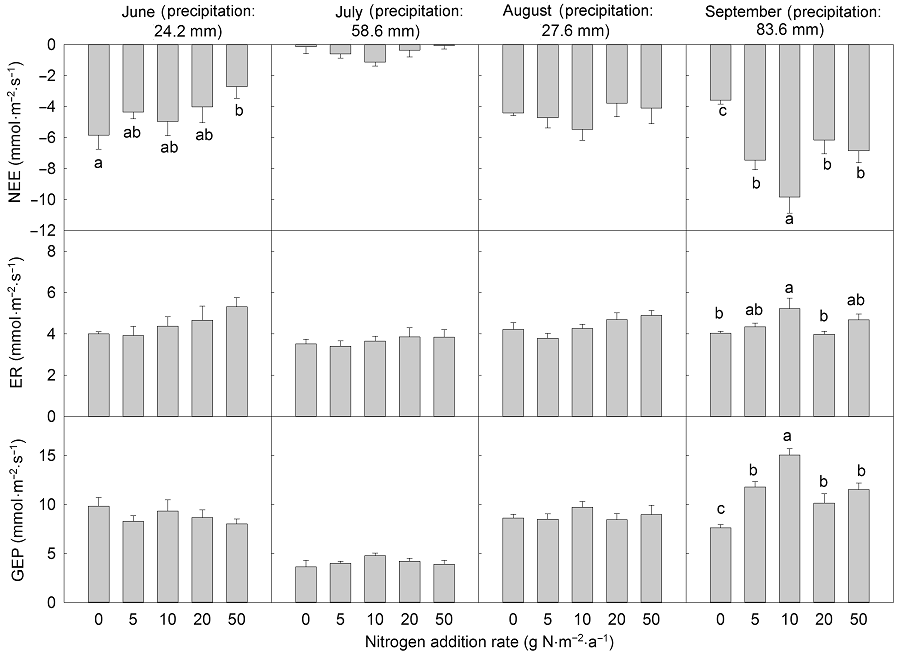
2.2 氮素处理对生态系统CO2交换的影响在整个生长季测定时间内, 净生态系统CO2交换最大值出现在生长季末的9月(图4)。重复测定方差分析结果表明, 尿素处理对NEE、ER和GEP没有显著影响, 添加缓释尿素时NEE和GEP显著增加。进一步的单因素多重比较分析结果显示, 缓释尿素10 g N·m-2·a-1处理显著增加了NEE和GEP, 分别增加62.59%和33.64%, 但对ER的影响不显著(表1, 表2)。月动态分析结果(图5, 图6)显示, 生长季初期(6月)氮素(尿素和缓释尿素)添加浓度为50 g N·m-2·a-1时, ER分别显著增加了32.77%和32.72%, GEP没有显著变化, NEE分别显著降低49.63%和53.65%, 其它氮素添加水平对ER具有增加趋势, 对GEP具降低趋势, 但在统计学上没有显著差异。生长季中期(7月和8月)氮素(尿素和缓释尿素)添加对NEE、ER和GEP均没有显著影响。生长季末期(9月)随着氮素(尿素和缓释尿素)添加量的增加, NEE和GEP显著增加。不同尿素添加水平下(5.0、10.0、20.0和50.0 g N·m-2·a-1), NEE分别显著增加了54.33%、103.38%、80.06%和94.46%, GEP除了5 g N·m-2·a-1处理外, 其余处理(10.0、20.0和50.0 g N·m-2·a-1)分别显著增加53.52%、43.52%和51%; 不同剂量缓释尿素处理下, NEE分别显著增加了108.15%、174.33%、71.76%和91.11%, GEP分别显著增加了54.97%、97.57%、33.08%和51.38%。无论是尿素还是缓释尿素, 当氮素添加量达到10 g N·m-2·a-1时, NEE和GEP的响应趋于饱和, 即随着氮素添加水平的进一步提高, 其NEE和GEP的增加趋势减弱。ER对尿素处理的响应不显著, 对缓释尿素处理仅在10 g N·m-2·a-1氮素添加水平时显著增加了29.37% (图5, 图6)。配对T检验结果表明, 2种氮肥(尿素和缓释尿素)对生态系统CO2的影响在施氮量为5 g N·m-2·a-1时差异显著, 缓释尿素对NEE和GEP的促进作用显著大于尿素(P<0.05), 在10和20 g N·m-2·a-1处理水平下缓释尿素效应边缘显著大于尿素(P<0.01), 当施肥量为50 g N·m-2·a-1时, 2种氮肥的影响没有显著差异。当施肥量为5 g N·m-2·a-1时, 缓释尿素对ER的影响显著大于尿素(P<0.05), 其它施氮水平时2种氮肥对生态系统CO2的影响没有显著差异。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_5.png<b>图4</b> 不同氮素添加浓度下生态系统净CO<sub>2</sub>交换、生态系统呼吸和生态系统总光合季节动态(平均值±标准误)<br/>(A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。<br/><b>Figure 4</b> Seasonal variations in net ecosystem CO<sub>2</sub> exchange, ecosystem respiration and gross ecosystem photosynthesis under different nitrogen addition rates (means±SE)<br/>(A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>. Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_5.png<b>图4</b> 不同氮素添加浓度下生态系统净CO<sub>2</sub>交换、生态系统呼吸和生态系统总光合季节动态(平均值±标准误)<br/>(A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。<br/><b>Figure 4</b> Seasonal variations in net ecosystem CO<sub>2</sub> exchange, ecosystem respiration and gross ecosystem photosynthesis under different nitrogen addition rates (means±SE)<br/>(A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>.
图4 不同氮素添加浓度下生态系统净CO2交换、生态系统呼吸和生态系统总光合季节动态(平均值±标准误) (A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同表1。 Figure 4 Seasonal variations in net ecosystem CO2 exchange, ecosystem respiration and gross ecosystem photosynthesis under different nitrogen addition rates (means±SE) (A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see Table 1.
图5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_6.png<b>图5</b> 不同浓度尿素处理时生态系统净CO<sub>2</sub>交换、生态系统呼吸和生态系统总光合月动态(平均值±标准误, <i>n</i>=4)<br/>NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。不同小写字母表示处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 5</b> Seasonal patterns of net ecosystem CO<sub>2</sub> exchange, ecosystem respiration and gross ecosystem photosynthesis at different nitrogen addition rates with urea in each month (means±SE, <i>n</i>=4)<br/>NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>. Different lowercase letters among columns indicate significant differences among treatments (<i>P</i><0.05). Figure 5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_6.png<b>图5</b> 不同浓度尿素处理时生态系统净CO<sub>2</sub>交换、生态系统呼吸和生态系统总光合月动态(平均值±标准误, <i>n</i>=4)<br/>NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。不同小写字母表示处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 5</b> Seasonal patterns of net ecosystem CO<sub>2</sub> exchange, ecosystem respiration and gross ecosystem photosynthesis at different nitrogen addition rates with urea in each month (means±SE, <i>n</i>=4)<br/>NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>. Different lowercase letters among columns indicate significant differences among treatments (<i>P</i><0.05).
图5 不同浓度尿素处理时生态系统净CO2交换、生态系统呼吸和生态系统总光合月动态(平均值±标准误, n=4) NEE、ER和GEP同表1。不同小写字母表示处理间差异显著(P<0.05)。 Figure 5 Seasonal patterns of net ecosystem CO2 exchange, ecosystem respiration and gross ecosystem photosynthesis at different nitrogen addition rates with urea in each month (means±SE, n=4) NEE, ER and GEP see Table 1. Different lowercase letters among columns indicate significant differences among treatments (P<0.05).
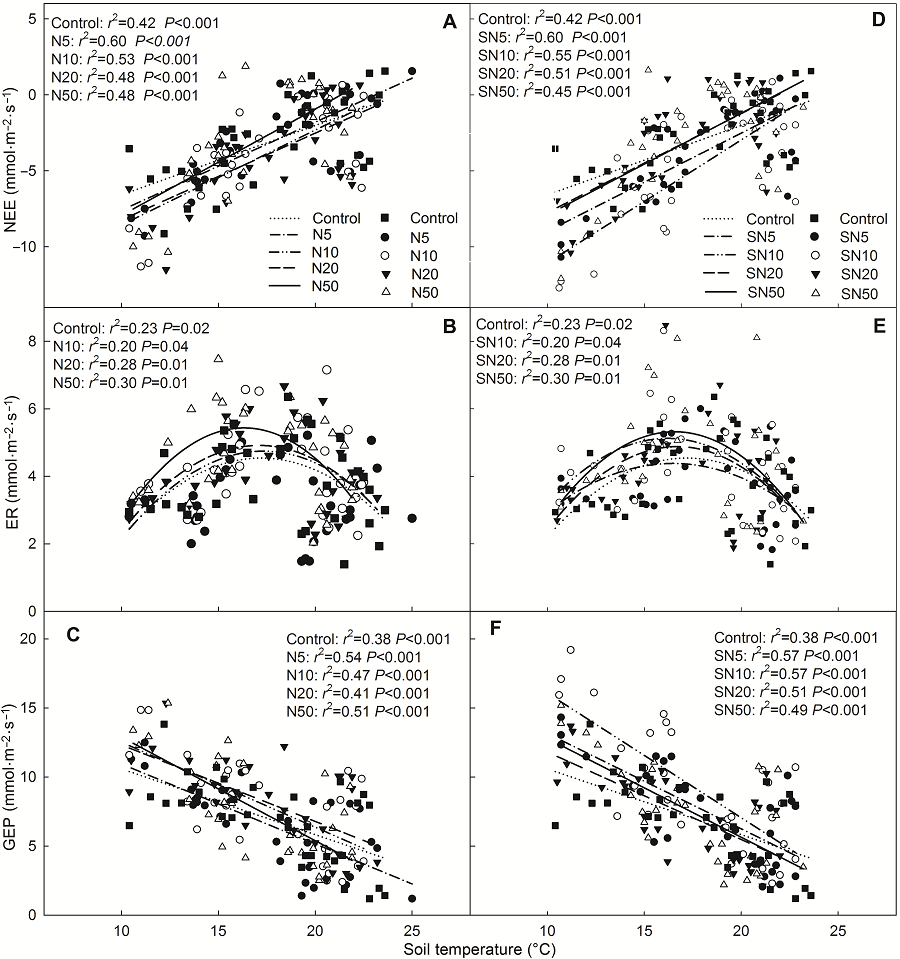
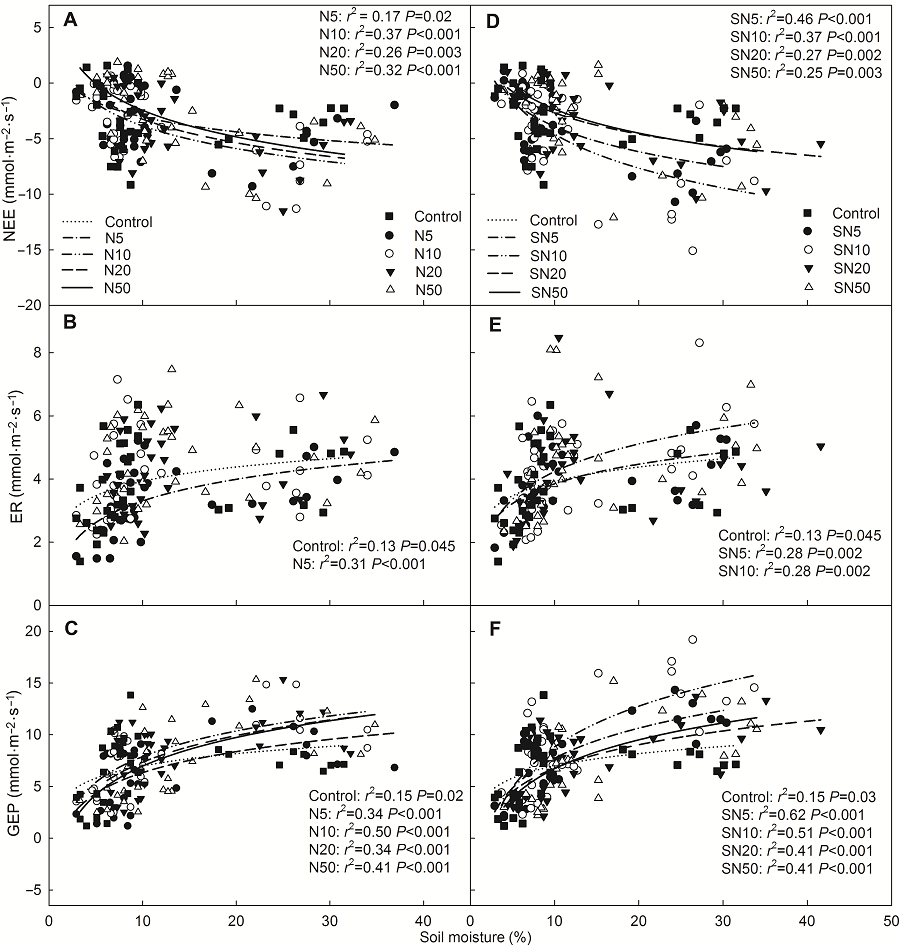
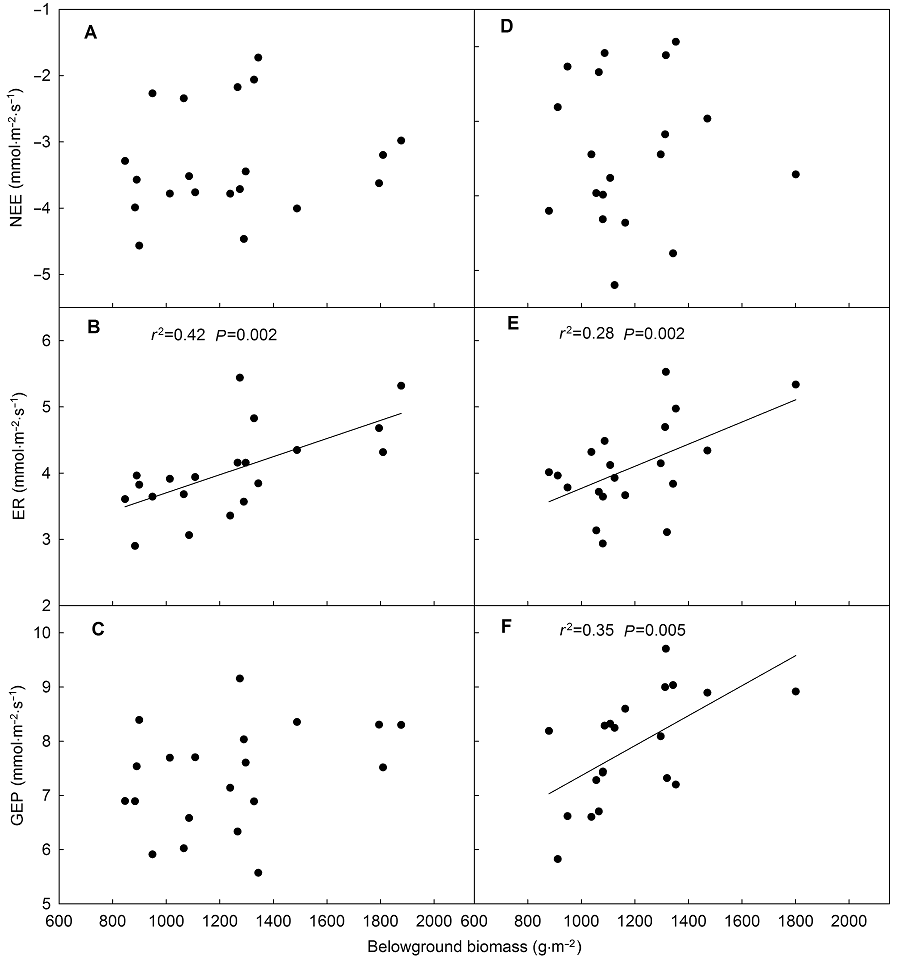
2.3 环境因子对生态系统CO2交换的调控本研究对该地区生态系统CO2交换与土壤温度和土壤含水量之间的关系进行了分析。结果显示, 氮素(尿素和缓释尿素)添加时NEE和GEP随着土壤温度的增加呈线性增加, ER则呈先增加后降低的二项式关系(图7), ER最适温度范围为15-20.6°C, 低于或高于这个范围时ER具有降低趋势。NEE、ER和GEP与土壤含水量之间存在显著对数关系(y=aLn(x)+b) (图8)。该方程中斜率(a值)被认为是生态系统CO2交换对土壤含水量的敏感系数(Yan et al., 2011)。因此, GEP的水分敏感性显著高于ER的水分敏感性(P<0.05); 氮素添加(无论是尿素还是缓释尿素)显著增加了GEP的水分敏感性(表4)。在尿素处理时, 地下总生物量(BGB)可以解释42%的ER变化, BGB与ER呈正相关(图9); 缓释尿素处理时分别可以解释28%和35%的ER与GEP变化, 并且BGB与ER和GEP均表现为正相关关系(图9)。 图6https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_7.png<b>图6</b> 不同浓度缓释尿素处理时生态系统净CO<sub>2</sub>交换、生态系统呼吸和生态系统总光合的月动态(平均值±标准误, <i>n</i>=4)<br/>NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。不同小写字母表示处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 6</b> Seasonal patterns of net ecosystem CO<sub>2</sub> exchange, ecosystem respiration and gross ecosystem photosynthesis at different nitrogen addition rates with slow-release urea in each month (means±SE, <i>n</i>=4) <br/>NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>. Different lowercase letters among columns indicate significant differences among treatments (<i>P</i><0.05). Figure 6https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_7.png<b>图6</b> 不同浓度缓释尿素处理时生态系统净CO<sub>2</sub>交换、生态系统呼吸和生态系统总光合的月动态(平均值±标准误, <i>n</i>=4)<br/>NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。不同小写字母表示处理间差异显著(<i>P</i><0.05)。<br/><b>Figure 6</b> Seasonal patterns of net ecosystem CO<sub>2</sub> exchange, ecosystem respiration and gross ecosystem photosynthesis at different nitrogen addition rates with slow-release urea in each month (means±SE, <i>n</i>=4) <br/>NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>. Different lowercase letters among columns indicate significant differences among treatments (<i>P</i><0.05).
图6 不同浓度缓释尿素处理时生态系统净CO2交换、生态系统呼吸和生态系统总光合的月动态(平均值±标准误, n=4) NEE、ER和GEP同表1。不同小写字母表示处理间差异显著(P<0.05)。 Figure 6 Seasonal patterns of net ecosystem CO2 exchange, ecosystem respiration and gross ecosystem photosynthesis at different nitrogen addition rates with slow-release urea in each month (means±SE, n=4) NEE, ER and GEP see Table 1. Different lowercase letters among columns indicate significant differences among treatments (P<0.05).
图7https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_8.png<b>图7</b> 生态系统净CO<sub>2</sub>交换、生态系统呼吸及生态系统总光合与表层(0-10 cm)土壤温度的相关性<br/>(A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。<br/><b>Figure 7</b> Relationships of net ecosystem CO<sub>2 </sub>exchange, ecosystem respiration or gross ecosystem photosynthesis with 0-10 cm soil temperature<br/>(A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>. Figure 7https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_8.png<b>图7</b> 生态系统净CO<sub>2</sub>交换、生态系统呼吸及生态系统总光合与表层(0-10 cm)土壤温度的相关性<br/>(A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。<br/><b>Figure 7</b> Relationships of net ecosystem CO<sub>2 </sub>exchange, ecosystem respiration or gross ecosystem photosynthesis with 0-10 cm soil temperature<br/>(A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>.
图7 生态系统净CO2交换、生态系统呼吸及生态系统总光合与表层(0-10 cm)土壤温度的相关性 (A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同表1。 Figure 7 Relationships of net ecosystem CO2 exchange, ecosystem respiration or gross ecosystem photosynthesis with 0-10 cm soil temperature (A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see Table 1.
图8https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_9.png<b>图8</b> 生态系统净CO<sub>2</sub>交换、生态系统呼吸及生态系统总光合与表层(0-10 cm)土壤含水量的相关性<br/>(A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。<br/><b>Figure 8</b> Relationships of net ecosystem CO<sub>2</sub> exchange, ecosystem respiration or gross ecosystem photosynthesis with 0-10 cm soil moisture<br/>(A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>. Figure 8https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_9.png<b>图8</b> 生态系统净CO<sub>2</sub>交换、生态系统呼吸及生态系统总光合与表层(0-10 cm)土壤含水量的相关性<br/>(A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。<br/><b>Figure 8</b> Relationships of net ecosystem CO<sub>2</sub> exchange, ecosystem respiration or gross ecosystem photosynthesis with 0-10 cm soil moisture<br/>(A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>.
图8 生态系统净CO2交换、生态系统呼吸及生态系统总光合与表层(0-10 cm)土壤含水量的相关性 (A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同表1。 Figure 8 Relationships of net ecosystem CO2 exchange, ecosystem respiration or gross ecosystem photosynthesis with 0-10 cm soil moisture (A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see Table 1.
图9https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_10.png<b>图9</b> 生态系统净CO<sub>2</sub>交换、生态系统呼吸及生态系统总光合与地下总生物量的相关性<br/>(A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。<br/><b>Figure 9</b> Relationships of net ecosystem CO<sub>2</sub> exchange, ecosystem respiration or gross ecosystem photosynthesis with belowground biomass<br/>(A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>. Figure 9https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-27/img_10.png<b>图9</b> 生态系统净CO<sub>2</sub>交换、生态系统呼吸及生态系统总光合与地下总生物量的相关性<br/>(A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同<xref ref-type="table" rid="T1-1674-3466-53-1-27">表1</xref>。<br/><b>Figure 9</b> Relationships of net ecosystem CO<sub>2</sub> exchange, ecosystem respiration or gross ecosystem photosynthesis with belowground biomass<br/>(A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see <xref ref-type="table" rid="T1-1674-3466-53-1-27">Table 1</xref>.
图9 生态系统净CO2交换、生态系统呼吸及生态系统总光合与地下总生物量的相关性 (A)-(C) 添加尿素; (D)-(F) 添加缓释尿素。NEE、ER和GEP同表1。 Figure 9 Relationships of net ecosystem CO2 exchange, ecosystem respiration or gross ecosystem photosynthesis with belowground biomass (A)-(C) Addition with urea; (D)-(F) Addition with slow-release urea. NEE, ER and GEP see Table 1.
表4 Table 4 表4 表4 不同氮素(尿素和缓释尿素)添加水平下生态系统呼吸(ER)和生态系统总光合(GEP)对土壤含水量的敏感性(平均值±标准误, n=4) Table 4 Sensitivity of ecosystem respiration (ER) and gross ecosystem photosynthesis (GEP) to soil moisture under different nitrogen addition rates (means±SE, n=4)
N addition rates (g N·m-2·a-1)
ER
GEP
Urea
0
0.72a±0.09
1.86a±0.64
5
0.95a±0.16
3.23ab±0.62
10
0.59a±0.15
3.72ab±0.71
20
0.76a±0.19
3.67ab±1.35
50
0.74a±0.09
4.54b±0.34
Slow-release urea
0
0.72a±0.09
1.86a±0.64
5
0.89a±0.14
4.21b±0.16
10
1.43b±0.27
5.86c±0.61
20
0.69a±0.12
3.46b±0.62
50
0.92a±0.12
4.99bc±0.36
Different lowercase letters in a column indicate significant differences among different N addition rates at P<0.05. 同列不同小写字母表示各处理间差异显著(P<0.05)。
表4 不同氮素(尿素和缓释尿素)添加水平下生态系统呼吸(ER)和生态系统总光合(GEP)对土壤含水量的敏感性(平均值±标准误, n=4) Table 4 Sensitivity of ecosystem respiration (ER) and gross ecosystem photosynthesis (GEP) to soil moisture under different nitrogen addition rates (means±SE, n=4)
3 讨论本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响。前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009)。当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015)。模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993)。本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011)。降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低。生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加。缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4)。因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强。这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致。本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关。另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009)。由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低。 本研究结果也在一定程度上表明, 缓释尿素氮利用效率高于尿素。缓释尿素通过改变化学成分或在尿素表面包涂半透水性或不透水性物质使其有效成分缓慢释放(陈德明等, 2002), 所以缓释尿素比尿素有效期长。本研究采用的缓释尿素全部释放所用时间大约为3-4个月。尿素一般由于降雨的淋溶、气态氮的挥发、硝态氮的反硝化损失, 导致尿素氮利用率低(吴平霄等, 2000), 而缓释尿素氮肥效果显著大于尿素。另外, 当氮肥(尿素和缓释尿素)添加量较大时(如本研究中的50.0 g N·m-2·a-1), 由于土壤pH值显著降低(P<0.05)导致土壤酸化(图3), 对植物产生毒害作用, 从而抑制生态系统CO2交换(Yan et al., 2010)。
AiresLMI, PioCA, PereiraJS (2008). Carbon dioxide exchange above a Mediterranean C3/C4 grassland during two climatologically contrasting years.Global Change Biol 14, 539-555. [本文引用: 1]
[9]
ArensSJT, SullivanPF, WelkerJM (2008). Nonlinear responses to nitrogen and strong interactions with nitrogen and phosphorus additions drastically alter the structure and function of a high arctic ecosystem. J Geophys Res 113, G03S09. [本文引用: 4]
[10]
BaiYF, WuJG, ClarkCM, NaeemS, PanQM, HuangJH, ZhangLX, HanXG (2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and eco- system functioning: evidence from inner Mongolia grass- lands.Global Change Biol 16, 358-372. [本文引用: 1]
[11]
BaiYF, WuJG, XingQ, PanQM, HuangJH, YangDL, HanXG (2008). Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau.Ecology 89, 2140-2153. [本文引用: 1]
[12]
BubierJL, MooreTR, BledzkiLA (2007). Effects of nutrient addition on vegetation and carbon cycling in an ombrotro- phic bog.Global Change Biol 13, 1168-1186. [本文引用: 1]
[13]
ChenSP, LinGH, HuangJH, JeneretteGD (2009). Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe.Global Change Biol 15, 2450-2461. [本文引用: 1]
[14]
ElserJJ, BrackenMES, ClelandEE, GrunerDS, HarpoleWS, HillebrandH, NgaiJT, SeabloomEW, ShurinJB, SmithJE (2007). Global analysis of nitrogen and phospho- rus limitation of primary producers in freshwater, marine and terrestrial ecosystems.Ecol Lett 10, 1135-1142. [本文引用: 1]
[15]
GruberN, GallowayJN (2008). An earth-system perspec- tive of the global nitrogen cycle.Nature 451, 293-296. [本文引用: 1]
[16]
HarpoleWS, PottsDL, SudingKN (2007). Ecosystem responses to water and nitrogen amendment in a Cali- fornia grassland.Global Change Biol 13, 2341-2348. [本文引用: 3]
[17]
HooperDU, JohnsonL (1999). Nitrogen limitation in dryland ecosystems: responses to geographical and temporal va- riation in precipitation.Biogeochemistry 46, 247-293. [本文引用: 1]
[18]
HuxmanTE, SmithMD, FayPA, KnappAK, ShawMR, LoikME, SmithSD, TissueDT, ZakJC, WeltzinJF, PockmanWT, SalaOE, HaddadBM, HarteJ, KochGW, SchwinningS, SmallEE, WilliamsDG (2004). Conver- gence across biomes to a common rain-use efficiency.Nature 429, 651-654. [本文引用: 1]
[19]
HyvönenR, AgrenGI, LinderS, PerssonT, CotrufoMF, EkbladA, FreemanM, GrelleA, JanssensIA, JarvisPG, KellomäkiS, LindrothA, LoustauD, LundmarkT, NorbyRJ, OrenR, PilegaardK, RyanMG, SigurdssonBD, StrömgrenM, van OijenM, WallinG (2007). The likely impact of elevated [CO2], nitrogen deposition, in- creased temperature and management on carbon seq- uestration in temperate and boreal forest ecosystems: a literature review.New Phytol 173, 463-480. [本文引用: 1]
[20]
JasoniRL, SmithSD, ArnoneJA (2005). Net ecosystem CO2 exchange in Mojave desert shrublands during the eighth year of exposure to elevated CO2.Global Change Biol 11, 749-756. [本文引用: 1]
[21]
KwonH, PendallE, EwersBE, ClearyM, NaithaniK (2008). Spring drought regulates summer net ecosystem CO2 exchange in a sagebrush-steppe ecosystem.Agric Forest Meteor 148, 381-391. [本文引用: 1]
[22]
LeBauerDS, TresederKK (2008). Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed.Ecology 89, 371-379. [本文引用: 1]
[23]
LüFM, LüXT, LiuW, HanX, ZhangGM, KongDL, HanXG (2011). Carbon and nitrogen storage in plant and soil as related to nitrogen and water amendment in a temperate steppe of northern China.Biol Fertility Soils 47, 187-196. [本文引用: 1]
[24]
NiuSL, WuMY, HanYI, XiaJY, LiLH, WanSQ (2008). Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe.New Phytol 177, 209-219. [本文引用: 3]
[25]
NiuSL, WuMY, HanYI, XiaJY, ZhangZHE, YangHJ, WanSQ (2010). Nitrogen effects on net ecosystem carbon exchange in a temperate steppe.Global Change Biol 16, 144-155. [本文引用: 2]
[26]
NiuSL, YangHJ, ZhangZ, WuMY, LuQ, LiLH, HanXG, WanSQ (2009). Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe.Ecosystems 12, 915-926. [本文引用: 2]
[27]
PatrickL, CableJ, PottsD, lgnaceD, Barron-GaffordG, GriffithA, AlpertH, Van GestelN, RobertsonT, HuxmanTE, ZakJ, LoikME, TissueD (2007). Effects of an increase in summer precipitation on leaf, soil, and ecosystem fluxes of CO2 and H2O in a sotol grassland in Big Bend National Park, Texas.Oecologia 151, 704-718. [本文引用: 1]
[28]
PepperDA, Del GrossoSJ, McMurtrieRE, PartonWJ (2005). Simulated carbon sink response of shortgrass steppe, tallgrass prairie and forest ecosystems to rising [CO2], temperature and nitrogen input. Global Biogeochem Cycl 19, GB1004. [本文引用: 1]
[29]
SaarnioS, JärviöS, SaarinenT, VasanderH, SilvolaJ (2003). Minor changes in vegetation and carbon gas balance in a boreal mire under a raised CO2 or NH4NO3 supply.Ecosystems 6, 46-60. [本文引用: 2]
[30]
SeagleSW, McNaughtonSJ (1993). Simulated effects of precipitation and nitrogen on serengeti grassland produc- tivity.Biogeochemistry 22, 157-178. [本文引用: 1]
[31]
ShaverGR, JohnsonLC, CadesDH, MurrayG, LaundreJA, RastetterEB, NadelhofferKJ, GiblinAE (1998). Biomass and CO2 flux in wet sedge tundras: responses to nutrients, temperature, and light.Ecol Monogr 68, 75-97. [本文引用: 1]
[32]
TianDS, NiuSL, PanQM, RenTT, ChenSP, BaiYF, HanXG (2016). Nonlinear responses of ecosystem carbon fluxes and water-use efficiency to nitrogen addition in Inner Mongolia grassland.Funct Ecol 30, 490-499. [本文引用: 3]
VitousekPM, AberJD, HowarthRW, LikensGE, MatsonPA, SchindlerDW, SchlesingerWH, TilmanD (1997). Human alteration of the global nitrogen cycle: sources and consequences.Ecol Appl 7, 737-750. [本文引用: 1]
[35]
VoseJM, ElliottKJ, JohnsonDW, TingeyDT, JohnsonMG (1997). Soil respiration response to three years of elevated CO2 and N fertilization in ponderosa pine ( Pinus ponderosa Dong. ex Laws.). Plant Soil 190, 19-28. [本文引用: 1]
[36]
WangLX, D'odoricoP, O'halloranLR, CaylorK, MackoS (2010). Combined effects of soil moisture and nitrogen availability variations on grass productivity in African sa- vannas.Plant Soil 328, 95-108. [本文引用: 1]
[37]
XiaJY, NiuSL, WanSQ (2009). Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a tem- perate steppe.Global Change Biol 15, 1544-1556. [本文引用: 2]
[38]
XiaJY, WanSQ (2008). Global response patterns of terr- estrial plant species to nitrogen addition.New Phytol 179, 428-439. [本文引用: 2]
[39]
YanLM, ChenSP, HuangJH, LinGH (2010). Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate step- pe.Global Change Biol 16, 2345-2357. [本文引用: 1]
[40]
YanLM, ChenSP, HuangJH, LinGH (2011). Increasing water and nitrogen availability enhanced net ecosystem CO2 assimilation of a temperate semiarid steppe.Plant Soil 349, 227-240. [本文引用: 3]
[41]
ZhangXL, TanYL, LiA, RenTT, ChenSP, WangLX, HuangJH (2015). Water and nitrogen availability co- control ecosystem CO2 exchange in a semiarid temperate steppe.Sci Rep 5, 15549. [本文引用: 1]
缓释和控释尿素的研究与开发综述 1 2002
... 本研究结果也在一定程度上表明, 缓释尿素氮利用效率高于尿素.缓释尿素通过改变化学成分或在尿素表面包涂半透水性或不透水性物质使其有效成分缓慢释放(陈德明等, 2002), 所以缓释尿素比尿素有效期长.本研究采用的缓释尿素全部释放所用时间大约为3-4个月.尿素一般由于降雨的淋溶、气态氮的挥发、硝态氮的反硝化损失, 导致尿素氮利用率低(吴平霄等, 2000), 而缓释尿素氮肥效果显著大于尿素.另外, 当氮肥(尿素和缓释尿素)添加量较大时(如本研究中的50.0 g N·m-2·a-1), 由于土壤pH值显著降低(P<0.05)导致土壤酸化(图3), 对植物产生毒害作用, 从而抑制生态系统CO2交换(Yan et al., 2010). ...
1 2000
... 草地生态系统以40%的陆地面积储存了陆地生态系统中近1/3的有机碳, 维持着30%的净初级生产力, 是陆地生态系统的重要组成部分(陈佐忠和汪诗平, 2000).截至目前, 国内外在氮素与碳循环方面的研究大多集中在植物生长与净初级生产力(NPP) (El- ser et al., 2007; LeBauer and Treseder, 2008; Xia et al., 2009), 这些研究都表明氮添加通过刺激植物生长和积累生物量来增加NPP (Xia and Wan, 2008), 进而显著增加陆地生态系统净初级生产力.然而, 生态系统碳平衡是由碳吸收与碳释放2个过程共同决定, 即生态系统通过光合作用(GEP)固定大气中的CO2, 并通过呼吸作用(ER)使得部分C返回到大气中(Niu et al., 2010). ...
干旱对亚热带人工针叶林碳交换的影响 1 2008
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
改性尿素的肥效及淋溶特性研究初探 1 2000
... 本研究结果也在一定程度上表明, 缓释尿素氮利用效率高于尿素.缓释尿素通过改变化学成分或在尿素表面包涂半透水性或不透水性物质使其有效成分缓慢释放(陈德明等, 2002), 所以缓释尿素比尿素有效期长.本研究采用的缓释尿素全部释放所用时间大约为3-4个月.尿素一般由于降雨的淋溶、气态氮的挥发、硝态氮的反硝化损失, 导致尿素氮利用率低(吴平霄等, 2000), 而缓释尿素氮肥效果显著大于尿素.另外, 当氮肥(尿素和缓释尿素)添加量较大时(如本研究中的50.0 g N·m-2·a-1), 由于土壤pH值显著降低(P<0.05)导致土壤酸化(图3), 对植物产生毒害作用, 从而抑制生态系统CO2交换(Yan et al., 2010). ...
氮添加对内蒙古温带典型草原生态系统碳交换的影响 1 2016
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
氮输入对沼泽湿地碳平衡的影响 1 2006
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
1 1998
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
1 2008
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
4 2008
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ... ... ).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ... ... ; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ... ... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 2010
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 2008
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
1 2007
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
1 2009
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 2007
... 草地生态系统以40%的陆地面积储存了陆地生态系统中近1/3的有机碳, 维持着30%的净初级生产力, 是陆地生态系统的重要组成部分(陈佐忠和汪诗平, 2000).截至目前, 国内外在氮素与碳循环方面的研究大多集中在植物生长与净初级生产力(NPP) (El- ser et al., 2007; LeBauer and Treseder, 2008; Xia et al., 2009), 这些研究都表明氮添加通过刺激植物生长和积累生物量来增加NPP (Xia and Wan, 2008), 进而显著增加陆地生态系统净初级生产力.然而, 生态系统碳平衡是由碳吸收与碳释放2个过程共同决定, 即生态系统通过光合作用(GEP)固定大气中的CO2, 并通过呼吸作用(ER)使得部分C返回到大气中(Niu et al., 2010). ...
1 2008
... 由于人类社会发展以及大量施肥和化石燃料的燃烧, 近年来通过大气沉降返回陆地生态系统的氮(N)元素急剧增加(Vitousek et al., 1997; Tilman et al., 2001; Gruber and Galloway, 2008).氮素是陆地生态系统植物生长的限制因子(Hooper and John- son, 1999; Xia and Wan, 2008), 因此大气氮沉降的增加将对陆地生态系统碳循环带来剧烈的影响. ...
3 2007
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ... ... ; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ... ... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 1999
... 由于人类社会发展以及大量施肥和化石燃料的燃烧, 近年来通过大气沉降返回陆地生态系统的氮(N)元素急剧增加(Vitousek et al., 1997; Tilman et al., 2001; Gruber and Galloway, 2008).氮素是陆地生态系统植物生长的限制因子(Hooper and John- son, 1999; Xia and Wan, 2008), 因此大气氮沉降的增加将对陆地生态系统碳循环带来剧烈的影响. ...
1 2004
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 2007
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 2008
... 草地生态系统以40%的陆地面积储存了陆地生态系统中近1/3的有机碳, 维持着30%的净初级生产力, 是陆地生态系统的重要组成部分(陈佐忠和汪诗平, 2000).截至目前, 国内外在氮素与碳循环方面的研究大多集中在植物生长与净初级生产力(NPP) (El- ser et al., 2007; LeBauer and Treseder, 2008; Xia et al., 2009), 这些研究都表明氮添加通过刺激植物生长和积累生物量来增加NPP (Xia and Wan, 2008), 进而显著增加陆地生态系统净初级生产力.然而, 生态系统碳平衡是由碳吸收与碳释放2个过程共同决定, 即生态系统通过光合作用(GEP)固定大气中的CO2, 并通过呼吸作用(ER)使得部分C返回到大气中(Niu et al., 2010). ...
1 2011
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
3 2008
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ... ... ; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ... ... ), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
2 2010
... 草地生态系统以40%的陆地面积储存了陆地生态系统中近1/3的有机碳, 维持着30%的净初级生产力, 是陆地生态系统的重要组成部分(陈佐忠和汪诗平, 2000).截至目前, 国内外在氮素与碳循环方面的研究大多集中在植物生长与净初级生产力(NPP) (El- ser et al., 2007; LeBauer and Treseder, 2008; Xia et al., 2009), 这些研究都表明氮添加通过刺激植物生长和积累生物量来增加NPP (Xia and Wan, 2008), 进而显著增加陆地生态系统净初级生产力.然而, 生态系统碳平衡是由碳吸收与碳释放2个过程共同决定, 即生态系统通过光合作用(GEP)固定大气中的CO2, 并通过呼吸作用(ER)使得部分C返回到大气中(Niu et al., 2010). ... ... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
2 2009
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ... ... ; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 2007
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 2005
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
2 2003
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ... ... ; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
1 1993
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 1998
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
3 2016
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ... ... ; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ... ... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 2001
... 由于人类社会发展以及大量施肥和化石燃料的燃烧, 近年来通过大气沉降返回陆地生态系统的氮(N)元素急剧增加(Vitousek et al., 1997; Tilman et al., 2001; Gruber and Galloway, 2008).氮素是陆地生态系统植物生长的限制因子(Hooper and John- son, 1999; Xia and Wan, 2008), 因此大气氮沉降的增加将对陆地生态系统碳循环带来剧烈的影响. ...
1 1997
... 由于人类社会发展以及大量施肥和化石燃料的燃烧, 近年来通过大气沉降返回陆地生态系统的氮(N)元素急剧增加(Vitousek et al., 1997; Tilman et al., 2001; Gruber and Galloway, 2008).氮素是陆地生态系统植物生长的限制因子(Hooper and John- son, 1999; Xia and Wan, 2008), 因此大气氮沉降的增加将对陆地生态系统碳循环带来剧烈的影响. ...
1 1997
... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
1 2010
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
2 2009
... 草地生态系统以40%的陆地面积储存了陆地生态系统中近1/3的有机碳, 维持着30%的净初级生产力, 是陆地生态系统的重要组成部分(陈佐忠和汪诗平, 2000).截至目前, 国内外在氮素与碳循环方面的研究大多集中在植物生长与净初级生产力(NPP) (El- ser et al., 2007; LeBauer and Treseder, 2008; Xia et al., 2009), 这些研究都表明氮添加通过刺激植物生长和积累生物量来增加NPP (Xia and Wan, 2008), 进而显著增加陆地生态系统净初级生产力.然而, 生态系统碳平衡是由碳吸收与碳释放2个过程共同决定, 即生态系统通过光合作用(GEP)固定大气中的CO2, 并通过呼吸作用(ER)使得部分C返回到大气中(Niu et al., 2010). ... ... 一般认为, 氮素输入对草地生态系统C吸收和排放2个关键过程都有影响.早期国内外****对氮素添加对生态系统CO2交换的影响进行了一系列研究, 这些实验涉及苔原(Shaver et al., 1998)、北方泥炭地(Saarnio et al., 2003)、沼泽湿地(张丽华等, 2006)、一年生(Harpole et al., 2007)及多年生(Niu et al., 2010)草地和温带草原(Xia et al., 2009)等不同生态系统类型.也有利用模型预测(Pepper et al., 2005)和样带研究(Hyvönen et al., 2007)等方法来评估氮素添加对生态系统CO2交换的影响.然而, 生态系统CO2交换如何响应N素添加至今仍没有得到统一的结论.Shaver等(1998)、Niu等(2009)、Xia等(2009)和Yan等(2011)的研究均表明, 氮素添加对生态系统净CO2交换(NEE)具有显著正效应.另一些研究则发现, 氮素添加对生态系统NEE没有显著影响, 或甚至有负效应(Vose et al., 1997; Saarnio et al., 2003; Bu- bier et al., 2007; Harpole et al., 2007).另外, 当前研究大多侧重于单一剂量氮素添加条件下生态系统CO2的动态, 而不同类型和剂量氮素对生态系统CO2交换的研究却很少(Arens et al., 2008; Tian et al., 2016; 游成铭等, 2016).氮沉降是一个持续的过程, 预计以非线性方式影响生态系统C循环(Arens et al., 2008; Bai et al., 2008; Tian et al., 2016).高氮输入导致饱和响应, 生态系统过程不会随氮素的增加而无限制增加, 因为氮供应充足情况下资源限制可能从氮素转向其它因素, 比如水分、磷素和光照等(Aber et al., 1998; Arens et al., 2008).本研究通过在内蒙古草甸草原开展氮素添加实验, 研究生态系统CO2交换对氮素添加的响应, 以期探明不同剂量氮素(尿素和缓释尿素)添加对草甸草原生态系统CO2交换的影响. ...
2 2008
... 由于人类社会发展以及大量施肥和化石燃料的燃烧, 近年来通过大气沉降返回陆地生态系统的氮(N)元素急剧增加(Vitousek et al., 1997; Tilman et al., 2001; Gruber and Galloway, 2008).氮素是陆地生态系统植物生长的限制因子(Hooper and John- son, 1999; Xia and Wan, 2008), 因此大气氮沉降的增加将对陆地生态系统碳循环带来剧烈的影响. ... ... 草地生态系统以40%的陆地面积储存了陆地生态系统中近1/3的有机碳, 维持着30%的净初级生产力, 是陆地生态系统的重要组成部分(陈佐忠和汪诗平, 2000).截至目前, 国内外在氮素与碳循环方面的研究大多集中在植物生长与净初级生产力(NPP) (El- ser et al., 2007; LeBauer and Treseder, 2008; Xia et al., 2009), 这些研究都表明氮添加通过刺激植物生长和积累生物量来增加NPP (Xia and Wan, 2008), 进而显著增加陆地生态系统净初级生产力.然而, 生态系统碳平衡是由碳吸收与碳释放2个过程共同决定, 即生态系统通过光合作用(GEP)固定大气中的CO2, 并通过呼吸作用(ER)使得部分C返回到大气中(Niu et al., 2010). ...
1 2010
... 本研究结果也在一定程度上表明, 缓释尿素氮利用效率高于尿素.缓释尿素通过改变化学成分或在尿素表面包涂半透水性或不透水性物质使其有效成分缓慢释放(陈德明等, 2002), 所以缓释尿素比尿素有效期长.本研究采用的缓释尿素全部释放所用时间大约为3-4个月.尿素一般由于降雨的淋溶、气态氮的挥发、硝态氮的反硝化损失, 导致尿素氮利用率低(吴平霄等, 2000), 而缓释尿素氮肥效果显著大于尿素.另外, 当氮肥(尿素和缓释尿素)添加量较大时(如本研究中的50.0 g N·m-2·a-1), 由于土壤pH值显著降低(P<0.05)导致土壤酸化(图3), 对植物产生毒害作用, 从而抑制生态系统CO2交换(Yan et al., 2010). ...
3 2011
... 式中, F表示GEP或者ER (μmol·m-2·s-1), θν为表层0-10 cm的土壤含水量, a和b是参数.斜率(a值)可以认为是生态系统CO2交换对土壤含水量的敏感系数(Yan et al., 2011). ... ... 本研究对该地区生态系统CO2交换与土壤温度和土壤含水量之间的关系进行了分析.结果显示, 氮素(尿素和缓释尿素)添加时NEE和GEP随着土壤温度的增加呈线性增加, ER则呈先增加后降低的二项式关系(图7), ER最适温度范围为15-20.6°C, 低于或高于这个范围时ER具有降低趋势.NEE、ER和GEP与土壤含水量之间存在显著对数关系(y=aLn(x)+b) (图8).该方程中斜率(a值)被认为是生态系统CO2交换对土壤含水量的敏感系数(Yan et al., 2011).因此, GEP的水分敏感性显著高于ER的水分敏感性(P<0.05); 氮素添加(无论是尿素还是缓释尿素)显著增加了GEP的水分敏感性(表4).在尿素处理时, 地下总生物量(BGB)可以解释42%的ER变化, BGB与ER呈正相关(图9); 缓释尿素处理时分别可以解释28%和35%的ER与GEP变化, 并且BGB与ER和GEP均表现为正相关关系(图9). ... ... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...
1 2015
... 本研究结果显示, 该草甸草原生态系统CO2交换对氮素添加的响应强烈地受降水事件及降雨量的影响.前人的研究结果也表明, 水分是半干旱草原生态系统CO2交换的一个主要调节因子(Aires et al., 2008; Niu et al., 2008, 2009).当降水量低于年平均值时, 氮素添加抑制生态系统CO2交换; 而当降水量高于年平均值时, 氮素添加促进生态系统CO2交换(Zhang et al., 2015).模型研究也表明, 当降雨量高于平均降雨时, 氮素促进生产力, 说明生产力首先受水分有效性的限制, 其次受氮素有效性的限制(Seagle and McNau- ghton, 1993).本研究表明, 在生长季初期和中期, 由于干旱导致氮素(尿素和缓释尿素)对生态系统CO2交换并没有显著影响, 甚至产生抑制作用, 这与半干旱草原、非洲热带稀树草原和亚热带人工针叶林在降水量较低的条件下, 生态系统CO2交换对氮素添加效果不显著或显著下降的结果相似(顾峰雪等, 2008; Wang et al., 2010; Lü et al., 2011).降雨量低、温度高引起气孔早期关闭以及气孔导度和比叶面积下降(Kwon et al., 2008; Niu et al., 2008), 土壤水分强烈蒸发导致土壤含水量较低, 进而制约生态系统对水分的利用(Niu et al., 2008), 最终导致生态系统CO2交换降低.生长季末期(9月), 由于降雨量增加导致生态系统CO2交换显著增加.缓释尿素的添加显著增加了GEP的水分敏感性, 但对ER的水分敏感性没有显著影响; 尿素添加对GEP水分敏感性有显著增加作用, 并在添加剂量为50 g N·m-2·a-1时达到显著水平(表4).因此, 在降水增加的情况下, GEP对氮素添加的响应要显著高于ER, 从而使该生态系统的NEE显著增强.这与在半干旱草原地区(Chen et al., 2009; Niu et al.,2009; Yan et al., 2011)以及其它地区(Huxman et al., 2004; Harpole et al., 2007; Patrick et al., 2007)的研究结果一致.本研究结果显示, 生长季末期, 当氮素(尿素和缓释尿素)添加达到10 g N·m-2·a-1时, NEE和GEP对氮的响应达到饱和, 此结果与前人在其它草原生态系统所做的氮添加对生态系统CO2交换与生产力影响的实验结果(Arens et al., 2008; Bai et al., 2010; Tian et al., 2016)一致, 但饱和阈值在不同研究中有较大差异, 这也与不同生态系统中水分条件的差异有关.另外, 降水量的季节分配对植物生长和生物量积累至关重要(Niu et al., 2009).由于本年度生长旺季降雨量低, 导致本研究草地生态系统生长季NEE较低, 尽管生长季末(9月)降雨量增加, 在一定程度上促进NEE, 但是其处于生长季末期, 该增加效应无法弥补生长旺季较低的NEE, 使整个生长季NEE总体较低. ...

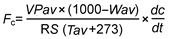

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}