党成强 ,
黄慧敏 ,
董蓉 ,
陈淼 ,
高婷 ,
陶建平 * , 西南大学生命科学学院, 三峡库区生态环境教育部重点实验室, 重庆市三峡库区植物生态与环境资源重点实验室, 重庆 400715
Dang Chengqiang ,
Huang Huimin ,
Dong Rong ,
Chen Miao ,
Gao Ting ,
Tao Jianping * , Chongqing Key Laboratory of Plant Ecology and Resources Research in Three Gorges Reservoir Region, Key Laboratory of Eco-Environments in Three Gorges Reservoir Region, Ministry of Education, School of Life Sciences, Southwest University, Chongqing 400715, China 引用本文 党成强 ,
黄慧敏 ,
董蓉 ,
陈淼 ,
高婷 ,
陶建平 . 重庆中梁山不同石漠化生境中石生南亚毛灰藓 斑块的空间分布格局. , 2017, 52(5): 598-607
贡献者
* 通讯作者。E-mail: taojp@swu.edu.cn 基金资助
国家重点研发计划(No.2016YFC0502304) ;
接受日期:2016-09-25接受日期:2017-03-6网络出版日期:2017-09-1
-->Copyright
2017《植物学报》编辑部
Contributors
* Author for correspondence. E-mail: taojp@swu.edu.cn History
Received :
Accepted :
Online :
摘要: 基于重庆市中梁山喀斯特(Karst)地区3种不同石漠化生境(潜在石漠化、轻度石漠化和中度石漠化)中0.75 hm2 样地的数据, 以石生南亚毛灰藓(Homomallium simlaense )的3种斑块(小斑块、中斑块和大斑块)为研究对象, 运用点格局O-ring的单变量和双变量统计方法对斑块空间格局以及斑块间的关联性进行生态学特性与形成过程分析, 取得以下结果。(1) 南亚毛灰藓总斑块数量关系为轻度石漠化>潜在石漠化>中度石漠化; 在3种石漠化生境中, 各种类型的斑块均呈现出小斑块>中斑块>大斑块的数量变化。(2) 小斑块和中斑块在小尺度上主要为聚集分布, 其它尺度上为随机分布; 大斑块在整个尺度上都呈随机分布。(3) 中斑块与小斑块在3种生境中均在小尺度上有显著的关联性; 大斑块与中斑块仅在潜在石漠化生境中的小尺度上有关联性, 其它斑块间在不同生境与尺度上均无关联性。(4) 环境因子中坡度和石缝数对斑块的分布起很大作用, 但是相对湿度是影响分布的主要因子。(5) 在相对湿度和郁闭度胁迫方面, 小斑块和中斑块随环境胁迫增大而趋于聚集分布。研究结果表明, 苔藓斑块大小及其分布式样在石漠化地区裸露岩石的生态恢复中具有重要指示意义。 关键词 :
石漠化生境 ;
斑块大小 ;
空间格局 ;
胁迫梯度假说 Abstract :
The present study used O-ring Point Pattern Analysis to analyze the spatial distribution patterns of three sizes of moss patches in the Karst area of Zhongliang Mountain, Chongqing, to understand the ecological characteristics and process of the common epilithic moss species Homomallium simlaense . Data were from 0.75 hm2 sample plots in three rocky desertification habitats (potential, slight, and medium rocky desertification). The total number of H. simlaense patches was higher in slight rocky desertification plots, then potential rock desertification plots; medium rock desertification plots had lower patch number. In the three habitats, the changes in patch number were from small patches>medium patches>large patches. Small and medium patches were significantly clustered at small scales but randomly distributed at other scales; however, the large patches randomly distributed at all scales. We found a positive correlation between medium and small patches at small scales in the three habitats and no significant correlation between large and medium patches at most scales, except small scales in potential rocky desertification plots. Relative humidity was the most important environmental factor affecting the moss patch distribution, and the slope and number of rock fracture also played a significant role. Small and medium patches tended to aggregate with increasing moisture and shade stress. Moss patch size and its distribution pattern are good predictors of the ecological restoration in Karst rocky desertification areas. Key words :
rocky desertification habitat ;
patch size ;
spatial distribution pattern ;
stress gradient hypothesis 植物种群的空间分布格局是指个体在种群的二维空间水平结构中的分布位置、方式与分布特征。种群的分布格局研究不仅可以揭示种群的形成过程和维持机制, 而且还可以根据种群形成和维持机制推测不同尺度范围内的种群格局、成因和规律, 同时可以对种群的演替方向做出预判(
王鑫厅等, 2011 )。种群分布格局受到不同尺度上生物因子的影响(
Nathan, 2006 ;
高福元和石福习, 2015 ), 同时还受到非生物因子长期相互作用的影响(
Bai et al., 2014 )。
在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 )。在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 )。在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 )。而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少。
喀斯特脆弱的生态系统一直是中外****的研究热点, 而我国西南地区是全球喀斯特地貌较为集中的区域。由于岩石裸露率高及土层瘠薄等条件限制, 加之人为干扰, 石漠化不断加重的问题亟待解决(
郭柯等, 2011 ;
张军以等, 2015 )。南亚毛灰藓(
Homoma- llium simlaense )作为喀斯特地区常见的一种藓类植物, 主要以平铺的生活型(
Mägdefrau,1982 )生长在完全裸露的岩石上, 极具耐旱性。南亚毛灰藓能够成片地生长在岩石表面, 形成不同的斑块, 并积累养分改善微环境, 为其它被子植物的着生提供适宜的环境条件。同时, 它还作为喀斯特地区演替中的岩溶先锋植物扮演着非常重要的角色(
Glime, 2006 )。
本研究基于二阶统计方法O-ring统计(
张金屯, 1998 ;
Wiegand and Moloney, 2004 )的单变量和双变量分析南亚毛灰藓在不同尺度上的分布格局、不同斑块间的空间关联性及不同石漠化生境中斑块的空间分布规律; 同时应用主成分分析(principal component analysis, PCA)探讨环境因子对不同斑块南亚毛灰藓分布格局的影响。通过尝试从不同大小斑块分布格局及其影响因素的角度来揭示石漠化地区南亚毛灰藓的生态学特性和形成过程, 以期为喀斯特石漠化地区裸露岩石的生态恢复及环境指示提供理论依据。
1 材料与方法1.1 研究区概况研究区域位于重庆市沙坪坝区中梁山中梁镇海石公园(106°24′31″E, 29°41′36″N)。该公园由24个山头组成, 其喀斯特地貌各有特色, 海拔范围为480-690 m。研究区属于中亚热带湿润季风气候, 年平均温度为18 °C, 年均降水量为1 000-1 300 mm, 冬季湿度大且多雾, 夏季高温多雨。该区域在20世纪中期以前, 是郁闭度较高的以柏木(
Cupressus funebris Endl.)、构树(
Broussonetia papyrifera (Linnaeus) L'Heritier ex Ventenat)及棕榈(
Trachycarpus fortunei (Hook.) H. Wendl.)为优势种的亚热带次生林。但由于人为破坏, 目前该区植被主要以草本和1-2 m高的次生灌丛为主。在灌木丛和人工林下的裸露岩石上分布着较多的呈不同斑块状的石生苔藓。
1.2 研究方法根据喀斯特石漠化等级的划分规则(
熊康宁, 2002 ), 选取潜在石漠化、轻度石漠化和中度石漠化3种生境的样地。每个生境设立50 m×50 m的样方, 总样方面积为0.75 hm
2 。各样地概况见
表1 。在进行实地调查后, 记录样地内所有南亚毛灰藓(
Homomallium simlaense )斑块的盖度和形状变化的范围。依据整体数据将群落中的南亚毛灰藓以盖度大小分为3个等级: 小斑块(<0.014 m
2 )、中斑块(0.014-0.028 m
2 )和大斑块(>0.028 m
2 )。由于苔藓斑块的形状不规则, 以不规则形状的几何中心为南亚毛灰藓斑块在样方中的坐标位置, 记录坐标值(x, y) (位置记录精度为±5 cm)。每个样方以西南角为原点开始记录。在记录坐标的同时, 记录其盖度和5个环境因子: 坡度、郁闭度、相对湿度、石缝数和小生境。其中石缝数指在苔藓斑块下所有长>2 cm、宽>1 mm、深>2 mm的石缝数目。小生境分为石面、石壁、石沟、石坑和石洞5种(
张天汉等, 2014 )。
表1 Table 1 表1 表1 3种石漠化生境概况
Table 1 Survey of three types of rocky desertification habitat
Item Plots Potential rocky desertification (A) Slight rocky desertification (B) Medium rocky desertification (C) Elevation (m) 552 564 585 Slope (°) 26.3 22.1 31.5 Shade degree 0.55 0.35 0.10 Aspect South Southeast Northeast Geographical coordinates 29°41′40″N 29°41′53″N 29°41′57″N Vegetation type Broadleaved deciduous forest Thick growth of grass Thick growth of grass
表1
3种石漠化生境概况
Table 1
Survey of three types of rocky desertification habitat
样地中, 乔木层有棕榈、盐肤木(
Rhus chinensis Mill.)和女贞(
Ligustrum lucidum Ait.); 灌木丛有火棘(
Pyracantha fortuneana (Maxim.) Li)、云实(
Caes- alpinia decapetala (Roth) Alston)、小果蔷薇(
Rosa cymosa Tratt.)、金樱子(
Rosa laevigata Michx
. )、山麻杆(
Alchornea davidii Franch.)、铁仔(
Myrsine africana Linn
. )和南天竹(
Nandina domestica Thunb.); 藤本有川莓(
Rubus setchuenensis Bureau et Franch.)、白花银背藤(
Argyreia seguinii (Levl.) Van. ex Levl)、常春油麻藤(
Mucuna sempervirens Hemsl.)、铁线莲(
Clematis florida Thunb.)和菝葜(
Smilax china L.); 草本层有五节芒(
Miscanthus floridulus (Labill.) Warb. ex Schum. et Laut.)、白茅(
Imperata cylindrica (L.) Beauv.)和野菊(
Dendranthema indicum (L.) Des Mo- ul
. ); 苔藓类有大羽藓(
Thuidium cymbifolium (Dozy et Molk.) Dozy et Molk
. )、真藓(
Bryum argenteum Hedw.)、绢藓(
Entodon cladorrhizans (Hedw.) C. Muell.)、大灰藓(
Hypnum plumaeforme Wils
. )、青藓(
Brachythecium pulchellum Broth. et Par.)、小凤尾藓(
Fissidens bryoides Hedw.)、短尖美喙藓(
Eurhynchium angustirete (Broth.) T. Kop.)、南亚毛灰藓(
Homomallium simlaense (Mitt.) Broth.)和鳞叶藓(
Taxiphyllum taxirameum (Mitt.) Fleisch.)。
1.3 数据分析本研究中单个对象的空间分布格局采用单变量O-ring分析方法来完成。若O(r)值高于置信区间上限, 则该对象在该尺度上是聚集分布; 若O(r)值在置信区间之内则为随机分布; 若O(r)值低于置信区间的下限则为均匀分布。对于不同斑块间的关联性, 采用双变量O-ring统计方法中的点格局分析。因为喀斯特的生境异质性较大, 所以采用异质性Poisson过程的零假设分析法, 即大斑块和中斑块位置不变, 小斑块的位置随机变化。在对南亚毛灰藓3个斑块间的关联性进行分析时, 若O(r)值高于置信区间的上限则呈显著正关联; 若低于置信区间的下限则呈显著负关联; 若在置信区间内则表明2个斑块之间相互独立或无显著关联性。依据各斑块的数量特征, 分别在3个石漠化生境中随机选取大中小南亚毛灰藓斑块各20个, 使用主成分分析法(PCA)分析斑块与环境因子的关系。
在分析南亚毛灰藓空间格局和空间关联性过程中, 为精确分析设定栅格大小为1 m×1 m, 圆环宽度为3 m, 设置0-50 m的研究尺度, 并经过99次Monte Carlo模拟得到95%的置信区间。数据分析采用Programita (
Wiegand and Moloney, 2004 )软件完成。使用Excel 2013软件处理数据。使用Origin 9.0软件作图。主成分分析使用CANOCO软件完成。
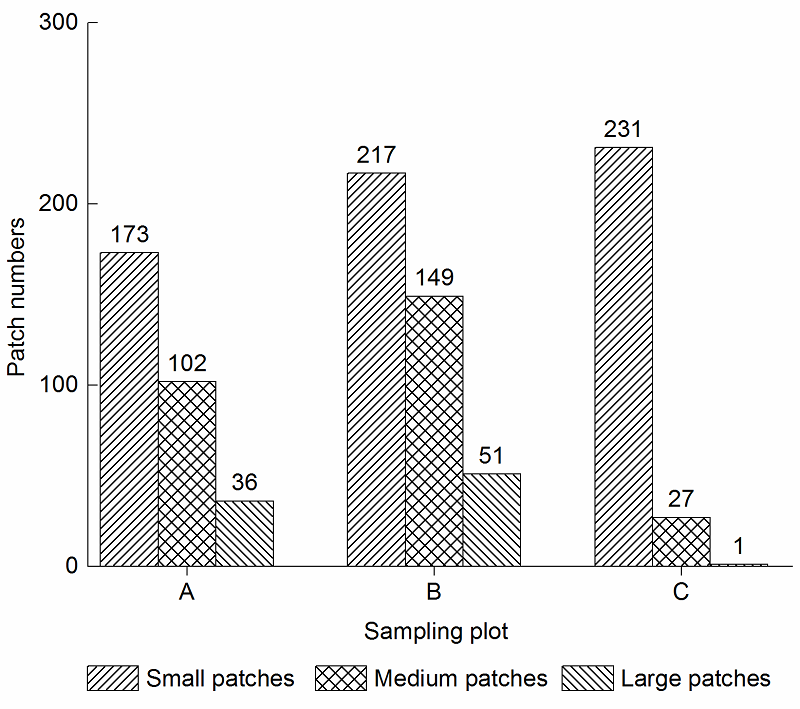
2 结果与讨论2.1 南亚毛灰藓斑块数量特征在3种石漠化生境中, 南亚毛灰藓斑块的总数量关系为轻度石漠化(B)>潜在石漠化(A)>中度石漠化(C)。各种类型的斑块数量均呈现出小斑块>中斑块>大斑块的规律(
图1 )。斑块密度大小分别为, 小斑块: C (924块·hm
-2 )>B (868块·hm
-2 )>A (692块·hm
-2 ); 中斑块: B (596块·hm
2 )>A (408块·hm
-2 )>C (108块·hm
-2 ); 大斑块: B (204块·hm
-2 )>A (144块·hm
-2 )>C (4块·hm
-2 ) (
图2 )。
图1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_1.png <b>图1</b> 3种石漠化生境中南亚毛灰藓斑块的数量关系<br/>A: 潜在石漠化; B: 轻度石漠化; C: 中度石漠化<br/><b>Figure 1</b> The quantitative relation of <i>Homomallium simlaense </i>patches in 3 types of rocky desertification habitat <br/>A: Potential rocky desertification; B: Slight rocky desertification; C: Medium rocky desertification Figure 1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_1.png <b>图1</b> 3种石漠化生境中南亚毛灰藓斑块的数量关系<br/>A: 潜在石漠化; B: 轻度石漠化; C: 中度石漠化<br/><b>Figure 1</b> The quantitative relation of <i>Homomallium simlaense </i>patches in 3 types of rocky desertification habitat <br/>A: Potential rocky desertification; B: Slight rocky desertification; C: Medium rocky desertification 在新窗口打开 下载 下载原图ZIP 生成PPT 图1
3种石漠化生境中南亚毛灰藓斑块的数量关系
A: 潜在石漠化; B: 轻度石漠化; C: 中度石漠化
Figure 1
The quantitative relation of
Homomallium simlaense patches in 3 types of rocky desertification habitat
A: Potential rocky desertification; B: Slight rocky desertification; C: Medium rocky desertification
图2 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_2.png <b>图2</b> 南亚毛灰藓斑块在3种石漠化生境中的空间分布散点图 <br/>(A) 潜在石漠化; (B) 轻度石漠化; (C) 中度石漠化<br/>+ Small patches ○ Medium patches ▲ Large patches<br/><b>Figure 2</b> Scatterplot of spatial distributions of <i>Homomallium simlaense </i>patches in 3 types of rocky desertification habitat<br/>(A) Potential rocky desertification; (B) Slight rocky desertification; (C) Medium rocky desertification Figure 2 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_2.png <b>图2</b> 南亚毛灰藓斑块在3种石漠化生境中的空间分布散点图 <br/>(A) 潜在石漠化; (B) 轻度石漠化; (C) 中度石漠化<br/>+ Small patches ○ Medium patches ▲ Large patches<br/><b>Figure 2</b> Scatterplot of spatial distributions of <i>Homomallium simlaense </i>patches in 3 types of rocky desertification habitat<br/>(A) Potential rocky desertification; (B) Slight rocky desertification; (C) Medium rocky desertification 在新窗口打开 下载 下载原图ZIP 生成PPT 图2
南亚毛灰藓斑块在3种石漠化生境中的空间分布散点图
(A) 潜在石漠化; (B) 轻度石漠化; (C) 中度石漠化
+ Small patches ○ Medium patches ▲ Large patches
Figure 2
Scatterplot of spatial distributions of
Homomallium simlaense patches in 3 types of rocky desertification habitat
(A) Potential rocky desertification; (B) Slight rocky desertification; (C) Medium rocky desertification
2.2 3种石漠化生境中斑块的空间分布我们运用单变量O-ring法统计各样地中南亚毛灰藓不同斑块的空间分布。结果(
图3 )显示, A样地小斑块在0-3 m的尺度上呈明显的聚集分布; 中斑块在0-2.4 m的尺度上呈聚集分布, 在10-11 m的尺度上呈规则分布; 大斑块在整个尺度上呈随机分布。B样地小斑块在0-2 m的尺度上呈显著的聚集分布; 中斑块在0-2.5 m的尺度上也呈聚集分布; 大斑块在整个尺度上呈随机分布。C样地小斑块在0-4 m的尺度上呈明显的聚集分布; 中斑块为随机分布, 大斑块只有1个, 故未做格局分析。以上结果表明, 3种石漠化生境中, 小斑块在小尺度上多为聚集分布; 大斑块在整个尺度上都是随机分布; 而中斑块只有在A和B样地小尺度上为聚集分布。
图3 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_3.png <b>图3</b> 南亚毛灰藓斑块在3种石漠化生境中的分布格局<br/>(A1)-(A3) 潜在石漠化; (B1)-(B3) 轻度石漠化; (C1), (C2) 中度石漠化<br/><b>Figure 3</b> Spatial distribution pattern of <i>Homomallium simlaense</i> patches in 3 types of rocky desertification habitat<br/>(A1)-(A3) Potential rocky desertification; (B1)-(B3) Slight rocky desertification; (C1), (C2) Medium rocky desertification Figure 3 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_3.png <b>图3</b> 南亚毛灰藓斑块在3种石漠化生境中的分布格局<br/>(A1)-(A3) 潜在石漠化; (B1)-(B3) 轻度石漠化; (C1), (C2) 中度石漠化<br/><b>Figure 3</b> Spatial distribution pattern of <i>Homomallium simlaense</i> patches in 3 types of rocky desertification habitat<br/>(A1)-(A3) Potential rocky desertification; (B1)-(B3) Slight rocky desertification; (C1), (C2) Medium rocky desertification 在新窗口打开 下载 下载原图ZIP 生成PPT 图3
南亚毛灰藓斑块在3种石漠化生境中的分布格局
(A1)-(A3) 潜在石漠化; (B1)-(B3) 轻度石漠化; (C1), (C2) 中度石漠化
Figure 3
Spatial distribution pattern of
Homomallium simlaense patches in 3 types of rocky desertification habitat
(A1)-(A3) Potential rocky desertification; (B1)-(B3) Slight rocky desertification; (C1), (C2) Medium rocky desertification
2.3 3种石漠化生境中不同斑块的空间关联我们运用双变量O-ring统计法分析南亚毛灰藓在3个样地中不同斑块的空间关联性。结果(
图4 )显示, A样地中, 大斑块和小斑块在20.5-21.5 m的尺度上为负关联, 大斑块和中斑块在0-2 m的尺度上具有显著的正关联; 中斑块和小斑块在0-3 m的尺度上有显著的正关联。B样地中, 大斑块和小斑块与大斑块和中斑块都无关联性; 只有中斑块和小斑块在0-3 m的尺度上呈显著关联。在C样地中, 中斑块和小斑块在0-2 m的尺度上显著关联, 在16-17.5 m的尺度上有关联; 在其它尺度上均无关联。以上结果表明, 中斑块和小斑块在3个样地中都有显著的关联性, 说明中斑块与小斑块有相互的关联作用, 而大斑块仅在样地A中与中斑块有显著关联。
图4 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_4.png <b>图4</b> 3种石漠化生境中南亚毛灰藓不同斑块的空间关联<br/>(A1)-(A3) 潜在石漠化; (B1)-(B3) 轻度石漠化; (C1) 中度石漠化<br/><b>Figure 4</b> Spatial associations of different patches of <i>Homomallium simlaense</i> in 3 types of rocky desertification habitat<br/>(A1)-(A3) Potential rocky desertification; (B1)-(B3) Slight rocky desertification; (C1) Medium rocky desertification Figure 4 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_4.png <b>图4</b> 3种石漠化生境中南亚毛灰藓不同斑块的空间关联<br/>(A1)-(A3) 潜在石漠化; (B1)-(B3) 轻度石漠化; (C1) 中度石漠化<br/><b>Figure 4</b> Spatial associations of different patches of <i>Homomallium simlaense</i> in 3 types of rocky desertification habitat<br/>(A1)-(A3) Potential rocky desertification; (B1)-(B3) Slight rocky desertification; (C1) Medium rocky desertification 在新窗口打开 下载 下载原图ZIP 生成PPT 图4
3种石漠化生境中南亚毛灰藓不同斑块的空间关联
(A1)-(A3) 潜在石漠化; (B1)-(B3) 轻度石漠化; (C1) 中度石漠化
Figure 4
Spatial associations of different patches of
Homomallium simlaense in 3 types of rocky desertification habitat
(A1)-(A3) Potential rocky desertification; (B1)-(B3) Slight rocky desertification; (C1) Medium rocky desertification
2.4 3种石漠化生境中不同斑块与环境的关系通过主成分分析前4轴的统计特征(
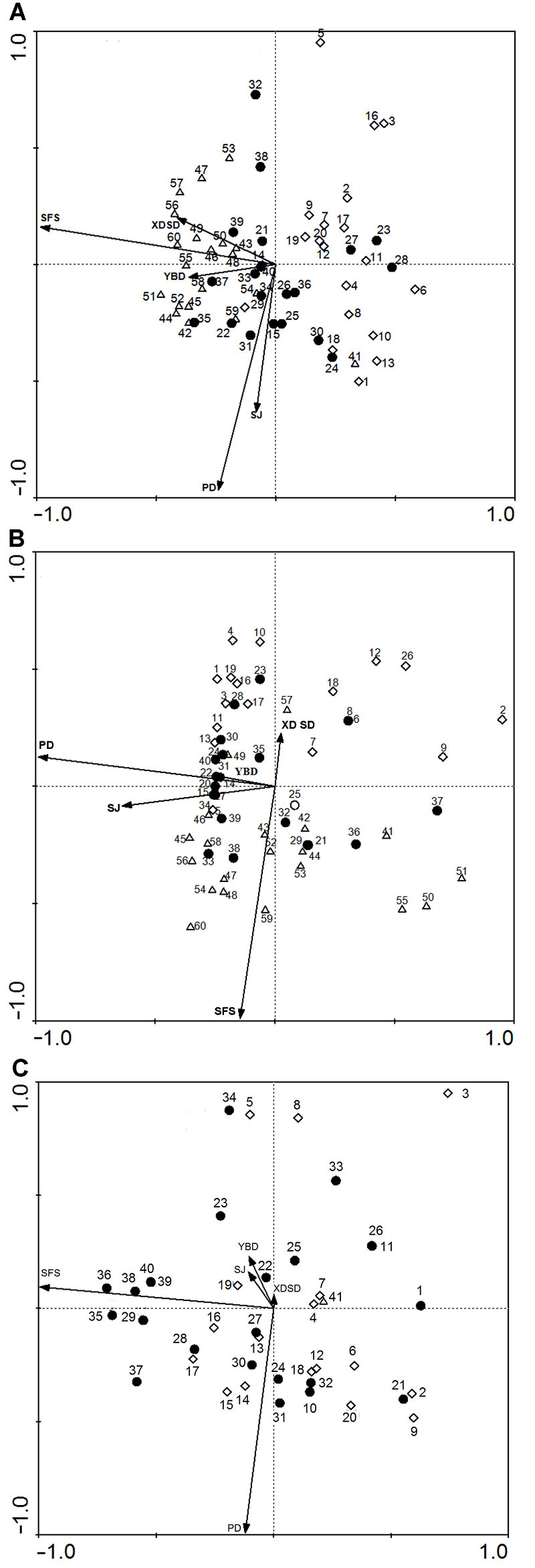
表2 ), 表明石缝数在样地A和C与第1轴及样地B与第2轴呈显著负相关。这表明随着石缝数增多, 斑块面积也会随之变大。坡度在样地A和C与第2轴及样地B与第1轴呈显著负相关。在一定坡度分布的大斑块和中斑块较为集中, 而小斑块的分布对坡度没有很强的依赖性。相对湿度在样地A与第1轴呈负相关, 在样地B和C与第2轴呈正相关。在样地A相对湿度大的地方分布着较多的大斑块和中斑块, 3个样地中, 随着湿度降低小斑块分布渐多。郁闭度在样地A和B与第1轴呈负相关, 在样地C与第2轴呈正相关。在郁闭度大的地方分布较多的大斑块, 且随着郁闭度的降低中斑块和小斑块分布渐多。生境在样地A与第2轴呈负相关, 在样地B和C与第1轴、第2轴呈正相关。大斑块和中斑块多分布在石面上, 而小斑块多分布在石壁、石沟及石坑等小生境中。以上结果(
图5 )表明, 在3种石漠化生境中, 南亚毛灰藓在湿度大、石缝数多、郁闭度高和坡度小的石面生境分布着大斑块; 其受到环境因子的限制较多, 而环境胁迫较小。中斑块与大斑块呈现出相似的趋势, 但是各个环境因子对中斑块的限制程度小于大斑块。小斑块多分布在郁闭度较小、相对湿度小、石缝数少、坡度和生境因子限制较小的环境中, 其生长受到的环境胁迫较大。
表2 Table 2 表2 表2 3种石漠化生境中主成分分析前4轴的统计特征
Table 2 Statistical characteristics of the first 4 axes of the principal component analysis in 3 types of rocky desertification habitat
Plots Axis 1 2 3 4 Potential rocky desertification (A) Eigenvalues 0.515 0.329 0.123 0.028 Cumulative percentage variance of species data 51.5 84.4 96.6 99.4 Slight rocky desertification (B) Eigenvalues 0.527 0.398 0.057 0.016 Cumulative percentage variance of species data 52.7 92.5 98.2 99.8 Medium rocky desertification (C) Eigenvalues 0.461 0.385 0.141 0.011 Cumulative percentage variance of species data 46.1 84.7 98.7 99.8
表2
3种石漠化生境中主成分分析前4轴的统计特征
Table 2
Statistical characteristics of the first 4 axes of the principal component analysis in 3 types of rocky desertification habitat
图5 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_5.png <b>图5</b> 3种石漠化生境中南亚毛灰藓不同斑块与环境因子的关系 <br/>(A) 潜在石漠化; (B) 轻度石漠化; (C) 中度石漠化。1、2、3……为斑块号。SFS: 石缝数; SJ: 生境(石面、石壁、石沟、石坑和石洞); PD: 坡度: YBD: 郁闭度; XDSD: 相对湿度<br/>◇Small patches ●Medium patches △Large patches<br/><b>Figure 5</b> Different patches of <i>Homomallium simlaense</i> rela- tionship with environmental factors in 3 types of rocky desertification habitat<br/>(A) Potential rocky desertification; (B) Slight rocky desertification; (C) Medium rocky desertification. 1, 2, 3……No. of patches. SFS: Rock fracture; SJ: Microhabitat (stone facing, cliff, stony gully, stone pit, and grotto); PD: Gradient; YBD: Crown density; XDSD: Relative humidity Figure 5 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-598/img_5.png <b>图5</b> 3种石漠化生境中南亚毛灰藓不同斑块与环境因子的关系 <br/>(A) 潜在石漠化; (B) 轻度石漠化; (C) 中度石漠化。1、2、3……为斑块号。SFS: 石缝数; SJ: 生境(石面、石壁、石沟、石坑和石洞); PD: 坡度: YBD: 郁闭度; XDSD: 相对湿度<br/>◇Small patches ●Medium patches △Large patches<br/><b>Figure 5</b> Different patches of <i>Homomallium simlaense</i> rela- tionship with environmental factors in 3 types of rocky desertification habitat<br/>(A) Potential rocky desertification; (B) Slight rocky desertification; (C) Medium rocky desertification. 1, 2, 3……No. of patches. SFS: Rock fracture; SJ: Microhabitat (stone facing, cliff, stony gully, stone pit, and grotto); PD: Gradient; YBD: Crown density; XDSD: Relative humidity 在新窗口打开 下载 下载原图ZIP 生成PPT 图5
3种石漠化生境中南亚毛灰藓不同斑块与环境因子的关系
(A) 潜在石漠化; (B) 轻度石漠化; (C) 中度石漠化。1、2、3……为斑块号。SFS: 石缝数; SJ: 生境(石面、石壁、石沟、石坑和石洞); PD: 坡度: YBD: 郁闭度; XDSD: 相对湿度
◇Small patches ●Medium patches △Large patches
Figure 5
Different patches of
Homomallium simlaense rela- tionship with environmental factors in 3 types of rocky desertification habitat
(A) Potential rocky desertification; (B) Slight rocky desertification; (C) Medium rocky desertification. 1, 2, 3……No. of patches. SFS: Rock fracture; SJ: Microhabitat (stone facing, cliff, stony gully, stone pit, and grotto); PD: Gradient; YBD: Crown density; XDSD: Relative humidity
2.5 讨论2.5.1 南亚毛灰藓在样地内的分布
苔藓植物斑块大小主要与其所处不同尺度上的生境有密切关系(
籍烨和张朝晖, 2014 )。在3种石漠化生境中, 潜在石漠化样地的整体郁闭度大于轻度石漠化样地, 但是3种斑块的数量都少于轻度石漠化样地。在实地调查中, 我们发现潜在石漠化样地中草本和灌木可以完全覆盖在整个石面上, 对苔藓植物的生长和繁殖产生一定的抑制作用。从
图2 可以看出, 斑块在3种石漠化生境中的空间分布有明显的空白处, 潜在石漠化生境中多为密度大的草本和灌木; 轻度和中度石漠化生境空白多为郁闭度为0的完全裸露的岩石。因此南亚毛灰藓不同斑块的数量变化和分布受大生境与小生境的影响而发生变化。
2.5.2 不同斑块的空间分布与空间关联
物种的空间分布格局与尺度有着密切联系, 并随着尺度的变化而变化。在小尺度上, 空间格局和空间关联性可能是由于生境、竞争以及种子扩散等因素造成。而在大尺度上, 则可能是生境异质性和种子的扩散限制导致的(
Rayburn et al., 2011 ;
Shen et al., 2013 )。南亚毛灰藓孢子小且轻, 局部的扩散会使多数孢子落在配子体附近。重庆中梁山喀斯特地区地形复杂, 小生境异质性大, 石沟、石槽使孢子在扩散过程中容易被阻挡。扩散限制和生境条件的异质性使得小斑块和中斑块在小尺度上为聚集分布。而大斑块占据着水分条件相对优越且其它条件也较好的生境, 抗旱能力增强、相互依赖性减弱, 从而使竞争力增大, 因而大斑块多为随机分布。
由于群落生境的差异影响了物种的分布, 关联性有助于进一步认识群落的结构、类型及群落在生境中相互作用所形成的有机联系, 体现了物种之间的竞争关系以及对生境分化的反应(
王琳和张金屯, 2004 ;
姜俊和赵秀海, 2011 ;
郭垚鑫等, 2014 )。本研究中, 中斑块与小斑块之间通过改变微环境相互庇护, 抵御恶劣的环境以提高存活机会, 这种小尺度上正向的相互关联性可能是其应对喀斯特严峻的生境异质性以及应对极端环境的生态对策之一。而大斑块对中、小斑块无关联性, 其占据适宜的环境后, 在生长过程中分泌酸性物质, 促进岩面溶解而自身残骸堆积在岩面上, 有助于加快岩石表面碳循环速率、富集营养元素、涵养水分及土壤的沉积, 为其它高等植物着生在岩石上创造了必要条件(
《广西西南喀斯特生物多样性》编委会, 2011 )。因此在喀斯特地区, 南亚毛灰藓以小斑块和中斑块的状态广泛分布, 存活率较高, 而大斑块可为演替初期做铺垫。
2.5.3 不同斑块的南亚毛灰藓与环境的关系
随着环境的变化苔藓植物自身的生长和分布呈现出不同的响应。在不同的生境下, 其分布、群落大小以及盖度都会有明显的差异, 即使在同种生境下其生长也有所不同(
张元明等, 2003 ;
吉雪花等, 2013 ;
皮春燕和刘艳, 2014 )。不同的环境因子对植物群落的格局也有一定的影响(
刘艳等, 2008 ;
宋同清等, 2010 ;
李军峰等, 2015 )。本研究中, 石缝数和坡度都与南亚毛灰藓的3种斑块有很强的关联性, 但是苔藓植物作为变水植物(poikilohydric plants)对水分有很强的依赖性, 石生南亚毛灰藓平铺的生活型通过内吸水和外吸水来增加对水分的涵养。因此相对湿度是石生苔藓斑块大小变化和格局分布的重要因素之一。
胁迫梯度假说(stress gradient hypothesis, SGH)认为, 随胁迫强度的增加, 植物间正相互作用增强(
Callaway and Walker, 1997 ;
Callaway, 2007 ;
Brooker et al., 2008 ;
张炜平等, 2013 )。综合本研究基于不同尺度下不同斑块的空间分布与环境的关系, 从相对湿度和郁闭度胁迫来说, 大斑块在环境因子中受到的环境胁迫较小, 小斑块最大。因此小斑块和中斑块随环境胁迫增大而趋于聚集分布。这基本符合胁迫梯度假说, 也为其提供了数据支撑, 但仍需要进行受控实验以确定斑块间相互作用特性及其随环境胁迫变化的生态机制。以上结果表明, 不同斑块的分布也受到一定的环境胁迫影响。
自然条件下, 种群的结构和空间分布往往是种群生物学特性与环境相互作用的结果(
Condit et al., 2000 )。不同斑块南亚毛灰藓的分布规律揭示了在喀斯特地区岩石先锋种苔藓植物在自然演替中的生态学进程, 同时不同斑块与环境变化的关系阐释了南亚毛灰藓在喀斯特地区的适应性。石生南亚毛灰藓不同斑块的分布不仅受孢子扩散和生境异质性的影响, 而且不同的环境因子和一定的环境胁迫也会对南亚毛灰藓的分布起重要作用。对于生态恢复来说, 我国西南部石漠化地区的生态恢复多是以种植经济林和速生林为主(
张军以等, 2015 ), 忽略了裸露岩石的恢复。而根据喀斯特石生苔藓的分布特点, 在郁闭度和湿度条件较好的石面上可培养大斑块的石生苔藓, 为景观或其它植物的生存创造条件。而在郁闭度低、湿度低和石漠化严重的区域, 可在环境条件相对优越的小生境中以小斑块散点模式培养苔藓, 以提高成活率; 同时待苔藓成活后可改变微环境使其相互庇护, 提高抗干旱和高温等环境胁迫的能力。
致谢 重庆师范大学生命科学学院刘艳副教授帮助鉴定苔藓植物; 重庆文理学院林学与生命科学学院王微副教授指导论文撰写; 西南大学生命科学学院钱凤博士对论文提出修改建议。在此一并表示衷心感谢。 The authors have declared that no competing interests exist. 参考文献 文献选项 原文顺序 文献年度倒序 文中引用次数倒序 被引期刊影响因子 [1] 高福元 , 石福习 (2015 ). 基于不同零模型的三江平原沼泽湿地主要物种小尺度点格局分析. 生态学报 35, 2029 -2037 .[2] 《广西西南喀斯特生物多样性》编委会 (2011 ). 广西西南喀斯特生物多样性 . 北京: 中国大百科全书出版社. pp.106 -108 .[3] 郭柯 , 刘长成 , 董鸣 (2011 ). 我国西南喀斯特植物生态适应性与石漠化治理. 植物生态学报 35, 991 -999 .[4] 郭水良 , 曹同 (2000 ). 长白山地区森林生态系统树附生苔藓植物群落分布格局研究. 植物生态学报 24, 442 -450 .[5] 郭垚鑫 , 胡有宁 , 李刚 , 王得祥 , 杨吉健 , 杨改河 (2014 ). 太白山红桦种群不同发育阶段的空间格局与关联性. 林业科学 50, 9 -14 .[6] 郭屹立 , 王斌 , 向悟生 , 丁涛 , 陆树华 , 黄俞淞 , 黄甫昭 , 李冬兴 , 李先琨 (2015 ). 广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析. 生物多样性 23, 183 -191 .[7] 韩文衡 , 向悟生 , 叶铎 , 吕仕洪 , 丁涛 , 李先琨 (2010 ). 广西木论保护区喀斯特常绿落叶阔叶混交林优势种空间格局及其相关性. 应用生态学报 21, 2769 -2776 .[8] 吉雪花 , 张元明 , 陶冶 , 周小兵 , 张静 (2013 ). 藓类结皮斑块面积与环境因子的关系. 中国沙漠 33, 1803 -1809 .[9] 吉雪花 , 张元明 , 周小兵 , 吴林 , 张静 (2014 ). 不同尺度苔藓结皮土壤性状的空间分布特征. 生态学报 34, 4006 -4016 .[10] 籍烨 , 张朝晖 (2014 ). 喀斯特石漠生态系统不同自然演替阶段中苔藓植物多样性特征分析. 植物科学学报 32, 577 -585 .[11] 姜俊 , 赵秀海 (2011 ). 吉林蛟河针阔混交林群落优势种群种间联结性. 林业科学 47, 149 -153 .[12] 李军峰 , 贾少华 , 王智慧 , 张朝晖 (2015 ). 喀斯特石漠化过程中苔藓植物多样性及分布与环境关系. 生态科学 34, 68 -73 .[13] 刘艳 , 曹同 , 王剑 , 曹阳 (2008 ). 杭州市区土生苔藓植物分布与生态因子的关系. 应用生态学报 19, 775 -781 .[14] 皮春燕 , 刘艳 (2014 ). 重庆主城区住宅小区苔藓组成与多样性. 生物多样性 22, 583 -588 .[15] 宋同清 , 彭晚霞 , 曾馥平 , 王克林 , 覃文更 , 谭卫宁 , 刘璐 , 杜虎 , 鹿士杨 (2010 ). 木论喀斯特峰丛洼地森林群落空间格局及环境解释. 植物生态学报 34, 298 -308 .[16] 王琳 , 张金屯 (2004 ). 历山山地草甸优势种的种间关联和相关分析. 西北植物学报 24, 1435 -1440 .[17] 王晓雨 , 于大炮 , 周莉 , 周旺明 , 吴志军 , 郭焱 , 包也 , 孟莹莹 , 代力民 (2015 ). 长白山北坡林线岳桦种群空间分布格局. 生态学报 35, 116 -124 .[18] 王鑫厅 , 侯亚丽 , 刘芳 , 常英 , 王炜 , 梁存柱 , 苗百岭 (2011 ). 羊草+大针茅草原退化群落优势种群空间点格局分析. 植物生态学报 35, 1281 -1289 .[19] 熊康宁 (2002 ). 喀斯特石漠化的遥感——GIS典型研究: 以贵州省为例 . 北京: 地质出版社. pp. 23 -25 .[20] 徐晟翀 , 曹同 , 于晶 , 陈怡 , 宋国元 (2006 ). 上海市树附生苔藓植物分布格局研究. 西北植物学报 26, 1053 -1058 .[21] 许强 , 吕金枝 , 苗艳明 , 毕润成 (2016 ). 翅果油树群落主要物种空间分布格局及其关联性. 植物学报 51, 49 -57 .[22] 杨洪晓 , 张金屯 , 吴波 , 李晓松 , 张友炎 (2006 ). 毛乌素沙地油蒿种群点格局分析. 植物生态学报 30, 563 -570 .[23] 张金屯 (1998 ). 植物种群空间分布的点格局分析. 植物生态学报 22, 344 -349 .[24] 张军以 , 戴明宏 , 王腊春 , 苏维词 , 曹立国 (2015 ). 西南喀斯特石漠化治理植物选择与生态适应性. 地球与环境 43, 269 -278 .[25] 张明娟 , 刘茂松 , 徐驰 , 池婷 , 洪超 (2012 ). 不同密度条件下芨芨草空间格局对环境胁迫的响应. 生态学报 32, 595 -604 .[26] 张天汉 , 代玉 , 王智慧 , 张朝晖 (2014 ). 贵州关岭县喀斯特峰丛石漠区苔藓群落生态特征. 中国岩溶 33, 192 -200 .[27] 张炜平 , 潘莎 , 贾昕 , 储诚进 , 肖洒 , 林玥 , 白燕远 , 王根轩 (2013 ). 植物间正相互作用对种群动态和群落结构的影响: 基于个体模型的研究进展. 植物生态学报 37, 571 -582 .[28] 张元明 , 曹同 , 潘伯荣 (2003 ). 新疆博格达山地面生苔藓植物物种多样性研究. 应用生态学报 14, 887 -891 .[29] 张忠华 , 胡刚 , 倪健 (2015 ). 茂兰喀斯特常绿落叶阔叶混交林树种的空间分布格局及其分形特征. 生态学报 35, 8221 -8230 .[30] Bai C Yan M BI R He YH 2014 ). Spatial pattern analysis of dominant species in Exochorda giraldii community in Xingtang Temple of Taiyue Mountains, Shanxi, China.Chin J Plant Ecol 38, 1283 -1295 .[31] Brooker RW Maestre FT Callaway RM Lortie CL Cavieres LA Kunstler G Liancourt P Tielböerger K Travis JMJ Anthelme F Armas C Coll L Corcket E Delzon S Forey E Kikvidze Z Olofsson J Pugnaire FI Quiroz CL Saccone P Schiffers K Seifan M Touzard B Michalet R 2008 ). Facilitation in plant comm- unities: the past, the present, and the future.J Ecol 96, 18 -34 .[32] Callaway RM 2007 ). Positive Interactions and Interdependence in Plant Communities. Dordrecht: Springer. pp. 443 -444 .[33] Callaway RM Walker LR 1997 ). Competition and facilitation: a synthetic approach to interactions in plant communities.Ecology 78, 1958 -1965 .[34] Condit R Ashton PS Baker P Bunyavejchewin S Gunatilleke S Gunatilleke N Hubbell SP Foster RB Itoh A LaFrankie JV Lee HS Losos E Manokaran N Sukumar R Yamakura T 2000 ). Spatial patterns in the distribution of tropical tree species.Science 288, 1414 -1418 .[35] Glime JM 2006 ). Bryophyte Ecology. .URL [36] Mägdefrau K 1982 ). Life-forms of Bryophytes. In: Smith AJE, ed. Bryophyte Ecology. Dordrecht: Springer . pp. 45 -58 .[37] Nathan R 2006 ). Long-distance dispersal of plants.Science 313, 786 -788 .[38] Rayburn AP Schiffers K Schupp EW 2011 ). Use of precise spatial data for describing spatial patterns and plant interactions in a diverse Great Basin shrub community.Plant Ecol 212, 585 -594 .[39] Shen GC He FL Waagepetersen R Sun IF Hao ZQ Chen ZS Yu MJ 2013 ). Quantifying effects of habitat heterogeneity and other clustering processes on spatial distributions of tree species.Ecology 94, 2436 -2443 .[40] Wiegand T Moloney KA 2004 ). Rings, circles, and null- models for point pattern analysis in ecology.Oikos 104, 209 -229 .
基于不同零模型的三江平原沼泽湿地主要物种小尺度点格局分析
1
2015
... 植物种群的空间分布格局是指个体在种群的二维空间水平结构中的分布位置、方式与分布特征.种群的分布格局研究不仅可以揭示种群的形成过程和维持机制, 而且还可以根据种群形成和维持机制推测不同尺度范围内的种群格局、成因和规律, 同时可以对种群的演替方向做出预判(
王鑫厅等, 2011 ).种群分布格局受到不同尺度上生物因子的影响(
Nathan, 2006 ;
高福元和石福习, 2015 ), 同时还受到非生物因子长期相互作用的影响(
Bai et al., 2014 ). ...
1
2011
... 由于群落生境的差异影响了物种的分布, 关联性有助于进一步认识群落的结构、类型及群落在生境中相互作用所形成的有机联系, 体现了物种之间的竞争关系以及对生境分化的反应(
王琳和张金屯, 2004 ;
姜俊和赵秀海, 2011 ;
郭垚鑫等, 2014 ).本研究中, 中斑块与小斑块之间通过改变微环境相互庇护, 抵御恶劣的环境以提高存活机会, 这种小尺度上正向的相互关联性可能是其应对喀斯特严峻的生境异质性以及应对极端环境的生态对策之一.而大斑块对中、小斑块无关联性, 其占据适宜的环境后, 在生长过程中分泌酸性物质, 促进岩面溶解而自身残骸堆积在岩面上, 有助于加快岩石表面碳循环速率、富集营养元素、涵养水分及土壤的沉积, 为其它高等植物着生在岩石上创造了必要条件(
《广西西南喀斯特生物多样性》编委会, 2011 ).因此在喀斯特地区, 南亚毛灰藓以小斑块和中斑块的状态广泛分布, 存活率较高, 而大斑块可为演替初期做铺垫. ...
我国西南喀斯特植物生态适应性与石漠化治理
1
2011
... 喀斯特脆弱的生态系统一直是中外****的研究热点, 而我国西南地区是全球喀斯特地貌较为集中的区域.由于岩石裸露率高及土层瘠薄等条件限制, 加之人为干扰, 石漠化不断加重的问题亟待解决(
郭柯等, 2011 ;
张军以等, 2015 ).南亚毛灰藓(
Homoma- llium simlaense )作为喀斯特地区常见的一种藓类植物, 主要以平铺的生活型(
Mägdefrau,1982 )生长在完全裸露的岩石上, 极具耐旱性.南亚毛灰藓能够成片地生长在岩石表面, 形成不同的斑块, 并积累养分改善微环境, 为其它被子植物的着生提供适宜的环境条件.同时, 它还作为喀斯特地区演替中的岩溶先锋植物扮演着非常重要的角色(
Glime, 2006 ). ...
长白山地区森林生态系统树附生苔藓植物群落分布格局研究
1
2000
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
太白山红桦种群不同发育阶段的空间格局与关联性
1
2014
... 由于群落生境的差异影响了物种的分布, 关联性有助于进一步认识群落的结构、类型及群落在生境中相互作用所形成的有机联系, 体现了物种之间的竞争关系以及对生境分化的反应(
王琳和张金屯, 2004 ;
姜俊和赵秀海, 2011 ;
郭垚鑫等, 2014 ).本研究中, 中斑块与小斑块之间通过改变微环境相互庇护, 抵御恶劣的环境以提高存活机会, 这种小尺度上正向的相互关联性可能是其应对喀斯特严峻的生境异质性以及应对极端环境的生态对策之一.而大斑块对中、小斑块无关联性, 其占据适宜的环境后, 在生长过程中分泌酸性物质, 促进岩面溶解而自身残骸堆积在岩面上, 有助于加快岩石表面碳循环速率、富集营养元素、涵养水分及土壤的沉积, 为其它高等植物着生在岩石上创造了必要条件(
《广西西南喀斯特生物多样性》编委会, 2011 ).因此在喀斯特地区, 南亚毛灰藓以小斑块和中斑块的状态广泛分布, 存活率较高, 而大斑块可为演替初期做铺垫. ...
广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析
1
2015
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
广西木论保护区喀斯特常绿落叶阔叶混交林优势种空间格局及其相关性
1
2010
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
藓类结皮斑块面积与环境因子的关系
1
2013
... 随着环境的变化苔藓植物自身的生长和分布呈现出不同的响应.在不同的生境下, 其分布、群落大小以及盖度都会有明显的差异, 即使在同种生境下其生长也有所不同(
张元明等, 2003 ;
吉雪花等, 2013 ;
皮春燕和刘艳, 2014 ).不同的环境因子对植物群落的格局也有一定的影响(
刘艳等, 2008 ;
宋同清等, 2010 ;
李军峰等, 2015 ).本研究中, 石缝数和坡度都与南亚毛灰藓的3种斑块有很强的关联性, 但是苔藓植物作为变水植物(poikilohydric plants)对水分有很强的依赖性, 石生南亚毛灰藓平铺的生活型通过内吸水和外吸水来增加对水分的涵养.因此相对湿度是石生苔藓斑块大小变化和格局分布的重要因素之一. ...
不同尺度苔藓结皮土壤性状的空间分布特征
1
2014
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
喀斯特石漠生态系统不同自然演替阶段中苔藓植物多样性特征分析
1
2014
... 苔藓植物斑块大小主要与其所处不同尺度上的生境有密切关系(
籍烨和张朝晖, 2014 ).在3种石漠化生境中, 潜在石漠化样地的整体郁闭度大于轻度石漠化样地, 但是3种斑块的数量都少于轻度石漠化样地.在实地调查中, 我们发现潜在石漠化样地中草本和灌木可以完全覆盖在整个石面上, 对苔藓植物的生长和繁殖产生一定的抑制作用.从
图2 可以看出, 斑块在3种石漠化生境中的空间分布有明显的空白处, 潜在石漠化生境中多为密度大的草本和灌木; 轻度和中度石漠化生境空白多为郁闭度为0的完全裸露的岩石.因此南亚毛灰藓不同斑块的数量变化和分布受大生境与小生境的影响而发生变化. ...
吉林蛟河针阔混交林群落优势种群种间联结性
1
2011
... 由于群落生境的差异影响了物种的分布, 关联性有助于进一步认识群落的结构、类型及群落在生境中相互作用所形成的有机联系, 体现了物种之间的竞争关系以及对生境分化的反应(
王琳和张金屯, 2004 ;
姜俊和赵秀海, 2011 ;
郭垚鑫等, 2014 ).本研究中, 中斑块与小斑块之间通过改变微环境相互庇护, 抵御恶劣的环境以提高存活机会, 这种小尺度上正向的相互关联性可能是其应对喀斯特严峻的生境异质性以及应对极端环境的生态对策之一.而大斑块对中、小斑块无关联性, 其占据适宜的环境后, 在生长过程中分泌酸性物质, 促进岩面溶解而自身残骸堆积在岩面上, 有助于加快岩石表面碳循环速率、富集营养元素、涵养水分及土壤的沉积, 为其它高等植物着生在岩石上创造了必要条件(
《广西西南喀斯特生物多样性》编委会, 2011 ).因此在喀斯特地区, 南亚毛灰藓以小斑块和中斑块的状态广泛分布, 存活率较高, 而大斑块可为演替初期做铺垫. ...
喀斯特石漠化过程中苔藓植物多样性及分布与环境关系
1
2015
... 随着环境的变化苔藓植物自身的生长和分布呈现出不同的响应.在不同的生境下, 其分布、群落大小以及盖度都会有明显的差异, 即使在同种生境下其生长也有所不同(
张元明等, 2003 ;
吉雪花等, 2013 ;
皮春燕和刘艳, 2014 ).不同的环境因子对植物群落的格局也有一定的影响(
刘艳等, 2008 ;
宋同清等, 2010 ;
李军峰等, 2015 ).本研究中, 石缝数和坡度都与南亚毛灰藓的3种斑块有很强的关联性, 但是苔藓植物作为变水植物(poikilohydric plants)对水分有很强的依赖性, 石生南亚毛灰藓平铺的生活型通过内吸水和外吸水来增加对水分的涵养.因此相对湿度是石生苔藓斑块大小变化和格局分布的重要因素之一. ...
杭州市区土生苔藓植物分布与生态因子的关系
1
2008
... 随着环境的变化苔藓植物自身的生长和分布呈现出不同的响应.在不同的生境下, 其分布、群落大小以及盖度都会有明显的差异, 即使在同种生境下其生长也有所不同(
张元明等, 2003 ;
吉雪花等, 2013 ;
皮春燕和刘艳, 2014 ).不同的环境因子对植物群落的格局也有一定的影响(
刘艳等, 2008 ;
宋同清等, 2010 ;
李军峰等, 2015 ).本研究中, 石缝数和坡度都与南亚毛灰藓的3种斑块有很强的关联性, 但是苔藓植物作为变水植物(poikilohydric plants)对水分有很强的依赖性, 石生南亚毛灰藓平铺的生活型通过内吸水和外吸水来增加对水分的涵养.因此相对湿度是石生苔藓斑块大小变化和格局分布的重要因素之一. ...
重庆主城区住宅小区苔藓组成与多样性
1
2014
... 随着环境的变化苔藓植物自身的生长和分布呈现出不同的响应.在不同的生境下, 其分布、群落大小以及盖度都会有明显的差异, 即使在同种生境下其生长也有所不同(
张元明等, 2003 ;
吉雪花等, 2013 ;
皮春燕和刘艳, 2014 ).不同的环境因子对植物群落的格局也有一定的影响(
刘艳等, 2008 ;
宋同清等, 2010 ;
李军峰等, 2015 ).本研究中, 石缝数和坡度都与南亚毛灰藓的3种斑块有很强的关联性, 但是苔藓植物作为变水植物(poikilohydric plants)对水分有很强的依赖性, 石生南亚毛灰藓平铺的生活型通过内吸水和外吸水来增加对水分的涵养.因此相对湿度是石生苔藓斑块大小变化和格局分布的重要因素之一. ...
木论喀斯特峰丛洼地森林群落空间格局及环境解释
1
2010
... 随着环境的变化苔藓植物自身的生长和分布呈现出不同的响应.在不同的生境下, 其分布、群落大小以及盖度都会有明显的差异, 即使在同种生境下其生长也有所不同(
张元明等, 2003 ;
吉雪花等, 2013 ;
皮春燕和刘艳, 2014 ).不同的环境因子对植物群落的格局也有一定的影响(
刘艳等, 2008 ;
宋同清等, 2010 ;
李军峰等, 2015 ).本研究中, 石缝数和坡度都与南亚毛灰藓的3种斑块有很强的关联性, 但是苔藓植物作为变水植物(poikilohydric plants)对水分有很强的依赖性, 石生南亚毛灰藓平铺的生活型通过内吸水和外吸水来增加对水分的涵养.因此相对湿度是石生苔藓斑块大小变化和格局分布的重要因素之一. ...
历山山地草甸优势种的种间关联和相关分析
1
2004
... 由于群落生境的差异影响了物种的分布, 关联性有助于进一步认识群落的结构、类型及群落在生境中相互作用所形成的有机联系, 体现了物种之间的竞争关系以及对生境分化的反应(
王琳和张金屯, 2004 ;
姜俊和赵秀海, 2011 ;
郭垚鑫等, 2014 ).本研究中, 中斑块与小斑块之间通过改变微环境相互庇护, 抵御恶劣的环境以提高存活机会, 这种小尺度上正向的相互关联性可能是其应对喀斯特严峻的生境异质性以及应对极端环境的生态对策之一.而大斑块对中、小斑块无关联性, 其占据适宜的环境后, 在生长过程中分泌酸性物质, 促进岩面溶解而自身残骸堆积在岩面上, 有助于加快岩石表面碳循环速率、富集营养元素、涵养水分及土壤的沉积, 为其它高等植物着生在岩石上创造了必要条件(
《广西西南喀斯特生物多样性》编委会, 2011 ).因此在喀斯特地区, 南亚毛灰藓以小斑块和中斑块的状态广泛分布, 存活率较高, 而大斑块可为演替初期做铺垫. ...
长白山北坡林线岳桦种群空间分布格局
1
2015
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
羊草+大针茅草原退化群落优势种群空间点格局分析
1
2011
... 植物种群的空间分布格局是指个体在种群的二维空间水平结构中的分布位置、方式与分布特征.种群的分布格局研究不仅可以揭示种群的形成过程和维持机制, 而且还可以根据种群形成和维持机制推测不同尺度范围内的种群格局、成因和规律, 同时可以对种群的演替方向做出预判(
王鑫厅等, 2011 ).种群分布格局受到不同尺度上生物因子的影响(
Nathan, 2006 ;
高福元和石福习, 2015 ), 同时还受到非生物因子长期相互作用的影响(
Bai et al., 2014 ). ...
1
2002
... 根据喀斯特石漠化等级的划分规则(
熊康宁, 2002 ), 选取潜在石漠化、轻度石漠化和中度石漠化3种生境的样地.每个生境设立50 m×50 m的样方, 总样方面积为0.75 hm
2 .各样地概况见
表1 .在进行实地调查后, 记录样地内所有南亚毛灰藓(
Homomallium simlaense )斑块的盖度和形状变化的范围.依据整体数据将群落中的南亚毛灰藓以盖度大小分为3个等级: 小斑块(<0.014 m
2 )、中斑块(0.014-0.028 m
2 )和大斑块(>0.028 m
2 ).由于苔藓斑块的形状不规则, 以不规则形状的几何中心为南亚毛灰藓斑块在样方中的坐标位置, 记录坐标值(x, y) (位置记录精度为±5 cm).每个样方以西南角为原点开始记录.在记录坐标的同时, 记录其盖度和5个环境因子: 坡度、郁闭度、相对湿度、石缝数和小生境.其中石缝数指在苔藓斑块下所有长>2 cm、宽>1 mm、深>2 mm的石缝数目.小生境分为石面、石壁、石沟、石坑和石洞5种(
张天汉等, 2014 ). ...
上海市树附生苔藓植物分布格局研究
1
2006
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
翅果油树群落主要物种空间分布格局及其关联性
1
2016
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
毛乌素沙地油蒿种群点格局分析
1
2006
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
植物种群空间分布的点格局分析
1
1998
... 本研究基于二阶统计方法O-ring统计(
张金屯, 1998 ;
Wiegand and Moloney, 2004 )的单变量和双变量分析南亚毛灰藓在不同尺度上的分布格局、不同斑块间的空间关联性及不同石漠化生境中斑块的空间分布规律; 同时应用主成分分析(principal component analysis, PCA)探讨环境因子对不同斑块南亚毛灰藓分布格局的影响.通过尝试从不同大小斑块分布格局及其影响因素的角度来揭示石漠化地区南亚毛灰藓的生态学特性和形成过程, 以期为喀斯特石漠化地区裸露岩石的生态恢复及环境指示提供理论依据. ...
西南喀斯特石漠化治理植物选择与生态适应性
2
2015
... 喀斯特脆弱的生态系统一直是中外****的研究热点, 而我国西南地区是全球喀斯特地貌较为集中的区域.由于岩石裸露率高及土层瘠薄等条件限制, 加之人为干扰, 石漠化不断加重的问题亟待解决(
郭柯等, 2011 ;
张军以等, 2015 ).南亚毛灰藓(
Homoma- llium simlaense )作为喀斯特地区常见的一种藓类植物, 主要以平铺的生活型(
Mägdefrau,1982 )生长在完全裸露的岩石上, 极具耐旱性.南亚毛灰藓能够成片地生长在岩石表面, 形成不同的斑块, 并积累养分改善微环境, 为其它被子植物的着生提供适宜的环境条件.同时, 它还作为喀斯特地区演替中的岩溶先锋植物扮演着非常重要的角色(
Glime, 2006 ). ...
... 自然条件下, 种群的结构和空间分布往往是种群生物学特性与环境相互作用的结果(
Condit et al., 2000 ).不同斑块南亚毛灰藓的分布规律揭示了在喀斯特地区岩石先锋种苔藓植物在自然演替中的生态学进程, 同时不同斑块与环境变化的关系阐释了南亚毛灰藓在喀斯特地区的适应性.石生南亚毛灰藓不同斑块的分布不仅受孢子扩散和生境异质性的影响, 而且不同的环境因子和一定的环境胁迫也会对南亚毛灰藓的分布起重要作用.对于生态恢复来说, 我国西南部石漠化地区的生态恢复多是以种植经济林和速生林为主(
张军以等, 2015 ), 忽略了裸露岩石的恢复.而根据喀斯特石生苔藓的分布特点, 在郁闭度和湿度条件较好的石面上可培养大斑块的石生苔藓, 为景观或其它植物的生存创造条件.而在郁闭度低、湿度低和石漠化严重的区域, 可在环境条件相对优越的小生境中以小斑块散点模式培养苔藓, 以提高成活率; 同时待苔藓成活后可改变微环境使其相互庇护, 提高抗干旱和高温等环境胁迫的能力. ...
不同密度条件下芨芨草空间格局对环境胁迫的响应
1
2012
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
贵州关岭县喀斯特峰丛石漠区苔藓群落生态特征
1
2014
... 根据喀斯特石漠化等级的划分规则(
熊康宁, 2002 ), 选取潜在石漠化、轻度石漠化和中度石漠化3种生境的样地.每个生境设立50 m×50 m的样方, 总样方面积为0.75 hm
2 .各样地概况见
表1 .在进行实地调查后, 记录样地内所有南亚毛灰藓(
Homomallium simlaense )斑块的盖度和形状变化的范围.依据整体数据将群落中的南亚毛灰藓以盖度大小分为3个等级: 小斑块(<0.014 m
2 )、中斑块(0.014-0.028 m
2 )和大斑块(>0.028 m
2 ).由于苔藓斑块的形状不规则, 以不规则形状的几何中心为南亚毛灰藓斑块在样方中的坐标位置, 记录坐标值(x, y) (位置记录精度为±5 cm).每个样方以西南角为原点开始记录.在记录坐标的同时, 记录其盖度和5个环境因子: 坡度、郁闭度、相对湿度、石缝数和小生境.其中石缝数指在苔藓斑块下所有长>2 cm、宽>1 mm、深>2 mm的石缝数目.小生境分为石面、石壁、石沟、石坑和石洞5种(
张天汉等, 2014 ). ...
植物间正相互作用对种群动态和群落结构的影响: 基于个体模型的研究进展
1
2013
... 胁迫梯度假说(stress gradient hypothesis, SGH)认为, 随胁迫强度的增加, 植物间正相互作用增强(
Callaway and Walker, 1997 ;
Callaway, 2007 ;
Brooker et al., 2008 ;
张炜平等, 2013 ).综合本研究基于不同尺度下不同斑块的空间分布与环境的关系, 从相对湿度和郁闭度胁迫来说, 大斑块在环境因子中受到的环境胁迫较小, 小斑块最大.因此小斑块和中斑块随环境胁迫增大而趋于聚集分布.这基本符合胁迫梯度假说, 也为其提供了数据支撑, 但仍需要进行受控实验以确定斑块间相互作用特性及其随环境胁迫变化的生态机制.以上结果表明, 不同斑块的分布也受到一定的环境胁迫影响. ...
新疆博格达山地面生苔藓植物物种多样性研究
1
2003
... 随着环境的变化苔藓植物自身的生长和分布呈现出不同的响应.在不同的生境下, 其分布、群落大小以及盖度都会有明显的差异, 即使在同种生境下其生长也有所不同(
张元明等, 2003 ;
吉雪花等, 2013 ;
皮春燕和刘艳, 2014 ).不同的环境因子对植物群落的格局也有一定的影响(
刘艳等, 2008 ;
宋同清等, 2010 ;
李军峰等, 2015 ).本研究中, 石缝数和坡度都与南亚毛灰藓的3种斑块有很强的关联性, 但是苔藓植物作为变水植物(poikilohydric plants)对水分有很强的依赖性, 石生南亚毛灰藓平铺的生活型通过内吸水和外吸水来增加对水分的涵养.因此相对湿度是石生苔藓斑块大小变化和格局分布的重要因素之一. ...
茂兰喀斯特常绿落叶阔叶混交林树种的空间分布格局及其分形特征
1
2015
... 在空间分布格局研究方面, 乔木种群和群落的格局研究较多(
韩文衡等, 2010 ;
张忠华等, 2015 ;
郭屹立等, 2015 ;
王晓雨等, 2015 ;
许强等, 2016 ).在格局方法研究中有****将芨芨草(
Achnatherum splendens )看作一个“丛”进行格局分析(
张明娟等, 2012 ); 还有****对油蒿(
Artemisia ordosica )和苔藓以斑块形式进行空间分布特征研究(
杨洪晓等, 2006 ;
吉雪花等, 2014 ).在对苔藓植物格局的研究中, 有分析地被层中苔藓植物的分布格局(
郭水良和曹同, 2000 ); 还有对树附生苔藓植物的分布格局进行研究(
徐晟翀等, 2006 ).而使用点格局方法分析喀斯特(Karst)地区特殊的苔藓植物群落空间格局的研究相对较少. ...
1
2014
... 植物种群的空间分布格局是指个体在种群的二维空间水平结构中的分布位置、方式与分布特征.种群的分布格局研究不仅可以揭示种群的形成过程和维持机制, 而且还可以根据种群形成和维持机制推测不同尺度范围内的种群格局、成因和规律, 同时可以对种群的演替方向做出预判(
王鑫厅等, 2011 ).种群分布格局受到不同尺度上生物因子的影响(
Nathan, 2006 ;
高福元和石福习, 2015 ), 同时还受到非生物因子长期相互作用的影响(
Bai et al., 2014 ). ...
1
2008
... 胁迫梯度假说(stress gradient hypothesis, SGH)认为, 随胁迫强度的增加, 植物间正相互作用增强(
Callaway and Walker, 1997 ;
Callaway, 2007 ;
Brooker et al., 2008 ;
张炜平等, 2013 ).综合本研究基于不同尺度下不同斑块的空间分布与环境的关系, 从相对湿度和郁闭度胁迫来说, 大斑块在环境因子中受到的环境胁迫较小, 小斑块最大.因此小斑块和中斑块随环境胁迫增大而趋于聚集分布.这基本符合胁迫梯度假说, 也为其提供了数据支撑, 但仍需要进行受控实验以确定斑块间相互作用特性及其随环境胁迫变化的生态机制.以上结果表明, 不同斑块的分布也受到一定的环境胁迫影响. ...
1
2007
... 胁迫梯度假说(stress gradient hypothesis, SGH)认为, 随胁迫强度的增加, 植物间正相互作用增强(
Callaway and Walker, 1997 ;
Callaway, 2007 ;
Brooker et al., 2008 ;
张炜平等, 2013 ).综合本研究基于不同尺度下不同斑块的空间分布与环境的关系, 从相对湿度和郁闭度胁迫来说, 大斑块在环境因子中受到的环境胁迫较小, 小斑块最大.因此小斑块和中斑块随环境胁迫增大而趋于聚集分布.这基本符合胁迫梯度假说, 也为其提供了数据支撑, 但仍需要进行受控实验以确定斑块间相互作用特性及其随环境胁迫变化的生态机制.以上结果表明, 不同斑块的分布也受到一定的环境胁迫影响. ...
1
1997
... 胁迫梯度假说(stress gradient hypothesis, SGH)认为, 随胁迫强度的增加, 植物间正相互作用增强(
Callaway and Walker, 1997 ;
Callaway, 2007 ;
Brooker et al., 2008 ;
张炜平等, 2013 ).综合本研究基于不同尺度下不同斑块的空间分布与环境的关系, 从相对湿度和郁闭度胁迫来说, 大斑块在环境因子中受到的环境胁迫较小, 小斑块最大.因此小斑块和中斑块随环境胁迫增大而趋于聚集分布.这基本符合胁迫梯度假说, 也为其提供了数据支撑, 但仍需要进行受控实验以确定斑块间相互作用特性及其随环境胁迫变化的生态机制.以上结果表明, 不同斑块的分布也受到一定的环境胁迫影响. ...
1
2000
... 自然条件下, 种群的结构和空间分布往往是种群生物学特性与环境相互作用的结果(
Condit et al., 2000 ).不同斑块南亚毛灰藓的分布规律揭示了在喀斯特地区岩石先锋种苔藓植物在自然演替中的生态学进程, 同时不同斑块与环境变化的关系阐释了南亚毛灰藓在喀斯特地区的适应性.石生南亚毛灰藓不同斑块的分布不仅受孢子扩散和生境异质性的影响, 而且不同的环境因子和一定的环境胁迫也会对南亚毛灰藓的分布起重要作用.对于生态恢复来说, 我国西南部石漠化地区的生态恢复多是以种植经济林和速生林为主(
张军以等, 2015 ), 忽略了裸露岩石的恢复.而根据喀斯特石生苔藓的分布特点, 在郁闭度和湿度条件较好的石面上可培养大斑块的石生苔藓, 为景观或其它植物的生存创造条件.而在郁闭度低、湿度低和石漠化严重的区域, 可在环境条件相对优越的小生境中以小斑块散点模式培养苔藓, 以提高成活率; 同时待苔藓成活后可改变微环境使其相互庇护, 提高抗干旱和高温等环境胁迫的能力. ...
1
2006
... 喀斯特脆弱的生态系统一直是中外****的研究热点, 而我国西南地区是全球喀斯特地貌较为集中的区域.由于岩石裸露率高及土层瘠薄等条件限制, 加之人为干扰, 石漠化不断加重的问题亟待解决(
郭柯等, 2011 ;
张军以等, 2015 ).南亚毛灰藓(
Homoma- llium simlaense )作为喀斯特地区常见的一种藓类植物, 主要以平铺的生活型(
Mägdefrau,1982 )生长在完全裸露的岩石上, 极具耐旱性.南亚毛灰藓能够成片地生长在岩石表面, 形成不同的斑块, 并积累养分改善微环境, 为其它被子植物的着生提供适宜的环境条件.同时, 它还作为喀斯特地区演替中的岩溶先锋植物扮演着非常重要的角色(
Glime, 2006 ). ...
1
1982
... 喀斯特脆弱的生态系统一直是中外****的研究热点, 而我国西南地区是全球喀斯特地貌较为集中的区域.由于岩石裸露率高及土层瘠薄等条件限制, 加之人为干扰, 石漠化不断加重的问题亟待解决(
郭柯等, 2011 ;
张军以等, 2015 ).南亚毛灰藓(
Homoma- llium simlaense )作为喀斯特地区常见的一种藓类植物, 主要以平铺的生活型(
Mägdefrau,1982 )生长在完全裸露的岩石上, 极具耐旱性.南亚毛灰藓能够成片地生长在岩石表面, 形成不同的斑块, 并积累养分改善微环境, 为其它被子植物的着生提供适宜的环境条件.同时, 它还作为喀斯特地区演替中的岩溶先锋植物扮演着非常重要的角色(
Glime, 2006 ). ...
1
2006
... 植物种群的空间分布格局是指个体在种群的二维空间水平结构中的分布位置、方式与分布特征.种群的分布格局研究不仅可以揭示种群的形成过程和维持机制, 而且还可以根据种群形成和维持机制推测不同尺度范围内的种群格局、成因和规律, 同时可以对种群的演替方向做出预判(
王鑫厅等, 2011 ).种群分布格局受到不同尺度上生物因子的影响(
Nathan, 2006 ;
高福元和石福习, 2015 ), 同时还受到非生物因子长期相互作用的影响(
Bai et al., 2014 ). ...
1
2011
... 物种的空间分布格局与尺度有着密切联系, 并随着尺度的变化而变化.在小尺度上, 空间格局和空间关联性可能是由于生境、竞争以及种子扩散等因素造成.而在大尺度上, 则可能是生境异质性和种子的扩散限制导致的(
Rayburn et al., 2011 ;
Shen et al., 2013 ).南亚毛灰藓孢子小且轻, 局部的扩散会使多数孢子落在配子体附近.重庆中梁山喀斯特地区地形复杂, 小生境异质性大, 石沟、石槽使孢子在扩散过程中容易被阻挡.扩散限制和生境条件的异质性使得小斑块和中斑块在小尺度上为聚集分布.而大斑块占据着水分条件相对优越且其它条件也较好的生境, 抗旱能力增强、相互依赖性减弱, 从而使竞争力增大, 因而大斑块多为随机分布. ...
1
2013
... 物种的空间分布格局与尺度有着密切联系, 并随着尺度的变化而变化.在小尺度上, 空间格局和空间关联性可能是由于生境、竞争以及种子扩散等因素造成.而在大尺度上, 则可能是生境异质性和种子的扩散限制导致的(
Rayburn et al., 2011 ;
Shen et al., 2013 ).南亚毛灰藓孢子小且轻, 局部的扩散会使多数孢子落在配子体附近.重庆中梁山喀斯特地区地形复杂, 小生境异质性大, 石沟、石槽使孢子在扩散过程中容易被阻挡.扩散限制和生境条件的异质性使得小斑块和中斑块在小尺度上为聚集分布.而大斑块占据着水分条件相对优越且其它条件也较好的生境, 抗旱能力增强、相互依赖性减弱, 从而使竞争力增大, 因而大斑块多为随机分布. ...
2
2004
... 本研究基于二阶统计方法O-ring统计(
张金屯, 1998 ;
Wiegand and Moloney, 2004 )的单变量和双变量分析南亚毛灰藓在不同尺度上的分布格局、不同斑块间的空间关联性及不同石漠化生境中斑块的空间分布规律; 同时应用主成分分析(principal component analysis, PCA)探讨环境因子对不同斑块南亚毛灰藓分布格局的影响.通过尝试从不同大小斑块分布格局及其影响因素的角度来揭示石漠化地区南亚毛灰藓的生态学特性和形成过程, 以期为喀斯特石漠化地区裸露岩石的生态恢复及环境指示提供理论依据. ...
... 在分析南亚毛灰藓空间格局和空间关联性过程中, 为精确分析设定栅格大小为1 m×1 m, 圆环宽度为3 m, 设置0-50 m的研究尺度, 并经过99次Monte Carlo模拟得到95%的置信区间.数据分析采用Programita (
Wiegand and Moloney, 2004 )软件完成.使用Excel 2013软件处理数据.使用Origin 9.0软件作图.主成分分析使用CANOCO软件完成. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}