张盛春, 李清明, 阳成伟*, 华南师范大学生命科学学院, 广东省植物发育生物工程重点实验室, 广州 510631 ZhangShengchun, LiQingming, YangChengwei*, Guangdong Provincial Key Laboratory of Biotechnology for Plant Development, College of Life Sciences, South China Normal University, Guangzhou 510631, China 引用本文 张盛春, 李清明, 阳成伟. 拟南芥金属蛋白酶FtSH4通过生长素与活性氧调控叶片衰老. , 2017, 52(4): 453-464
Abstract: The plant metalloproteinases FtSH gene family has 12 members in Arabidopsis, and their functions are still unclear. In the present study, we analyzed the function of FtSH4 on leaf senescence using cell biology and genetics methods. The ftsh4-4 mutant displayed a premature leaf senescence phenotype with increased H2O2 content and cell death rate, decreased chlorophyll content, increased peroxidase gene expression and peroxidase activity. The ftsh4-4 leaf senescence phenotype could be rescued by applying the exogenous antioxidant AsA and endogenous or exogenous auxin by decreasing H2O2 content, peroxidase gene expression level and peroxidase activity. The expression of auxin response factor genes ARF2 and ARF7 was increased in the ftsh4-4 mutant and was reduced by exogenous auxin or AsA. Moreover, H2O2 content and the senescence phenotype of ftsh4-4 could be rescued by the arf2-8 mutant. These results indicate that FtSH4 gene plays an important role in the regulation of leaf senescence through auxin and reactive oxygen species.
植物叶片衰老是一种依赖年龄的细胞程序性退化和降解过程, 能够引起植物体内的代谢紊乱并最终导致植物器官或整株死亡(Lim et al., 2007)。植物叶片在衰老过程中, 由于细胞质膜和液泡膜以及叶绿体等细胞器发生瓦解, 破坏了细胞的内稳态, 最终导致衰老叶片细胞死亡。叶片衰老相关代谢变化包括叶绿素降解、叶片黄化, 还有蛋白质、脂质、核酸以及其它生物大分子的水解(Ren et al., 2010; Watanabe et al., 2013)。叶片衰老以依赖年龄的方式进行, 属于程序性细胞死亡的一种, 受到各种外部环境因素和内部因素的诱导(Guo and Gan, 2012)。影响植物叶片衰老的内部因素包括各种植物激素、植物体内的氧化还原状态以及依赖年龄的内源发育信号(Lim et al., 2007)。
植物中FtSH (filamentation temperature sensitive H)等ATP依赖的金属蛋白酶在植物生长发育过程中, 尤其是维持叶绿体和线粒体膜蛋白的完整性方面起重要作用(Nolden et al., 2005)。拟南芥基因组共编码12个FtSH蛋白(Garcia-Lorenzo et al., 2006), 其中有8个定位在叶绿体(FtSH1、2、5-9和12), 3个定位在线粒体中(FtSH3、4和10), FtSH11同时存在于叶绿体和线粒体中(Kolodziejczak et al., 2002)。我们前期的研究表明, 线粒体FtSH4基因突变会导致H2O2大量积累(Zhang et al., 2014), 叶片出现早衰表型(Zhang et al., 2017)。本研究表明, H2O2清除剂和生长素能延缓ftsh4突变体的早衰表型, 提示该基因可能在H2O2与生长素调控叶片衰老过程中发挥重要作用。本研究通过细胞生物学和遗传学等方法, 在前期研究的基础上, 进一步对线粒体FtSH4通过生长素与H2O2调控叶片衰老的分子机制进行了深入研究。
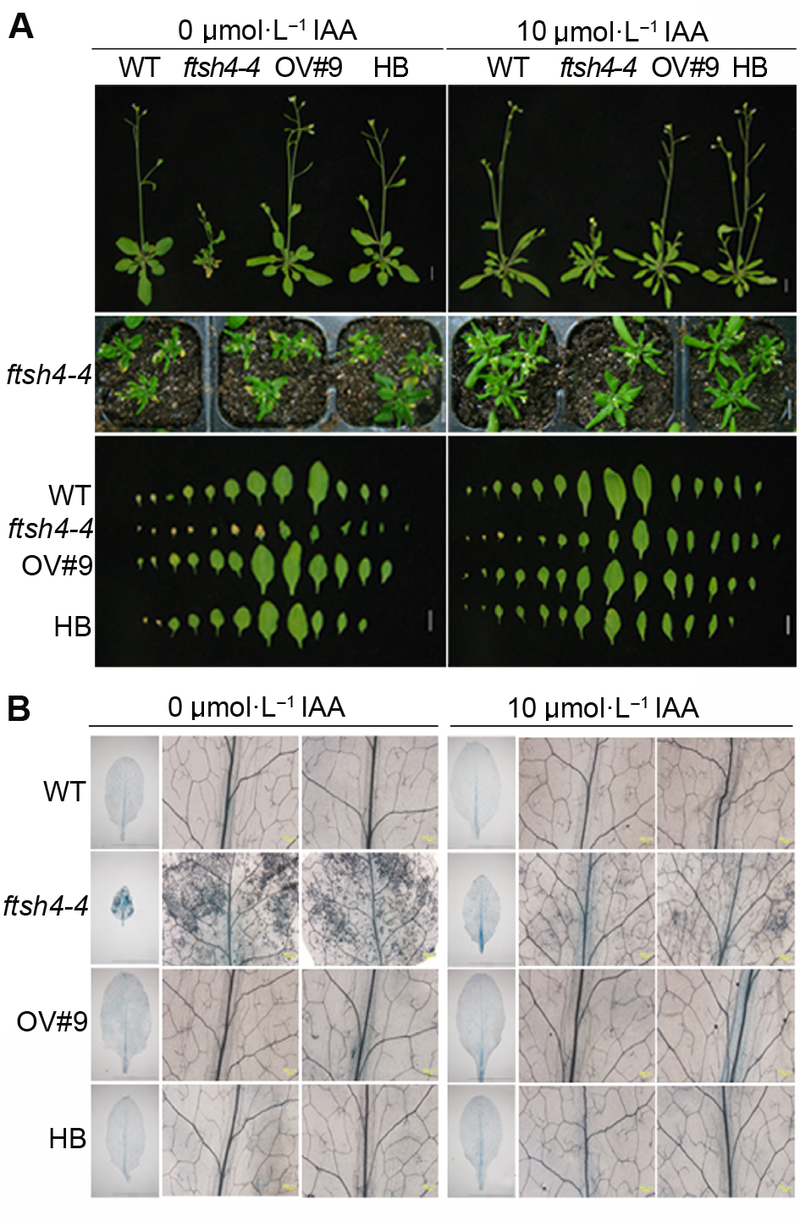
图3 外源生长素(IAA)能恢复拟南芥ftsh4-4突变体早衰表型^(A) 外源IAA处理能恢复ftsh4-4突变体早衰表型(Bar=1 cm); (B) 外源IAA处理能减少ftsh4-4突变体中细胞死亡数目 Figure 3 Exogenous IAA restored the leaf senescence phenotype of Arabidopsis ftsh4-4 mutant^(A) Exogenous IAA restored the leaf senescence phenotype of ftsh4-4 mutant (Bar=1 cm); (B) Exogenous IAA reduced the cell death of ftsh4-4 mutant
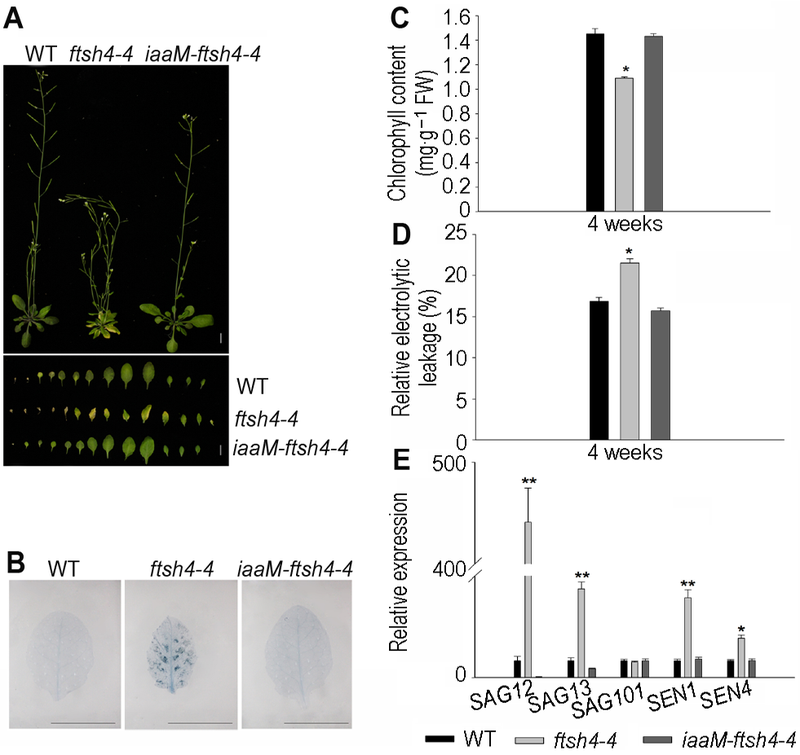
为了进一步确定ftsh4-4突变体表现出的早衰表型是由IAA含量减少引起的, 我们构建了iaaM-ftsh4-4转基因植株(Zhang et al., 2014)。iaaM编码色氨酸单加氧酶(IAAM), IAAM是生长素合成途径吲哚乙酰胺途径中的关键酶, 能催化色氨酸转化为吲哚乙酰胺(IAM), 随后IAM再通过吲哚乙酰胺水解酶(IAAH)水解成IAA (Camilleri and Jouanin, 1991; Romano et al., 1995)。与ftsh4-4突变体相比, iaaM-ftsh4-4转基因植株中内源IAA含量增加(Zhang et al., 2014)。iaaM-ftsh4-4能恢复ftsh4-4突变体的叶片早衰表型, 表现出与WT一致的表型(图4A)。对4周龄拟南芥植株莲座叶进行乳酚-台盼蓝染色, 结果显示, 与ftsh4-4突变体相比, iaaM-ftsh4-4转基因植物细胞死亡数目明显减少(图4B), 并且iaaM-ftsh4-4植株的叶绿素 含量(图4C)及相对电导率(图4D)等衰老生理指标均 恢复到与野生型相同的正常水平。我们还利用qRT- PCR检测了生长4周的WT、ftsh4-4和iaaM-ftsh4-4植株莲座叶中衰老相关标记基因SAG12、SAG13、SAG101、SEN1及SEN4的表达水平, 发现与WT相比, ftsh4-4突变体植株莲座叶中SAG12、SAG13、SEN1及SEN4表达量分别增加了440倍、4.2倍、3.7倍和1.3倍, 而iaaM-ftsh4-4能明显将ftsh4-4突变体中SAG12、SAG13、SEN1及SEN4的表达水平降低到WT水平(图4E)。以上结果表明, 增加内源生长素含量能恢复ftsh4-4突变体的早衰表型, 即ftsh4-4突变体表现出的早衰表型确实是由于IAA含量减少引起的。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-453/img_5.png<b>图4</b> 增加内源生长素含量能恢复拟南芥<i>ftsh4-4</i>突变体早衰表型^(A) 增加内源IAA含量能恢复<i>ftsh4-4</i>突变体早衰表型(Bar=1 cm); (B) <i>iaaM-ftsh4-4</i>中细胞死亡数目减少(Bar=1 cm); (C) <i>iaaM-ftsh4-4</i>中叶绿素含量恢复到WT水平; (D) <i>iaaM-ftsh4-4</i>中相对电导率恢复到WT水平; (E) 内源激素降低<i>ftsh4-4</i>突变体中衰老基因<i>SAG12</i>、<i>SAG13</i>、<i>SAG101</i>、<i>SEN1</i>及<i>SEN4</i>的表达。* 表示差异显著(<i>P</i><0.05); ** 表示差异极显著(<i>P</i><0.01) (Student’s<i> t</i>-test)。<br/><b>Figure 4</b> Increasing endogenous IAA restored the leaf senescence phenotype of Arabidopsis<i> ftsh4-4</i> mutant^(A) Increasing endogenous IAA restored the leaf senescence phenotype of <i>ftsh4-4</i> mutant (Bar=1 cm); (B) Cell death decreased in <i>iaaM-ftsh4-4</i> (Bar=1 cm); (C) The chlorophyll content of <i>iaaM-ftsh4-4</i> transgeneic line restored to the wild type level; (D) The relative electrolytic leakage of <i>iaaM- ftsh4-4</i> transgeneic line restored to the wild type level; (E) Endogenous IAA decreased the expression of senescence-associated genes <i>SAG12</i>,<i> SAG13</i>, <i>SAG101</i>,<i> SEN1</i> and <i>SEN4</i> in <i>ftsh4-4</i> mutant. * indicates significant difference at <i>P</i><0.05; ** indicates significant difference at <i>P</i><0.01 (Student’s<i> t</i>-test). Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-453/img_5.png<b>图4</b> 增加内源生长素含量能恢复拟南芥<i>ftsh4-4</i>突变体早衰表型^(A) 增加内源IAA含量能恢复<i>ftsh4-4</i>突变体早衰表型(Bar=1 cm); (B) <i>iaaM-ftsh4-4</i>中细胞死亡数目减少(Bar=1 cm); (C) <i>iaaM-ftsh4-4</i>中叶绿素含量恢复到WT水平; (D) <i>iaaM-ftsh4-4</i>中相对电导率恢复到WT水平; (E) 内源激素降低<i>ftsh4-4</i>突变体中衰老基因<i>SAG12</i>、<i>SAG13</i>、<i>SAG101</i>、<i>SEN1</i>及<i>SEN4</i>的表达。* 表示差异显著(<i>P</i><0.05); ** 表示差异极显著(<i>P</i><0.01) (Student’s<i> t</i>-test)。<br/><b>Figure 4</b> Increasing endogenous IAA restored the leaf senescence phenotype of Arabidopsis<i> ftsh4-4</i> mutant^(A) Increasing endogenous IAA restored the leaf senescence phenotype of <i>ftsh4-4</i> mutant (Bar=1 cm); (B) Cell death decreased in <i>iaaM-ftsh4-4</i> (Bar=1 cm); (C) The chlorophyll content of <i>iaaM-ftsh4-4</i> transgeneic line restored to the wild type level; (D) The relative electrolytic leakage of <i>iaaM- ftsh4-4</i> transgeneic line restored to the wild type level; (E) Endogenous IAA decreased the expression of senescence-associated genes <i>SAG12</i>,<i> SAG13</i>, <i>SAG101</i>,<i> SEN1</i> and <i>SEN4</i> in <i>ftsh4-4</i> mutant. * indicates significant difference at <i>P</i><0.05; ** indicates significant difference at <i>P</i><0.01 (Student’s<i> t</i>-test).
图4 增加内源生长素含量能恢复拟南芥ftsh4-4突变体早衰表型^(A) 增加内源IAA含量能恢复ftsh4-4突变体早衰表型(Bar=1 cm); (B) iaaM-ftsh4-4中细胞死亡数目减少(Bar=1 cm); (C) iaaM-ftsh4-4中叶绿素含量恢复到WT水平; (D) iaaM-ftsh4-4中相对电导率恢复到WT水平; (E) 内源激素降低ftsh4-4突变体中衰老基因SAG12、SAG13、SAG101、SEN1及SEN4的表达。* 表示差异显著(P<0.05); ** 表示差异极显著(P<0.01) (Student’s t-test)。 Figure 4 Increasing endogenous IAA restored the leaf senescence phenotype of Arabidopsis ftsh4-4 mutant^(A) Increasing endogenous IAA restored the leaf senescence phenotype of ftsh4-4 mutant (Bar=1 cm); (B) Cell death decreased in iaaM-ftsh4-4 (Bar=1 cm); (C) The chlorophyll content of iaaM-ftsh4-4 transgeneic line restored to the wild type level; (D) The relative electrolytic leakage of iaaM- ftsh4-4 transgeneic line restored to the wild type level; (E) Endogenous IAA decreased the expression of senescence-associated genes SAG12, SAG13, SAG101, SEN1 and SEN4 in ftsh4-4 mutant. * indicates significant difference at P<0.05; ** indicates significant difference at P<0.01 (Student’s t-test).
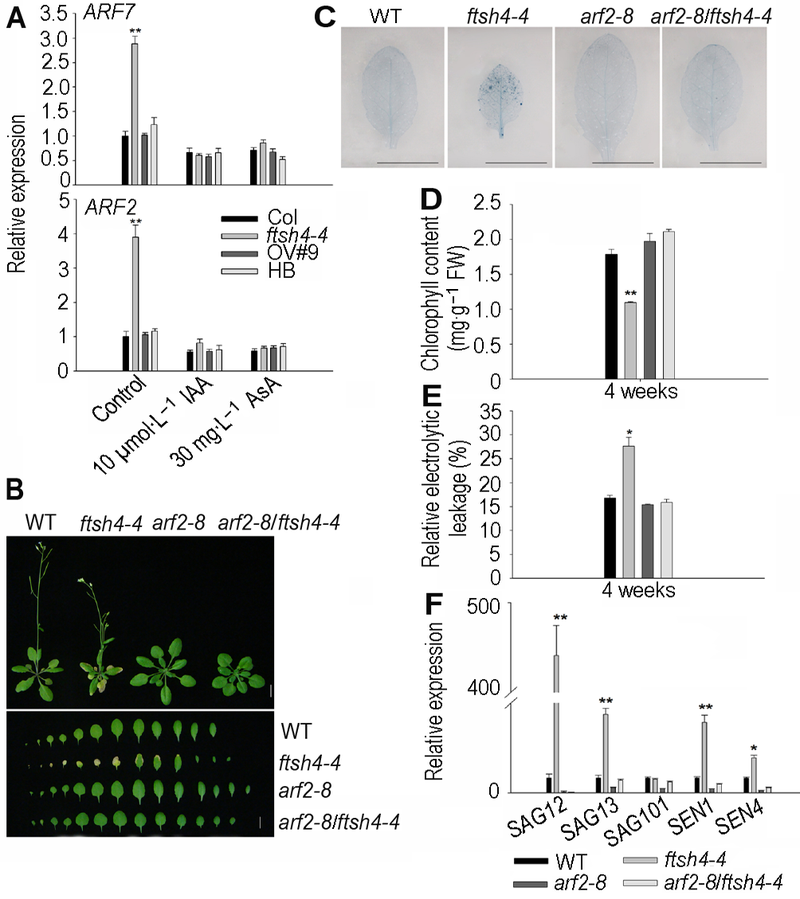
2.4 ARF2参与FtSH4介导的植物叶片衰老过程为了分析ftsh4-4突变体出现的早衰表型是否与生长素响应因子ARF有关, 我们利用qRT-PCR技术检测了ARF2和ARF7在ftsh4-4突变体莲座叶中的表达变化。在3周龄植物中, ARF2和ARF7在ftsh4-4中的表达明显比WT高(图5A)。为了进一步探明ftsh4-4突变体的早衰表型是否是由于ARF2和ARF7表达异常所致, 我们检测了10 µmol∙L-1 IAA和30 mg∙L-1 AsA处理后不同FtSH4基因型植株莲座叶中ARF2和ARF7的表达变化。结果表明, IAA或者AsA处理后, ftsh4-4突变体莲座叶中ARF2和ARF7的表达与其它基因型植物相比没有明显差异(图5A)。由此推测, 生长素响应因子基因ARF2和ARF7可能参与了FtSH4介导的植物叶片衰老过程。 图5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-453/img_6.png<b>图5</b> <i>ARF2</i>参与<i>FtSH4</i>介导的拟南芥叶片衰老过程^(A) 生长素响应因子基因<i>ARF2</i>和<i>ARF7</i>在<i>ftsh4-4</i>中上调表达并受外源IAA和AsA抑制; (B) <i>ARF2</i>突变抑制了<i>ftsh4-4</i>的叶片衰老(Bar=1 cm); (C) <i>ARF2</i>突变使<i>ftsh4-4</i>中细胞死亡数目减少(Bar=1 cm); (D) <i>ARF2</i>突变使<i>ftsh4-4</i>中叶绿素含量增加; (E) <i>ARF2</i>突变使<i>ftsh4-4</i>中相对电导率降低; (F) <i>ARF2</i>突变使<i>ftsh4-4</i>中衰老相关标记基因<i>SAG12</i>、<i>SAG13</i>、<i>SAG101</i>、<i>SEN1</i>及<i>SEN4</i>下调表达。* 表示差异显著(<i>P</i><0.05); ** 表示差异极显著(<i>P</i><0.01) (Student’s<i> t</i>-test)。<br/><b>Figure 5</b> <i>ARF2 </i>is involved in <i>FtSH4</i>-mediated leaf senescence of Arabidopsis^(A) The expression levels of <i>ARF2</i> and <i>ARF7</i> increased in the <i>ftsh4-4</i> and were inhibited by the exogenous IAA and AsA; (B) <i>ARF2</i> mutation rescued the leaf senescence of <i>ftsh4-4 </i>mutant (Bar=1 cm); (C)<i> ARF2</i> mutation reduced the cell death of<i> ftsh4-4 </i>mutant (Bar=1 cm); (D) <i>ARF2</i> mutation increased the chlorophyll content of<i> ftsh4-4 </i>mutant; (E)<i> ARF2</i> mutation reduced the relative electrolytic leakage of<i> ftsh4-4 </i>mutant; (F) <i>ARF2</i> mutation reduced the expression of senescence-associated genes <i>SAG12</i>, <i>SAG13</i>, <i>SAG101</i>, <i>SEN1</i> and <i>SEN4</i>. * indicates significant difference at <i>P</i><0.05; ** indicates significant difference at <i>P</i><0.01 (Student’s<i> t</i>-test). Figure 5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-453/img_6.png<b>图5</b> <i>ARF2</i>参与<i>FtSH4</i>介导的拟南芥叶片衰老过程^(A) 生长素响应因子基因<i>ARF2</i>和<i>ARF7</i>在<i>ftsh4-4</i>中上调表达并受外源IAA和AsA抑制; (B) <i>ARF2</i>突变抑制了<i>ftsh4-4</i>的叶片衰老(Bar=1 cm); (C) <i>ARF2</i>突变使<i>ftsh4-4</i>中细胞死亡数目减少(Bar=1 cm); (D) <i>ARF2</i>突变使<i>ftsh4-4</i>中叶绿素含量增加; (E) <i>ARF2</i>突变使<i>ftsh4-4</i>中相对电导率降低; (F) <i>ARF2</i>突变使<i>ftsh4-4</i>中衰老相关标记基因<i>SAG12</i>、<i>SAG13</i>、<i>SAG101</i>、<i>SEN1</i>及<i>SEN4</i>下调表达。* 表示差异显著(<i>P</i><0.05); ** 表示差异极显著(<i>P</i><0.01) (Student’s<i> t</i>-test)。<br/><b>Figure 5</b> <i>ARF2 </i>is involved in <i>FtSH4</i>-mediated leaf senescence of Arabidopsis^(A) The expression levels of <i>ARF2</i> and <i>ARF7</i> increased in the <i>ftsh4-4</i> and were inhibited by the exogenous IAA and AsA; (B) <i>ARF2</i> mutation rescued the leaf senescence of <i>ftsh4-4 </i>mutant (Bar=1 cm); (C)<i> ARF2</i> mutation reduced the cell death of<i> ftsh4-4 </i>mutant (Bar=1 cm); (D) <i>ARF2</i> mutation increased the chlorophyll content of<i> ftsh4-4 </i>mutant; (E)<i> ARF2</i> mutation reduced the relative electrolytic leakage of<i> ftsh4-4 </i>mutant; (F) <i>ARF2</i> mutation reduced the expression of senescence-associated genes <i>SAG12</i>, <i>SAG13</i>, <i>SAG101</i>, <i>SEN1</i> and <i>SEN4</i>. * indicates significant difference at <i>P</i><0.05; ** indicates significant difference at <i>P</i><0.01 (Student’s<i> t</i>-test).
图5 ARF2参与FtSH4介导的拟南芥叶片衰老过程^(A) 生长素响应因子基因ARF2和ARF7在ftsh4-4中上调表达并受外源IAA和AsA抑制; (B) ARF2突变抑制了ftsh4-4的叶片衰老(Bar=1 cm); (C) ARF2突变使ftsh4-4中细胞死亡数目减少(Bar=1 cm); (D) ARF2突变使ftsh4-4中叶绿素含量增加; (E) ARF2突变使ftsh4-4中相对电导率降低; (F) ARF2突变使ftsh4-4中衰老相关标记基因SAG12、SAG13、SAG101、SEN1及SEN4下调表达。* 表示差异显著(P<0.05); ** 表示差异极显著(P<0.01) (Student’s t-test)。 Figure 5 ARF2 is involved in FtSH4-mediated leaf senescence of Arabidopsis^(A) The expression levels of ARF2 and ARF7 increased in the ftsh4-4 and were inhibited by the exogenous IAA and AsA; (B) ARF2 mutation rescued the leaf senescence of ftsh4-4 mutant (Bar=1 cm); (C) ARF2 mutation reduced the cell death of ftsh4-4 mutant (Bar=1 cm); (D) ARF2 mutation increased the chlorophyll content of ftsh4-4 mutant; (E) ARF2 mutation reduced the relative electrolytic leakage of ftsh4-4 mutant; (F) ARF2 mutation reduced the expression of senescence-associated genes SAG12, SAG13, SAG101, SEN1 and SEN4. * indicates significant difference at P<0.05; ** indicates significant difference at P<0.01 (Student’s t-test).
ApelK, HirtH (2004). Reactive oxygen species: metabolism, oxidative stress, and signal transduction.Annu Rev Plant Biol 55, 373-399. [本文引用: 1]
[2]
ArnonDI (1949). Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol 24, 1-15. [本文引用: 1]
[3]
BashandyT, GuilleminotJ, VernouxT, Caparros-RuizD, LjungK, MeyerY, ReichheldJP (2010). Interplay bet- ween the NADP-linked thioredoxin and glutathione systems in Arabidopsis auxin signaling.Plant Cell 22 376-391. [本文引用: 1]
[4]
BlomsterT, SalojarviJ, SipariN, BroscheM, AhlforsR, KeinanenM, OvermyerK, KangasjarviJ (2011). Apo- plastic reactive oxygen species transiently decrease auxin signaling and cause stress-induced morphogenic res- ponse in Arabidopsis.Plant Physiol 157, 1866-1883. [本文引用: 2]
[5]
CamilleriC, JouaninL (1991). The TR-DNA region carrying the auxin synthesis genes of the Agrobacterium rhizo- genes agropine-type plasmid pRiA4: nucleotide sequence analysis and introduction into tobacco plants.Mol Plant Microbe Interact 4, 155-162. [本文引用: 1]
[6]
ChenGH, LiuCP, ChenSC, WangLC (2012). Role of ARABIDOPSIS A-FIFTEEN in regulating leaf senescence involves response to reactive oxygen species and is dependent on ETHYLENE INSENSITIVE2.J Exp Bot 63, 275-292. [本文引用: 1]
[7]
ChenJP, BurkeJJ, VeltenJ, XinZU (2006). FtsH11 protease plays a critical role in Arabidopsis thermotolerance.Plant J 48, 73-84. [本文引用: 1]
[8]
EllisCM, NagpalP, YoungJC, HagenG, GuilfoyleTJ, ReedJW (2005). AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 regulate senescence and floral organ abscission in Arabidopsis thaliana.Development 132, 4563-4574. [本文引用: 2]
[9]
Garcia-LorenzoM, SjodinA, JanssonS, FunkC (2006). Protease gene families in Populus and Arabidopsis.BMC Plant Biol 6, 30. [本文引用: 1]
[10]
GazarianIG, LagriminiLM, MellonFA, NaldrettMJ, AshbyGA, ThorneleyRN (1998). Identification of skatolyl hydroperoxide and its role in the peroxidase-catalysed oxidation of indol-3-yl acetic acid.Biochem J 333, 223-232. [本文引用: 1]
[11]
GibalaM, KiciaM, SakamotoW, GolaEM, KubrakiewiczJ, SmakowskaE, JanskaH (2009). The lack of mitochondrial AtFtsH4 protease alters Arabidopsis leaf morphology at the late stage of rosette development under short-day photoperiod.Plant J 59, 685-699. [本文引用: 1]
[12]
GuoY, GanSS (2012). Convergence and divergence in gene expression profiles induced by leaf senescence and 27 senescence-promoting hormonal, pathological and environmental stress treatments.Plant Cell Environ 35, 644-655. [本文引用: 1]
[13]
HeJ, DuanY, HuaD, FanG, WangL, LiuY, ChenZ, HanL, QuLJ, GongZ (2012). DEXH box RNA helicase- mediated mitochondrial reactive oxygen species production in Arabidopsis mediates crosstalk between abscisic acid and auxin signaling.Plant Cell 24, 1815-1833. [本文引用: 2]
[14]
HouK, WuW, GanSS (2013). SAUR36, a small auxin up RNA gene, is involved in the promotion of leaf senescence in Arabidopsis.Plant Physiol 161, 1002-1009. [本文引用: 1]
[15]
HuangYC, ChangYL, HsuJJ, ChuangHW (2008). Trans- criptome analysis of auxin-regulated genes of Arabidopsis thaliana.Gene 420, 118-124. [本文引用: 1]
[16]
JibranR, HunterDA, DijkwelPP (2013). Hormonal regulation of leaf senescence through integration of developmental and stress signals.Plant Mol Biol 82, 547-561. [本文引用: 1]
[17]
JooJH, BaeYS, LeeJS (2001). Role of auxin-induced reactive oxygen species in root gravitropism.Plant Physiol 126, 1055-1060. [本文引用: 1]
[18]
KantS, BiYM, ZhuT, RothsteinSJ (2009). SAUR39, a small auxin-up RNA gene, acts as a negative regulator of auxin synthesis and transport in rice.Plant Physiol 151, 691-701. [本文引用: 1]
[19]
KatoY, MiuraE, IdoK, IfukuK, SakamotoW (2009). The variegated mutants lacking chloroplastic FtsHs are defective in D1 degradation and accumulate reactive oxygen sp- ecies.Plant Physiol 151, 1790-1801. [本文引用: 1]
LimPO, LeeIC, KimJ, KimHJ, RyuJS, WooHR, NamHG (2010). Auxin response factor 2 (ARF2) plays a major role in regulating auxin-mediated leaf longevity.J Exp Bot 61, 1419-1430. [本文引用: 1]
[26]
LjungK, HullAK, KowalczykM, MarchantA, CelenzaJ, CohenJD, SandbergG (2002). Biosynthesis, conjugation, catabolism and homeostasis of indole-3-acetic acid in Arabidopsis thaliana. Plant Mol Biol 49, 249-272. [本文引用: 1]
[27]
MalnoeA, WangF, Girard-BascouJ, WollmanFA, de VitryC (2014). Thylakoid FtsH protease contributes to photosystem II and cytochrome b6f remodeling in Chlamydomonas reinhardtii under stress conditions.Plant Cell 26, 373-390. [本文引用: 1]
[28]
MeudtWJ, GainesTP (1967). Studies on the oxidation of indole-3-acetic acid by peroxidase enzymes. I. Colorimetric determination of indole-3-acetic acid oxidation produ- cts.Plant Physiol 144, 118-128. [本文引用: 1]
[29]
MittlerR, VanderauweraS, SuzukiN, MillerG, TognettiVB, VandepoeleK, GolleryM, ShulaevV, Van Breu- segemF (2011). ROS signaling: the new wave?Trends Plant Sci 16, 300-309. [本文引用: 1]
[30]
NakagamiH, SoukupovaH, SchikoraA, ZarskyV, HirtH (2006). A mitogen-activated protein kinase kinase kinase mediates reactive oxygen species homeostasis in Arabidopsis.J Biol Chem 281, 38697-38704. [本文引用: 1]
[31]
NoldenM, EhsesS, KoppenM, BernacchiaA, RugarliEI, LangerT (2005). The m-AAA protease defective in hereditary spastic paraplegia controls ribosome assembly in mitochondria.Cell 123, 277-289. [本文引用: 1]
[32]
OkushimaY, FukakiH, OnodaM, TheologisA, TasakaM (2007). ARF7 and ARF19 regulate lateral root formation via direct activation ofLBD/ASL genes in Arabidopsis. Pla- nt Cell 19, 118-130. [本文引用: 1]
[33]
PiechotaJ, KolodziejczakM, JuszczakI, SakamotoW, JanskaH (2010). Identification and characterization of high molecular weight complexes formed by matrix AAA proteases and prohibitins in mitochondria ofArabidopsis thaliana. J Biol Chem 285, 12512-12521. [本文引用: 1]
[34]
PottersG, PasternakTP, GuisezY, PalmeKJ, JansenMA (2007). Stress-induced morphogenic responses: growing out of trouble?Trends Plant Sci 12, 98-105. [本文引用: 1]
[35]
QuevalG, Issakidis-BourguetE, HoeberichtsFA, VandorpeM, GakiereB, VanackerH, Miginiac-MaslowM, Van BreusegemF, NoctorG (2007). Conditional oxidative stress responses in the Arabidopsis photorespiratory mutant cat2 demonstrate that redox state is a key modulator of daylength-dependent gene expression, and define photoperiod as a crucial factor in the regulation of H2O2- induced cell death.Plant J 52, 640-657. [本文引用: 1]
[36]
RenG, ZhouQ, WuS, ZhangY, ZhangL, HuangJ, SunZ, KuaiB (2010). Reverse genetic identification of CRN1 and its distinctive role in chlorophyll degradation in Arabidopsis.J Integr Plant Biol 52, 496-504. [本文引用: 1]
[37]
RomanoCP, RobsonPR, SmithH, EstelleM, KleeH (1995). Transgene-mediated auxin overproduction in Ara- bidopsis: hypocotyl elongation phenotype and interactions with the hy6-1 hypocotyl elongation and axr1 auxin-resis- tant mutants.Plant Mol Biol 27, 1071-1083. [本文引用: 1]
[38]
SakamotoW, TamuraT, Hanba-TomitaY, MurataM (2002). The VAR1 locus of Arabidopsis encodes a chloroplastic FtsH and is responsible for leaf variegation in the mutant alleles.Genes Cells 7, 769-780. [本文引用: 1]
[39]
SallehFM, EvansK, GoodallB, MachinH, MowlaSB, MurLA, RunionsJ, TheodoulouFL, FoyerCH, RogersHJ (2012). A novel function for a redox-related LEA protein (SAG21/AtLEA5) in root development and biotic stress re- sponses.Plant Cell Environ 35, 418-429. [本文引用: 1]
[40]
SavitskyPA, GazaryanIG, TishkovVI, LagriminiLM, RuzgasT, GortonL (1999). Oxidation of indole-3-acetic acid by dioxygen catalysed by plant peroxidases: specifi- city for the enzyme structure.Biochem J 340, 579-583. [本文引用: 1]
[41]
SierlaM, RahikainenM, SalojarviJ, KangasjarviJ, KangasjarviS (2013). Apoplastic and chloroplastic redox signaling networks in plant stress responses.Antioxid Re- dox Signal 18, 2220-2239. [本文引用: 1]
WagnerR, AignerH, PruzinskaA, JankanpaaHJ, JanssonS, FunkC (2011). Fitness analyses of Arabidopsis thaliana mutants depleted of FtsH metalloproteases and characterization of three FtsH6 deletion mutants exposed to high light stress, senescence and chilling. New Phytol 191, 449-458. [本文引用: 1]
[44]
WangP, SongCP (2008). Guard-cell signaling for hydrogen peroxide and abscisic acid.New Phytol 178, 703-718. [本文引用: 1]
[45]
WatanabeM, BalazadehS, TohgeT, ErbanA, GiavaliscoP, KopkaJ, Mueller-RoeberB, FernieAR, HoefgenR (2013). Comprehensive dissection of spatiotemporal meta- bolic shifts in primary, secondary, and lipid metabolism during developmental senescence in Arabidopsis.Plant Physiol 162, 1290-1310. [本文引用: 1]
ZimmermannP, HeinleinC, OrendiG, ZentgrafU (2006). Senescence-specific regulation of catalases in Arabidopsis thaliana(L.) Heynh.Plant Cell Environ 29, 1049-1060. [本文引用: 1]
1 2004
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
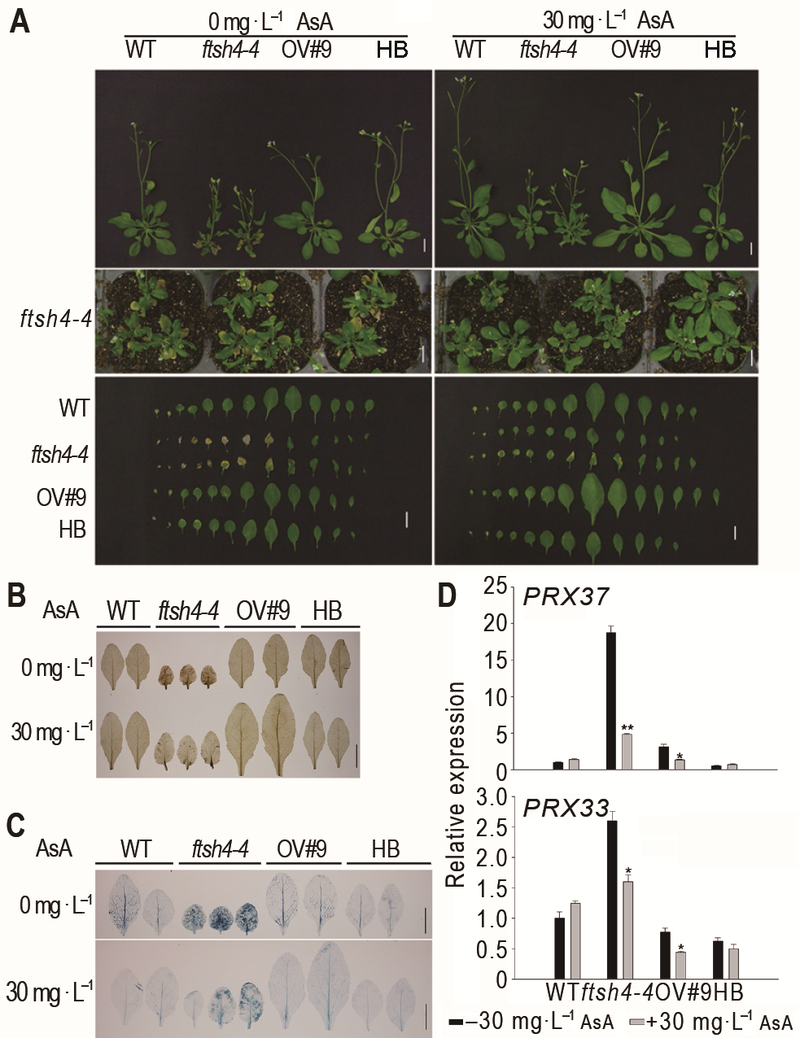
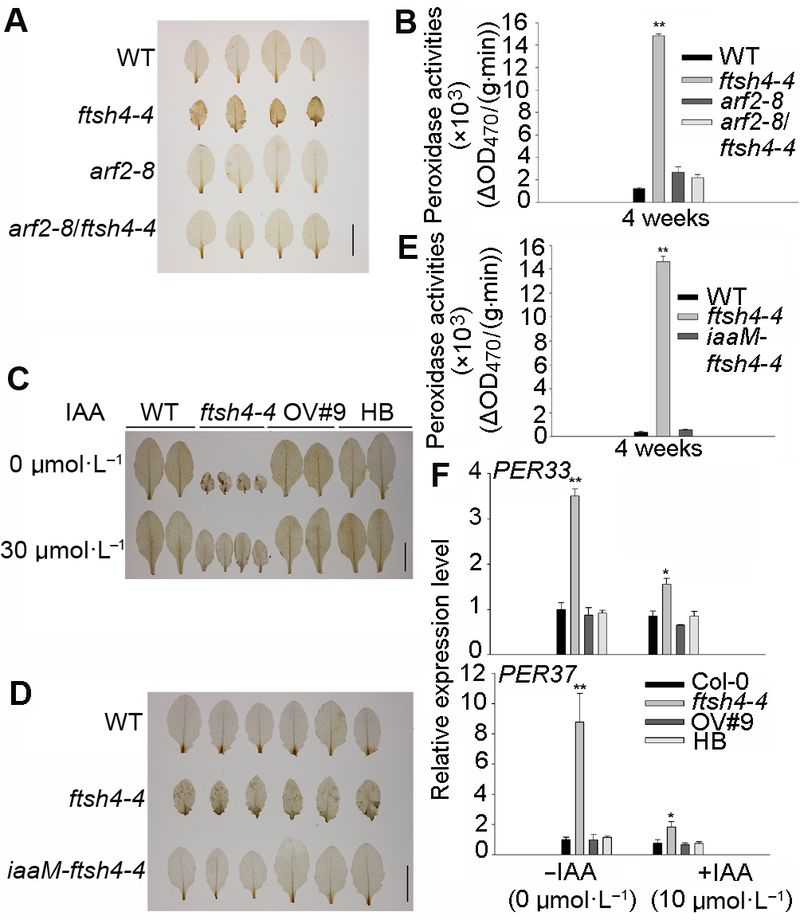
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
2 2011
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ... ... ).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 1991
... 为了进一步确定ftsh4-4突变体表现出的早衰表型是由IAA含量减少引起的, 我们构建了iaaM-ftsh4-4转基因植株(Zhang et al., 2014).iaaM编码色氨酸单加氧酶(IAAM), IAAM是生长素合成途径吲哚乙酰胺途径中的关键酶, 能催化色氨酸转化为吲哚乙酰胺(IAM), 随后IAM再通过吲哚乙酰胺水解酶(IAAH)水解成IAA (Camilleri and Jouanin, 1991; Romano et al., 1995).与ftsh4-4突变体相比, iaaM-ftsh4-4转基因植株中内源IAA含量增加(Zhang et al., 2014).iaaM-ftsh4-4能恢复ftsh4-4突变体的叶片早衰表型, 表现出与WT一致的表型(图4A).对4周龄拟南芥植株莲座叶进行乳酚-台盼蓝染色, 结果显示, 与ftsh4-4突变体相比, iaaM-ftsh4-4转基因植物细胞死亡数目明显减少(图4B), 并且iaaM-ftsh4-4植株的叶绿素 含量(图4C)及相对电导率(图4D)等衰老生理指标均 恢复到与野生型相同的正常水平.我们还利用qRT- PCR检测了生长4周的WT、ftsh4-4和iaaM-ftsh4-4植株莲座叶中衰老相关标记基因SAG12、SAG13、SAG101、SEN1及SEN4的表达水平, 发现与WT相比, ftsh4-4突变体植株莲座叶中SAG12、SAG13、SEN1及SEN4表达量分别增加了440倍、4.2倍、3.7倍和1.3倍, 而iaaM-ftsh4-4能明显将ftsh4-4突变体中SAG12、SAG13、SEN1及SEN4的表达水平降低到WT水平(图4E).以上结果表明, 增加内源生长素含量能恢复ftsh4-4突变体的早衰表型, 即ftsh4-4突变体表现出的早衰表型确实是由于IAA含量减少引起的. ...
1 2012
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
1 2006
... 目前, 关于拟南芥FtSH蛋白酶功能的研究主要集中在定位于叶绿体的FtSH蛋白, 而线粒体定位的FtSH蛋白酶的具体功能及其分子机制还不清楚.FtSH1通过清除D1片段调节光系统II的降解(Malnoe et al., 2014).FtSH2和FtSH5参与类囊体蛋白质的降解及质体发育过程, 2个基因的功能缺失会引起光保护机制的破坏和内囊体的发育异常, 并导致叶斑表型(Saka- moto et al., 2002).FtSH8和FtSH1分别与FtSH2和FtSH5存在功能冗余, 且FtSH2/FtSH8以及FtSH5/ FtSH1形成的复合体参与了光系统II反应中心蛋白D1的降解(Kato et al., 2009).2个不同的ftsh11突变体均表现出热敏感, 表明FtSH11可能在应对高温胁迫中发挥重要作用(Chen et al., 2006).相比其它类型的FtSH蛋白, FtSH2、3、5、10、11在植物生长发育及对环境的适应性中表现出更为重要的作用(Wagner et al., 2011).线粒体定位的FtSH3和FtSH10通过影响氧化磷酸化途径中相关复合体的活性来调控植物的正常发育(Piechota et al., 2010).在短日照条件下, 线粒体定位的FtSH4蛋白酶缺失突变体ftsh4-1中ROS (H2O2)含量增加, 造成拟南芥莲座叶形态发生改变(Gibala et al., 2009), 并且FtSH4能够通过影响过氧化物酶的积累来调控生长素的体内平衡, 进而调控拟南芥的株型(Zhang et al., 2014). ...
2 2005
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ... ... 本研究表明, FtSH4基因插入突变体ftsh4-4在长日照条件下具有早衰表型, 且其叶片早衰表型能够被外源生长素所抑制, 提示FtSH4可能通过生长素正调控植物叶片衰老过程.生长素在调控植物叶片衰老中的功能一直存在争议.有研究认为, 生长素能促进植物叶片衰老.例如, 大豆(Glycine max)中衰老相关受体类激酶(GmSARK)的表达受到生长素的诱导, Gm- SARK在拟南芥中过量表达能够诱导植物叶片早衰, 生长素缺失突变体能延缓GmSARK过量表达植株的早衰表型(Xu et al., 2011); 拟南芥中受生长素诱导表达的基因SAUR36能够促进植物叶片衰老, SAUR- 36在衰老叶片中表达明显升高, 其功能缺失突变体表现出衰老延缓表型, 而过量表达植株则表现出早衰表型(Hou et al., 2013).另有研究则认为, 生长素能够抑制植物叶片衰老.例如, 拟南芥黄素单氧合酶基因YUCCA6过量表达植株35S:YUC6和激活突变体yuc6-1D植株中自由生长素含量增加, 能够延缓黑暗诱导的叶片衰老, 降低衰老相关标记基因SAG12的表达(Kim et al., 2011); SAUR39过量表达能导致植物体内生长素含量减少并且表现出早衰表型(Kant et al., 2009); 拟南芥生长素响应因子ARF2缺失突变体的莲座叶表现出衰老延缓表型(Ellis et al., 2005; Lim et al., 2010).此外, 生长素响应因子基因ARF1、ARF7以及ARF19也可促进叶片衰老(Li et al., 2012).本研究表明, 在ftsh4-4突变体中突变ARF2后能够恢复ftsh4-4的早衰表型, 表明ftsh4-4的叶片早衰表型也是由于高水平表达ARF2造成的. ...
1 2006
... 植物中FtSH (filamentation temperature sensitive H)等ATP依赖的金属蛋白酶在植物生长发育过程中, 尤其是维持叶绿体和线粒体膜蛋白的完整性方面起重要作用(Nolden et al., 2005).拟南芥基因组共编码12个FtSH蛋白(Garcia-Lorenzo et al., 2006), 其中有8个定位在叶绿体(FtSH1、2、5-9和12), 3个定位在线粒体中(FtSH3、4和10), FtSH11同时存在于叶绿体和线粒体中(Kolodziejczak et al., 2002).我们前期的研究表明, 线粒体FtSH4基因突变会导致H2O2大量积累(Zhang et al., 2014), 叶片出现早衰表型(Zhang et al., 2017).本研究表明, H2O2清除剂和生长素能延缓ftsh4突变体的早衰表型, 提示该基因可能在H2O2与生长素调控叶片衰老过程中发挥重要作用.本研究通过细胞生物学和遗传学等方法, 在前期研究的基础上, 进一步对线粒体FtSH4通过生长素与H2O2调控叶片衰老的分子机制进行了深入研究. ...
1 1998
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2009
... 目前, 关于拟南芥FtSH蛋白酶功能的研究主要集中在定位于叶绿体的FtSH蛋白, 而线粒体定位的FtSH蛋白酶的具体功能及其分子机制还不清楚.FtSH1通过清除D1片段调节光系统II的降解(Malnoe et al., 2014).FtSH2和FtSH5参与类囊体蛋白质的降解及质体发育过程, 2个基因的功能缺失会引起光保护机制的破坏和内囊体的发育异常, 并导致叶斑表型(Saka- moto et al., 2002).FtSH8和FtSH1分别与FtSH2和FtSH5存在功能冗余, 且FtSH2/FtSH8以及FtSH5/ FtSH1形成的复合体参与了光系统II反应中心蛋白D1的降解(Kato et al., 2009).2个不同的ftsh11突变体均表现出热敏感, 表明FtSH11可能在应对高温胁迫中发挥重要作用(Chen et al., 2006).相比其它类型的FtSH蛋白, FtSH2、3、5、10、11在植物生长发育及对环境的适应性中表现出更为重要的作用(Wagner et al., 2011).线粒体定位的FtSH3和FtSH10通过影响氧化磷酸化途径中相关复合体的活性来调控植物的正常发育(Piechota et al., 2010).在短日照条件下, 线粒体定位的FtSH4蛋白酶缺失突变体ftsh4-1中ROS (H2O2)含量增加, 造成拟南芥莲座叶形态发生改变(Gibala et al., 2009), 并且FtSH4能够通过影响过氧化物酶的积累来调控生长素的体内平衡, 进而调控拟南芥的株型(Zhang et al., 2014). ...
1 2012
... 植物叶片衰老是一种依赖年龄的细胞程序性退化和降解过程, 能够引起植物体内的代谢紊乱并最终导致植物器官或整株死亡(Lim et al., 2007).植物叶片在衰老过程中, 由于细胞质膜和液泡膜以及叶绿体等细胞器发生瓦解, 破坏了细胞的内稳态, 最终导致衰老叶片细胞死亡.叶片衰老相关代谢变化包括叶绿素降解、叶片黄化, 还有蛋白质、脂质、核酸以及其它生物大分子的水解(Ren et al., 2010; Watanabe et al., 2013).叶片衰老以依赖年龄的方式进行, 属于程序性细胞死亡的一种, 受到各种外部环境因素和内部因素的诱导(Guo and Gan, 2012).影响植物叶片衰老的内部因素包括各种植物激素、植物体内的氧化还原状态以及依赖年龄的内源发育信号(Lim et al., 2007). ...
2 2012
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ... ... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2013
... 本研究表明, FtSH4基因插入突变体ftsh4-4在长日照条件下具有早衰表型, 且其叶片早衰表型能够被外源生长素所抑制, 提示FtSH4可能通过生长素正调控植物叶片衰老过程.生长素在调控植物叶片衰老中的功能一直存在争议.有研究认为, 生长素能促进植物叶片衰老.例如, 大豆(Glycine max)中衰老相关受体类激酶(GmSARK)的表达受到生长素的诱导, Gm- SARK在拟南芥中过量表达能够诱导植物叶片早衰, 生长素缺失突变体能延缓GmSARK过量表达植株的早衰表型(Xu et al., 2011); 拟南芥中受生长素诱导表达的基因SAUR36能够促进植物叶片衰老, SAUR- 36在衰老叶片中表达明显升高, 其功能缺失突变体表现出衰老延缓表型, 而过量表达植株则表现出早衰表型(Hou et al., 2013).另有研究则认为, 生长素能够抑制植物叶片衰老.例如, 拟南芥黄素单氧合酶基因YUCCA6过量表达植株35S:YUC6和激活突变体yuc6-1D植株中自由生长素含量增加, 能够延缓黑暗诱导的叶片衰老, 降低衰老相关标记基因SAG12的表达(Kim et al., 2011); SAUR39过量表达能导致植物体内生长素含量减少并且表现出早衰表型(Kant et al., 2009); 拟南芥生长素响应因子ARF2缺失突变体的莲座叶表现出衰老延缓表型(Ellis et al., 2005; Lim et al., 2010).此外, 生长素响应因子基因ARF1、ARF7以及ARF19也可促进叶片衰老(Li et al., 2012).本研究表明, 在ftsh4-4突变体中突变ARF2后能够恢复ftsh4-4的早衰表型, 表明ftsh4-4的叶片早衰表型也是由于高水平表达ARF2造成的. ...
1 2008
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2013
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
1 2001
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2009
... 本研究表明, FtSH4基因插入突变体ftsh4-4在长日照条件下具有早衰表型, 且其叶片早衰表型能够被外源生长素所抑制, 提示FtSH4可能通过生长素正调控植物叶片衰老过程.生长素在调控植物叶片衰老中的功能一直存在争议.有研究认为, 生长素能促进植物叶片衰老.例如, 大豆(Glycine max)中衰老相关受体类激酶(GmSARK)的表达受到生长素的诱导, Gm- SARK在拟南芥中过量表达能够诱导植物叶片早衰, 生长素缺失突变体能延缓GmSARK过量表达植株的早衰表型(Xu et al., 2011); 拟南芥中受生长素诱导表达的基因SAUR36能够促进植物叶片衰老, SAUR- 36在衰老叶片中表达明显升高, 其功能缺失突变体表现出衰老延缓表型, 而过量表达植株则表现出早衰表型(Hou et al., 2013).另有研究则认为, 生长素能够抑制植物叶片衰老.例如, 拟南芥黄素单氧合酶基因YUCCA6过量表达植株35S:YUC6和激活突变体yuc6-1D植株中自由生长素含量增加, 能够延缓黑暗诱导的叶片衰老, 降低衰老相关标记基因SAG12的表达(Kim et al., 2011); SAUR39过量表达能导致植物体内生长素含量减少并且表现出早衰表型(Kant et al., 2009); 拟南芥生长素响应因子ARF2缺失突变体的莲座叶表现出衰老延缓表型(Ellis et al., 2005; Lim et al., 2010).此外, 生长素响应因子基因ARF1、ARF7以及ARF19也可促进叶片衰老(Li et al., 2012).本研究表明, 在ftsh4-4突变体中突变ARF2后能够恢复ftsh4-4的早衰表型, 表明ftsh4-4的叶片早衰表型也是由于高水平表达ARF2造成的. ...
1 2009
... 目前, 关于拟南芥FtSH蛋白酶功能的研究主要集中在定位于叶绿体的FtSH蛋白, 而线粒体定位的FtSH蛋白酶的具体功能及其分子机制还不清楚.FtSH1通过清除D1片段调节光系统II的降解(Malnoe et al., 2014).FtSH2和FtSH5参与类囊体蛋白质的降解及质体发育过程, 2个基因的功能缺失会引起光保护机制的破坏和内囊体的发育异常, 并导致叶斑表型(Saka- moto et al., 2002).FtSH8和FtSH1分别与FtSH2和FtSH5存在功能冗余, 且FtSH2/FtSH8以及FtSH5/ FtSH1形成的复合体参与了光系统II反应中心蛋白D1的降解(Kato et al., 2009).2个不同的ftsh11突变体均表现出热敏感, 表明FtSH11可能在应对高温胁迫中发挥重要作用(Chen et al., 2006).相比其它类型的FtSH蛋白, FtSH2、3、5、10、11在植物生长发育及对环境的适应性中表现出更为重要的作用(Wagner et al., 2011).线粒体定位的FtSH3和FtSH10通过影响氧化磷酸化途径中相关复合体的活性来调控植物的正常发育(Piechota et al., 2010).在短日照条件下, 线粒体定位的FtSH4蛋白酶缺失突变体ftsh4-1中ROS (H2O2)含量增加, 造成拟南芥莲座叶形态发生改变(Gibala et al., 2009), 并且FtSH4能够通过影响过氧化物酶的积累来调控生长素的体内平衡, 进而调控拟南芥的株型(Zhang et al., 2014). ...
1 2011
... 本研究表明, FtSH4基因插入突变体ftsh4-4在长日照条件下具有早衰表型, 且其叶片早衰表型能够被外源生长素所抑制, 提示FtSH4可能通过生长素正调控植物叶片衰老过程.生长素在调控植物叶片衰老中的功能一直存在争议.有研究认为, 生长素能促进植物叶片衰老.例如, 大豆(Glycine max)中衰老相关受体类激酶(GmSARK)的表达受到生长素的诱导, Gm- SARK在拟南芥中过量表达能够诱导植物叶片早衰, 生长素缺失突变体能延缓GmSARK过量表达植株的早衰表型(Xu et al., 2011); 拟南芥中受生长素诱导表达的基因SAUR36能够促进植物叶片衰老, SAUR- 36在衰老叶片中表达明显升高, 其功能缺失突变体表现出衰老延缓表型, 而过量表达植株则表现出早衰表型(Hou et al., 2013).另有研究则认为, 生长素能够抑制植物叶片衰老.例如, 拟南芥黄素单氧合酶基因YUCCA6过量表达植株35S:YUC6和激活突变体yuc6-1D植株中自由生长素含量增加, 能够延缓黑暗诱导的叶片衰老, 降低衰老相关标记基因SAG12的表达(Kim et al., 2011); SAUR39过量表达能导致植物体内生长素含量减少并且表现出早衰表型(Kant et al., 2009); 拟南芥生长素响应因子ARF2缺失突变体的莲座叶表现出衰老延缓表型(Ellis et al., 2005; Lim et al., 2010).此外, 生长素响应因子基因ARF1、ARF7以及ARF19也可促进叶片衰老(Li et al., 2012).本研究表明, 在ftsh4-4突变体中突变ARF2后能够恢复ftsh4-4的早衰表型, 表明ftsh4-4的叶片早衰表型也是由于高水平表达ARF2造成的. ...
1 2002
... 植物中FtSH (filamentation temperature sensitive H)等ATP依赖的金属蛋白酶在植物生长发育过程中, 尤其是维持叶绿体和线粒体膜蛋白的完整性方面起重要作用(Nolden et al., 2005).拟南芥基因组共编码12个FtSH蛋白(Garcia-Lorenzo et al., 2006), 其中有8个定位在叶绿体(FtSH1、2、5-9和12), 3个定位在线粒体中(FtSH3、4和10), FtSH11同时存在于叶绿体和线粒体中(Kolodziejczak et al., 2002).我们前期的研究表明, 线粒体FtSH4基因突变会导致H2O2大量积累(Zhang et al., 2014), 叶片出现早衰表型(Zhang et al., 2017).本研究表明, H2O2清除剂和生长素能延缓ftsh4突变体的早衰表型, 提示该基因可能在H2O2与生长素调控叶片衰老过程中发挥重要作用.本研究通过细胞生物学和遗传学等方法, 在前期研究的基础上, 进一步对线粒体FtSH4通过生长素与H2O2调控叶片衰老的分子机制进行了深入研究. ...
1 2000
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
2 2012
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ... ... 本研究表明, FtSH4基因插入突变体ftsh4-4在长日照条件下具有早衰表型, 且其叶片早衰表型能够被外源生长素所抑制, 提示FtSH4可能通过生长素正调控植物叶片衰老过程.生长素在调控植物叶片衰老中的功能一直存在争议.有研究认为, 生长素能促进植物叶片衰老.例如, 大豆(Glycine max)中衰老相关受体类激酶(GmSARK)的表达受到生长素的诱导, Gm- SARK在拟南芥中过量表达能够诱导植物叶片早衰, 生长素缺失突变体能延缓GmSARK过量表达植株的早衰表型(Xu et al., 2011); 拟南芥中受生长素诱导表达的基因SAUR36能够促进植物叶片衰老, SAUR- 36在衰老叶片中表达明显升高, 其功能缺失突变体表现出衰老延缓表型, 而过量表达植株则表现出早衰表型(Hou et al., 2013).另有研究则认为, 生长素能够抑制植物叶片衰老.例如, 拟南芥黄素单氧合酶基因YUCCA6过量表达植株35S:YUC6和激活突变体yuc6-1D植株中自由生长素含量增加, 能够延缓黑暗诱导的叶片衰老, 降低衰老相关标记基因SAG12的表达(Kim et al., 2011); SAUR39过量表达能导致植物体内生长素含量减少并且表现出早衰表型(Kant et al., 2009); 拟南芥生长素响应因子ARF2缺失突变体的莲座叶表现出衰老延缓表型(Ellis et al., 2005; Lim et al., 2010).此外, 生长素响应因子基因ARF1、ARF7以及ARF19也可促进叶片衰老(Li et al., 2012).本研究表明, 在ftsh4-4突变体中突变ARF2后能够恢复ftsh4-4的早衰表型, 表明ftsh4-4的叶片早衰表型也是由于高水平表达ARF2造成的. ...
2 2007
... 植物叶片衰老是一种依赖年龄的细胞程序性退化和降解过程, 能够引起植物体内的代谢紊乱并最终导致植物器官或整株死亡(Lim et al., 2007).植物叶片在衰老过程中, 由于细胞质膜和液泡膜以及叶绿体等细胞器发生瓦解, 破坏了细胞的内稳态, 最终导致衰老叶片细胞死亡.叶片衰老相关代谢变化包括叶绿素降解、叶片黄化, 还有蛋白质、脂质、核酸以及其它生物大分子的水解(Ren et al., 2010; Watanabe et al., 2013).叶片衰老以依赖年龄的方式进行, 属于程序性细胞死亡的一种, 受到各种外部环境因素和内部因素的诱导(Guo and Gan, 2012).影响植物叶片衰老的内部因素包括各种植物激素、植物体内的氧化还原状态以及依赖年龄的内源发育信号(Lim et al., 2007). ... ... ).影响植物叶片衰老的内部因素包括各种植物激素、植物体内的氧化还原状态以及依赖年龄的内源发育信号(Lim et al., 2007). ...
1 2010
... 本研究表明, FtSH4基因插入突变体ftsh4-4在长日照条件下具有早衰表型, 且其叶片早衰表型能够被外源生长素所抑制, 提示FtSH4可能通过生长素正调控植物叶片衰老过程.生长素在调控植物叶片衰老中的功能一直存在争议.有研究认为, 生长素能促进植物叶片衰老.例如, 大豆(Glycine max)中衰老相关受体类激酶(GmSARK)的表达受到生长素的诱导, Gm- SARK在拟南芥中过量表达能够诱导植物叶片早衰, 生长素缺失突变体能延缓GmSARK过量表达植株的早衰表型(Xu et al., 2011); 拟南芥中受生长素诱导表达的基因SAUR36能够促进植物叶片衰老, SAUR- 36在衰老叶片中表达明显升高, 其功能缺失突变体表现出衰老延缓表型, 而过量表达植株则表现出早衰表型(Hou et al., 2013).另有研究则认为, 生长素能够抑制植物叶片衰老.例如, 拟南芥黄素单氧合酶基因YUCCA6过量表达植株35S:YUC6和激活突变体yuc6-1D植株中自由生长素含量增加, 能够延缓黑暗诱导的叶片衰老, 降低衰老相关标记基因SAG12的表达(Kim et al., 2011); SAUR39过量表达能导致植物体内生长素含量减少并且表现出早衰表型(Kant et al., 2009); 拟南芥生长素响应因子ARF2缺失突变体的莲座叶表现出衰老延缓表型(Ellis et al., 2005; Lim et al., 2010).此外, 生长素响应因子基因ARF1、ARF7以及ARF19也可促进叶片衰老(Li et al., 2012).本研究表明, 在ftsh4-4突变体中突变ARF2后能够恢复ftsh4-4的早衰表型, 表明ftsh4-4的叶片早衰表型也是由于高水平表达ARF2造成的. ...
1 2002
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2014
... 目前, 关于拟南芥FtSH蛋白酶功能的研究主要集中在定位于叶绿体的FtSH蛋白, 而线粒体定位的FtSH蛋白酶的具体功能及其分子机制还不清楚.FtSH1通过清除D1片段调节光系统II的降解(Malnoe et al., 2014).FtSH2和FtSH5参与类囊体蛋白质的降解及质体发育过程, 2个基因的功能缺失会引起光保护机制的破坏和内囊体的发育异常, 并导致叶斑表型(Saka- moto et al., 2002).FtSH8和FtSH1分别与FtSH2和FtSH5存在功能冗余, 且FtSH2/FtSH8以及FtSH5/ FtSH1形成的复合体参与了光系统II反应中心蛋白D1的降解(Kato et al., 2009).2个不同的ftsh11突变体均表现出热敏感, 表明FtSH11可能在应对高温胁迫中发挥重要作用(Chen et al., 2006).相比其它类型的FtSH蛋白, FtSH2、3、5、10、11在植物生长发育及对环境的适应性中表现出更为重要的作用(Wagner et al., 2011).线粒体定位的FtSH3和FtSH10通过影响氧化磷酸化途径中相关复合体的活性来调控植物的正常发育(Piechota et al., 2010).在短日照条件下, 线粒体定位的FtSH4蛋白酶缺失突变体ftsh4-1中ROS (H2O2)含量增加, 造成拟南芥莲座叶形态发生改变(Gibala et al., 2009), 并且FtSH4能够通过影响过氧化物酶的积累来调控生长素的体内平衡, 进而调控拟南芥的株型(Zhang et al., 2014). ...
1 1967
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2011
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
1 2006
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2005
... 植物中FtSH (filamentation temperature sensitive H)等ATP依赖的金属蛋白酶在植物生长发育过程中, 尤其是维持叶绿体和线粒体膜蛋白的完整性方面起重要作用(Nolden et al., 2005).拟南芥基因组共编码12个FtSH蛋白(Garcia-Lorenzo et al., 2006), 其中有8个定位在叶绿体(FtSH1、2、5-9和12), 3个定位在线粒体中(FtSH3、4和10), FtSH11同时存在于叶绿体和线粒体中(Kolodziejczak et al., 2002).我们前期的研究表明, 线粒体FtSH4基因突变会导致H2O2大量积累(Zhang et al., 2014), 叶片出现早衰表型(Zhang et al., 2017).本研究表明, H2O2清除剂和生长素能延缓ftsh4突变体的早衰表型, 提示该基因可能在H2O2与生长素调控叶片衰老过程中发挥重要作用.本研究通过细胞生物学和遗传学等方法, 在前期研究的基础上, 进一步对线粒体FtSH4通过生长素与H2O2调控叶片衰老的分子机制进行了深入研究. ...
1 2007
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
1 2010
... 目前, 关于拟南芥FtSH蛋白酶功能的研究主要集中在定位于叶绿体的FtSH蛋白, 而线粒体定位的FtSH蛋白酶的具体功能及其分子机制还不清楚.FtSH1通过清除D1片段调节光系统II的降解(Malnoe et al., 2014).FtSH2和FtSH5参与类囊体蛋白质的降解及质体发育过程, 2个基因的功能缺失会引起光保护机制的破坏和内囊体的发育异常, 并导致叶斑表型(Saka- moto et al., 2002).FtSH8和FtSH1分别与FtSH2和FtSH5存在功能冗余, 且FtSH2/FtSH8以及FtSH5/ FtSH1形成的复合体参与了光系统II反应中心蛋白D1的降解(Kato et al., 2009).2个不同的ftsh11突变体均表现出热敏感, 表明FtSH11可能在应对高温胁迫中发挥重要作用(Chen et al., 2006).相比其它类型的FtSH蛋白, FtSH2、3、5、10、11在植物生长发育及对环境的适应性中表现出更为重要的作用(Wagner et al., 2011).线粒体定位的FtSH3和FtSH10通过影响氧化磷酸化途径中相关复合体的活性来调控植物的正常发育(Piechota et al., 2010).在短日照条件下, 线粒体定位的FtSH4蛋白酶缺失突变体ftsh4-1中ROS (H2O2)含量增加, 造成拟南芥莲座叶形态发生改变(Gibala et al., 2009), 并且FtSH4能够通过影响过氧化物酶的积累来调控生长素的体内平衡, 进而调控拟南芥的株型(Zhang et al., 2014). ...
1 2007
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2007
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
1 2010
... 植物叶片衰老是一种依赖年龄的细胞程序性退化和降解过程, 能够引起植物体内的代谢紊乱并最终导致植物器官或整株死亡(Lim et al., 2007).植物叶片在衰老过程中, 由于细胞质膜和液泡膜以及叶绿体等细胞器发生瓦解, 破坏了细胞的内稳态, 最终导致衰老叶片细胞死亡.叶片衰老相关代谢变化包括叶绿素降解、叶片黄化, 还有蛋白质、脂质、核酸以及其它生物大分子的水解(Ren et al., 2010; Watanabe et al., 2013).叶片衰老以依赖年龄的方式进行, 属于程序性细胞死亡的一种, 受到各种外部环境因素和内部因素的诱导(Guo and Gan, 2012).影响植物叶片衰老的内部因素包括各种植物激素、植物体内的氧化还原状态以及依赖年龄的内源发育信号(Lim et al., 2007). ...
1 1995
... 为了进一步确定ftsh4-4突变体表现出的早衰表型是由IAA含量减少引起的, 我们构建了iaaM-ftsh4-4转基因植株(Zhang et al., 2014).iaaM编码色氨酸单加氧酶(IAAM), IAAM是生长素合成途径吲哚乙酰胺途径中的关键酶, 能催化色氨酸转化为吲哚乙酰胺(IAM), 随后IAM再通过吲哚乙酰胺水解酶(IAAH)水解成IAA (Camilleri and Jouanin, 1991; Romano et al., 1995).与ftsh4-4突变体相比, iaaM-ftsh4-4转基因植株中内源IAA含量增加(Zhang et al., 2014).iaaM-ftsh4-4能恢复ftsh4-4突变体的叶片早衰表型, 表现出与WT一致的表型(图4A).对4周龄拟南芥植株莲座叶进行乳酚-台盼蓝染色, 结果显示, 与ftsh4-4突变体相比, iaaM-ftsh4-4转基因植物细胞死亡数目明显减少(图4B), 并且iaaM-ftsh4-4植株的叶绿素 含量(图4C)及相对电导率(图4D)等衰老生理指标均 恢复到与野生型相同的正常水平.我们还利用qRT- PCR检测了生长4周的WT、ftsh4-4和iaaM-ftsh4-4植株莲座叶中衰老相关标记基因SAG12、SAG13、SAG101、SEN1及SEN4的表达水平, 发现与WT相比, ftsh4-4突变体植株莲座叶中SAG12、SAG13、SEN1及SEN4表达量分别增加了440倍、4.2倍、3.7倍和1.3倍, 而iaaM-ftsh4-4能明显将ftsh4-4突变体中SAG12、SAG13、SEN1及SEN4的表达水平降低到WT水平(图4E).以上结果表明, 增加内源生长素含量能恢复ftsh4-4突变体的早衰表型, 即ftsh4-4突变体表现出的早衰表型确实是由于IAA含量减少引起的. ...
1 2002
... 目前, 关于拟南芥FtSH蛋白酶功能的研究主要集中在定位于叶绿体的FtSH蛋白, 而线粒体定位的FtSH蛋白酶的具体功能及其分子机制还不清楚.FtSH1通过清除D1片段调节光系统II的降解(Malnoe et al., 2014).FtSH2和FtSH5参与类囊体蛋白质的降解及质体发育过程, 2个基因的功能缺失会引起光保护机制的破坏和内囊体的发育异常, 并导致叶斑表型(Saka- moto et al., 2002).FtSH8和FtSH1分别与FtSH2和FtSH5存在功能冗余, 且FtSH2/FtSH8以及FtSH5/ FtSH1形成的复合体参与了光系统II反应中心蛋白D1的降解(Kato et al., 2009).2个不同的ftsh11突变体均表现出热敏感, 表明FtSH11可能在应对高温胁迫中发挥重要作用(Chen et al., 2006).相比其它类型的FtSH蛋白, FtSH2、3、5、10、11在植物生长发育及对环境的适应性中表现出更为重要的作用(Wagner et al., 2011).线粒体定位的FtSH3和FtSH10通过影响氧化磷酸化途径中相关复合体的活性来调控植物的正常发育(Piechota et al., 2010).在短日照条件下, 线粒体定位的FtSH4蛋白酶缺失突变体ftsh4-1中ROS (H2O2)含量增加, 造成拟南芥莲座叶形态发生改变(Gibala et al., 2009), 并且FtSH4能够通过影响过氧化物酶的积累来调控生长素的体内平衡, 进而调控拟南芥的株型(Zhang et al., 2014). ...
1 2012
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
1 1999
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2013
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2011
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2011
... 目前, 关于拟南芥FtSH蛋白酶功能的研究主要集中在定位于叶绿体的FtSH蛋白, 而线粒体定位的FtSH蛋白酶的具体功能及其分子机制还不清楚.FtSH1通过清除D1片段调节光系统II的降解(Malnoe et al., 2014).FtSH2和FtSH5参与类囊体蛋白质的降解及质体发育过程, 2个基因的功能缺失会引起光保护机制的破坏和内囊体的发育异常, 并导致叶斑表型(Saka- moto et al., 2002).FtSH8和FtSH1分别与FtSH2和FtSH5存在功能冗余, 且FtSH2/FtSH8以及FtSH5/ FtSH1形成的复合体参与了光系统II反应中心蛋白D1的降解(Kato et al., 2009).2个不同的ftsh11突变体均表现出热敏感, 表明FtSH11可能在应对高温胁迫中发挥重要作用(Chen et al., 2006).相比其它类型的FtSH蛋白, FtSH2、3、5、10、11在植物生长发育及对环境的适应性中表现出更为重要的作用(Wagner et al., 2011).线粒体定位的FtSH3和FtSH10通过影响氧化磷酸化途径中相关复合体的活性来调控植物的正常发育(Piechota et al., 2010).在短日照条件下, 线粒体定位的FtSH4蛋白酶缺失突变体ftsh4-1中ROS (H2O2)含量增加, 造成拟南芥莲座叶形态发生改变(Gibala et al., 2009), 并且FtSH4能够通过影响过氧化物酶的积累来调控生长素的体内平衡, 进而调控拟南芥的株型(Zhang et al., 2014). ...
1 2008
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
1 2013
... 植物叶片衰老是一种依赖年龄的细胞程序性退化和降解过程, 能够引起植物体内的代谢紊乱并最终导致植物器官或整株死亡(Lim et al., 2007).植物叶片在衰老过程中, 由于细胞质膜和液泡膜以及叶绿体等细胞器发生瓦解, 破坏了细胞的内稳态, 最终导致衰老叶片细胞死亡.叶片衰老相关代谢变化包括叶绿素降解、叶片黄化, 还有蛋白质、脂质、核酸以及其它生物大分子的水解(Ren et al., 2010; Watanabe et al., 2013).叶片衰老以依赖年龄的方式进行, 属于程序性细胞死亡的一种, 受到各种外部环境因素和内部因素的诱导(Guo and Gan, 2012).影响植物叶片衰老的内部因素包括各种植物激素、植物体内的氧化还原状态以及依赖年龄的内源发育信号(Lim et al., 2007). ...
1 2005
... 植物通过各种机制调控体内IAA的平衡, 如生长素的生物合成、降解和转运以及IAA的氧化(Meudt and Gaines, 1967; Woodward and Bartel, 2005).许多研究认为, ROS是植物以及其它生物不同代谢过程中的重要信号分子, 可与植物激素及其它信号分子相互作用, 参与病原体防御、非生物胁迫耐受、气孔调节、发育、生长以及细胞增殖等代谢过程(Blomster et al., 2011; Suzuki et al., 2011; Sierla et al., 2013).生物/非生物胁迫能诱导植物体ROS的产生, 增加的ROS能改变体内生长素平衡, 进而导致植株株型发生变化(Potters et al., 2007).ROS不仅可通过调节参与生长素合成的相关酶活性来影响生长素的体内平衡, 同时也可以通过依赖H2O2产生的过氧化物酶氧化降解生长素(Ljung et al., 2002).植物过氧化物酶具有生长素氧化酶活性, 能够催化IAA与氧分子发生氧化反应, 从而氧化IAA (Savitsky et al., 1999).另外, H2O2也能够氧化植物体内的IAA, 催化其降解, 进而扰乱植物体内生长素的稳态平衡(Gazarian et al., 1998); 同时, H2O2能通过激活拟南芥中特殊的促分裂原活化蛋白激酶ANP1来抑制生长素活性(Ko- vtun et al., 2000); 而且H2O2能够以依赖蛋白酶体的方式调节MAPK级联信号, 降低生长素信号途径相关基因的表达, 从而负调控植物生长素信号途径(Naka- gami et al., 2006).拟南芥ntra/ntrb/cad2三突变体中ROS清除剂硫氧还蛋白和谷胱甘肽的含量降低, ROS含量升高, 内源生长素含量减少且表现出生长素缺陷相关表型(Bashandy et al., 2010).外源ROS处理能短暂地抑制拟南芥中的生长素信号, 且引发胁迫诱导的表型响应(Blomster et al., 2011).拟南芥线粒体产生的ROS能介导ABA和生长素相互作用, 调控主根生长和种子萌发(He et al., 2012).ftsh4-4突变体中有大量的H2O2积累, 过氧化物酶相关基因PRX33及PRX37的表达明显上调且过氧化物酶活性明显增强, 外施抗氧化剂能够抑制ftsh4-4突变体中的早衰表型.结合外源或者内源生长素均能恢复ftsh4-4的衰老表型, 暗示外源抗氧化剂可能是通过降低突变体内的ROS含量及过氧化物酶的活性, 进而恢复ftsh4-4体内的生长素水平并抑制ftsh4-4叶片的衰老.也有研究显示, 植物氧化胁迫相关基因的表达受到生长素的调控(Huang et al., 2008), 生长素能够调节植物体内抗氧化酶的含量(Joo et al., 2001).本研究还表明, 外源生长素处理或者提高内源生长素后不仅恢复了ftsh4-4突变体的表型, 而且降低了ftsh4-4突变体植株中H2O2含量以及过氧化物酶活性, 暗示着生长素在维持植物体内氧化还原平衡中可能也起重要作用.生长素如何调控植物体内ROS的含量是一个非常有趣的科学问题.在ftsh4-4中, 由于ARF2突变能够降低ftsh4-4的H2O2含量, 因此生长素很有可能是通过诸如ARF2等生长素响应因子来调控植物体内的ROS含量. ...
1 2011
... 本研究表明, FtSH4基因插入突变体ftsh4-4在长日照条件下具有早衰表型, 且其叶片早衰表型能够被外源生长素所抑制, 提示FtSH4可能通过生长素正调控植物叶片衰老过程.生长素在调控植物叶片衰老中的功能一直存在争议.有研究认为, 生长素能促进植物叶片衰老.例如, 大豆(Glycine max)中衰老相关受体类激酶(GmSARK)的表达受到生长素的诱导, Gm- SARK在拟南芥中过量表达能够诱导植物叶片早衰, 生长素缺失突变体能延缓GmSARK过量表达植株的早衰表型(Xu et al., 2011); 拟南芥中受生长素诱导表达的基因SAUR36能够促进植物叶片衰老, SAUR- 36在衰老叶片中表达明显升高, 其功能缺失突变体表现出衰老延缓表型, 而过量表达植株则表现出早衰表型(Hou et al., 2013).另有研究则认为, 生长素能够抑制植物叶片衰老.例如, 拟南芥黄素单氧合酶基因YUCCA6过量表达植株35S:YUC6和激活突变体yuc6-1D植株中自由生长素含量增加, 能够延缓黑暗诱导的叶片衰老, 降低衰老相关标记基因SAG12的表达(Kim et al., 2011); SAUR39过量表达能导致植物体内生长素含量减少并且表现出早衰表型(Kant et al., 2009); 拟南芥生长素响应因子ARF2缺失突变体的莲座叶表现出衰老延缓表型(Ellis et al., 2005; Lim et al., 2010).此外, 生长素响应因子基因ARF1、ARF7以及ARF19也可促进叶片衰老(Li et al., 2012).本研究表明, 在ftsh4-4突变体中突变ARF2后能够恢复ftsh4-4的早衰表型, 表明ftsh4-4的叶片早衰表型也是由于高水平表达ARF2造成的. ...
1 2013
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
1 2010
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...
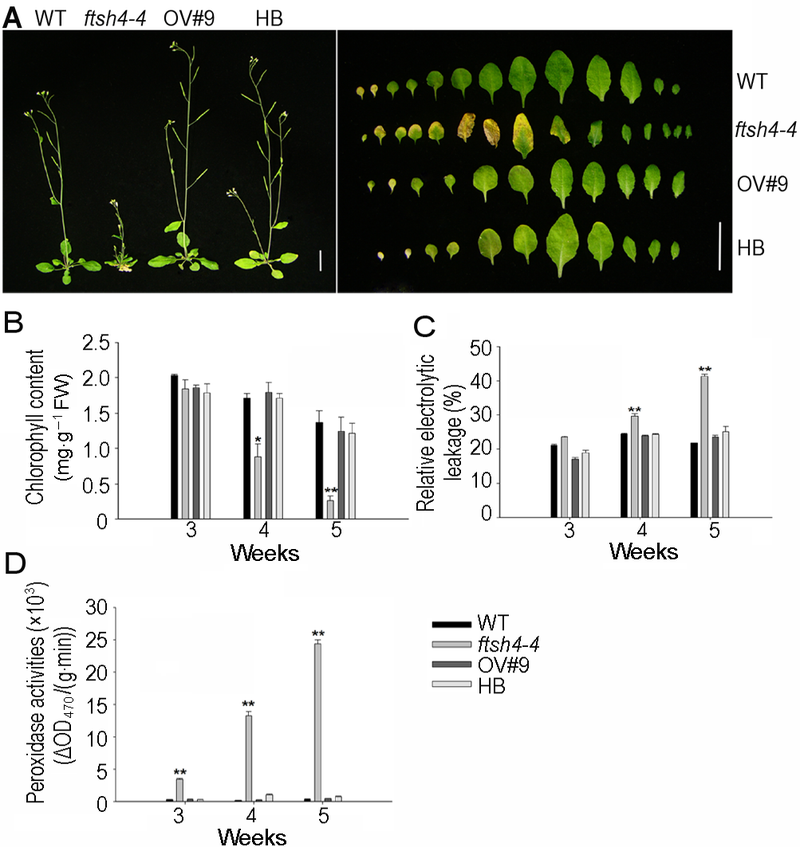
6 2014
... 植物中FtSH (filamentation temperature sensitive H)等ATP依赖的金属蛋白酶在植物生长发育过程中, 尤其是维持叶绿体和线粒体膜蛋白的完整性方面起重要作用(Nolden et al., 2005).拟南芥基因组共编码12个FtSH蛋白(Garcia-Lorenzo et al., 2006), 其中有8个定位在叶绿体(FtSH1、2、5-9和12), 3个定位在线粒体中(FtSH3、4和10), FtSH11同时存在于叶绿体和线粒体中(Kolodziejczak et al., 2002).我们前期的研究表明, 线粒体FtSH4基因突变会导致H2O2大量积累(Zhang et al., 2014), 叶片出现早衰表型(Zhang et al., 2017).本研究表明, H2O2清除剂和生长素能延缓ftsh4突变体的早衰表型, 提示该基因可能在H2O2与生长素调控叶片衰老过程中发挥重要作用.本研究通过细胞生物学和遗传学等方法, 在前期研究的基础上, 进一步对线粒体FtSH4通过生长素与H2O2调控叶片衰老的分子机制进行了深入研究. ... ... 长日照条件下, 生长4周的FtSH4基因T-DNA插入突变体ftsh4-4植株与野生型(WT)相比, 莲座叶表现出明显的早衰表型, 而ftsh4-4突变体互补植株(FtSH4-ftsh4- 4, HB)能够完全恢复其早衰表型, FtSH4过表达植株(OV#9)生长情况与WT基本一致(图1A).长日照条件下不同生长时期(3、4和5周)不同FtSH4基因型植株莲座叶中叶绿素含量以及相对电导率的测定结果显示, 长日照条件下生长4周时, ftsh4-4突变体中叶绿素含量开始明显下降(图1B), 相对电导率明显上升(图1C).前期研究表明, ftsh4-4突变体中H2O2积累明显升高, 外源FtSH4基因能够降低ftsh4-4突变体高含量H2O2的积累, 并且ftsh4-4突变体中过氧化物酶基因PRX33、PRX34和PRX37的表达均明显高于WT (Zhang et al., 2014).为了进一步确定FtSH4与H2O2的关系, 我们检测了长日照条件下不同生长时期(3、4和5周)不同FtSH4基因型植株莲座叶中过氧化物酶的活性.结果显示, ftsh4-4突变体中过氧化物酶活性明显高于WT、OV#9和HB植株; 相比同期WT植株, 3、4和5周龄ftsh4-4突变体中过氧化物酶活性分别增加了约10倍、36倍和62倍(图1D).以上结果表明, FtSH4突变能够诱导H2O2大量积累, 并且可能通过上调过 氧化物酶的活性参与对H2O2的调控, 进而影响叶片衰老. ... ... 前期研究表明, 与野生型相比, ftsh4-4突变体中IAA含量减少(Zhang et al., 2014).为了验证ftsh4-4突变体的早衰表型是否是由植物体内IAA含量减少所引起的, 我们利用外源生长素处理ftsh4-4突变体.结果发现, 外源生长素能明显延缓ftsh4-4突变体的早衰表型(图3A).对IAA处理2周后的莲座叶进行乳酚-台盼蓝染色, 结果显示, 10 µmol∙L-1 IAA处理后ftsh4-4突变体中细胞死亡数目明显比未处理植株少, 而WT、OV#9和HB植株处理前后没有明显差异(图3B).结果表明生长素在FtSH4介导的植物叶片衰老调控中发挥重要作用. ... ... 为了进一步确定ftsh4-4突变体表现出的早衰表型是由IAA含量减少引起的, 我们构建了iaaM-ftsh4-4转基因植株(Zhang et al., 2014).iaaM编码色氨酸单加氧酶(IAAM), IAAM是生长素合成途径吲哚乙酰胺途径中的关键酶, 能催化色氨酸转化为吲哚乙酰胺(IAM), 随后IAM再通过吲哚乙酰胺水解酶(IAAH)水解成IAA (Camilleri and Jouanin, 1991; Romano et al., 1995).与ftsh4-4突变体相比, iaaM-ftsh4-4转基因植株中内源IAA含量增加(Zhang et al., 2014).iaaM-ftsh4-4能恢复ftsh4-4突变体的叶片早衰表型, 表现出与WT一致的表型(图4A).对4周龄拟南芥植株莲座叶进行乳酚-台盼蓝染色, 结果显示, 与ftsh4-4突变体相比, iaaM-ftsh4-4转基因植物细胞死亡数目明显减少(图4B), 并且iaaM-ftsh4-4植株的叶绿素 含量(图4C)及相对电导率(图4D)等衰老生理指标均 恢复到与野生型相同的正常水平.我们还利用qRT- PCR检测了生长4周的WT、ftsh4-4和iaaM-ftsh4-4植株莲座叶中衰老相关标记基因SAG12、SAG13、SAG101、SEN1及SEN4的表达水平, 发现与WT相比, ftsh4-4突变体植株莲座叶中SAG12、SAG13、SEN1及SEN4表达量分别增加了440倍、4.2倍、3.7倍和1.3倍, 而iaaM-ftsh4-4能明显将ftsh4-4突变体中SAG12、SAG13、SEN1及SEN4的表达水平降低到WT水平(图4E).以上结果表明, 增加内源生长素含量能恢复ftsh4-4突变体的早衰表型, 即ftsh4-4突变体表现出的早衰表型确实是由于IAA含量减少引起的. ... ... 转基因植株中内源IAA含量增加(Zhang et al., 2014).iaaM-ftsh4-4能恢复ftsh4-4突变体的叶片早衰表型, 表现出与WT一致的表型(图4A).对4周龄拟南芥植株莲座叶进行乳酚-台盼蓝染色, 结果显示, 与ftsh4-4突变体相比, iaaM-ftsh4-4转基因植物细胞死亡数目明显减少(图4B), 并且iaaM-ftsh4-4植株的叶绿素 含量(图4C)及相对电导率(图4D)等衰老生理指标均 恢复到与野生型相同的正常水平.我们还利用qRT- PCR检测了生长4周的WT、ftsh4-4和iaaM-ftsh4-4植株莲座叶中衰老相关标记基因SAG12、SAG13、SAG101、SEN1及SEN4的表达水平, 发现与WT相比, ftsh4-4突变体植株莲座叶中SAG12、SAG13、SEN1及SEN4表达量分别增加了440倍、4.2倍、3.7倍和1.3倍, 而iaaM-ftsh4-4能明显将ftsh4-4突变体中SAG12、SAG13、SEN1及SEN4的表达水平降低到WT水平(图4E).以上结果表明, 增加内源生长素含量能恢复ftsh4-4突变体的早衰表型, 即ftsh4-4突变体表现出的早衰表型确实是由于IAA含量减少引起的. ... ... 目前, 关于拟南芥FtSH蛋白酶功能的研究主要集中在定位于叶绿体的FtSH蛋白, 而线粒体定位的FtSH蛋白酶的具体功能及其分子机制还不清楚.FtSH1通过清除D1片段调节光系统II的降解(Malnoe et al., 2014).FtSH2和FtSH5参与类囊体蛋白质的降解及质体发育过程, 2个基因的功能缺失会引起光保护机制的破坏和内囊体的发育异常, 并导致叶斑表型(Saka- moto et al., 2002).FtSH8和FtSH1分别与FtSH2和FtSH5存在功能冗余, 且FtSH2/FtSH8以及FtSH5/ FtSH1形成的复合体参与了光系统II反应中心蛋白D1的降解(Kato et al., 2009).2个不同的ftsh11突变体均表现出热敏感, 表明FtSH11可能在应对高温胁迫中发挥重要作用(Chen et al., 2006).相比其它类型的FtSH蛋白, FtSH2、3、5、10、11在植物生长发育及对环境的适应性中表现出更为重要的作用(Wagner et al., 2011).线粒体定位的FtSH3和FtSH10通过影响氧化磷酸化途径中相关复合体的活性来调控植物的正常发育(Piechota et al., 2010).在短日照条件下, 线粒体定位的FtSH4蛋白酶缺失突变体ftsh4-1中ROS (H2O2)含量增加, 造成拟南芥莲座叶形态发生改变(Gibala et al., 2009), 并且FtSH4能够通过影响过氧化物酶的积累来调控生长素的体内平衡, 进而调控拟南芥的株型(Zhang et al., 2014). ...
1 2017
... 植物中FtSH (filamentation temperature sensitive H)等ATP依赖的金属蛋白酶在植物生长发育过程中, 尤其是维持叶绿体和线粒体膜蛋白的完整性方面起重要作用(Nolden et al., 2005).拟南芥基因组共编码12个FtSH蛋白(Garcia-Lorenzo et al., 2006), 其中有8个定位在叶绿体(FtSH1、2、5-9和12), 3个定位在线粒体中(FtSH3、4和10), FtSH11同时存在于叶绿体和线粒体中(Kolodziejczak et al., 2002).我们前期的研究表明, 线粒体FtSH4基因突变会导致H2O2大量积累(Zhang et al., 2014), 叶片出现早衰表型(Zhang et al., 2017).本研究表明, H2O2清除剂和生长素能延缓ftsh4突变体的早衰表型, 提示该基因可能在H2O2与生长素调控叶片衰老过程中发挥重要作用.本研究通过细胞生物学和遗传学等方法, 在前期研究的基础上, 进一步对线粒体FtSH4通过生长素与H2O2调控叶片衰老的分子机制进行了深入研究. ...
1 2006
... 植物激素是叶片衰老的重要调节者, 改变植物体内的激素信号会导致叶片衰老表型发生变化(Li et al., 2012).植物激素可诱导或抑制叶片衰老, 乙烯(ethy- lene)、茉莉酸(JA)、脱落酸(ABA)和水杨酸(SA)可促进叶片衰老; 而生长素(auxin)、赤霉素(GA)和细胞分裂素(CKs)则延缓叶片衰老(Jibran et al., 2013).虽然已知生长素作为调控植物生长发育的重要激素参与叶片衰老调控, 但是相关的分子机制仍不清楚.有限的研究表明, 模式植物拟南芥(Arabidopsis thaliana)中, 生长素响应因子ARF2和ARF7参与植物衰老调控(Ellis et al., 2005; Okushima et al., 2007).活性氧(reactive oxygen species, ROS)的积累是植物细胞应对各种生物和非生物胁迫以及自然衰老过程做出的响应, 自然状态下的衰老及环境胁迫诱导的衰老过程均会产生大量的ROS (Apel and Hirt, 2004).其中, H2O2 (hydrogen peroxide)被认为是调控细胞程序性死亡的信号中介, 衰老叶片中H2O2含量明显增加(Zimmermann et al., 2006; Queval et al., 2007), 许多叶片衰老调控基因(如WRKY53和SAG21)的表达均受H2O2的诱导(Zentgraf et al., 2010; Salleh et al., 2012).ROS信号能够与植物激素信号形成复杂的调控网络, 在调控植物生长发育、衰老以及环境应答等过程中起重要作用(Mittler et al., 2011).如ROS信号与ABA和乙烯等激素相互作用调控正常及逆境胁迫条件下的植物叶片衰老过程(Wang and Song, 2008; Chen et al., 2012); 与生长素相互作用调控植物主根和侧根生长、种子萌发以及金属胁迫响应等过程(He et al., 2012; Yuan et al., 2013).但是ROS如何与生长素相互作用调控植物叶片衰老的分子机制尚不清楚. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}