 ,1,*, 李彪1,2, 韩一萱1,2, 顾秋锦1,3, 万洪秀1
,1,*, 李彪1,2, 韩一萱1,2, 顾秋锦1,3, 万洪秀1Responses of freshwater ecosystems to global change: research progress and outlook
Peng XING,1,*, Biao LI1,2, Yi-Xuan HAN1,2, Qiu-Jin GU1,3, Hong-Xiu WAN1通讯作者: * Email:pxing@niglas.ac.cn
编委: 牛书丽
责任编辑: 李 敏
收稿日期:2020-01-4接受日期:2020-04-19网络出版日期:2020-05-20
| 基金资助: |
Received:2020-01-4Accepted:2020-04-19Online:2020-05-20
| Fund supported: |

摘要
全球变化已经通过提高水温、改变降水格局和水流状况、促进物种入侵、增加极端事件, 对不同的淡水生态系统造成严重的威胁。该文将全球变化背景下淡水生态学的主要研究内容归纳为: (1)全球变化各要素对个体、种群、群落及至生态系统水平的影响; (2)全球变化过程中生态系统生物地球化学循环的改变; (3)淡水生态系统对全球变化的适应对策。最近10-15年淡水生态系统与全球变化研究快速发展, 取得的重要突破有: (1)阐明淡水生态系统结构与功能对全球气候变化尤其是水温升高的响应过程与机制; (2)揭示淡水生态系统(湿地、湖泊、河流等)是全球碳循环的重要组成部分, 在全球变化因素的影响下呈现有机碳埋藏减少和矿化速率提高。今后的研究中, 需要进一步加强对淡水生态系统全要素的系统观测与整合; 开展以“河流”为介质耦合多系统的碳输运和转化过程研究; 强化基础理论研究揭示淡水生态系统对全球变化的适应机制。
关键词:
Abstract
Global change has already posed a serious threat to different freshwater ecosystems by raising water temperatures, changing precipitation patterns and water flow conditions, enhancing species invasion, and increasing extreme events. In order to identify the major works carried out and highlights of the outcomes of research in freshwater ecology in the context of global change, we conducted literature search and analysis of papers published during 1900-2018 via Web of Science. In this review, the major researches in freshwater ecology in the context of global change are categorized into: (1) the effects of various global change factors on individuals, populations, communities and ecosystems; (2) changes in biogeochemical cycles of ecosystems in the process of global change; and (3) adaptation strategies of freshwater ecosystems to global changes. Over the past 10-15 years, research in freshwater ecosystems and global change progressed rapidly and showed breakthroughs in the following aspects: (1) elucidated the response processes and mechanisms of the structure and function of freshwater ecosystems to global climate change, in particular rising water temperatures; (2) revealed that freshwater ecosystems (wetlands, lakes, rivers, etc.) are important components of the global carbon cycle, such that under the influence of global change factors organic carbon burial decreased and mineralization rate increased. In future research, it is necessary to strengthen the systematic observations and integration of the total elements of freshwater ecosystems, to conduct research on carbon transport and transformation processes mediated by the river-connected multi-systems, and to strengthen basic theoretical research for uncovering the adaptation mechanisms of freshwater ecosystems to global change.
Keywords:
PDF (1067KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
邢鹏, 李彪, 韩一萱, 顾秋锦, 万洪秀. 淡水生态系统对全球变化的响应: 研究进展与展望. 植物生态学报, 2020, 44(5): 565-574. DOI: 10.17521/cjpe.2020.0009
XING Peng, LI Biao, HAN Yi-Xuan, GU Qiu-Jin, WAN Hong-Xiu.
1 基本概念及研究现状的文献计量分析
1.1 淡水生态系统与全球变化
淡水生态系统是指由淡水生物群落与水环境所组成的一类生态系统, 包括流水生态系统和静水生态系统。流水生态系统是指由流动水体构成的淡水生态系统, 如江河、溪流、水沟、水渠等; 静水生态系统是指由相对静止水体(流动和更换缓慢)构成的淡水生态系统, 如湖泊、水库、池塘等。冰川作为地球固态淡水资源的主体, 其演变也可以纳入淡水生态系统研究的范畴。淡水生态系统主要以水作为其环境介质, 而陆地生态系统主要以空气、陆地或土壤作为其环境介质。正是由于这些环境介质理化特征的不同, 使水、陆两类生态系统在系统的结构和功能上存在着许多明显的差异。全球变化是指由自然和人文因素引起的地球系统结构和功能的变化以及这些变化产生的影响, 其主要内容包括大气成分变化、全球气候变化、海平面上升、生物多样性以及生物入侵、人口增长、经济技术发展、土地利用和土地覆盖变化等(于振良, 2017)。
淡水生态系统作为地球系统的组成部分, 对自然和人文因素引起的地球系统结构和功能的变化极为敏感。淡水只占世界水资源的0.01%, 约占地球表面的0.8%, 但却维系着全球6%的已知生物物种 (Dudgeon et al., 2006)。淡水生态系统提供了广泛的生态系统服务, 如洪水调节、食物供应和文化重要性, 这与现有生物具有的功能多样性有着内在的联系, 因此可以连接人类社会与他们的生存环境。由于土地利用的集约化、点源和非点源污染、河道改造和过度开发, 淡水生态系统是地球上退化最严重的栖息地(V?r?smarty et al., 2010)。21世纪, 气候变化和人为因素不仅对受威胁和濒临灭绝的淡水物种, 而且对生态系统的功能产生深远的影响。为了维持生物多样性和关键的生态系统进程, 淡水生态系统的保护和恢复必须是一项最优先事项。
1.2 全球变化淡水生态学及其研究内容
生态学研究已经从传统的关注有机体扩展到关注作为一个完整生态系统的地球。在卫星遥感和全球气候计算机模型的辅助下, 我们可以记录地球的基本参数, 包括它的净初级生产力、生物地球化学循环和人类对地球的影响。另外, 分子技术使我们能够认识微生物群落, 它们主导着地球上如此多的生物地球化学循环; 质谱分析稳定同位素在不同来源和流通中的比例, 彻底改变了我们对生物在控制地球化学循环中重要性的认识。在此背景下, 全球变化生态学应运而生, 它是一门宏观与微观相互交叉、多学科相互渗透的前沿科学, 重点研究全球变化领域的基本生态学问题及其相互关系, 解决大尺度环境问题(Schlesinger, 2006)。淡水生态系统与全球变化关系的研究内容主要包括以下三点: (1)研究全球变化各要素对个体、种群、群落及至生态系统水平的影响; (2)探讨全球变化过程中生态系统生物地球化学循环的改变; (3)研究淡水生态系统对全球变化的适应对策。1.3 国际研究文献计量分析
本文研究数据来源于Web of Science (WOS)中的科学引文索引扩展版(Science Citation Index Expanded, SCI-E, 简称SCI), 分别以主题词: “global change” & lake or wetland or river or reservoir or glacier, 对SCI数据库时间范围1900-2018年的文献进行检索。检索时间为2019年11月20日, 检索文献类型界定为“Article (论文)”和“Review (综述)”, 不包括会议录文献、会议摘要、书评、信函、社论材料等。共检索到文献14 427篇, 其中最近10年(2009- 2018年)发表文章10 804篇, 占总发表文章数量的74.9%。淡水生态学在2005年前后进入指数增长期(R2 = 0.987, p < 0.05), 我国相关主题论文发表数量在2010年后开始迅速增长(图1), 2018年发表论文数量接近全球论文总量的1/3。淡水生态系统与全球变化研究涉及环境科学与生态学、地质学、水资源、气象学、自然地理学等相关学科(图2)。以检索的文献为研究对象, 使用数据库平台中的分析功能、DDA软件的清洗功能以及Excel软件的绘图和统计功能等, 结合文献阅读和淡水生态学领域专家的建议, 对检索结果进行数据合并、去重等清洗后进行各指标定量分析(表1)。本文在对国内外全球变化研究成果综合分析的基础上, 总结了淡水生态系统与全球变化研究的阶段性研究成果, 并对未来发展趋势进行简单归纳。图1
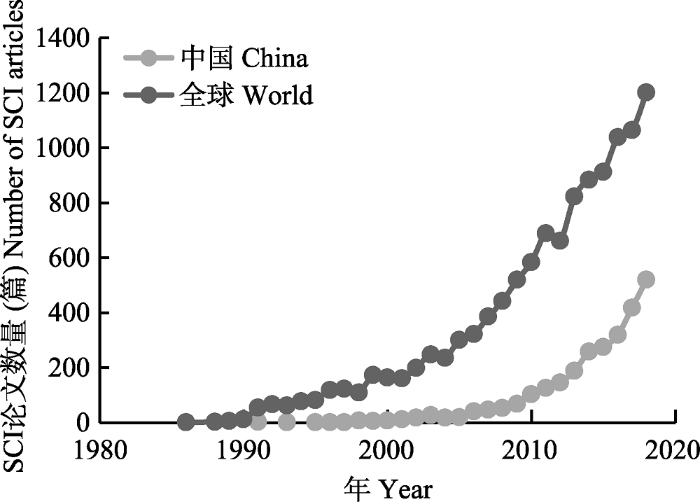 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1全球变化淡水生态系统研究发表论文数量。
Fig. 1Papers published within the scope of freshwater ecosystem research under global change.
图2
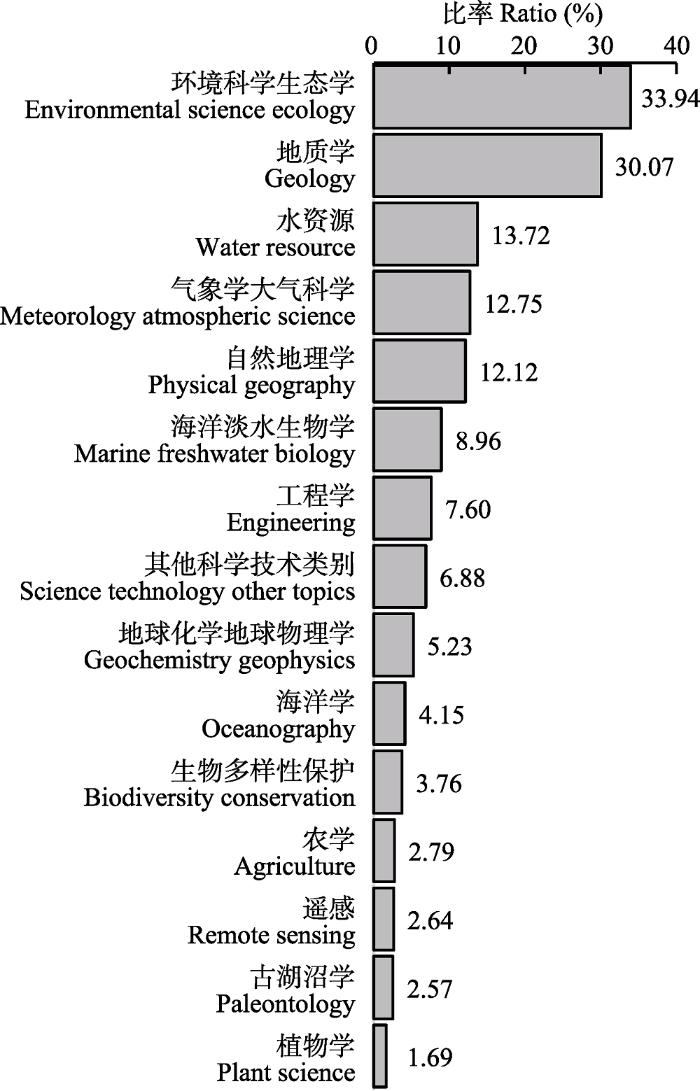 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2全球变化淡水生态系统研究涉及的学科领域。
Fig. 2Research areas and relative contributions in the responses of freshwater ecosystems to global change.
Table 1
表1
表1近10年全球变化淡水生态系统研究TOP 20热点关键词
Table 1
| 排序 Order | 关键词 Key | 中文释义 Chinese | 被引频次 Citation |
|---|---|---|---|
| 1 | Climate change | 气候变化 | 1 883 |
| 2 | Global change | 全球变化 | 349 |
| 3 | Greenhouse gas | 温室气体 | 319 |
| 4 | Global warming | 全球变暖 | 303 |
| 5 | Hydrology | 水文学 | 266 |
| 6 | Carbon | 碳 | 211 |
| 7 | Land use | 土地利用 | 200 |
| 8 | Temperature | 温度 | 183 |
| 9 | Remote sensing | 遥感 | 166 |
| 10 | Eutrophication | 富营养化 | 154 |
| 11 | Precipitation | 降水 | 152 |
| 12 | Biodiversity | 生物多样性 | 146 |
| 13 | Methane | 甲烷 | 133 |
| 14 | Drought | 干旱 | 120 |
| 15 | Flooding | 洪水 | 117 |
| 16 | Sea level rise | 海平面上升 | 117 |
| 17 | Water resources | 水资源 | 114 |
| 18 | Water quality | 水质 | 106 |
| 19 | Modeling | 建模 | 103 |
| 20 | Ecosystem services | 生态系统服务 | 100 |
新窗口打开|下载CSV
2 研究现状分析
2.1 变化要素的生态效应
在淡水生态系统面临的一系列复杂威胁中, 气候变化是一个重要的压力来源。气候变化已经通过提高水温, 改变水流状况, 增加极端事件(如洪水、干旱、野火)和促进物种入侵, 对不同的淡水生态系统造成了压力。人们普遍预测在21世纪, 气候变化更会全面超越那些较为局部化的影响, 如人类活动造成的栖息地丧失和破碎化、富营养化和酸化等(V?r?smarty et al., 2010)。2.1.1 全球变暖
气候变化导致的水温升高将改变基本的生态过程和水生物种的地理分布。水生物种能否成功地在陆地上迁移将取决于是否存在扩散的通道, 这些通道因地域而异并且经常遭到人类活动的破坏。目前分布在低地的溪流和河流中但无法向北或者高纬度扩散的, 以及仅能适应冷水的鱼类(如鳟鱼和鲑鱼), 水温升高增加了其物种灭绝和生物多样性丧失的可能性(Clark et al., 2001)。相比之下, 许多喜欢温暖水域的鱼类, 如大口黑鲈(Micropterus salmoides)和鲤鱼(Cyprinus carpio), 随着地表水变暖, 可能会扩大它们在北美地区的分布范围(Eaton & Scheller, 1996)。
水温升高导致水生生物个体小型化。Daufresne等(2009)通过对长期调查、实验数据和已发表的结果整合分析, 研究了气候变化对变温水生生物(细菌、浮游植物、浮游动物和鱼类)体型从群落到个体水平的影响。结果显示小型物种和幼龄物种的比例显著增加, 而幼龄物种的尺寸也减小了。研究提供的证据表明, 体型变小是暨“物种向更高的海拔和纬度的转移”和“生命周期事件的季节性变化”之后, 水生系统对全球变暖的第三个普遍的生态反应。在此之前, 关于全球变暖对生物群的影响一直缺乏一般的规律。鱼类体积变小会对生态环境产生严重影响, 如体积变小将使鱼类的产卵量下降, 从而打破食物链和生态系统的平衡。
水温升高简化了淡水溪流食物网结构。气候变暖增加了消费者的新陈代谢需求, 加强了它们的摄食相互作用。这可能会改变能量流, 甚至扩大食物网中的灭绝率。O’Gorman等(2019)研究证实, 较高的温度简化了食物网络结构, 缩短了消费者和资源之间的能量流动路径。Hansson等(2013)在建模、监测和实验数据的基础上, 研究发现淡水群落对全球变化的反应是由食物链长度决定的。某一营养级是否获益与其所在的层次有关, 以长度为三级的食物链为例, 最上层和最下层营养级将受益于气候变化, 而中间营养级将受到不利影响。不过, 研究还发现与其他类群相比, 蓝藻(产生毒素的有害浮游植物)却可以受益于较高的温度和腐殖质含量, 而与食物链的组成无关。气候变化和水体腐殖质含量增加(褐变)之间的协同作用将对未来的水生态系统产生相当大的影响。
气候变暖改变全球湖泊湖水混合特征。全球变暖导致湖泊冰雪覆盖层快速融化、湖面水温升高, 影响湖泊的混合特征。Woolway和Merchant (2019)构建了一维湖泊数学模型, 将两种排放模式下4种最优模型计算的结果作为输入参数, 开展全球635个湖泊湖水混合特征变化预测。在高排放模型情景下——典型浓度路径(RCP) 6.0, 湖泊冰雪覆盖层将大幅减少, 预计最迟到2100年约1/4的季节性冰雪覆盖湖泊将不再出现冰封期。湖泊表层水温预计将升高, 中等变暖条件下升高约2.5 ℃, 极端变暖条件下升高约5.5 ℃。635个湖泊中约有100个湖泊的湖水混合状态将发生变化, 其中约有1/4的湖泊由单混合(即每年只发生一次湖水上下混合)变为永久分层湖泊, 约有1/6的湖泊由双混合(即每年湖水上下混合二次)将成为单混合。气候变化背景下, 湖泊总体呈现混合频率减弱的趋势, 这无疑将影响湖泊内部以及湖泊与其他系统之间的生物地球化学循环。不仅如此, 全球受洪水影响的程度将随气候变暖的程度而增加。只有实施额外的洪水风险管理措施, 才有可能减少气候变暖导致的洪水泛滥损失, 保障国计民生(Willner et al., 2018)。
2.1.2 降水和径流变化
降水和径流的变化改变了水生生物栖息地的数量和质量, 从而间接影响了生态系统的生产力和多样性。
冰川径流量持续增加。受冰川影响的大型流域, 覆盖了格陵兰岛和南极洲以外的26%的全球陆地表面, 以及全球近1/3的人口。由于气候环境的影响, 全球高山冰川预计在21世纪初的9-11年间, 经历大规模的退缩和质量损失。全球冰川变化的最新估计显示, 冰川质量损失速度为(259 ± 28) Gt·a-1, 全球冰川径流超过1 350 km3·a-1, 冰川消融引起的水文和河流地貌变化预计将对河流生态系统产生重大影响。Brown等(2018)通过功能性状分析揭示了生物多样性对环境变化反应背后的全球机制。从3个大洲的9个生物地理区域的河流中收集了包括123万无脊椎动物在内的363项记录, 对这些记录进行评估, 结果表明, 不同区域群落特征组成和多样性对冰川覆盖梯度表现出了一致的反应。在考虑了纬度的系统区域效应后, 河流无脊椎动物功能多样性的形成过程在全球范围内是一致的。嵌套在单个区域内的分析发现, 随着冰川覆盖面积的减少, 功能多样性有所增加。尽管环境过滤在高度冰川化的盆地中也很明显, 群落聚集模型表明扩散限制是这些模式下的主导过程。这些发现表明, 冰川覆盖度变化威胁着河流无脊椎动物的生物多样性及其在多种生态系统功能和服务中的中心作用。
湿地是经历快速变化的水文复杂系统。据估计, 大多数国家30%-90%的湿地已被破坏或高度改造(Davidson, 2014)。湿地独特的属性影响着全球生物地球化学和水循环, 并在大尺度上提供生态系统服务(Barbier, 2011)。而气候变化预计会通过水文变化、温度升高和海平面上升给湿地增加额外的压力(Junk et al., 2013)。预期这些压力会对湿地产生不同的影响, 需要特定的研究和管理来了解它们保护的特殊需求(Erwin, 2009)。
降水格局的变化导致干旱和半干旱地区的封闭流域成为全球无机碳汇。盆地终端湖泊中可能形成大量溶解无机碳的封存。几十年来, 由于人类活动和气候的密集化改变, 国际重要湖泊湿地的萎缩和干涸加剧。面积大于1 km2的湖泊从20世纪80年代后期的785个降低至2010年的577个, 其中中国内蒙古降低速率(34%)高于蒙古国(17.6%), 面积大于10 km2的湖泊数量同期也减少了30% (Tao et al., 2015)。未来几十年, 不仅由于气候变化, 而且由于对高原地下矿物和地下水资源的不断开发, 预计温带湖泊的生态环境恶化还将继续。由于湖泊和草原的退化, 内蒙古高原已成为中国北方沙尘暴的主要来源。研究者对我国西北干旱地区石羊河流域终端区地下水、河水和沉积物进行了同位素、放射性碳和化学分析, 并以全球11个内流盆地沉积资料为基础, 对全球内流盆地溶解性无机碳的封存进行了推断。结果表明, 内流盆地是全球范围内重要的碳汇, 其规模与深海碳埋藏相当(Li et al., 2017)。干旱和半干旱生态系统日益被认为是重要的碳储存场所。
由气候变化引起的水资源短缺和水温上升, 将会直接威胁水-能源联系。全球大多数热电厂所处的地区, 平均年流量会下降的同时水温也会大幅上升, 这两种情况都限制了冷却水使用。对于大多数地区来说, 即便将电厂效率提高20%, 仍不足以缓解气候变化下冷却水使用潜力的总体下降。此外, 地下水枯竭已经成为全球性问题, 它直接关系到人类赖以生存的水和粮食安全(Aeschbach-Hertig & Gleeson, 2012)。研究者结合遥感数据归一化植被指数(NDVI)和长期的水平衡蒸散(ET)测量得出半湿润和半干旱盆地“绿化”将消耗更多的水, 导致河流流量显著减少(24%-28%)。同时, CO2浓度升高也会加剧植被用水量, 从而进一步减少缺水地区旳径流量(Pall et al., 2011)。
2.1.3 土地利用变化
全球土地利用正在以惊人的速度变化, 而且在未来很长一段时间内可能还会继续变化。与人类土地利用有关的对河流生态系统改变, 包括水文变化和沉积物再悬浮, 河岸植被组成, 大型无脊椎动物群落, 营养盐浓度和有机质动态变化等等(Griffiths et al., 2009)。此外, 特定的土地利用变化, 例如森林用地转为农业用地或农业用地转为郊区用地, 可能对流入河流的有机质动态产生多重影响, 并可能对水生食物网产生连锁影响。例如, 与土地利用变化相关的河岸物种的组成变化可以改变凋落物的组成, 进而影响水生生态系统中有机物的处理效率(Tank et al., 2010)。
在湖泊中, Cl-在低浓度时是一种相对无害的离子, 但当浓度升至100-1 000 mg·L-1 时, 开始对生态系统产生影响。Dugan等(2017)在全球湖泊密度最高的北温带调查了371个淡水湖, 分析其中氯化物浓度的长期变化趋势。结果显示不断增加的城市化和相关含氯径流会使淡水盐碱化, 并威胁到湖泊的水质和湖泊提供的许多生态系统服务。研究预测, 如果目前的趋势继续下去, 在未来50年内, 绝大部分湖泊将超过美国环境保护署(EPA)规定的长期氯暴露的水生生物阈值标准(230 mg·L-1)。
2.1.4 生物入侵
气候变化和人类活动为外来生物扩散和入侵提供了有利的途径, 未来气候变化和生物入侵可能相互作用成为全球生物多样性的主要威胁。近年来, 大量研究开始关注生物入侵如何改变淡水生态系统结构, 影响生物多样性以及生态系统服务功能(吴昊和丁建清, 2014)。气候变化导致本地生境的环境紊乱度增加, 提升外来物种的入侵风险。Sorte等(2013)采用meta分析, 评估了157个入侵物种和204个本地物种对气候条件(不同温度、CO2和降水条件)变化的反应。结果显示在陆地(主要是植物)生态系统中, 本地和入侵物种对环境变化的反应相似。相比之下, 在水生(主要是动物)生态系统中, 温度上升和CO2浓度升高在很大程度上抑制了本地物种。面对气候变化, 入侵物种表现出更强烈的反应, 包括对更有利条件做出更积极的反应以及对较不利条件做出更消极的反应。因此, 随着气候变化的加剧, 淡水生态系统可能特别容易受到生物入侵的威胁。此外, 全球变化下频繁的人类活动(经济一体化、国际交往)给外来种提供了多种入侵机会和渠道, 并且国际贸易、旅游等行为引起的生物入侵危害具有潜伏性和持续性。
2.1.5 其他因素
(1)重大生态工程。人类不仅通过增加有机碳和营养物质对河流造成负荷, 而且通过大规模建造大坝深刻地改变了碳固定、矿化和沿着河流连续体储存有机碳之间的平衡(Shields et al., 2017; Veldkamp et al., 2017)。在过去的一个世纪里, 由于修建了5万座大坝, 导致世界上大约2/3的河流都受到了损害。大坝水库是沿河流连续介质沉积、初级生产力和有机碳矿化的热点地区。大坝的建造还会引起水库内淹没的生物量增加和土壤有机质的释放。这些过程改变了下游有机碳和养分的转移, 从而改变了河流系统的营养状态, 这可以从许多河流在大坝关闭后CO2分压的下降中看出。目前由于筑坝而引起的河流有机碳通量的全球变化仍然难以量化。
(2)富营养化。大量研究表明, 富营养化、CO2浓度上升和全球变暖可能会增加全球许多水生生态系统中蓝藻繁殖的频率、强度和持续时间(Ho et al., 2019)。蓝藻水华暴发对水生食物网的生物多样性和功能产生负面影响, 并威胁到湖泊生态系统饮用水、渔业和其他景观用途。近几十年来, 全球氮肥的使用逐渐超过了磷肥的使用, 氮负荷的增加和氮磷比的提高可能改变蓝藻水华的物种组成。特别是可以增加微囊藻、浮游菌等非固氮蓝藻的数量。近期的研究更打破了蓝藻通常被认为只在低浓度CO2和高pH下是优越竞争者的范式, 蓝藻碳酸氢盐吸收系统的遗传多样性和生理灵活性, 使蓝藻快速适应CO2浓度的升高(Huisman et al., 2018)。
2.2 生物地球化学循环
2.2.1 碳循环内陆水体储存了陆地生态系统近一半的碳, 并参与碳的形态转化和气体释放, 对于全球碳循环发挥了重要作用(Tranvik et al., 2009)。陆地向内陆水体的碳输入往往和陆地净生产力处于相同数量级, 因此, 忽略内陆水体碳收支中的作用将会高估陆地温室气体吸收和储存量。
湖泊沉积物中有机碳的矿化受到温度的控制(Gudasz et al., 2010)。湖泊和水库中有机碳的年埋藏量超过了海洋沉积物, 结果表明, 湖泊沉积物中有机碳的矿化与温度呈强正相关关系, 说明水温升高导致有机碳矿化程度升高, 有机碳埋藏程度降低。该研究预计温度升高(IPCC提出的气候变化情景)将导致北方湖泊有机碳埋藏量减少4%-27% (相当于0.9-6.4 Tg C·a-1)。有机碳矿化速率提高导致湖泊等内陆水体成为温室气体排放的热点区域。湖泊仅占全球陆地面积的3%, 但每年CH4排放量达到0.65 Pg C·a -1 (换算成CO2当量)。根据估算, 湖泊释放的CH4和CO2等温室气体总量, 可以相当于79%的陆地温室气体碳汇。2011年发表在Science上的报道引起人们对湖泊在全球气候变化中作用的高度关注(Bastviken et al., 2011)。
淡水生态系统温室气体由于来源和产生过程复杂, 估算和预测其规模都是很大的挑战。冰川融化导致的径流量增加使得陆源有机质在河流系统中的分解转化过程增强(Datry et al., 2018)。目前河流和溪流自生代谢产生的CO2, 还未包括在更大尺度的碳收支估算及碳循环和归趋研究中(Hotchkiss et al. 2015)。湿地CH4排放是全球最大的CH4自然来源, 约占自然和人为排放总量的1/3。然而, 国际耦合模式比较计划第五阶段(CMIP5)中未考虑自然湿地和冻土融化释放的CO2、CH4对温度升高存在的正反馈效应(McCalley et al., 2014; Yvon-Durocher et al., 2017)。湿地(永久或季节性饱和的陆地区域)未来扩张和CH4排放将演变到何种程度以及如何推动气候反馈仍然是一个重大问题(Comyn-Platt et al., 2018)。
此外, 河流和冰川径流形成的碳输移对于相邻生态系统产生了显著的影响。冰川径流流量和时间的变化, 改变了海洋生态系统中碳源可用性和异养生产力, 特别是在具有重要商业价值的渔业产区(例如北大西洋的阿拉斯加湾流域), 这种陆源碳输入效应尤为明显。格陵兰冰盖径流增加导致阿拉斯加湾流域渔业产量在1961-2003年增加了41%。
2.2.2 氮循环
近年来, 淡水生态系统内部的氮循环过程研究得到了长足的发展, 但是相对于碳循环, 氮循环观测数据严重不足, 尚无法形成全球和区域的氮循环参数的大尺度格局。已有证据显示河流NO3-浓度的增加并不一定会导致N2O释放的增加, 但河流日益严重的缺氧可能造成N2O释放增加。N2O是一种强效的温室气体, 农业排放的N2O中有17%来自小溪、河流和河口, 而这又是无机氮通过浸出、径流和污水输入的结果。政府间气候变化和全球N2O估算小组假设, 河流N2O的排放量随着溶解的无机氮负荷呈线性增加。Rosamond等(2012)报道了对加拿大Grand River为期两年的N2O排放量测量, 结果显示城市地区和夏季夜间, 河流N2O排放量显著偏高。此外, 基于溶解无机氮负荷的年排放量估算高估了湿润年份的测量排放量, 而低估了干燥年份的排放量。更为重要的是, 他们并未发现N2O排放与硝酸盐或溶解无机氮的相关性, 但却检测到与溶解氧负相关。
2.2.3 碳-氮-磷交互作用
生态学家逐渐认识到, 生态系统碳循环并不是独立发生的, 而是与其他元素如氮、磷循环密切耦合在一起。碳-氮-磷循环的交互作用是当前生物地球化学研究中的热点(Schlesinger et al., 2011)。以富营养化与蓝藻水华控制为例, 究竟是控磷还是氮磷双控一直是国际湖沼学界长期争而未决的问题。目前, 富营养化控制策略主要基于小水体的营养盐添加模拟外源输入实验, 但忽略了营养盐在湖泊内的生物地球化学循环过程。通过收集整理全球573个湖泊的形态与营养状况数据, 比较湖泊混合层深度(epilimnion)与平均和最大水深, 将湖泊分为浅水湖泊(混合深度>最大深度)、深水湖泊(混合深度<平均深度)和过渡型湖泊(平均深度≤混合深度≤最大深度)。以TN:TP (质量比)作为湖泊氮磷限制指标, 当N:P < 9时为氮限制, 当9 ≤ N:P < 22.6时为氮磷双限制, 当N:P ≥ 22.6时为磷限制。meta分析结果显示, 磷控制虽然可以减轻大多数湖泊的富营养化, 但是在富营养化湖泊中, 尤其是在浅水湖泊(或海湾)中, 可能需要氮和磷双控制才能有效控制富营养化和藻类水华。因此, 湖泊富营养化控制需要根据湖泊的水深和水体混合特性采取不同的控制策略(Qin et al., 2020)。
2.3 全球变化大型控制实验和数据积累
全球变化生态学研究手段与传统淡水生态学交叉融合, 呈现出蓬勃发展的态势:(1)模拟复杂生态系统对全球变化响应。随着生态系统弹性理论、稳态转换理论、食物网上行和下行效应等理论的提出, 以及预测未来生态系统变化趋势的需要, 近年来科研人员着手建立尺度更大、生态复杂性更高的模拟实验体系, 以研究增温对群落和水域生态系统稳态转换的影响。例如2003年由EUROLIMPACS资助在丹麦建立的人工池实验一直运行到现在, 这些装置模拟并检测增温和多环境胁迫因子对生态结构的影响, 长期运行积累的数据成为全球变化淡水生态系统响应研究的第一手资料, 是丰富模型参数和提高模型预测能力的重要依据。
(2)建立全球资料和信息系统。淡水生态系统需要借助全球模式来定量分析系统内物理、化学和生物过程的相互作用, 估计未来变化的可能影响, 因此建立全球资料和信息的处理、贮存和交流系统是全球变化研究基本的出发点。基于长时间和大空间尺度的湖泊生态系统对全球变化响应趋势研究, 是当前国际学术界关注的热点, 并成为一些重要生态学理论发展的基础。国际湖泊生态学研究人员于2005年发起成立的全球湖泊生态系统观测网络(Global Lake Ecosystem Observation Network, GLEON), 通过强化湖沼学研究的5个基础: 自然历史、多尺度数据、实验、理论和比较研究(尤其强调多尺度数据和比较研究), 在湖泊生态系统全球观测方面取得重大进展(Hanson et al., 2016)。中国科学院组织院内所属的17个野外站(点), 通过与其他系统的野外站联合组建“高寒区地表过程与环境监测研究网络(http://www.horn.ac.cn/)”在全球气候变化敏感区开展联网观测, 为揭示大江大河源头区气候变化规律和水资源形成转化规律、合理开发利用水资源等提供数据支持。
2.4 新技术与新方法
在这个大数据的新时代, 我们正处在能够将生态学和许多完全不同领域的其他学科结合起来的难得机遇期。新技术的巨大进步, 包括遥感和地理信息系统(GIS)技术、民众可以参与的从本地到全球的监测活动, 分子生态学中快速增长的下一代测序(NGS)和环境DNA (eDNA)分析能力, 以及处理复杂驱动力系统响应的建模能力, 都将有助于满足全球变化研究对于跨越空间、环境梯度的大数据收集需求的快速增长。这将提高我们目前非常有限的对生物多样性不同水平反应的预测能力, 为实现从基因-生态网络-生态系统-气候变化的系统研究提供重要的新见解。3 总结和展望
本文综述了淡水生态学与全球变化研究当前发展的主要态势, 可以看出该领域在几个关键方向上已经取得了快速进步, 但必须承认对该领域的认识在很大程度上仍然是未知的。在今后的研究中需要加强“多元”方法的集成, 建立起在时间与空间尺度上、生态与进化现象之间目前仍然缺失的联系。这其中的某些方面可以通过重新整合现有的研究方法来完成, 但其他方面可能需要引入全新的方法和思路。3.1 对生态系统全要素的系统观测和整合
由于淡水生态系统极度的多样化和复杂性, 且各个全球变化要素之间存在的交互作用, 使得淡水生态系统与全球变化研究中面临着诸多的挑战。(1)淡水生态系统特别容易受到气候变化的不同组成部分的影响, 但我们仍然对这些环境驱动因素及其与其他压力源的相互作用的后果了解有限, 特别是在更高的、多物种的、组织性的层次上。目前普遍采用的模拟实验方法具有较好的重复性, 能较直接地揭示气候对环境和生物产生影响的机理, 但无法完全模拟自然系统的复杂性, 而且由于对湖泊、湿地等在内的淡水生态系统地表过程认识不足, 尚不能全面揭示生态系统水平的响应特征。(2)全球变化研究要求野外观测可以实现多站点联网、基于统一的方法在不同站点开展长期的生态系统观测。对于高度多样化的淡水生态系统, 针对全球变化过程的联网研究尚处于摸索阶段。(3)生态系统响应全球变化的模式取决于系统本身的演化过程, 对事件发生的响应在短时间无法找到可侦测的表现, 只有通过长期演化过程的研究才能揭示其敏感性、抵抗力与弹性和突变的阈值等(Wang et al., 2012)。如何从生态系统缓慢变化过程中提取增量的信息, 判识突变或灾变发生的早期信号, 是现代生态学研究已经关注、但尚未解决的关键科学问题。3.2 不同生态系统之间的耦合研究
近年来, 内陆水体(特别是河流)对陆源碳的跨系统输送在碳循环研究中受到越来越多的关注。内陆水体每年传输、矿化和埋藏的碳约2.7 Pg (Battin et al., 2009), 约占全球化石燃料燃烧排放碳的31%。由于受到全球变化和人类活动的双重驱动, 陆地-河流-近海的碳输运发生了显著的变化, 这将可能对区域乃至全球的碳循环产生影响。我国河流分布广泛, 流域覆盖人口最为稠密、工农业活动频繁的地区, 加之土地利用变化、水利工程建设以及降水格局变化引起的干旱和洪涝灾害日益频繁, 使得开展陆地-河流-近海的碳输运研究需要考虑的全球变化要素异常复杂。相对其他的碳循环过程, 目前对跨系统的碳输运过程及其对全球变化影响的研究还处于起步阶段。3.3 淡水生态学的基础研究
准确评价、预测淡水生态系统对气候变化的响应和发展趋势, 必须加强生态系统的演变过程研究。例如, 在过去的20年, 人们付出了巨大的努力研究生物多样性-生态系统功能, 以及后来的生物多样性-生态系统服务的关系(Cardinale et al., 2012)。但是, 这些研究都还主要是在随机的物种损失条件下进行的, 未与任何确定的环境驱动力, 尤其是那些与气候变化有关的驱动力相联系。此外, 全球变化研究存在驱动要素与生态系统响应无法一一对应的情况。是否可以通过淡水生态学的基础研究找到适合的用于反映全球变化影响的生态系统“属性”, 是一个值得探讨的问题。这一问题的解决, 有助于开发基于过程的模型, 优化现有模型的预测精度。
致谢
感谢厦门大学史大林教授对文章撰写提出的宝贵意见, 感谢东南大学赵晟明在资料收集整理工作中给予的帮助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1038/NGEO1617URL [本文引用: 1]

Groundwater-the world's largest freshwater resource-is critically important for irrigated agriculture and hence for global food security. Yet depletion is widespread in large groundwater systems in both semi-arid and humid regions of the world. Excessive extraction for irrigation where groundwater is slowly renewed is the main cause of the depletion, and climate change has the potential to exacerbate the problem in some regions. Globally aggregated groundwater depletion contributes to sea-level rise, and has accelerated markedly since the mid-twentieth century. But its impacts on water resources are more obvious at the regional scale, for example in agriculturally important parts of India, China and the United States. Food production in such regions can only be made sustainable in the long term if groundwater levels are stabilized. To this end, a transformation is required in how we value, manage and characterize groundwater systems. Technical approaches-such as water diversion, artificial groundwater recharge and efficient irrigation-have failed to balance regional groundwater budgets. They need to be complemented by more comprehensive strategies that are adapted to the specific social, economic, political and environmental settings of each region.
[本文引用: 1]
URLPMID:21212349 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1017/S1464793105006950URLPMID:16336747 [本文引用: 1]
Freshwater biodiversity is the over-riding conservation priority during the International Decade for Action - 'Water for Life' - 2005 to 2015. Fresh water makes up only 0.01% of the World's water and approximately 0.8% of the Earth's surface, yet this tiny fraction of global water supports at least 100000 species out of approximately 1.8 million - almost 6% of all described species. Inland waters and freshwater biodiversity constitute a valuable natural resource, in economic, cultural, aesthetic, scientific and educational terms. Their conservation and management are critical to the interests of all humans, nations and governments. Yet this precious heritage is in crisis. Fresh waters are experiencing declines in biodiversity far greater than those in the most affected terrestrial ecosystems, and if trends in human demands for water remain unaltered and species losses continue at current rates, the opportunity to conserve much of the remaining biodiversity in fresh water will vanish before the 'Water for Life' decade ends in 2015. Why is this so, and what is being done about it? This article explores the special features of freshwater habitats and the biodiversity they support that makes them especially vulnerable to human activities. We document threats to global freshwater biodiversity under five headings: overexploitation; water pollution; flow modification; destruction or degradation of habitat; and invasion by exotic species. Their combined and interacting influences have resulted in population declines and range reduction of freshwater biodiversity worldwide. Conservation of biodiversity is complicated by the landscape position of rivers and wetlands as 'receivers' of land-use effluents, and the problems posed by endemism and thus non-substitutability. In addition, in many parts of the world, fresh water is subject to severe competition among multiple human stakeholders. Protection of freshwater biodiversity is perhaps the ultimate conservation challenge because it is influenced by the upstream drainage network, the surrounding land, the riparian zone, and - in the case of migrating aquatic fauna - downstream reaches. Such prerequisites are hardly ever met. Immediate action is needed where opportunities exist to set aside intact lake and river ecosystems within large protected areas. For most of the global land surface, trade-offs between conservation of freshwater biodiversity and human use of ecosystem goods and services are necessary. We advocate continuing attempts to check species loss but, in many situations, urge adoption of a compromise position of management for biodiversity conservation, ecosystem functioning and resilience, and human livelihoods in order to provide a viable long-term basis for freshwater conservation. Recognition of this need will require adoption of a new paradigm for biodiversity protection and freshwater ecosystem management - one that has been appropriately termed 'reconciliation ecology'.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1890/07-1876.1URLPMID:19323178 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:29946124 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:25341787 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:23062213 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:28232747 [本文引用: 1]
URLPMID:20882010 [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Regional strategies for the accelerating global problem of groundwater depletion
1
2012
... 由气候变化引起的水资源短缺和水温上升, 将会直接威胁水-能源联系.全球大多数热电厂所处的地区, 平均年流量会下降的同时水温也会大幅上升, 这两种情况都限制了冷却水使用.对于大多数地区来说, 即便将电厂效率提高20%, 仍不足以缓解气候变化下冷却水使用潜力的总体下降.此外, 地下水枯竭已经成为全球性问题, 它直接关系到人类赖以生存的水和粮食安全(
Wetlands as natural assets
1
2011
... 湿地是经历快速变化的水文复杂系统.据估计, 大多数国家30%-90%的湿地已被破坏或高度改造(
Freshwater methane emissions offset the continental carbon sink
1
2011
... 湖泊沉积物中有机碳的矿化受到温度的控制(
The boundless carbon cycle
1
2009
... 近年来, 内陆水体(特别是河流)对陆源碳的跨系统输送在碳循环研究中受到越来越多的关注.内陆水体每年传输、矿化和埋藏的碳约2.7 Pg (
Functional diversity and community assembly of river invertebrates show globally consistent responses to decreasing glacier cover
1
2018
... 冰川径流量持续增加.受冰川影响的大型流域, 覆盖了格陵兰岛和南极洲以外的26%的全球陆地表面, 以及全球近1/3的人口.由于气候环境的影响, 全球高山冰川预计在21世纪初的9-11年间, 经历大规模的退缩和质量损失.全球冰川变化的最新估计显示, 冰川质量损失速度为(259 ± 28) Gt·a-1, 全球冰川径流超过1 350 km3·a-1, 冰川消融引起的水文和河流地貌变化预计将对河流生态系统产生重大影响.
Biodiversity loss and its impact on humanity
1
2012
... 准确评价、预测淡水生态系统对气候变化的响应和发展趋势, 必须加强生态系统的演变过程研究.例如, 在过去的20年, 人们付出了巨大的努力研究生物多样性-生态系统功能, 以及后来的生物多样性-生态系统服务的关系(
Predicting climate change effects on Appalachian Trout: combining GIS and individual-based modeling
1
2001
... 气候变化导致的水温升高将改变基本的生态过程和水生物种的地理分布.水生物种能否成功地在陆地上迁移将取决于是否存在扩散的通道, 这些通道因地域而异并且经常遭到人类活动的破坏.目前分布在低地的溪流和河流中但无法向北或者高纬度扩散的, 以及仅能适应冷水的鱼类(如鳟鱼和鲑鱼), 水温升高增加了其物种灭绝和生物多样性丧失的可能性(
Carbon budgets for 1.5 and 2 °C targets lowered by natural wetland and permafrost feedbacks
1
2018
... 淡水生态系统温室气体由于来源和产生过程复杂, 估算和预测其规模都是很大的挑战.冰川融化导致的径流量增加使得陆源有机质在河流系统中的分解转化过程增强(
A global analysis of terrestrial plant litter dynamics in non-perennial waterways
1
2018
... 淡水生态系统温室气体由于来源和产生过程复杂, 估算和预测其规模都是很大的挑战.冰川融化导致的径流量增加使得陆源有机质在河流系统中的分解转化过程增强(
Global warming benefits the small in aquatic ecosystems
1
2009
... 水温升高导致水生生物个体小型化.
How much wetland has the world lost? Long-term and recent trends in global wetland area
1
2014
... 湿地是经历快速变化的水文复杂系统.据估计, 大多数国家30%-90%的湿地已被破坏或高度改造(
Freshwater biodiversity: importance, threats, status and conservation challenges
1
2006
... 淡水生态系统作为地球系统的组成部分, 对自然和人文因素引起的地球系统结构和功能的变化极为敏感.淡水只占世界水资源的0.01%, 约占地球表面的0.8%, 但却维系着全球6%的已知生物物种 (
Salting our freshwater lakes
1
2017
... 在湖泊中, Cl-在低浓度时是一种相对无害的离子, 但当浓度升至100-1 000 mg·L-1 时, 开始对生态系统产生影响.
Effects of climate warming on fish thermal habitat in streams of the United States
1
1996
... 气候变化导致的水温升高将改变基本的生态过程和水生物种的地理分布.水生物种能否成功地在陆地上迁移将取决于是否存在扩散的通道, 这些通道因地域而异并且经常遭到人类活动的破坏.目前分布在低地的溪流和河流中但无法向北或者高纬度扩散的, 以及仅能适应冷水的鱼类(如鳟鱼和鲑鱼), 水温升高增加了其物种灭绝和生物多样性丧失的可能性(
Wetlands and global climate change: the role of wetland restoration in a changing world
1
2009
... 湿地是经历快速变化的水文复杂系统.据估计, 大多数国家30%-90%的湿地已被破坏或高度改造(
Rapid decomposition of maize detritus in agricultural headwater streams
1
2009
... 全球土地利用正在以惊人的速度变化, 而且在未来很长一段时间内可能还会继续变化.与人类土地利用有关的对河流生态系统改变, 包括水文变化和沉积物再悬浮, 河岸植被组成, 大型无脊椎动物群落, 营养盐浓度和有机质动态变化等等(
Temperature-controlled organic carbon mineralization in lake sediments
1
2010
... 湖泊沉积物中有机碳的矿化受到温度的控制(
Networked lake science: How the Global Lake Ecological Observatory Network (GLEON) works to understand, predict, and communicate lake ecosystem response to global change
1
2016
... (2)建立全球资料和信息系统.淡水生态系统需要借助全球模式来定量分析系统内物理、化学和生物过程的相互作用, 估计未来变化的可能影响, 因此建立全球资料和信息的处理、贮存和交流系统是全球变化研究基本的出发点.基于长时间和大空间尺度的湖泊生态系统对全球变化响应趋势研究, 是当前国际学术界关注的热点, 并成为一些重要生态学理论发展的基础.国际湖泊生态学研究人员于2005年发起成立的全球湖泊生态系统观测网络(Global Lake Ecosystem Observation Network, GLEON), 通过强化湖沼学研究的5个基础: 自然历史、多尺度数据、实验、理论和比较研究(尤其强调多尺度数据和比较研究), 在湖泊生态系统全球观测方面取得重大进展(
Food-chain length alters community responses to global change in aquatic systems
1
2013
... 水温升高简化了淡水溪流食物网结构.气候变暖增加了消费者的新陈代谢需求, 加强了它们的摄食相互作用.这可能会改变能量流, 甚至扩大食物网中的灭绝率.
Widespread global increase in intense lake phytoplankton blooms since the 1980s
1
2019
... (2)富营养化.大量研究表明, 富营养化、CO2浓度上升和全球变暖可能会增加全球许多水生生态系统中蓝藻繁殖的频率、强度和持续时间(
Sources of and processes controlling CO2 emissions change with the size of streams and rivers
1
2015
... 淡水生态系统温室气体由于来源和产生过程复杂, 估算和预测其规模都是很大的挑战.冰川融化导致的径流量增加使得陆源有机质在河流系统中的分解转化过程增强(
Cyanobacterial blooms
1
2018
... (2)富营养化.大量研究表明, 富营养化、CO2浓度上升和全球变暖可能会增加全球许多水生生态系统中蓝藻繁殖的频率、强度和持续时间(
Current state of knowledge regarding the world’s wetlands and their future under global climate change: a synthesis
1
2013
... 湿地是经历快速变化的水文复杂系统.据估计, 大多数国家30%-90%的湿地已被破坏或高度改造(
Substantial inorganic carbon sink in closed drainage basins globally
1
2017
... 降水格局的变化导致干旱和半干旱地区的封闭流域成为全球无机碳汇.盆地终端湖泊中可能形成大量溶解无机碳的封存.几十年来, 由于人类活动和气候的密集化改变, 国际重要湖泊湿地的萎缩和干涸加剧.面积大于1 km2的湖泊从20世纪80年代后期的785个降低至2010年的577个, 其中中国内蒙古降低速率(34%)高于蒙古国(17.6%), 面积大于10 km2的湖泊数量同期也减少了30% (
Methane dynamics regulated by microbial community response to permafrost thaw
1
2014
... 淡水生态系统温室气体由于来源和产生过程复杂, 估算和预测其规模都是很大的挑战.冰川融化导致的径流量增加使得陆源有机质在河流系统中的分解转化过程增强(
A simple model predicts how warming simplifies wild food webs
1
2019
... 水温升高简化了淡水溪流食物网结构.气候变暖增加了消费者的新陈代谢需求, 加强了它们的摄食相互作用.这可能会改变能量流, 甚至扩大食物网中的灭绝率.
Anthropogenic greenhouse gas contribution to flood risk in England and Wales in autumn 2000
1
2011
... 由气候变化引起的水资源短缺和水温上升, 将会直接威胁水-能源联系.全球大多数热电厂所处的地区, 平均年流量会下降的同时水温也会大幅上升, 这两种情况都限制了冷却水使用.对于大多数地区来说, 即便将电厂效率提高20%, 仍不足以缓解气候变化下冷却水使用潜力的总体下降.此外, 地下水枯竭已经成为全球性问题, 它直接关系到人类赖以生存的水和粮食安全(
Water depth underpins the relative roles and fates of nitrogen and phosphorus in lakes
1
2020
... 生态学家逐渐认识到, 生态系统碳循环并不是独立发生的, 而是与其他元素如氮、磷循环密切耦合在一起.碳-氮-磷循环的交互作用是当前生物地球化学研究中的热点(
Dependence of riverine nitrous oxide emissions on dissolved oxygen levels
1
2012
... 近年来, 淡水生态系统内部的氮循环过程研究得到了长足的发展, 但是相对于碳循环, 氮循环观测数据严重不足, 尚无法形成全球和区域的氮循环参数的大尺度格局.已有证据显示河流NO3-浓度的增加并不一定会导致N2O释放的增加, 但河流日益严重的缺氧可能造成N2O释放增加.N2O是一种强效的温室气体, 农业排放的N2O中有17%来自小溪、河流和河口, 而这又是无机氮通过浸出、径流和污水输入的结果.政府间气候变化和全球N2O估算小组假设, 河流N2O的排放量随着溶解的无机氮负荷呈线性增加.
Global change ecology
1
2006
... 生态学研究已经从传统的关注有机体扩展到关注作为一个完整生态系统的地球.在卫星遥感和全球气候计算机模型的辅助下, 我们可以记录地球的基本参数, 包括它的净初级生产力、生物地球化学循环和人类对地球的影响.另外, 分子技术使我们能够认识微生物群落, 它们主导着地球上如此多的生物地球化学循环; 质谱分析稳定同位素在不同来源和流通中的比例, 彻底改变了我们对生物在控制地球化学循环中重要性的认识.在此背景下, 全球变化生态学应运而生, 它是一门宏观与微观相互交叉、多学科相互渗透的前沿科学, 重点研究全球变化领域的基本生态学问题及其相互关系, 解决大尺度环境问题(
Introduction to coupled biogeochemical cycles
1
2011
... 生态学家逐渐认识到, 生态系统碳循环并不是独立发生的, 而是与其他元素如氮、磷循环密切耦合在一起.碳-氮-磷循环的交互作用是当前生物地球化学研究中的热点(
Carbon storage in the Mississippi River delta enhanced by environmental engineering
1
2017
... (1)重大生态工程.人类不仅通过增加有机碳和营养物质对河流造成负荷, 而且通过大规模建造大坝深刻地改变了碳固定、矿化和沿着河流连续体储存有机碳之间的平衡(
Poised to prosper? A cross-system comparison of climate change effects on native and non-native species performance
1
2013
... 气候变化和人类活动为外来生物扩散和入侵提供了有利的途径, 未来气候变化和生物入侵可能相互作用成为全球生物多样性的主要威胁.近年来, 大量研究开始关注生物入侵如何改变淡水生态系统结构, 影响生物多样性以及生态系统服务功能(
A review of allochthonous organic matter dynamics and metabolism in streams
1
2010
... 全球土地利用正在以惊人的速度变化, 而且在未来很长一段时间内可能还会继续变化.与人类土地利用有关的对河流生态系统改变, 包括水文变化和沉积物再悬浮, 河岸植被组成, 大型无脊椎动物群落, 营养盐浓度和有机质动态变化等等(
Rapid loss of lakes on the Mongolian Plateau
1
2015
... 降水格局的变化导致干旱和半干旱地区的封闭流域成为全球无机碳汇.盆地终端湖泊中可能形成大量溶解无机碳的封存.几十年来, 由于人类活动和气候的密集化改变, 国际重要湖泊湿地的萎缩和干涸加剧.面积大于1 km2的湖泊从20世纪80年代后期的785个降低至2010年的577个, 其中中国内蒙古降低速率(34%)高于蒙古国(17.6%), 面积大于10 km2的湖泊数量同期也减少了30% (
Lakes and reservoirs as regulators of carbon cycling and climate
1
2009
... 内陆水体储存了陆地生态系统近一半的碳, 并参与碳的形态转化和气体释放, 对于全球碳循环发挥了重要作用(
Water scarcity hotspots travel downstream due to human interventions in the 20th and 21st century
1
2017
... (1)重大生态工程.人类不仅通过增加有机碳和营养物质对河流造成负荷, 而且通过大规模建造大坝深刻地改变了碳固定、矿化和沿着河流连续体储存有机碳之间的平衡(
Global threats to human water security and river biodiversity
2
2010
... 淡水生态系统作为地球系统的组成部分, 对自然和人文因素引起的地球系统结构和功能的变化极为敏感.淡水只占世界水资源的0.01%, 约占地球表面的0.8%, 但却维系着全球6%的已知生物物种 (
... 在淡水生态系统面临的一系列复杂威胁中, 气候变化是一个重要的压力来源.气候变化已经通过提高水温, 改变水流状况, 增加极端事件(如洪水、干旱、野火)和促进物种入侵, 对不同的淡水生态系统造成了压力.人们普遍预测在21世纪, 气候变化更会全面超越那些较为局部化的影响, 如人类活动造成的栖息地丧失和破碎化、富营养化和酸化等(
Flickering gives early warning signals of a critical transition to a eutrophic lake state
1
2012
... 由于淡水生态系统极度的多样化和复杂性, 且各个全球变化要素之间存在的交互作用, 使得淡水生态系统与全球变化研究中面临着诸多的挑战.(1)淡水生态系统特别容易受到气候变化的不同组成部分的影响, 但我们仍然对这些环境驱动因素及其与其他压力源的相互作用的后果了解有限, 特别是在更高的、多物种的、组织性的层次上.目前普遍采用的模拟实验方法具有较好的重复性, 能较直接地揭示气候对环境和生物产生影响的机理, 但无法完全模拟自然系统的复杂性, 而且由于对湖泊、湿地等在内的淡水生态系统地表过程认识不足, 尚不能全面揭示生态系统水平的响应特征.(2)全球变化研究要求野外观测可以实现多站点联网、基于统一的方法在不同站点开展长期的生态系统观测.对于高度多样化的淡水生态系统, 针对全球变化过程的联网研究尚处于摸索阶段.(3)生态系统响应全球变化的模式取决于系统本身的演化过程, 对事件发生的响应在短时间无法找到可侦测的表现, 只有通过长期演化过程的研究才能揭示其敏感性、抵抗力与弹性和突变的阈值等(
Global economic response to river floods
1
2018
... 气候变暖改变全球湖泊湖水混合特征.全球变暖导致湖泊冰雪覆盖层快速融化、湖面水温升高, 影响湖泊的混合特征.
Worldwide alteration of lake mixing regimes in response to climate change
1
2019
... 气候变暖改变全球湖泊湖水混合特征.全球变暖导致湖泊冰雪覆盖层快速融化、湖面水温升高, 影响湖泊的混合特征.
入侵生态学最新研究动态
1
2014
... 气候变化和人类活动为外来生物扩散和入侵提供了有利的途径, 未来气候变化和生物入侵可能相互作用成为全球生物多样性的主要威胁.近年来, 大量研究开始关注生物入侵如何改变淡水生态系统结构, 影响生物多样性以及生态系统服务功能(
入侵生态学最新研究动态
1
2014
... 气候变化和人类活动为外来生物扩散和入侵提供了有利的途径, 未来气候变化和生物入侵可能相互作用成为全球生物多样性的主要威胁.近年来, 大量研究开始关注生物入侵如何改变淡水生态系统结构, 影响生物多样性以及生态系统服务功能(
1
2017
... 全球变化是指由自然和人文因素引起的地球系统结构和功能的变化以及这些变化产生的影响, 其主要内容包括大气成分变化、全球气候变化、海平面上升、生物多样性以及生物入侵、人口增长、经济技术发展、土地利用和土地覆盖变化等(
1
2017
... 全球变化是指由自然和人文因素引起的地球系统结构和功能的变化以及这些变化产生的影响, 其主要内容包括大气成分变化、全球气候变化、海平面上升、生物多样性以及生物入侵、人口增长、经济技术发展、土地利用和土地覆盖变化等(
Long-term warming amplifies shifts in the carbon cycle of experimental ponds
1
2017
... 淡水生态系统温室气体由于来源和产生过程复杂, 估算和预测其规模都是很大的挑战.冰川融化导致的径流量增加使得陆源有机质在河流系统中的分解转化过程增强(

{kind=link}
{kind=link}
{kind=link}
{kind=link}