 ,, 朱淑霞, 李涌福, 伊贤贵, 段一凡, 王贤荣
,, 朱淑霞, 李涌福, 伊贤贵, 段一凡, 王贤荣 ,*南京林业大学南方现代林业协同创新中心, 生物与环境学院, 亚热带森林生物多样性保护国家林草局重点实验室, 南京 210037
,*南京林业大学南方现代林业协同创新中心, 生物与环境学院, 亚热带森林生物多样性保护国家林草局重点实验室, 南京 210037Leaf phenotypic variation in natural populations of Cerasus dielsiana
ZHU Hong,, ZHU Shu-Xia, LI Yong-Fu, YI Xian-Gui, DUAN Yi-Fan, WANG Xian-Rong,*Co-Innovation Center for Sustainable Forestry in Southern China, College of Biology and the Environment, Key Laboratory of State Forestry and Grassland Administration on Subtropical Forest Biodiversity Conservation, Nanjing Forestry University, Nanjing 210037, China通讯作者:
编委: 王国宏
责任编辑: 李敏
收稿日期:2018-08-10修回日期:2018-11-2网络出版日期:2018-12-20
| 基金资助: |
Received:2018-08-10Revised:2018-11-2Online:2018-12-20

摘要
关键词:
Abstract
Keywords:
PDF (1251KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
朱弘, 朱淑霞, 李涌福, 伊贤贵, 段一凡, 王贤荣. 尾叶樱桃天然种群叶表型性状变异研究. 植物生态学报[J], 2018, 42(12): 1168-1178 doi:10.17521/cjpe.2018.0196
ZHU Hong.
植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(方炎明和丁雨龙, 2016), 它主要研究同一物种不同种群在分布区内不同环境条件下的变异样式, 相较于细胞学、生化和分子标记手段, 具有测量简便、快速、经济的优点。数量分类学是开展植物表型研究最主要的手段, 即通过有效的样品采样、合理的数理统计方法, 揭示植物表型性状的变异大小和遗传规律, 客观地评价遗传多样性, 已经被许多****广泛应用于经济作物(Ali, 2018)、林木(江锡兵等, 2013; Li et al., 2018)、园艺品种(张往祥等, 2017)和入侵种(Wang et al., 2018)等研究中。其中植物叶表型性状不仅是植物分类学中最直观与可操作的分类性状, 同时也反映了植物对生长环境的响应和适应, 有助于认识环境筛选过滤对植物功能性状的作用以及由此产生的物种生物地理分布格局(Wright et al., 2007)。
尾叶樱桃(Cerasus dielsiana)隶属蔷薇科樱属, 为我国特有落叶小乔木, 主要分布在我国亚热带季风区的各省山区, 分布海拔500-1 400 m; 因其树形优美、早春先花后叶、花色多样, 具有极高的观赏价值, 加上生态适应性广、抗逆性强, 市场开发前景十分广阔(王贤荣, 2014)。由于尾叶樱桃分布区生境多样, 受局部地形及小气候的影响, 加上种间、种内存在潜在的自然杂交, 导致该物种不同地理种群间在区域尺度上产生了一定的形态分化, 目前尚无针对尾叶樱桃天然种群地理变异、叶表型多样性的系统报道。本文通过对区域尺度的尾叶樱桃天然分布区8个代表种群的叶表型性状多样性进行初步评估, 从形态学水平上揭示该种种群内和种群间地理变异样式, 探明叶表型性状与地理-气候因子的相关关系, 研究结果可为今后开展尾叶樱桃分类鉴定、种质资源收集保育与开发利用提供理论依据。
1 材料和方法
1.1 气象和地理资料收集
本研究的气候数据来源于世界气候数据库WorldClim (http://www.worldclim.org/), 主要是根据1950-2000年世界各地气象站的气候观测记录, 采用插值法生成的空间分辨率为2.5° (约5 km2)的全球气候数据包; 各种群的经度、纬度、海拔等地理信息由手持GPS仪(UNI-T UT379B, 优利德, 广东东莞)野外测定。利用DIVA-GIS软件(Hijmans et al., 2001)提取8个研究区对应的19个通用气候变量(bio1-bio19)。为减少各变量间共线性带来的干扰, 参照朱弘等(2017)的方法, 对所有环境变量做主成分分析(PCA)降维处理, 最终筛选出代表第1主成分(贡献率达97.459%)的4个变量, 即bio4、bio12、bio16和bio18作为影响尾叶樱桃种群分布的主导气候因子用于后续分析(表1)。Table 1
表1
表1主成分分析(PCA)筛选后影响尾叶樱桃的4个主导气候变量及其贡献率
Table 1
| 代码 Code | 气候变量 Climatic variables | 特征值 Eigenvalue | 作为第1主成分的贡献率 Variance contribution rate as first principal component (%) |
|---|---|---|---|
| bio4 | 气温季节变化 Temperature seasonality (SD × 100) | -0.151 | 97.459% |
| bio12 | 年降水量 Annual precipitation (mm) | 0.784 | |
| bio16 | 最湿季降水量 Precipitation of wettest quarter (mm) | 0.420 | |
| bio18 | 最暖季降水量 Precipitation of warmest quarter (mm) | 0.403 |
新窗口打开|下载CSV
1.2 材料来源与样品采集
表型性状测量材料为课题组在2014-2018年春季全面开展野外调查、种群采样基础上获得的凭证标本; 为了尽可能反映各分布区及其生态环境的特点, 自西向东选出代表核心分区的四川(峨眉山) 1个种群, 湖北(神农架、星斗山)、湖南(张家界、莽山)、江西(金竹飞瀑、仙姑坛)各2个种群和新记录的台湾(阿里山) 1个种群, 共计8个种群; 由于种群数量较小及取样困难, 每个种群随机选取5株发育良好的代表株, 单株间的水平距离尽量在30 m以上, 以减少亲缘关系的影响; 选择每个单株阳面中部以上完整无损的1个枝条作为凭证标本, 每份标本中再随机各取5枚完整无损的成龄叶片, 每个种群共计25枚。取样种群的地理位置及主导气候因子特征详见表2。Table 2
表2
表2尾叶樱桃8个天然种群的地理位置及主导气候因子特征
Table 2
| 种群 Population | 纬度 Latitude (N) | 经度 Longitude (E) | 平均海拔 Mean altitude (m) | 气温季节变化 Temperature Seasonality bio4 (SD × 100) | 年降水量 Annual precipitation bio12 (mm) | 最湿季降水量 Precipitation of wettest quarter bio16 (mm) | 最暖季降水量 Precipitation of warmest quarter bio18 (mm) |
|---|---|---|---|---|---|---|---|
| 四川峨眉山 Mt. Emei, Sichuan (EMS) | 29.57° | 103.37° | 1 365 | 7.53 | 1 264.00 | 92.13 | 710.00 |
| 湖北神农架 Shennongjia, Hubei (SNJ) | 31.45° | 110.25° | 860 | 8.15 | 1 696.00 | 51.61 | 557.00 |
| 湖北星斗山 Mt. Xingdou, Hubei (XDS) | 30.02° | 109.12° | 839 | 8.19 | 1 772.00 | 60.22 | 568.00 |
| 湖南莽山 Mt. Mangshan, Hunan (MS) | 24.97° | 112.88° | 1 228 | 7.66 | 1 492.00 | 56.44 | 601.00 |
| 湖南张家界 Zhangjiajie, Hunan (ZJJ) | 29.32° | 110.42° | 1 118 | 7.91 | 1 591.00 | 62.14 | 556.00 |
| 江西金竹飞瀑 Jinzhufeipu, Jiangxi (JZFP) | 27.00° | 115.92° | 966 | 7.20 | 1 508.00 | 60.26 | 609.00 |
| 江西仙姑坛 Xiangutan, Jiangxi (XGT) | 28.45° | 114.38° | 1 364 | 6.86 | 1 405.00 | 62.72 | 591.00 |
| 台湾阿里山 Mt. Ali, Taiwan (ALS) | 23.50° | 120.80° | 1 182 | 8.20 | 3 747.00 | 80.66 | 1 780.00 |
新窗口打开|下载CSV
1.3 叶表型性状的选取与测量
将尾叶樱桃8个种群的200枚叶片利用佳能Lide 120扫描仪(Canon, B?c Ninh, Vietnam)逐一进行扫描, 参考戴志聪等(2009)的方法对获得的电子图像利用ImageJ 1.52c软件(https://imagej.nih.gov/ij/download. ?html)测定。参考王贤荣等(2007)对樱属表型研究的方法, 共选取11个参数, 其中直接测定的指标有8个, 分别为叶片长(叶基至叶尖距离)、叶片宽(叶片最宽处距离)、叶柄长、叶面积、叶周长、一级侧脉数(到达锯齿顶端的左右两侧叶脉总数)、叶尖夹角和叶基夹角。再根据上述指标, 分别计算叶长宽比=叶片长/叶片宽、叶形指数= 4π ×叶面积/叶周长平方和、叶脉密度=一级侧脉数/2倍叶片长3个指标(李梅等, 2005; Meng et al., 2017)。1.4 数据统计分析方法
观测记载的数据在Excel 2007软件中进行整理汇总, 根据各性状25个样品的平均值($\bar{X}$)和标准偏差(SD)计算变异系数(CV =标准偏差/平均值); 采用SPSS 16.0软件中的单因素方差分析(显著性检验采用多重比较Duncan法), 研究叶表型性状在不同种群间的变异水平; 采用巢式方差设计(李斌等, 2002)进一步探讨比较尾叶樱桃叶表型11个性状在种群间和种群内的来源和分布状况, 其线性模型为: Yijk = μ + Si + T(i)j + e(ij)k。其中Yijk为第i个种群第j个单株第k个观测值; μ为总体均值; Si为种群间的效应(固定); T(i)j为种群内单株效应(随机); e(ij)k为随机误差。为了与遗传分化系数(Gst)相对应, 参照葛颂等(1988)的表型分化系数公式: Vst = δ2t/s/(δ2t/s + δ2s), 其中(Vst)为表型分化系数, 表示种群间变异占遗传总变异的比例, 其中δ2t/s为种群间方差分量, δ2s为种群内方差分量。再通过Person相关分析及单尾检验得出叶表型性状对地理气候因子的响应, 为保证数据符合正态分布和方差齐性, 先对原始数据进行标准化处理; 通过PCA, 得出不同表型性状对种群变异贡献率; 上述分析在SPSS 16.0软件中完成; 为了更直观地反映各种群遗传距离远近, 采用PAST3 3.2软件包(http://folk.uio.no/ohammer/past/)绘制数据标准化后的基于环境变量与各性状平均值的联合矩阵的主坐标分析(PCoA)图和采用非加权配对算术平均法(UPGMA)的聚类分析图, 其中靴带值验证(bootstrap test) 1 000次。2 结果和分析
2.1 尾叶樱桃叶表型性状变异特征
从尾叶樱桃叶表型性状在各种群间的Duncan多重比较可以看出, 叶片最宽、叶柄最长、叶面积最大、叶周长最长、密度最小的均来自四川峨眉山种群; 叶形指数最大、一级侧脉数最多、叶尖夹角最大、叶基夹角最大、叶长宽比最小的均来自台湾阿里山种群; 叶长次长、叶长宽比最长、叶基夹角次小的来自湖南莽山种群; 叶形指数最小、叶尖夹角最小、一级侧脉数最多的均来自江西仙姑坛种群; 叶宽最短、叶周长最短、叶尖夹角最小、叶脉密度最大的均来自江西金竹飞瀑种群; 各叶表型性状在种群间显著差异, 但总体表现为连续变异。11个叶表型性状平均CV为22.44%, CV最大的为叶面积(CV = 50.83%), 最小的为一级侧脉数(CV = 7.96%); 通过变异系数的比较, 可以发现11个叶表型性状在种群间的差异程度依次为: 叶面积>叶柄长>叶宽>叶脉密度>叶周长>叶长>叶尖夹角>叶长宽比>叶基夹角>叶形指数>一级侧脉数(表3)。Table 3
表3
表3尾叶樱桃8个天然种群11个叶表型性状的变异情况
Table 3
| 叶表型性状 Leaf phenotypic trait | 种群 Population | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| EMS | SNJ | XDS | MS | ZJJ | JZFP | XGT | ALS | 平均值 Mean | 变异系数 CV (%) | |
| 叶长 LL (mm) | 88.80 ± 22.85a | 79.08 ± 8.62b | 83.40 ± 17.18ab | 87.48 ± 11.27a | 89.63 ± 12.56a | 54.57 ± 12.41d | 62.87 ± 8.19c | 69.17 ± 9.14c | 76.94 | 23.61 |
| 叶宽 LW (mm) | 43.95 ± 8.73a | 37.15 ± 5.44bc | 33.90 ± 9.02c | 34.93 ± 4.96bc | 38.92 ± 8.12b | 22.64 ± 6.34e | 26.68 ± 4.44d | 37.43 ± 3.93bc | 34.45 | 26.39 |
| 叶长宽比 LWR | 2.01 ± 0.21c | 2.15 ± 0.19c | 2.50 ± 0.27ab | 2.53 ± 0.35a | 2.36 ± 0.38b | 2.44 ± 0.27ab | 2.37 ± 0.19b | 1.86 ± 0.14d | 2.28 | 14.93 |
| 叶柄长 PL (mm) | 11.84 ± 3.86a | 8.86 ± 1.49b | 7.59 ± 2.46cd | 11.10 ± 2.11a | 8.25 ± 1.60bc | 6.63 ± 1.35d | 8.53 ± 0.89bc | 7.54 ± 0.89cd | 8.80 | 29.92 |
| 叶面积 LA (mm2) | 2 746.99 ± 1 337.93a | 1 870.77 ± 476.40bc | 2 045.84 ± 1 040.24bc | 2 020.51 ± 516.34bc | 2 306.43 ± 829.25a | 890.91 ± 463.20f | 1 095.62 ± 295.07e | 1 826.66 ± 482.46d | 1 846.13 | 50.83 |
| 叶周长 LP (mm) | 213.94 ± 50.90a | 183.08 ± 21.64cd | 191.98 ± 44.07bc | 198.99 ± 24.00abc | 207.05 ± 32.44ab | 127.23 ± 29.26e | 146.15 ± 18.91d | 171.85 ± 22.61c | 179.81 | 23.77 |
| 叶形指数 LI | 0.73 ± 0.04b | 0.69 ± 0.05c | 0.65 ± 0.05d | 0.63 ± 0.06d | 0.65 ± 0.07d | 0.64 ± 0.06d | 0.63 ± 0.03d | 0.77 ± 0.03a | 0.67 | 8.90 |
| 一级侧脉数 PLV | 11.00 ± 0.46c | 12.00 ± 0.71b | 12.00 ± 1.32b | 12.00 ± 0.44b | 12.00 ± 0.70b | 12.00 ± 0.77b | 13.00 ± 0.46a | 13.00 ± 0.34a | 11.88 | 7.96 |
| 叶脉密度 VD | 0.06 ± 0.01d | 0.07 ± 0.01c | 0.07 ± 0.01c | 0.07 ± 0.01cd | 0.07 ± 0.03cd | 0.11 ± 0.01a | 0.10 ± 0.01b | 0.10 ± 0.01b | 0.08 | 24.42 |
| 叶尖夹角 AA (°) | 27.57 ± 3.42cd | 28.99 ± 3.5bc | 28.54 ± 4.36bc | 24.56 ± 2.79e | 31.02 ± 5.67b | 25.22 ± 7.02de | 22.99 ± 2.64e | 40.24 ± 4.46a | 28.64 | 22.87 |
| 叶基夹角 BA (°) | 114.47 ± 14.05b | 114.88 ± 11.04b | 113.87 ± 14.78b | 128.34 ± 8.24a | 103.60 ± 9.34c | 121.53 ± 17.54b | 116.68 ± 15.03b | 130.67 ± 13.33a | 116.72 | 13.31 |
| 平均值 Mean | - | - | - | - | - | - | - | - | - | 22.44 |
新窗口打开|下载CSV
利用巢氏方差对11个尾叶樱桃叶表型性状在种群间和种群内的差异检验结果表明, 除一级侧脉数和叶基夹角外, 其余在两个层次上均达到极显著差异(p < 0.01), 表明变异广泛(表4)。
Table 4
表4
表4尾叶樱桃8个天然种群11个叶表型性状的巢氏方差分析
Table 4
| 叶表型性状 Leaf phenotypic trait | 均方 Mean square | F值 F value | |||
|---|---|---|---|---|---|
| 种群间 Among polulations | 种群内 Within polulations | 随机误差 Random errors | 种群间 Among polulations | 种群内 Within polulations | |
| LL (mm) | 174.11 | 333.09 | 203.98 | 23.41** | 44.79** |
| LW (mm) | 41.10 | 83.58 | 47.29 | 27.00** | 54.91** |
| LWR | 0.16 | 0.12 | 0.07 | 21.94** | 16.46** |
| PL (mm) | 6.80 | 18.95 | 4.21 | 19.45** | 54.20** |
| LA (mm2) | 442 870.00 | 889 674.00 | 636 437.00 | 16.55** | 33.25** |
| LP (mm) | 262.46 | 1 858.26 | 1 160.49 | 22.60** | 117.18** |
| LI | 0.01 | 0.00 | 0.00 | 24.74** | 11.63** |
| PLV | 3.46 | 0.84 | 0.47 | 23.06** | 5.60 |
| VD | 0.00 | 0.00 | 0.00 | 43.74** | 55.25** |
| AA (°) | 30.31 | 42.98 | 19.13 | 39.61** | 56.17** |
| BA (°) | 905.20 | 227.54 | 159.85 | 10.97** | 2.76 |
新窗口打开|下载CSV
2.2 尾叶樱桃叶表型变异来源及种群间性状分化
巢氏方差分析结果表明, 种群间表型分化系数的变化范围是9.61%-66.69%,平均叶表型性状的分化系数为30.78%, 其中叶基夹角(66.69%)的表型分化系数最大, 其次为一级侧脉数(58.23%)与叶柄长(48.47%), 而叶尖夹角(9.61%)的表型分化系数最小; 11个叶表型性状在种群内和种群间的平均方差分量百分比分别为51.55%和22.55%, 种群内的方差分量远大于种群间的方差分量, 说明种群内的变异是尾叶樱桃叶表型变异的主要来源(表5)。Table 5
表5
表5尾叶樱桃叶表型性状的方差分量与表型分化系数
Table 5
| 叶表型性状 Leaf phenotypic trait | 方差分量 Variance component | 方差分量百分比 Percentage of variance component (%) | 表型分化系数 Phenotype differentiation coefficient (%) | ||||
|---|---|---|---|---|---|---|---|
| 种群间 Among populations | 种群内 Within populations | 随机误差 Random errors | 种群间 Among populations | 种群内 Within populations | 随机误差 Random Errors | ||
| LL (mm) | 6.359 | 25.822 | 13.324 | 13.98 | 56.75 | 29.28 | 19.76 |
| LW (mm) | 1.699 | 7.259 | 3.343 | 13.81 | 59.01 | 27.18 | 18.97 |
| LWR | 0.002 | 0.001 | 0.005 | 29.27 | 60.67 | 29.27 | 14.22 |
| PL (mm) | 0.486 | 0.517 | 0.272 | 38.13 | 40.54 | 21.33 | 48.47 |
| LA (mm2) | 17 872.160 | 50 647.400 | 35 586.960 | 17.17 | 48.65 | 34.18 | 26.08 |
| LP (mm) | 63.832 | 139.554 | 74.330 | 22.98 | 50.25 | 26.76 | 31.38 |
| LI | 0.000 | 0.001 | 0.000 | 22.49 | 55.32 | 22.19 | 28.91 |
| PLV | 0.105 | 0.075 | 0.034 | 15.74 | 35.19 | 15.74 | 58.23 |
| VD | 0.000 | 0.000 | 0.000 | 12.69 | 65.29 | 22.02 | 16.27 |
| AA (°) | 0.507 | 4.771 | 1.719 | 7.25 | 68.18 | 24.57 | 9.61 |
| BA (°) | 22.105 | 13.537 | 9.101 | 54.49 | 27.21 | 18.30 | 66.69 |
| 平均值 Mean | - | - | - | 22.55 | 51.55 | 24.62 | 30.78 |
新窗口打开|下载CSV
2.3 尾叶樱桃叶表型性状的主成分分析
主成分分析结果表明, 11个叶表型性状中的前3个主成分方差贡献率分别为73.242%、11.319%和7.839%, 累计贡献率达到92.400%, 能够代表叶表型特征变量的绝大部分信息; 第1主成分主要表征为“大小性状”, 包含叶面积、叶宽、叶周长、叶长和叶脉密度; 第2和第3主成分主要表征为“形状性状”, 分别包含叶柄长和叶尖夹角, 叶长宽比、叶形指数和叶基夹角(表6)。Table 6
表6
表6尾叶樱桃叶表型性状的主成分分析
Table 6
| 叶表型性状 Leaf phenotypic trait | 主成分 Main components | 叶表型性状 Leaf phenotypic trait | 主成分 Main components | ||||
|---|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | ||
| LL (mm) | 0.312 | -0.093 | -0.268 | PLV | -0.020 | 0.071 | 0.021 |
| LW (mm) | 0.367 | 0.149 | 0.163 | VD | -0.332 | 0.182 | 0.302 |
| LWR | -0.055 | -0.242 | -0.431 | AA (°) | 0.109 | 0.604 | 0.244 |
| PL (mm) | 0.186 | -0.667 | 0.650 | BA (°) | 0.022 | 0.102 | 0.242 |
| LA (mm2) | 0.706 | 0.117 | -0.027 | 特征值 Eigenvalue | 0.092 | 0.015 | 0.011 |
| LP (mm) | 0.327 | -0.039 | -0.139 | 贡献率 Variance contribution rate (%) | 73.242 | 11.319 | 7.839 |
| LI | 0.051 | 0.195 | 0.251 | 累计贡献率 Cumulative Contribution rate (%) | 73.242 | 84.561 | 92.400 |
新窗口打开|下载CSV
2.4 尾叶樱桃叶表型性状与地理、气候因子间的相关性
进一步对尾叶樱桃叶表型与采集地对应的地理、气候因子做了相关性分析, 结果表明: 经度与尾叶樱桃叶宽(r = -0.641)、叶面积(r = -0.658)呈显著负相关关系, 而与一级侧脉数(r = 0.659)呈显著正相关关系; 气温季节变化(bio4)分别与叶脉密度(r = 0.615)和叶尖夹角(r = 0.809)呈显著或极显著正相关关系, 与叶长(r = -0.631)呈显著负相关关系; 最湿季降水量(bio16)与叶长宽比(r = -0.600)和叶形指数(r = 0.651)呈显著负相关和正相关关系(表7)。Table 7
表7
表7尾叶樱桃叶表型性状与地理、气候因子的Person相关性分析
Table 7
| 叶表型性状 Leaf phenotypic trait | 纬度 Latitude (N) | 经度 Longitude (E) | 海拔 Altitude (m) | 气温季节变化Temperature seasonality bio4 (SD × 100) | 年降水量 Annual precipitation bio12 (mm) | 最湿季降水量 Precipitation of wettest quarter bio16 (mm) | 最暖季降水量 Precipitation of warmest quarter bio18 (mm) |
|---|---|---|---|---|---|---|---|
| LL (mm) | 0.163 | -0.554 | 0.091 | -0.613* | -0.008 | 0.221 | -0.017 |
| LW (mm) | 0.284 | -0.641* | 0.176 | 0.545 | 0.002 | 0.443 | 0.057 |
| LWR | -0.363 | 0.431 | -0.260 | -0.098 | -0.028 | -0.600* | -0.175 |
| PL (mm) | -0.094 | -0.469 | 0.430 | 0.006 | -0.151 | 0.482 | 0.040 |
| LA (mm2) | 0.227 | -0.658* | 0.162 | 0.551 | -0.016 | 0.459 | 0.037 |
| LP (mm) | 0.184 | -0.593 | 0.141 | 0.585 | -0.009 | 0.315 | 0.008 |
| LI | 0.347 | -0.535 | 0.093 | 0.340 | 0.084 | 0.651* | 0.206 |
| PLV | -0.248 | 0.659* | 0.087 | -0.395 | 0.118 | -0.499 | 0.017 |
| VD | -0.220 | 0.590 | -0.035 | 0.651* | 0.007 | -0.222 | 0.018 |
| AA (°) | 0.324 | -0.249 | -0.429 | 0.809** | 0.303 | 0.127 | 0.181 |
| BA (°) | -0.478 | 0.168 | 0.277 | -0.090 | 0.059 | -0.075 | 0.121 |
新窗口打开|下载CSV
2.5 基于尾叶樱桃叶表型性状和地理气候因子的主坐标分析与聚类分析
为了直观地显示尾叶樱桃种群间的亲缘关系, 选择尾叶樱桃8个种群代表前3个主成分的10个叶表型性状联合对应的地理-气候因子, 进行主PCoA, 散点图分布结果与种群所在的实际地理位置有着良好的对应关系(图1A)。进一步利用欧式平均距离, 采用UPGMA法, 对尾叶樱桃8个种群进行聚类分析(图1B)。在欧式距离500-1 000阈值间, 自上而下可将8个种群分为4支: 四川峨眉山为第1分支; 湖北神农架、湖北星斗山、湖南莽山和湖南张家界构成第2分支; 江西金竹飞瀑和仙姑坛构成第3分支; 台湾阿里山为第4分支。图1
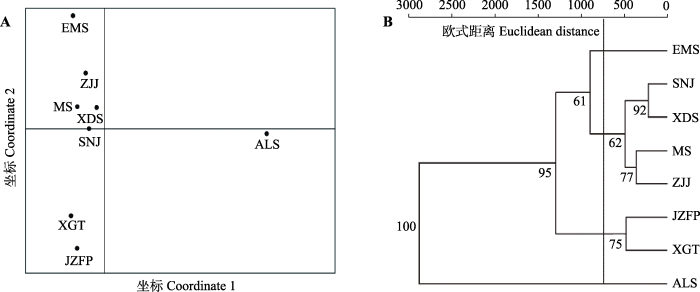 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1基于叶表型性状和地理-气候因子的8个尾叶樱桃天然种群关系示意图。A, 主坐标分析输出结果。B, 非加权配对算术平均法聚类分析输出结果。ALS, 台湾阿里山; EMS, 四川峨眉山; JZFP, 江西金竹飞瀑; MS, 湖南莽山; SNJ, 湖北神农架; XDS, 湖北星斗山; XGT, 江西仙姑坛; ZJJ, 湖南张家界。
Fig. 1Relationships between leaf phenotypic traits and geographic-climatic factors of the eight natural populations of Cerasus dielsiana. A, Output from principal coordinate analysis. B, Output from unweighted pair-group method with arithmetic mean cluster analysis. ALS, Mt. Ali, Taiwan. EMS, Mt. Emei, Sichuan; JZFP, Jinzhufeipu, Jiangxi; MS, Mt. Mangshan, Hunan; SNJ, Shennongjia, Hubei; XDS, Mt. Xingdou; Hubei; XGT, Xiangutan, Jiangxi; ZJJ, Zhangjiajie, Hunan.
3 结论和讨论
3.1 尾叶樱桃叶表型变异的多样性
植物表型多样性是遗传适应性和环境异质性长期共同作用的结果, 植物表型变异程度越丰富, 越能提高在不同环境下的适应程度(Verbeeck et al., 2014; Saenger & West, 2018)。鉴于叶表型性状是最直观的外在表现, 本研究基于尾叶樱桃天然分布区采集的蜡叶标本进行统计分析, 大部分叶表型性状在种群间和种群内的差异达到极显著(p < 0.01)水平, 体现了其叶片表型地理变异的丰富性; 变异系数最大的为叶面积(CV = 50.83%), 表明其性状可塑性最大, 易受到个体发育和环境的双重影响; 变异系数最小的为一级侧脉数(CV = 7.96%), 方差分析在种群间达到极显著差异而在种群内未达到显著性差异, 说明其在遗传上兼具种群间的变异性与物种间的稳定性, 例如, 尾叶樱桃一级侧脉数平均在12对左右(四川峨眉山最少有11对, 台湾阿里山和湖南莽山的侧脉最多均有13对), 在每个种群内部都是比较稳定的, 与其他樱属植物——例如迎春樱桃(C. discoidea)有8-10对, 华中樱桃(C. conradinae)有7-9对, 存在数量上的间断分布, 因此, 可以作为一个樱属数量分类的重要参考指标。主成分分析结果(表5)进一步说明, 在尾叶樱桃叶表型形态分化中, “大小性状”尤其是叶面积(特征值为0.706)起主要贡献作用, 也验证了变异系数的结果。从多重比较结果(表3)不难看到, 不同种群的尾叶樱桃叶表型均有一定差异, 但这些差异在数量性状上均表现出一定的连续性。3.2 尾叶樱桃叶表型变异的来源与分化程度
在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(葛颂等, 1988)。11个尾叶樱桃种群平均叶表型性状分化系数为30.78%, 低于无患子(Sapindus saponaria)(刁松锋等, 2014, 62.21%)、长柄扁桃(Amygdalus pedunculata)(柳江群等, 2017, 45.90%)、浙江柿(Diospyros glaucifolia)(井振华等, 2010, 40.15%)、水青树(Tetracentron sinense)(李珊等, 2016, 46.69%)、白檀(Symplocos paniculata)(杨艳, 2015, 59.16%)、浙江楠(Phoebe chekiangensis)(陆云峰等, 2018, 41.43%)等, 高于青钱柳(Cyclocarya paliurus)(佘诚棋等, 2009, 20.54%)、印楝(Azadirachta indica)(彭兴民等, 2012, 11.89%)等, 与上述物种相比, 由于分布碎片化和地理隔离导致尾叶樱桃种群间的叶表型分化处于中等水平; 种群间的变异能更加真实地反映地理与生殖隔离上的差异, 也是种内多样性的重要组成部分(庞广昌和姜冬梅, 1995)。其中, 尾叶樱桃叶基夹角在各性状中的分化系数(66.69%)最高, 种群间的方差分量百分比(54.49%)是种群内(27.21%)的2倍, 说明其变异在种群间占优势; 而叶尖夹角的分化系数最低, 反映其“尾尖”的特性最稳定; 叶表型种群内的平均方差分量百分比(51.55%)高于种群间的平均方差分量百分比(22.55%), 反映了种群内是变异的主要来源, 这主要与其自身繁殖特性和基因流有关, 尾叶樱桃依靠虫媒近距离传粉为主, 种群内杂交频繁, 加上自花繁殖的能力强, 种子结实量大, 种群更新以实生苗为主, 植株抗逆性强, 寿命长, 均有利于遗传变异在种群中的保存; 而以果实为食的鸟兽与河流是其种子扩散的长距离方式, 理论上有利于基因流的扩散, 但通过实地观察, 尾叶樱桃种子通常在离开母体后常温下1周左右就会逐渐丧生活能力, 其实际的有效传播距离可能小于种群间的地理距离, 使得种群间种子交流较少, 基于SSR分子标记对近缘樱属植物的研究结果(陈娇等, 2013)也得到相似的结果。3.3 尾叶樱桃叶表型性状对地理、气候因子的响应
植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(Wright et al., 2007)。通过在较大尺度上针对特定植物类群基于种群水平的表型变异的分析, 研究表型性状与地理-气候间的关系, 往往可以更准确地揭示植物变异格局及其生态响应策略(杨贺雨等, 2016; Meng et al., 2017)。相关性分析结果(表7)表明, 尾叶樱桃叶宽(r = -0.641)、叶面积(r = -0.658)和一级侧脉数(r = 0.659)在地理空间上呈现以经度为主的单一变异模式, 即自西向东随经度的增加, 8个种群呈现叶片变窄、叶面积变小和一级侧脉数增多的趋势。这种在北半球随海洋向内陆的过渡中引起的地理经向变异规律也在麻栎(Quercus acutissima)(刘志龙等, 2011)、米老排(Mytilaria laosensis)(袁洁等, 2013)、红椿(Toona ciliata)(汪洋等, 2018)等物种中有所报道, 通常反映了水分和温度的梯度变化(Qian et al., 1998); 与此同时, 尾叶樱桃叶表型性状与气候因子中的气温季节变化(bio4)和最湿季降水量(bio16)之间存在统计上的相关性, 而与年降水量(bio12)和最暖季降水量(bio18)无显著关系, 亦表明“气温变异性”与“展叶期降水量”是驱动尾叶樱桃叶表型性状变异的主要因子, 具体来说: 温差越大, 叶长越小、叶尖夹角越大而叶脉密度越高; 展叶期降水量越大, 叶形越宽; 此外叶表型性状与纬度和海拔没有达到显著相关性, 这可能受局部环境条件如土壤、坡度、坡向等影响更大(杜会聪等, 2018), 有待日后从小尺度上开展研究。3.4 尾叶樱桃叶表型性状变异的适应性机制
植物形态特征变异往往具有适应性意义, 广阔分布区、多样的生境与气候异质性, 为研究尾叶樱桃叶表型地理变异模式提供了可能。从种群生态学的角度看, 本研究选取的8个代表种群分布于我国南方长江流域, 跨度范围为23.50°-31.45° N, 103.37°-120.80° E, 平均海拔范围839-1 365 m, 其中最西缘峨眉山种群属于横断山脉东部边缘的邛崃山系, 平均海拔1 365 m, 在各种群中海拔分布最高; 而最东南缘的台湾种群, 与祖国大陆隔海相望, 所在生境受亚热带季风气候影响(姜超等, 2017)程度最大, 与高山、海峡等天然屏障共同加剧了局域生境的隔离程度; 同时这也是尾叶樱桃叶各功能性状相互协调的结果, 从全球尺度到区域尺度, 叶片大小和结构与温度、水分、风速等气候条件之间存在显著的相互关系, 譬如: 叶大小的形成机制与适应价值普遍被认为是与叶的能量平衡变化有关(Niinemets et al., 2010); 应用热成像技术和图像分析技术对植物叶片样地数据定量研究显示, 在高温、低风速环境下, 叶片宽度每减少1 cm, 叶片表面温度降低约2.1 ℃ (李永华等, 2018); 植物叶片变小、变窄将有利于削弱自身的边界层阻力, 增强叶边缘对水热与CO2的传导能力, 进而降低呼吸和蒸腾的成本, 使得相对小的叶片更利于生长在热、干、高光和低养分的环境下(McDonald et al., 2003); 同时, 作为叶片中的维管束组织和木质部成分, 叶脉具有水分运输和支撑叶片的双重作用, 因此其密度随温度的升高和光照强度的增加而增加, 以适应叶片蒸腾作用的增强, 随降水的增加而增加, 以提高叶片的抗打击能力(李东胜等, 2013)。综上所述, 我们认为尾叶樱桃采取减小叶宽度, 缩小叶面积, 增加叶脉密度的关联组合策略以适应亚热带东部地区夏季强烈的高温高湿、强风降水与剧烈的干湿交替的气候特征。
3.5 尾叶樱桃天然种群保护与开发利用的建议与展望
尾叶樱桃作为优良的国产樱属代表性野生植物, 颇具观赏价值, 其市场潜在价值近年来已经逐渐被人们所认识。然而当前尾叶樱桃天然种群正不断受到人类经济活动的影响(如城市化、旅游开发), 面临生境遭到破坏、资源减少的威胁; 就本研究而言, 8个尾叶樱桃代表种群均位于人为干扰较少的自然保护区或保护较好的景区, 其他地区发现天然种群的概率越来越低。由于种群内变异是尾叶樱桃种群变异的主要来源, 在开展种质资源收集、迁地或就地保存与遗传改良时, 可适当减少抽样种群个体数, 增加核心种群内的取样密度, 尤其是湖北、湖南、江西三省的种群, 以增加遗传变异的丰富性; 同时, 种群间又存在中等程度的叶表型分化, 不同层次的变异为优质种质资源和生物多样性保护提供了物质基础(杨晓霞等, 2016; Humphrey et al., 2018)。结合地理位置、主坐标与聚类分析结果看, 四川峨眉山与台湾阿里山种群均远离尾叶樱桃多样性分布的中心, 基因交流受限, 明显分化为两个独立分支, 值得一提的是, 后者在中国植物志中英文版本中均未记录, 为笔者近年来调查偶然发现, 属于尾叶樱桃分布新记录。在种子资源就地保护或开发时, 也应该予以充分重视, 通过采种播种、采穗扦插方式, 加强特征种群野生种质选择和收集。尾叶樱桃天然种群表型地理变异格局上的差异是由其树种特性以及适应性进化历史等因素所决定的, 从形态学水平上对遗传变异进行探索, 但尚需整合生态地理学(生态位模型)或分子亲缘地理学(谱系地理)等多方面的研究, 以更深层次验证并阐明遗传变异格局的形成机制, 确定保护单元, 有效的制订保护策略。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
URL [本文引用: 1]

采用SSR分子标记技术对四川野生中国樱桃5个居群共133株的遗传多样性水平及居群的遗传结构进行了研究。结果显示:10对SSR引物共检测到78个等位基因,平均每位点等位基因7.8个。Nei’s基因多样性指数(H)为0.6112~0.6689,Shannon’s信息指数(I)为1.1984~1.3786。基于分子方差分析(AMOVA),92.53%的变异来自居群内,7.47%的遗传变异来自于居群间。居群间遗传距离(GD0.2416)、遗传一致度(GI0.7854)、遗传分化指数(Fst=0.0844)以及较强的基因流(Nm=2.7125)均表明居群间的遗传分化水平较低,居群内存在显著近交现象(Fis=0.3986),且居群在大多数位点上偏离Hardy-Weinberg平衡。基于上述结果,分析讨论了居群较高遗传多样性和居群间较低遗传分化形成的可能原因,并提出野生中国樱桃的保护利用策略。
URL [本文引用: 1]
采用SSR分子标记技术对四川野生中国樱桃5个居群共133株的遗传多样性水平及居群的遗传结构进行了研究。结果显示:10对SSR引物共检测到78个等位基因,平均每位点等位基因7.8个。Nei’s基因多样性指数(H)为0.6112~0.6689,Shannon’s信息指数(I)为1.1984~1.3786。基于分子方差分析(AMOVA),92.53%的变异来自居群内,7.47%的遗传变异来自于居群间。居群间遗传距离(GD0.2416)、遗传一致度(GI0.7854)、遗传分化指数(Fst=0.0844)以及较强的基因流(Nm=2.7125)均表明居群间的遗传分化水平较低,居群内存在显著近交现象(Fis=0.3986),且居群在大多数位点上偏离Hardy-Weinberg平衡。基于上述结果,分析讨论了居群较高遗传多样性和居群间较低遗传分化形成的可能原因,并提出野生中国樱桃的保护利用策略。
DOI:10.3969/j.issn.1000-3142.2009.03.013URL [本文引用: 1]
传统的纸样称重法用来测量离体叶片的面积,烦琐、耗时、精度不高.为寻求一种适合的方法,我们对离体叶片采用扫描仪获取叶片的数字图像,利用Image J软件测量叶片的长、宽、周长、面积及叶柄的长,并与传统的纸样称重测定叶面积法进行比较.结果表明,此方法具有低成本、快速、精确等特点,适用于植物形态学、植物生理生态学、林学及农业等对叶片形态特征的测量研究工作.
DOI:10.3969/j.issn.1000-3142.2009.03.013URL [本文引用: 1]
传统的纸样称重法用来测量离体叶片的面积,烦琐、耗时、精度不高.为寻求一种适合的方法,我们对离体叶片采用扫描仪获取叶片的数字图像,利用Image J软件测量叶片的长、宽、周长、面积及叶柄的长,并与传统的纸样称重测定叶面积法进行比较.结果表明,此方法具有低成本、快速、精确等特点,适用于植物形态学、植物生理生态学、林学及农业等对叶片形态特征的测量研究工作.
DOI:10.5846/stxb201306211756URL [本文引用: 1]
无患子(Sapindus mukorossi Gaertn.)是我国长江以南地区传统的重要绿化树种,其果皮富含皂苷,种仁富含油脂,是国家林业局审定的新型木本油料树种之一。为揭示无患子群体间和 群体内种实表型性状变异式样,采用单因素方差分析、巢式方差分析、相关分析和主成分分析等方法,对无患子13个天然群体的10个种实表型性状进行比较分 析,研究其群体间和群体内种实表型多样性以及表型变异与地理生态因子间的相关性。结果表明:无患子种实表型性状变异系数平均为7.34%,在群体间和群体 内均存在丰富的表型变异。表型性状分化系数平均为62.21%,群体间变异(39.93%)大于群体内(27.46%),是无患子种实表型性状变异的主要 来源。多数性状在群体间差异显著,不同程度的表现出边缘群体易于分化的特点,但地理变化规律不连续,而在群体内不同性状的差异性亦不一致。种子形态受群体 地理生态的影响较果实形态大,西北部群体种子趋于椭圆形,东部、南部则趋于圆球形;地理纬度、年平均气温与多数种实性状间呈显著相关,是无患子种实表型性 状变异的主要地理生态影响因子。可见,无患子种实表型性状在群体间、群体内变异都较为丰富,这些变异是由遗传和环境因素共同作用的结果。群体间和群体内多 层次的变异为无患子优良种质资源保育和利用提供了物质基础。
DOI:10.5846/stxb201306211756URL [本文引用: 1]
无患子(Sapindus mukorossi Gaertn.)是我国长江以南地区传统的重要绿化树种,其果皮富含皂苷,种仁富含油脂,是国家林业局审定的新型木本油料树种之一。为揭示无患子群体间和 群体内种实表型性状变异式样,采用单因素方差分析、巢式方差分析、相关分析和主成分分析等方法,对无患子13个天然群体的10个种实表型性状进行比较分 析,研究其群体间和群体内种实表型多样性以及表型变异与地理生态因子间的相关性。结果表明:无患子种实表型性状变异系数平均为7.34%,在群体间和群体 内均存在丰富的表型变异。表型性状分化系数平均为62.21%,群体间变异(39.93%)大于群体内(27.46%),是无患子种实表型性状变异的主要 来源。多数性状在群体间差异显著,不同程度的表现出边缘群体易于分化的特点,但地理变化规律不连续,而在群体内不同性状的差异性亦不一致。种子形态受群体 地理生态的影响较果实形态大,西北部群体种子趋于椭圆形,东部、南部则趋于圆球形;地理纬度、年平均气温与多数种实性状间呈显著相关,是无患子种实表型性 状变异的主要地理生态影响因子。可见,无患子种实表型性状在群体间、群体内变异都较为丰富,这些变异是由遗传和环境因素共同作用的结果。群体间和群体内多 层次的变异为无患子优良种质资源保育和利用提供了物质基础。
URL [本文引用: 1]
测定了浙江省(富阳的龙王坎、碧东山和五尖山以及临安的方山)4个蜡梅[Chimonanthus praecox(Linn.)Link]野生种群种实11个表型性状及土壤元素含量和p H值;在此基础上,对表型性状进行相关性分析和主成分分析,对供试种群进行聚类分析,并进行了表型性状与土壤相关因子的灰色关联度分析。结果表明:在11个表型性状中,除种子千粒质量外,种群间其他表型性状均有显著或极显著差异,其中,富阳碧东山种群的多项指标值均最大;各表型性状的变异系数差异明显(6.97%~45.72%,均值为19.98%),按变异程度由大到小依次排序为单果种子数、单果质量、单果果壳质量、果实长、果实宽、果形系数、果口直径、种形系数、种子宽、种子长、种子千粒质量;种群间表型分化系数均值为14.83%;11个表型性状中,种形系数和种子千粒质量与其他表型性状的相关性均较小,而果实长、果实宽、果口直径、单果质量、单果果壳质量和单果种子数间均存在极显著相关性。4个种群的土壤元素含量差异明显,土壤均呈弱碱性;各表型性状与同一土壤因子的关联度系数差异较小,但不同土壤因子与同一表型性状的关联度系数差异明显,关联度系数均值从大到小依次为p H值、Na含量、K含量、Cu含量、Mn含量、Ca含量、Mg含量、N含量、Fe含量、P含量、Zn含量。主成分分析结果表明:单果果壳质量、果实宽、单果质量、果实长、种子长、种形系数和种子千粒质量对蜡梅野生种群种实表型性状变异起主要作用。通过聚类分析,可将供试4个蜡梅野生种群分为3组,其中,临安方山和富阳龙王坎2个种群归为一组,另2个种群各自独立成组,聚类分析结果与种群间的地理距离无关。研究结果表明:浙江省蜡梅野生种群种实表型性状变异丰富,以种群内变异占主导地位,遗传特性是其表型性状变异的主要因子,且表型性状变异与多个土壤因子的综合作用相关。
URL [本文引用: 1]
测定了浙江省(富阳的龙王坎、碧东山和五尖山以及临安的方山)4个蜡梅[Chimonanthus praecox(Linn.)Link]野生种群种实11个表型性状及土壤元素含量和p H值;在此基础上,对表型性状进行相关性分析和主成分分析,对供试种群进行聚类分析,并进行了表型性状与土壤相关因子的灰色关联度分析。结果表明:在11个表型性状中,除种子千粒质量外,种群间其他表型性状均有显著或极显著差异,其中,富阳碧东山种群的多项指标值均最大;各表型性状的变异系数差异明显(6.97%~45.72%,均值为19.98%),按变异程度由大到小依次排序为单果种子数、单果质量、单果果壳质量、果实长、果实宽、果形系数、果口直径、种形系数、种子宽、种子长、种子千粒质量;种群间表型分化系数均值为14.83%;11个表型性状中,种形系数和种子千粒质量与其他表型性状的相关性均较小,而果实长、果实宽、果口直径、单果质量、单果果壳质量和单果种子数间均存在极显著相关性。4个种群的土壤元素含量差异明显,土壤均呈弱碱性;各表型性状与同一土壤因子的关联度系数差异较小,但不同土壤因子与同一表型性状的关联度系数差异明显,关联度系数均值从大到小依次为p H值、Na含量、K含量、Cu含量、Mn含量、Ca含量、Mg含量、N含量、Fe含量、P含量、Zn含量。主成分分析结果表明:单果果壳质量、果实宽、单果质量、果实长、种子长、种形系数和种子千粒质量对蜡梅野生种群种实表型性状变异起主要作用。通过聚类分析,可将供试4个蜡梅野生种群分为3组,其中,临安方山和富阳龙王坎2个种群归为一组,另2个种群各自独立成组,聚类分析结果与种群间的地理距离无关。研究结果表明:浙江省蜡梅野生种群种实表型性状变异丰富,以种群内变异占主导地位,遗传特性是其表型性状变异的主要因子,且表型性状变异与多个土壤因子的综合作用相关。
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 2]
本文应用同工酶分析技术结合形态、生长和物候等表型性状,分析了福建省沙县境内的5个马尾松天然小群体的遗传结构。结果表明,马尾松群体具有较高的遗传变异水平,5个小群体的多态位点百分率达64.5%,等位基因平均数为1.65,平均期望杂合度为0.216。小群体间的分化程度不高,5个小群体的平均遗传距离为0.0047;同工酶测定的总变异中,2.4%来自小群体间,其余的变异(97.6%)来自小群体内;而表型性状总的遗传变异中,6.5%左右来自小群体间,其余的变异(93.5%)来自小群体内。
URL [本文引用: 2]
本文应用同工酶分析技术结合形态、生长和物候等表型性状,分析了福建省沙县境内的5个马尾松天然小群体的遗传结构。结果表明,马尾松群体具有较高的遗传变异水平,5个小群体的多态位点百分率达64.5%,等位基因平均数为1.65,平均期望杂合度为0.216。小群体间的分化程度不高,5个小群体的平均遗传距离为0.0047;同工酶测定的总变异中,2.4%来自小群体间,其余的变异(97.6%)来自小群体内;而表型性状总的遗传变异中,6.5%左右来自小群体间,其余的变异(93.5%)来自小群体内。
[本文引用: 1]
DOI:10.1007/s00442-018-4116-4URLPMID:29603095 [本文引用: 1]
The biotic and the abiotic environment play a major role in shaping plant phenotypes and their geographic distributions. However, little is known about the extent to which plant phenotypes match local patterns of herbivory across fine-grained habitat mosaics, despite the strong effect of herbivory on plant fitness. Through a reciprocal transplant-common garden experiment with clonally propagated rhizomes, we tested for local phenotypic differentiation in bittercress (Brassicaceae: Cardamine cordifolia ) plants collected across an ecotonal habitat mosaic. We found that bittercress in sunny meadows (high herbivory) and shaded understories (low herbivory) have diverged in heritable growth and herbivore resistance phenotypes. The expression of these differences was habitat dependent, mirroring patterns of adaptive divergence in phenotypic plasticity between plant populations in meadow and understory habitats at broader geographic scales, and showed no evidence for a constraint imposed by growth-defense tradeoffs. Most notably, plants derived from shade habitats exhibited a weaker shade-induced elongation response (i.e., shade avoidance syndrome, SAS) and reduced resistance to herbivory, relative to plants derived from sun habitats, when both were grown in shade common gardens. Greenhouse experiments revealed that divergent SAS phenotypes in shade conditions were expressed in offspring grown from seed as well. Finally, we observed partially non-overlapping flowering phenology between habitat-types in the field, which may be at least one factor that helps to reinforce habitat-specific phenotypic divergence. Altogether, our study illuminates how a native plant may cope with overlapping biotic and abiotic stressors across a fine-grained habitat mosaic.
DOI:10.17521/cjpe.2017.0070URL [本文引用: 1]
亚洲是全球季风气候最典型的区域,也是热带植物分布距离赤道最远的地区。揭示季风对热带植物迁移与进化的影响规律,有利于深入认识东亚植被分布和生物多样性形成与维持机制。亚洲地区夏季盛行东亚季风、南亚季风和西北太平洋季风。历史上,季风盛行的时间与早期被子植物在东南亚群岛、华夏古陆起源的时间大致吻合,季风可能促进了被子植物的快速分化与扩散。季风是热带植物得以向北扩散到我国滇黔桂交界区和雅鲁藏布江河谷的根本原因,并导致了热带季节性雨林、热带季雨林、干旱河谷稀树灌丛或草原、海南岛西部滨海稀树草原等特殊植被的形成。亚洲的三大夏季风在高山纵横、大河奔流和石灰岩地貌密布的中国西南与中南半岛一带交汇、叠加,使之成为一些典型热带类群的物种多样与特有种分布中心。这可能是中国.缅甸生物多样性热点地区形成与维持的一个重要原因。随着全球气候变暖,季风可能促进热带植物的进一步北迁,增加中国南方植物区系的热带植物成分。
DOI:10.17521/cjpe.2017.0070URL [本文引用: 1]
亚洲是全球季风气候最典型的区域,也是热带植物分布距离赤道最远的地区。揭示季风对热带植物迁移与进化的影响规律,有利于深入认识东亚植被分布和生物多样性形成与维持机制。亚洲地区夏季盛行东亚季风、南亚季风和西北太平洋季风。历史上,季风盛行的时间与早期被子植物在东南亚群岛、华夏古陆起源的时间大致吻合,季风可能促进了被子植物的快速分化与扩散。季风是热带植物得以向北扩散到我国滇黔桂交界区和雅鲁藏布江河谷的根本原因,并导致了热带季节性雨林、热带季雨林、干旱河谷稀树灌丛或草原、海南岛西部滨海稀树草原等特殊植被的形成。亚洲的三大夏季风在高山纵横、大河奔流和石灰岩地貌密布的中国西南与中南半岛一带交汇、叠加,使之成为一些典型热带类群的物种多样与特有种分布中心。这可能是中国.缅甸生物多样性热点地区形成与维持的一个重要原因。随着全球气候变暖,季风可能促进热带植物的进一步北迁,增加中国南方植物区系的热带植物成分。
.
DOI:10.3969/j.issn.1001-1498.2013.01.019URL [本文引用: 1]
Idesia polycarpa is an important woody oil and biomass energy tree species in China, the development of germplasm resources and research of genetic variation on I. polycarpa are of great significance. The phenotypic traits were investigated and determined, and variation among and within populations were analyzed on nine natural groups in the distribution zone of I. polycarpa. The results indicated that there were significant differences in age, growth and yield traits of I. polycarpa from different population, especially in yield. There were rich variations in fifteen phenotypic traits including leaf length. These variation appeared among and within populations, and with similar contribution rate (31.99% and 30.82%). The variation of fruit traits were much more obvious in phenotypes, and the variation coefficients such as number of fruits per grain, which were closely related to yield, were up to 38%. The rich genetic variation of I. polycarpa provides many advantages for the development and utilization of germplasm resources, selecting and breeding.
DOI:10.3969/j.issn.1001-1498.2013.01.019URL [本文引用: 1]
Idesia polycarpa is an important woody oil and biomass energy tree species in China, the development of germplasm resources and research of genetic variation on I. polycarpa are of great significance. The phenotypic traits were investigated and determined, and variation among and within populations were analyzed on nine natural groups in the distribution zone of I. polycarpa. The results indicated that there were significant differences in age, growth and yield traits of I. polycarpa from different population, especially in yield. There were rich variations in fifteen phenotypic traits including leaf length. These variation appeared among and within populations, and with similar contribution rate (31.99% and 30.82%). The variation of fruit traits were much more obvious in phenotypes, and the variation coefficients such as number of fruits per grain, which were closely related to yield, were up to 38%. The rich genetic variation of I. polycarpa provides many advantages for the development and utilization of germplasm resources, selecting and breeding.
URL [本文引用: 1]
以采自中国东南部亚热带7个种群的210棵单株的叶、果实和种子的12个形态指标作为研究对象,运用变异系数、巢式方差分析、相关分析揭示浙江柿()表型遗传变异规律和变异程度,并且运用聚类分析进行了居群分类。结果表明:浙江柿种内表型性状在群体间和群体内存在极其丰富的遗传差异,群体内的多样性程度高于群体间的多样性,而且种子性状在群体间较稳定。12个表型形状相关性不显著,浙江柿种居群表型变异在空间分布上主要呈现以纬度和海拔为主的单向变异模式,利用欧氏平均距离聚类分析,基本可将浙江柿7个群体可划分为5个类群。
URL [本文引用: 1]
以采自中国东南部亚热带7个种群的210棵单株的叶、果实和种子的12个形态指标作为研究对象,运用变异系数、巢式方差分析、相关分析揭示浙江柿()表型遗传变异规律和变异程度,并且运用聚类分析进行了居群分类。结果表明:浙江柿种内表型性状在群体间和群体内存在极其丰富的遗传差异,群体内的多样性程度高于群体间的多样性,而且种子性状在群体间较稳定。12个表型形状相关性不显著,浙江柿种居群表型变异在空间分布上主要呈现以纬度和海拔为主的单向变异模式,利用欧氏平均距离聚类分析,基本可将浙江柿7个群体可划分为5个类群。
.
DOI:10.3321/j.issn:1005-0094.2002.02.008URL [本文引用: 1]
Totally 13 natural populations of Pinus bungeana were collected. Five characteristics, including seed length (SL), seed width (SW), cone length (FL), cone width (FW) and weight (WT) of 1000 seeds, were measured. The variance analysis of the five characteristics showed that there were significant differences among populations with the value from 2.44 to 14.68, and there were also significant differences among families within a population with the value from 7.48 to 44.53. The variance factors analysis indicated that the variation within a population was the main part of the phenotypic variation in ST =22.8%), which meant that there was a strong adaptability to environment pressure in ) and relative extreme value (i) within each population show similar tendency. Based on Duncan's Multi comparison analysis, all the 13 populations were divided into four groups. The populations in groups A and B have higher phenotypic diversity while the populations in group C and group D have lower phenotypic diversity. Seed characteristics display a significant ecological gradient variation. There is close relationship between phenotypic diversity of seeds and economic characteristics of progeny.
DOI:10.3321/j.issn:1005-0094.2002.02.008URL [本文引用: 1]
Totally 13 natural populations of Pinus bungeana were collected. Five characteristics, including seed length (SL), seed width (SW), cone length (FL), cone width (FW) and weight (WT) of 1000 seeds, were measured. The variance analysis of the five characteristics showed that there were significant differences among populations with the value from 2.44 to 14.68, and there were also significant differences among families within a population with the value from 7.48 to 44.53. The variance factors analysis indicated that the variation within a population was the main part of the phenotypic variation in ST =22.8%), which meant that there was a strong adaptability to environment pressure in ) and relative extreme value (i) within each population show similar tendency. Based on Duncan's Multi comparison analysis, all the 13 populations were divided into four groups. The populations in groups A and B have higher phenotypic diversity while the populations in group C and group D have lower phenotypic diversity. Seed characteristics display a significant ecological gradient variation. There is close relationship between phenotypic diversity of seeds and economic characteristics of progeny.
DOI:10.3724/SP.J.1258.2013.00083URL [本文引用: 1]
植物叶片的形态性状能够有效地反映生存环境的变化,并且影响植物 的基本行为和功能.该研究通过获取植物标本提供的叶片形态信息,结合相关分析和标准化主轴分析,探讨了南北样带暖温带区栎属(Quercus)树种叶片形 态性状对气候条件的响应及适应策略.结果表明:在南北样带暖温带区,随着气候条件的变化,栎属树种的叶片形态性状发生显著的变化.随着年平均气温的降低和 年日照时数的增加,栎属树种叶面积增加,以利于吸收更多的光照辐射,并增加叶片的边界层阻力,减少叶片热量的散失;而叶片分裂程度的增加不仅可以降低热量 的散失,也可以提高叶片液流的波动以增强叶片的生理活动;叶脉密度随温度的升高、光照强度和降水量的增加而增加,以响应叶片蒸腾作用的增强,提高水分的运 输能力和叶片的支撑能力.此外,为适应南北样带暖温带区气候条件的变化,栎属树种的叶片形成了一系列的形态性状组合,随着叶面积的增加,叶柄长度和叶片分 裂程度逐渐增加,而叶脉密度降低;随着叶片倾向于向长条状发展,叶柄长度和叶脉密度也随之增加.
DOI:10.3724/SP.J.1258.2013.00083URL [本文引用: 1]
植物叶片的形态性状能够有效地反映生存环境的变化,并且影响植物 的基本行为和功能.该研究通过获取植物标本提供的叶片形态信息,结合相关分析和标准化主轴分析,探讨了南北样带暖温带区栎属(Quercus)树种叶片形 态性状对气候条件的响应及适应策略.结果表明:在南北样带暖温带区,随着气候条件的变化,栎属树种的叶片形态性状发生显著的变化.随着年平均气温的降低和 年日照时数的增加,栎属树种叶面积增加,以利于吸收更多的光照辐射,并增加叶片的边界层阻力,减少叶片热量的散失;而叶片分裂程度的增加不仅可以降低热量 的散失,也可以提高叶片液流的波动以增强叶片的生理活动;叶脉密度随温度的升高、光照强度和降水量的增加而增加,以响应叶片蒸腾作用的增强,提高水分的运 输能力和叶片的支撑能力.此外,为适应南北样带暖温带区气候条件的变化,栎属树种的叶片形成了一系列的形态性状组合,随着叶面积的增加,叶柄长度和叶片分 裂程度逐渐增加,而叶脉密度降低;随着叶片倾向于向长条状发展,叶柄长度和叶脉密度也随之增加.
DOI:10.3321/j.issn:1000-1522.2005.05.002URL [本文引用: 1]
为了研究辽东栎天然种群在山西灵空山自然保护区内的表型变异特征和影响因素,在该地区内共抽取了5个辽东栎天然种群,分别测了叶片及顶芽共24项形态特征.方差分析结果表明,各表型性状在种群内均达到极显著水平(α=0.01),种群间共有5个性状指标在α=0.05水平上差异显著,这些差异一方面来自于遗传因素,另一方面可能来自于环境因素.经研究得出,辽东栎表型性状受坡向、坡度、种群内树木平均胸径、种群密度等间接生态因子的影响.种群内变异是辽东栎表型变异的主要部分,表型分化系数(Vst)为13.31%,分化程度低,但部分性状也有了较大的分化,如最底部锯齿宽(25.99%)、中间顶芽宽(25.23%)等.在种群
DOI:10.3321/j.issn:1000-1522.2005.05.002URL [本文引用: 1]
为了研究辽东栎天然种群在山西灵空山自然保护区内的表型变异特征和影响因素,在该地区内共抽取了5个辽东栎天然种群,分别测了叶片及顶芽共24项形态特征.方差分析结果表明,各表型性状在种群内均达到极显著水平(α=0.01),种群间共有5个性状指标在α=0.05水平上差异显著,这些差异一方面来自于遗传因素,另一方面可能来自于环境因素.经研究得出,辽东栎表型性状受坡向、坡度、种群内树木平均胸径、种群密度等间接生态因子的影响.种群内变异是辽东栎表型变异的主要部分,表型分化系数(Vst)为13.31%,分化程度低,但部分性状也有了较大的分化,如最底部锯齿宽(25.99%)、中间顶芽宽(25.23%)等.在种群
URL [本文引用: 1]
[目的]通过对濒危植物水青树叶表型性状的测量分析,研究其表型变异程度和变异规律,并从遗传和环境适应角度探讨其致濒机制,为其天然种群的有效保护及管理提供科学依据。[方法]对来自14个水青树天然种群的90个个体的17个叶形态及其表皮微形态性状进行测量,利用巢式方差分析、多重比较、主成分分析、聚类分析、相关分析等多种分析方法,探讨种群间和种群内的叶表型变异及其与地理、环境因子的相关性。[结果](1)水青树17个叶表型性状在种群内和种群间均存在显著差异;(2)17个性状的平均表型分化系数为46.69%,种群内的变异(31.83%)大于种群间的变异(28.85%),种群内变异是水青树叶表型变异的主要来源;(3)各性状平均变异系数(CV)为12.56%,变异幅度为4.17%~26.25%;(4)水青树叶表型变异与年均日照时数、7月均温及年均降雨量等环境因子有关,变异呈现出随经度、纬度、海拔的梯度渐变规律;(5)利用主成分分析结果进一步聚类,可将14个水青树天然种群分为3大类。[结论]水青树天然种群间叶表型分化处于中等水平,种群内变异程度较低,其对环境的适应范围缩减,适应能力较差,这可能是导致水青树濒危的重要原因。据此,提出了相应的保护措施以便对其进行有效保护。
URL [本文引用: 1]
[目的]通过对濒危植物水青树叶表型性状的测量分析,研究其表型变异程度和变异规律,并从遗传和环境适应角度探讨其致濒机制,为其天然种群的有效保护及管理提供科学依据。[方法]对来自14个水青树天然种群的90个个体的17个叶形态及其表皮微形态性状进行测量,利用巢式方差分析、多重比较、主成分分析、聚类分析、相关分析等多种分析方法,探讨种群间和种群内的叶表型变异及其与地理、环境因子的相关性。[结果](1)水青树17个叶表型性状在种群内和种群间均存在显著差异;(2)17个性状的平均表型分化系数为46.69%,种群内的变异(31.83%)大于种群间的变异(28.85%),种群内变异是水青树叶表型变异的主要来源;(3)各性状平均变异系数(CV)为12.56%,变异幅度为4.17%~26.25%;(4)水青树叶表型变异与年均日照时数、7月均温及年均降雨量等环境因子有关,变异呈现出随经度、纬度、海拔的梯度渐变规律;(5)利用主成分分析结果进一步聚类,可将14个水青树天然种群分为3大类。[结论]水青树天然种群间叶表型分化处于中等水平,种群内变异程度较低,其对环境的适应范围缩减,适应能力较差,这可能是导致水青树濒危的重要原因。据此,提出了相应的保护措施以便对其进行有效保护。
DOI:10.1007/s11676-017-0409-4URL [本文引用: 1]
Phoebe bournei(Hemsl.) Yang is a rare and protected plant in China. This study was conducted to determine the phenotypic variation in this species and to document phenotypic variation within and among populations of P. bournei. Nested analysis of variance, coefficient of variation, multiple comparisons, principal component analysis(PCA) and correlation analysis were used to analyze ten phenotypic traits in ten natural populations of P.bournei from both the northeastern and the primary region of the range of this species. Significant differences among and within populations were observed in leaf and seed phenotypic traits. Variation among populations(34.92%)was greater than that within populations(26.19%). The mean phenotypic differentiation coefficient was 53.77%among populations, indicating that variation among populations comprised the majority of the phenotypic variation of P. bournei. The coefficient of variance(CV) of ten traits varied from 6.44 to 18.45%, with an average of 12.03%.The CV of leaf traits among populations(15.64%) was higher than that of seed traits(8.60%), indicating that seed traits were more stable. The results from CV and PCA indicated that leaf area, leaf length and thousand seed weight were the main factors accounting for the observed phenotypic variations. Significant or highly significant correlations were observed among most leaf and/or in seed phenotypic traits, whereas no significant correlations were observed between phenotypic traits and geographic factors.Based on cluster analysis, the ten populations can be divided into three clusters. These clusters were not a result of geographic distances.
DOI:10.3773/j.issn.1005-264x.2010.06.011URL [本文引用: 1]
干旱区植物叶片形态可塑性是植物适应高温干旱环境的重要生存策略,但目前仍缺乏直观的数据予以证明。该研究应用热成像技术和图像分析技术,同步测定真实叶片与模拟叶片的叶温、形态及风速、辐射和温度等环境参数。研究结果显示:在干旱、高温环境下,除了蒸腾,叶片形态变化也是调控叶温的重要因子。干旱区植物叶片变小,有利于加速叶片与环境的物质及热量交换,从而达到降低叶温的目的。样地数据显示,在高温、低风速环境下,叶片宽度每减少1 cm,叶片表面温度降低约2.1℃,而模拟叶片叶宽度每减少1 cm,叶片表面温度降低0.60–0.86℃。该研究对深入理解植物生存策略与环境适能力具有重要意义。
DOI:10.3773/j.issn.1005-264x.2010.06.011URL [本文引用: 1]
干旱区植物叶片形态可塑性是植物适应高温干旱环境的重要生存策略,但目前仍缺乏直观的数据予以证明。该研究应用热成像技术和图像分析技术,同步测定真实叶片与模拟叶片的叶温、形态及风速、辐射和温度等环境参数。研究结果显示:在干旱、高温环境下,除了蒸腾,叶片形态变化也是调控叶温的重要因子。干旱区植物叶片变小,有利于加速叶片与环境的物质及热量交换,从而达到降低叶温的目的。样地数据显示,在高温、低风速环境下,叶片宽度每减少1 cm,叶片表面温度降低约2.1℃,而模拟叶片叶宽度每减少1 cm,叶片表面温度降低0.60–0.86℃。该研究对深入理解植物生存策略与环境适能力具有重要意义。
URL [本文引用: 1]
为揭示长柄扁桃(Amygdalus pedunculata)天然种群的表型多样性特点,通过方差分析、相关分析、聚类分析和t检验等方法对7个天然种群14个表型性状种群间和种群内的表型变异特征和地理变异模式进行了探讨。结果表明:长柄扁桃种群内变异为40.91%,种群间变异为35.29%,种群内大于种群间,种群内变异是主要变异来源;其平均表型分化系数为45.90%,各表型性状平均变异系数为15.59%,変幅9.39%–31.98%,表型变异在种群内和种群间均非常丰富。年平均气温、纬度、无霜期、经度和海拔5个主要地理生态因子对长柄扁桃表型性状影响显著或极显著,平均气温和无霜期是不同立地长柄扁桃表型差异的主要影响因子。主成分分析和聚类分析将7个种群区划为两大类,其中山地的长柄扁桃叶多为近圆形至长圆形,果多为近球形,果柄较短,果肉较厚,核多为宽卵形至圆球形;沙地的长柄扁桃叶多为长椭圆形或卵状披针形,果为卵球形,果柄较长,果肉较薄,核近宽卵形。探讨长柄扁桃天然种群表型多样性,可为顺利开展野生资源收集、良种选育等工作提供依据。
URL [本文引用: 1]
为揭示长柄扁桃(Amygdalus pedunculata)天然种群的表型多样性特点,通过方差分析、相关分析、聚类分析和t检验等方法对7个天然种群14个表型性状种群间和种群内的表型变异特征和地理变异模式进行了探讨。结果表明:长柄扁桃种群内变异为40.91%,种群间变异为35.29%,种群内大于种群间,种群内变异是主要变异来源;其平均表型分化系数为45.90%,各表型性状平均变异系数为15.59%,変幅9.39%–31.98%,表型变异在种群内和种群间均非常丰富。年平均气温、纬度、无霜期、经度和海拔5个主要地理生态因子对长柄扁桃表型性状影响显著或极显著,平均气温和无霜期是不同立地长柄扁桃表型差异的主要影响因子。主成分分析和聚类分析将7个种群区划为两大类,其中山地的长柄扁桃叶多为近圆形至长圆形,果多为近球形,果柄较短,果肉较厚,核多为宽卵形至圆球形;沙地的长柄扁桃叶多为长椭圆形或卵状披针形,果为卵球形,果柄较长,果肉较薄,核近宽卵形。探讨长柄扁桃天然种群表型多样性,可为顺利开展野生资源收集、良种选育等工作提供依据。
URL [本文引用: 1]
麻栎(Quercus acutissima Carr.)是优良的能源和用材树种,在我国分布广泛并已有悠久栽培利用的历史,研究其种子和苗木性状地理变异规律对选择优良种源和适生区十分必要.研究收集了13个省(区)36个种源的麻栎种子,在测定麻栎种子性状的基础上,在安徽省滁州市红琊山林场进行了苗期试验和苗木性状测定,并采用趋势面分析方法对这些性状在经纬2维方向的地理变异模式进行了剖析.结果表明:1)麻栎种源间种子长度、宽度、百粒重和营养内含物存在极显著差异.种子百粒重、长度和宽度总体表现双向渐变趋势,随经度增高而增大,随纬度增高而减小,主要受到经度的控制;以西南到东北为中间地带,可溶性糖含量向东南表现先下降后上升的趋势,向西北则相反;淀粉含量从西北到东南呈逐渐减小的趋势.2)麻栎种源苗高、地径、生物量、热值和木材化学组分存在极显著差异.苗高、地径和生物量均呈双向渐变,经正向变异且变化幅度较大,纬负向变异且变化幅度较小,经度影响大于纬度;热值拟合回归方程不显著,方程无意义.木质素含量北部大于南部,但北部以西北部最高,南部以东南部最高.3)研究显示,麻栎种子和苗木性状多数存在显著的地理变异模式,这也是麻栎在长期进化过程中为适应复杂多变的环境而产生与之相适应的遗传变异结果.
URL [本文引用: 1]
麻栎(Quercus acutissima Carr.)是优良的能源和用材树种,在我国分布广泛并已有悠久栽培利用的历史,研究其种子和苗木性状地理变异规律对选择优良种源和适生区十分必要.研究收集了13个省(区)36个种源的麻栎种子,在测定麻栎种子性状的基础上,在安徽省滁州市红琊山林场进行了苗期试验和苗木性状测定,并采用趋势面分析方法对这些性状在经纬2维方向的地理变异模式进行了剖析.结果表明:1)麻栎种源间种子长度、宽度、百粒重和营养内含物存在极显著差异.种子百粒重、长度和宽度总体表现双向渐变趋势,随经度增高而增大,随纬度增高而减小,主要受到经度的控制;以西南到东北为中间地带,可溶性糖含量向东南表现先下降后上升的趋势,向西北则相反;淀粉含量从西北到东南呈逐渐减小的趋势.2)麻栎种源苗高、地径、生物量、热值和木材化学组分存在极显著差异.苗高、地径和生物量均呈双向渐变,经正向变异且变化幅度较大,纬负向变异且变化幅度较小,经度影响大于纬度;热值拟合回归方程不显著,方程无意义.木质素含量北部大于南部,但北部以西北部最高,南部以东南部最高.3)研究显示,麻栎种子和苗木性状多数存在显著的地理变异模式,这也是麻栎在长期进化过程中为适应复杂多变的环境而产生与之相适应的遗传变异结果.
DOI:10.13287/j.1001-9332.201807.013URL [本文引用: 1]
浙江楠为商品材"金丝楠木"的原植物之一,天然分布区窄,属国家二级保护渐危种,在全球气候变化背景下,研究其天然种群所在森林群落的结构特征及其多样性具有重要意义.本研究以浙江楠天然分布区内13个典型种群为对象,研究其所在群落的群落结构特征、物种多样性和叶片表型变异特征.结果表明:浙江楠天然种群所在群落的结构复杂,物种多样性较高,13个群落的16块样地内共有维管束植物87科162属235种,其中种子植物79科151属221种;重度干扰导致浙江开化、浙江临安等群落的乔木层物种多样性指数显著低于其他群落,而中度干扰提高了福建建宁群落灌木层物种多样性,轻度干扰则利于浙江楠种群自然更新.叶片表型作为楠木属植物重要的分类依据,叶片表型性状在种群间、种群内均存在丰富变异,平均变异系数为17.2%,变异幅度为10.4%~27.5%.种群间的变异(53.6%)大于种群内的变异(17.0%),平均表型分化系数为75.1%,种群间变异是浙江楠叶片表型变异的主要来源.基于欧式距离(10 cm)将13个天然种群划分为两大类群,但变异呈随机性.
DOI:10.13287/j.1001-9332.201807.013URL [本文引用: 1]
浙江楠为商品材"金丝楠木"的原植物之一,天然分布区窄,属国家二级保护渐危种,在全球气候变化背景下,研究其天然种群所在森林群落的结构特征及其多样性具有重要意义.本研究以浙江楠天然分布区内13个典型种群为对象,研究其所在群落的群落结构特征、物种多样性和叶片表型变异特征.结果表明:浙江楠天然种群所在群落的结构复杂,物种多样性较高,13个群落的16块样地内共有维管束植物87科162属235种,其中种子植物79科151属221种;重度干扰导致浙江开化、浙江临安等群落的乔木层物种多样性指数显著低于其他群落,而中度干扰提高了福建建宁群落灌木层物种多样性,轻度干扰则利于浙江楠种群自然更新.叶片表型作为楠木属植物重要的分类依据,叶片表型性状在种群间、种群内均存在丰富变异,平均变异系数为17.2%,变异幅度为10.4%~27.5%.种群间的变异(53.6%)大于种群内的变异(17.0%),平均表型分化系数为75.1%,种群间变异是浙江楠叶片表型变异的主要来源.基于欧式距离(10 cm)将13个天然种群划分为两大类群,但变异呈随机性.
DOI:10.2307/3599027URL [本文引用: 1]
http://www.jstor.org/stable/3599027
DOI:10.1111/plb.12564URL [本文引用: 2]
Abstract 1.Intraspecific trait variation and trait-climate relationships are crucial for understanding a species' response to climate change. However, these phenomena have been rarely studied for tree species. Euptelea pleiospermum (E. pleiospermum) is a relic tree species with a wide distribution in China that offers a novel opportunity to examine such relationships. 2.Here, we measured 13 leaf traits of E. pleiospermum in 20 sites across its natural distribution in China. We investigated the extent of trait variation at local and regional scales, and developed geographic and climatic models to explain trait variation at the regional scale. 3.We documented intraspecific trait variation among leaf traits of E. pleiospermum at local and regional scales. Five traits exhibited relatively high trait variation: leaf area, leaf density and three leaf economic traits (leaf dry matter content, specific leaf area and leaf phosphorus concentration). Significant trait-geography correlations were mediated by local climate. Most leaf trait variation could be explained (proportions from 24% to 64%) by geographic or climatic variables, except leaf width, leaf thickness, leaf dry matter content and leaf length-width ratio. Latitude and temperature were the strongest predictors of trait variation throughout the distribution of E. pleiospermum in China, and temperature explained more leaf trait variation than precipitation. In particular, we showed that leaves had longer petiole lengths, higher specific leaf areas (SLA) and lower densities in northern E. pleiospermum populations. We suggest that northern E. pleiospermum populations are adapting to higher latitudinal environments via high growth rate (higher SLAs) and low construction investment strategies (lower leaf densities), benefitting northern migration. 4.Overall, we demonstrate that intraspecific trait variation reflects E. pleiospermum response to local environment. We call for consideration of intraspecific trait variation to examine specific climate response questions. In addition, provenance experiments using widely-distributed species are needed to separate trait variation resulting from genetic differentiation and plastic responses to environmental change. This article is protected by copyright. All rights reserved.
DOI:10.1111/j.1469-8137.2006.01741.xURLPMID:16771985 [本文引用: 1]
090004 The implications of extensive variation in leaf size for biomass distribution between physiological and support tissues and for overall leaf physiological activity are poorly understood. Here, we tested the hypotheses that increases in leaf size result in enhanced whole-plant support investments, especially in compound-leaved species, and that accumulation of support tissues reduces average leaf nitrogen (N) content per unit dry mass ( N M ), a proxy for photosynthetic capacity. 090004 Leaf biomass partitioning among the lamina, mid-rib and petiole, and whole-plant investments in leaf support (within-leaf and stem) were studied in 33 simple-leaved and 11 compound-leaved species. 090004 Support investments in mid-ribs and petioles increased with leaf size similarly in simple leaves and leaflets of compound leaves, but the overall support mass fraction within leaves was larger in compound-leaved species as a result of prominent rachises. Within-leaf and within-plant support mass investments were negatively correlated. Therefore, the total plant support fraction was independent of leaf size and lamina dissection. Because of the lower N M of support biomass, the difference in N M between the entire leaf and the photosynthetic lamina increased with leaf size. 090004 We conclude that whole-plant support costs are weakly size-dependent, but accumulation of support structures within the leaf decreases whole-leaf average N M , potentially reducing the integrated photosynthetic activity of larger leaves.
URL [本文引用: 1]
随着分子生物学技术、电泳技术、同工酶染色技术和RFLP等技术的不断改造和发展,结合有关数据分析方法的建立和完善,群体遗传多样性、群体遗传结构和群体遗传分化已经成为群体遗传学的主体和物种多样性的重要内容,同时也受到育种学家的高度重视,成为目前最热门的研究领域之一。将对国内外有关的模型和方法进行了综合评述,并就遗传变异在群体内和群体间的分布问题提出自己的观点。
URL [本文引用: 1]
随着分子生物学技术、电泳技术、同工酶染色技术和RFLP等技术的不断改造和发展,结合有关数据分析方法的建立和完善,群体遗传多样性、群体遗传结构和群体遗传分化已经成为群体遗传学的主体和物种多样性的重要内容,同时也受到育种学家的高度重视,成为目前最热门的研究领域之一。将对国内外有关的模型和方法进行了综合评述,并就遗传变异在群体内和群体间的分布问题提出自己的观点。
DOI:10.3724/SP.J.1258.2012.00560URL [本文引用: 1]
为了揭示印楝(Azadirachta indica)实生种群表型变异程度和变异规律,以云南引种印楝人工林为研究对象,基于9个种群90个单株14个表型性状严格细致的测量,采用单因素方差分析、巢式方差分析、相关分析、协方差主成分分析(S法)和非加权配对算术平均法(UPGMA)聚类分析等数理方法,分析了种群的表型变异。结果表明:印楝种内表型性状在种群间和种群内均存在着较丰富的差异,种群内的变异大于种群间的变异,种群间的分化相对较小。对表型性状进行的变异系数多重比较和协方差主成分分析(S法)均显示,结实和种子化学成分相关性状的变异是造成印楝表型变异的主要来源。利用种群间欧氏距离进行的UPGMA聚类分析结果进一步表明,印楝9个种群可以分为4类,表型性状并没有严格依地理距离而聚类。研究结果为印楝的遗传改良工作奠定了基础,为制定育种策略和人工经营对策提供了科学依据。
DOI:10.3724/SP.J.1258.2012.00560URL [本文引用: 1]
为了揭示印楝(Azadirachta indica)实生种群表型变异程度和变异规律,以云南引种印楝人工林为研究对象,基于9个种群90个单株14个表型性状严格细致的测量,采用单因素方差分析、巢式方差分析、相关分析、协方差主成分分析(S法)和非加权配对算术平均法(UPGMA)聚类分析等数理方法,分析了种群的表型变异。结果表明:印楝种内表型性状在种群间和种群内均存在着较丰富的差异,种群内的变异大于种群间的变异,种群间的分化相对较小。对表型性状进行的变异系数多重比较和协方差主成分分析(S法)均显示,结实和种子化学成分相关性状的变异是造成印楝表型变异的主要来源。利用种群间欧氏距离进行的UPGMA聚类分析结果进一步表明,印楝9个种群可以分为4类,表型性状并没有严格依地理距离而聚类。研究结果为印楝的遗传改良工作奠定了基础,为制定育种策略和人工经营对策提供了科学依据。
DOI:10.1023/a:1009756318848URL [本文引用: 1]
Spatial patterns of plant diversity in the North American boreal forest were examined according to three plant life forms (woody plants, herbaceous plants, and bryophytes) and two taxonomic levels (species and genus), using sixty 9-ha plots sampled in white spruce (Picea glauca (Moench) Voss) and black spruce (Picea mariana (P. Mill.) B.S.P.) ecosystems along a transcontinental transect from the Pacific coast eastwards to the Atlantic coast. The patterns of inventory diversity (represented by alpha diversity), differentiation diversity (represented by the similarity index, habitat-heterogeneity index, similarity decay rate, and length of the first axis in detrended correspondence analysis), and pattern diversity (represented by the mosaic diversity index) were assessed along the transect in both ecosystem types. At the stand level, central North America had the highest alpha diversity in terms of the number of species or genera, and western North America had a higher alpha diversity than eastern North America. At the continental scale, herbaceous plants had the highest beta diversity in terms of floristic change from the eastern to western North America, bryophytes had the lowest beta diversity, and woody plants were in the middle, regardless of ecosystem type and taxonomic level. Central North America had the lowest mosaic diversity across the boreal transect of North America. The white spruce ecosystems had a higher alpha diversity than the black spruce ecosystems regardless of plant life form, taxonomic level and geographic location. The white spruce ecosystems tended to have more bryophytes, less woody plants, and higher species: genus ratio than the black spruce ecosystems. In general, the white spruce and black spruce ecosystems shared the same patterns in diversity changes at different spatial scales, plant life forms, and taxonomic levels across the transect studied. The existing patterns of plant diversity in the North American boreal forest area resulted from a combination of ecological processes and spatial configuration.
DOI:10.1016/j.aquabot.2018.05.004URL [本文引用: 1]
Avicennia marina is the most widely distributed of all mangrove species. This study examined phenotypic characteristics of seedlings raised from seven provenances of the species widely spread around the Australian coast from tropical to temperate regions. Seedlings were grown together for 18 months in a glasshouse at Lismore in sub-tropical eastern Australia. The provenances displayed differences in propagule and seedling size and seedling leaf characteristics that could be related to geographic occurrence but showed no distinct geographical cline. Consistent with other phenotypic and genetic studies of A. marina , phenotypic variation was mainly between provenances and between individuals across the entire population, rather than between individuals within any one provenance. The results suggested that provenance populations may have developed individually, separated by geographical barriers. If it can be assumed that characteristics observed in the present trial are likely to expressed by provenances when grown away from their natural environment, the present work has identified provenances that might be explored to find desirable individuals with phenotypes that might have value in mangrove planting or breeding programmes.
URL [本文引用: 1]
以青钱柳分布区内的9个天然群体种子为试材,对种子千粒重、体积、直径、厚度和径厚比5个表型性状进行了系统的比较分析,采用方差分析、相关分析及聚类分析等方法对群体间和群体内的表型多样性进行探讨.结果表明:青钱柳种子千粒重、体积、直径及厚度在群体间和群体内均达到显著或极显著差异,而种子径厚比仅在群体内存在极显著差异,说明青钱柳种子性状在2个层次上都具有丰富的多样性.各性状的平均表型分化系数(V)为20.54%,即群体间的变异远低于群体内的变异(79.4.6%).种子性状与地理因子相关程度存在差别,且受年均气温的影响最大.利用群体间欧氏距离进行系统聚类分析,青钱柳群体可以划分为三类.
URL [本文引用: 1]
以青钱柳分布区内的9个天然群体种子为试材,对种子千粒重、体积、直径、厚度和径厚比5个表型性状进行了系统的比较分析,采用方差分析、相关分析及聚类分析等方法对群体间和群体内的表型多样性进行探讨.结果表明:青钱柳种子千粒重、体积、直径及厚度在群体间和群体内均达到显著或极显著差异,而种子径厚比仅在群体内存在极显著差异,说明青钱柳种子性状在2个层次上都具有丰富的多样性.各性状的平均表型分化系数(V)为20.54%,即群体间的变异远低于群体内的变异(79.4.6%).种子性状与地理因子相关程度存在差别,且受年均气温的影响最大.利用群体间欧氏距离进行系统聚类分析,青钱柳群体可以划分为三类.
URL [本文引用: 1]
The Congo Basin has a large secondary forest area. Nevertheless, global plant trait databases lack substantial data from this biome and functional trait diversity is largely unknown. We analysed a unique leaf trait dataset (specific leaf area, nutrient and isotope concentrations) collected from 88 individual trees belonging to lOtlifferent species in tropical lowland forest in the Democratic Republic of the Congo. The trait data were found to be consistent with global datasets. &0167 N was the only trait significantly influenced by plot location. For all other leaf traits, shade tolerance was a significant factor. The species factor was significant within each shade tolerance class. This shows that shade tolerance is an important but not exclusive factor determining functional diversity. Tree height had significant influence on &0106C, specific leaf area and area-based nutrient concentrations. Higher individual trees had thicker sun-adapted leaves, regardless of the species. A principal component analysis (PCA) resulted in three significant ordination axes: leaf-thickness, N-content and P-content. By hierarchical clustering of the PCA scores, four functional groups were distinguished. This showed that species with diverse strategies coexisted in the ecosystem.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.1673-5102.2007.06.018URL [本文引用: 1]
The measurement of numerical taxonomy was applied to study the morphological variations of Cerasus subhirtella var. ascendens in different populations between Fujian Wuyi Mountain and Jiangsu Baohua Mountain. The 35 morphological indices analysis showed,the individuals in 2 populations were incorporated into 2 types according to the results of clustering analysis and principal component analysis(PCA),and the morphological variations are naturally showed in the results. The PCA results indicated that the characteristics of length,breadth,the angle of leaf basal in lamina and the characteristics of calyx-tube,corolla dimension and inflorescence pedicel in the flower were the main reasons leading to the morphological variations,and the F-test of single factor variance analysis verified the conclusion. Finally,the cherry in Fujian Wuyi Mountain should be treated as a new variation of Cerasus subhirtella in taxonomy.
DOI:10.3969/j.issn.1673-5102.2007.06.018URL [本文引用: 1]
The measurement of numerical taxonomy was applied to study the morphological variations of Cerasus subhirtella var. ascendens in different populations between Fujian Wuyi Mountain and Jiangsu Baohua Mountain. The 35 morphological indices analysis showed,the individuals in 2 populations were incorporated into 2 types according to the results of clustering analysis and principal component analysis(PCA),and the morphological variations are naturally showed in the results. The PCA results indicated that the characteristics of length,breadth,the angle of leaf basal in lamina and the characteristics of calyx-tube,corolla dimension and inflorescence pedicel in the flower were the main reasons leading to the morphological variations,and the F-test of single factor variance analysis verified the conclusion. Finally,the cherry in Fujian Wuyi Mountain should be treated as a new variation of Cerasus subhirtella in taxonomy.
DOI:10.13324/j.cnki.jfcf.2018.03.009URL [本文引用: 1]
为揭示湖北地区红椿天然居群叶与种实表型性状变异趋势与地理分布区的关系,采用均值与标准差、变异系数、变异幅度等指标对湖北红椿(Toona ciliata Roem.)天然居群叶片及种实表型地理变异进行研究。结果表明:湖北红椿10个叶片表型性状和12个种实表型性状均存在显著差异;叶尖角2次地理趋势面方程拟合度达88.13%;果纵径和种纵径4次趋势面拟合度分别达99.77%和99.52%,其他性状2次趋势面拟合度分别为:果大小指数93.40%、单果重95.34%、单种重95.00%,种大小指数为94.46%;F值检验表明趋势面拟合达显著水平;其它叶片、种实表型性状趋势面拟合方程不显著。红椿表型性状与环境因子相关分析表明:经度与大部分表型性状相关性显著;无霜期、年平均温度与红椿表型性状相关性较高;纬度、海拔与红椿表型相关性较低;叶尖角、果纵径、单果重、单种重是较为重要和关键的表型性状。湖北红椿叶片、种实表型变异表现为一定连续性,存在显著地理经度变异模式。
DOI:10.13324/j.cnki.jfcf.2018.03.009URL [本文引用: 1]
为揭示湖北地区红椿天然居群叶与种实表型性状变异趋势与地理分布区的关系,采用均值与标准差、变异系数、变异幅度等指标对湖北红椿(Toona ciliata Roem.)天然居群叶片及种实表型地理变异进行研究。结果表明:湖北红椿10个叶片表型性状和12个种实表型性状均存在显著差异;叶尖角2次地理趋势面方程拟合度达88.13%;果纵径和种纵径4次趋势面拟合度分别达99.77%和99.52%,其他性状2次趋势面拟合度分别为:果大小指数93.40%、单果重95.34%、单种重95.00%,种大小指数为94.46%;F值检验表明趋势面拟合达显著水平;其它叶片、种实表型性状趋势面拟合方程不显著。红椿表型性状与环境因子相关分析表明:经度与大部分表型性状相关性显著;无霜期、年平均温度与红椿表型性状相关性较高;纬度、海拔与红椿表型相关性较低;叶尖角、果纵径、单果重、单种重是较为重要和关键的表型性状。湖北红椿叶片、种实表型变异表现为一定连续性,存在显著地理经度变异模式。
DOI:10.1093/aob/mcl066URLPMID:28029050663 [本文引用: 2]
61 Background and Aims When ecologically important plant traits are correlated they may be said to constitute an ecological 'strategy' dimension. Through identifying these dimensions and understanding their inter-relationships we gain insight into why particular trait combinations are favoured over others and into the implications of trait differences among species. Here we investigated relationships among several traits, and thus the strategy dimensions they represented, across 2134 woody species from seven Neotropical forests. 61 Methods Six traits were studied: specific leaf area (SLA), the average size of leaves, seed and fruit, typical maximum plant height, and wood density (WD). Trait relationships were quantified across species at each individual forest as well as across the dataset as a whole. 'Phylogenetic' analyses were used to test for correlations among evolutionary trait-divergences and to ascertain whether interspecific relationships were biased by strong taxonomic patterning in the traits. 61 Key Results The interspecific and phylogenetic analyses yielded congruent results. Seed and fruit size were expected, and confirmed, to be tightly related. As expected, plant height was correlated with each of seed and fruit size, albeit weakly. Weak support was found for an expected positive relationship between leaf and fruit size. The prediction that SLA and WD would be negatively correlated was not supported. Otherwise the traits were predicted to be largely unrelated, being representatives of putatively independent strategy dimensions. This was indeed the case, although WD was consistently, negatively related to leaf size. 61 Conclusions The dimensions represented by SLA, seed/fruit size and leaf size were essentially independent and thus conveyed largely independent information about plant strategies. To a lesser extent the same was true for plant height and WD. Our tentative explanation for negative WD-leaf size relationships, now also known from other habitats, is that the traits are indirectly linked via plant hydraulics.
DOI:10.11983/CBB15024URL [本文引用: 1]
表型可塑性是指同一基因型的生物个体在不同环境条件下产生不同表型的特性,这一特性促使物种适应异质环境,具有更宽的生态幅。居群和物种水平广义的表型可塑性研究可揭示植物在自然环境中的生态意义并丰富对植物种群分布的了解。华中五味子(Schisandra sphenanthera)是一种资源严重减少的常用野生药材物种,对其进行研究和保护非常必要。叶是华中五味子主要的光合作用器官,随环境改变叶表型有很大变化,因此从居群水平上对华中五味子叶表型可塑性进行分析,揭示叶表型可塑性大小,探究环境因子对叶表型的影响,可为华中五味子的保护提供依据。该研究运用GIS技术从较大尺度(9省市26县27个居群)对华中五味子叶表型可塑性进行分析,并用表型可塑性指数和变异系数2种方法对叶表型可塑性进行评价,运用隶属函数和最大熵模型将3类(土壤、气候和地形)13个环境因子对叶表型的影响进行分析。结果表明,华中五味子7个叶表型(叶长、叶宽、叶柄长、叶面积、叶形指数、叶齿数和二级叶脉数)均存在可塑性,其中叶面积可塑性最大,叶形指数可塑性最小。3类环境因子中对叶表型影响最大的为土壤因子,气候因子次之,最小为地形因子。权重分析显示,土壤全钾含量对叶长和叶面积影响最大,有机碳含量、海拔、土壤全磷含量、坡度和年降水量分别对叶宽、叶柄长、叶形指数、叶齿数和二级叶脉数影响最大。研究结果表明,对多个叶表型有影响的5个共有重要因子为土壤全钾含量、全磷含量、有机碳含量、年降水量和坡度,它们与对应叶表型间的隶属函数多为高斯型,具有最优值,可通过人工控制达到最优值来促进其生长。该结果为华中五味子野生抚育和迁地种植提供理论参考。
DOI:10.11983/CBB15024URL [本文引用: 1]
表型可塑性是指同一基因型的生物个体在不同环境条件下产生不同表型的特性,这一特性促使物种适应异质环境,具有更宽的生态幅。居群和物种水平广义的表型可塑性研究可揭示植物在自然环境中的生态意义并丰富对植物种群分布的了解。华中五味子(Schisandra sphenanthera)是一种资源严重减少的常用野生药材物种,对其进行研究和保护非常必要。叶是华中五味子主要的光合作用器官,随环境改变叶表型有很大变化,因此从居群水平上对华中五味子叶表型可塑性进行分析,揭示叶表型可塑性大小,探究环境因子对叶表型的影响,可为华中五味子的保护提供依据。该研究运用GIS技术从较大尺度(9省市26县27个居群)对华中五味子叶表型可塑性进行分析,并用表型可塑性指数和变异系数2种方法对叶表型可塑性进行评价,运用隶属函数和最大熵模型将3类(土壤、气候和地形)13个环境因子对叶表型的影响进行分析。结果表明,华中五味子7个叶表型(叶长、叶宽、叶柄长、叶面积、叶形指数、叶齿数和二级叶脉数)均存在可塑性,其中叶面积可塑性最大,叶形指数可塑性最小。3类环境因子中对叶表型影响最大的为土壤因子,气候因子次之,最小为地形因子。权重分析显示,土壤全钾含量对叶长和叶面积影响最大,有机碳含量、海拔、土壤全磷含量、坡度和年降水量分别对叶宽、叶柄长、叶形指数、叶齿数和二级叶脉数影响最大。研究结果表明,对多个叶表型有影响的5个共有重要因子为土壤全钾含量、全磷含量、有机碳含量、年降水量和坡度,它们与对应叶表型间的隶属函数多为高斯型,具有最优值,可通过人工控制达到最优值来促进其生长。该结果为华中五味子野生抚育和迁地种植提供理论参考。
DOI:10.3969/j.issn.1674-7895.2016.03.10URL [本文引用: 1]
为探究不同产地暴马丁香〔Syringa reticulata subsp. amurensis (Rupr .) P. S. Green et M. C. Chang〕 种子和幼 苗的表型性状变异规律及其与地理- 气候因子的关系, 以来源于黑龙江、 河北、 北京、内蒙古、 青海和山西的1 0个种 源( 包括6 个野生种源和4 个栽培种源)的种子和幼苗为实验材料, 对种子长和宽、 种翅长和宽、 百粒质量、 发芽率、 叶长和宽、 叶长与叶宽的比值、 叶柄长、 株高和地径1 2个表型性状进行测量;在此基础上, 进行了方差分析、 变异系 数分析、 主成分分析、 聚类分析及相关性分析.结果显示:暴马丁香不同种源间种子和幼苗的表型性状均有极显著 差异(P = 0 . 01 ), 差异较大的表型性状为发芽率、 百粒质量、 种翅长和种翅宽.种源内和种源间各表型性状的变异 系数(CV)均差异较大, 种源内CV值为1 . 3 9 % -4 5 . 5 6 %, 种源间CV值为6 . 4 5 % -4 2 .7 6% ;其中, 种源内种翅长和 种翅宽的CV平均值较大, 发芽率和百粒质量的C V平均值均较小, 而种源间发芽率和百粒质量的CV值则较大.主成分分析结果表明:暴马丁香表型性状间的差异主要体现在种子长、 种子宽和叶长, 其次为株高、 发芽率、 地径、 叶宽和种翅宽.根据聚类分析结果, 供试的1 0个种源被划分为4 类, 其中, 来源于北京和内蒙古及3 个来源于河北的 种源聚为I 类, 来源于黑龙江哈尔滨的2 个种源聚为Ⅱ类, 来源于山西的2 个种源聚为Ⅲ类, 来源于青海的1 个种 源单独聚为Ⅳ类, 聚类结果与各种源的地理分布有关.相关性分析结果表明:各表型性状间及表型性状与地理- 气候因子间均存在不同的相关性, 其中, 种子发芽率与种子的其他5 个表型性状均无显著相关性;经度和纬度与各表型性状均无显著相关性, 而海拔、 年均温和无霜期则是影响表型性状变异的主要因子.综合分析17
DOI:10.3969/j.issn.1674-7895.2016.03.10URL [本文引用: 1]
为探究不同产地暴马丁香〔Syringa reticulata subsp. amurensis (Rupr .) P. S. Green et M. C. Chang〕 种子和幼 苗的表型性状变异规律及其与地理- 气候因子的关系, 以来源于黑龙江、 河北、 北京、内蒙古、 青海和山西的1 0个种 源( 包括6 个野生种源和4 个栽培种源)的种子和幼苗为实验材料, 对种子长和宽、 种翅长和宽、 百粒质量、 发芽率、 叶长和宽、 叶长与叶宽的比值、 叶柄长、 株高和地径1 2个表型性状进行测量;在此基础上, 进行了方差分析、 变异系 数分析、 主成分分析、 聚类分析及相关性分析.结果显示:暴马丁香不同种源间种子和幼苗的表型性状均有极显著 差异(P = 0 . 01 ), 差异较大的表型性状为发芽率、 百粒质量、 种翅长和种翅宽.种源内和种源间各表型性状的变异 系数(CV)均差异较大, 种源内CV值为1 . 3 9 % -4 5 . 5 6 %, 种源间CV值为6 . 4 5 % -4 2 .7 6% ;其中, 种源内种翅长和 种翅宽的CV平均值较大, 发芽率和百粒质量的C V平均值均较小, 而种源间发芽率和百粒质量的CV值则较大.主成分分析结果表明:暴马丁香表型性状间的差异主要体现在种子长、 种子宽和叶长, 其次为株高、 发芽率、 地径、 叶宽和种翅宽.根据聚类分析结果, 供试的1 0个种源被划分为4 类, 其中, 来源于北京和内蒙古及3 个来源于河北的 种源聚为I 类, 来源于黑龙江哈尔滨的2 个种源聚为Ⅱ类, 来源于山西的2 个种源聚为Ⅲ类, 来源于青海的1 个种 源单独聚为Ⅳ类, 聚类结果与各种源的地理分布有关.相关性分析结果表明:各表型性状间及表型性状与地理- 气候因子间均存在不同的相关性, 其中, 种子发芽率与种子的其他5 个表型性状均无显著相关性;经度和纬度与各表型性状均无显著相关性, 而海拔、 年均温和无霜期则是影响表型性状变异的主要因子.综合分析17
DOI:10.13430/j.cnki.jpgr.2015.01.012URL [本文引用: 1]
为了解白檀在湖南地区不同环境的适应性及遗传变化程度,对湖南地区6个白檀天然居群的189个个体的11个表型性状进行形态遗传多样性分析.结果表明,11个表型性状均具有丰富的遗传多样性(H'=1.389),居群间的平均方差分量为52.60%,居群内的平均方差分量为37.03%,说明在白檀形态性状多样性分布上是居群间多样性程度大于居群内,即居群间的形态变异是白檀形态变异的主要来源.聚类分析结果表明,6个居群的表型性状并没有严格依地理距离聚类,主成分与相关分析结果显示在11个表型中树型因子是主要形态变异因子,年降雨量与树型因子呈显著正相关,其他表型变异受遗传的影响可能大于受环境的影响.
DOI:10.13430/j.cnki.jpgr.2015.01.012URL [本文引用: 1]
为了解白檀在湖南地区不同环境的适应性及遗传变化程度,对湖南地区6个白檀天然居群的189个个体的11个表型性状进行形态遗传多样性分析.结果表明,11个表型性状均具有丰富的遗传多样性(H'=1.389),居群间的平均方差分量为52.60%,居群内的平均方差分量为37.03%,说明在白檀形态性状多样性分布上是居群间多样性程度大于居群内,即居群间的形态变异是白檀形态变异的主要来源.聚类分析结果表明,6个居群的表型性状并没有严格依地理距离聚类,主成分与相关分析结果显示在11个表型中树型因子是主要形态变异因子,年降雨量与树型因子呈显著正相关,其他表型变异受遗传的影响可能大于受环境的影响.
DOI:10.3969/j.issn.1000-2561.2013.10.035URL [本文引用: 1]
In order to reveal the phenotypic variation patterns of the seed and fruit traits of natural M. laosensis populations in different distribution areas,an investigation was conducted on 6 seed and fruit traits of 8 natural M. laosensis populations. We examined the morphological diversity among and within populations based on analysis of 6 phenotypic traits. Nested variance analysis,multi-comparison,correlation analysis and hierarchical cluster analysis were used to analyze the experimental results. There were significant differences in 6 seed and fruit traits among and within the populations. The mean phenotypic differentiation coefficient for the six traits was 33.73% which suggesting that the variance within the populations was the main part of the phenotypic variation.The average coefficient of variance(CV) among 6 traits was 13.90%,and the range of variation was 9.73% to18.39%. The size coefficient of seed was positively correlated with thousand seed weight. In general,the spatial variation of seed traits of natural populations was strongly related to longitude. There was significant relationship between geographic distances and mahalanobis distances calculated from all phenotypic traits(r=0.459 4, p=0.02). According to UPGMA cluster analysis by mahalanobis distances,the 8 populations of M. laosensis could be divided into 4 groups. These findings will provide scientific basis for collecting,evaluating and utilizing the germplasm resources of M. laosensis.
DOI:10.3969/j.issn.1000-2561.2013.10.035URL [本文引用: 1]
In order to reveal the phenotypic variation patterns of the seed and fruit traits of natural M. laosensis populations in different distribution areas,an investigation was conducted on 6 seed and fruit traits of 8 natural M. laosensis populations. We examined the morphological diversity among and within populations based on analysis of 6 phenotypic traits. Nested variance analysis,multi-comparison,correlation analysis and hierarchical cluster analysis were used to analyze the experimental results. There were significant differences in 6 seed and fruit traits among and within the populations. The mean phenotypic differentiation coefficient for the six traits was 33.73% which suggesting that the variance within the populations was the main part of the phenotypic variation.The average coefficient of variance(CV) among 6 traits was 13.90%,and the range of variation was 9.73% to18.39%. The size coefficient of seed was positively correlated with thousand seed weight. In general,the spatial variation of seed traits of natural populations was strongly related to longitude. There was significant relationship between geographic distances and mahalanobis distances calculated from all phenotypic traits(r=0.459 4, p=0.02). According to UPGMA cluster analysis by mahalanobis distances,the 8 populations of M. laosensis could be divided into 4 groups. These findings will provide scientific basis for collecting,evaluating and utilizing the germplasm resources of M. laosensis.
DOI:10.3969/j.issn.1000-2006.201708012URL [本文引用: 1]
【目的】基于海棠种质基因库优良品种资源丰富的优势,开展了以‘紫王子’海棠(Malus‘Purple Prince’)为母本的自由授粉子代试验研究,探索其半同胞家系表型特征及育种成效。【方法】运用Shannon-Wiener(H'R)及Pielou(ER)等指数进行14个性状的表型多样性比较,采用频率分布函数法进行亲子代间性状变异趋势及超亲性分析,通过层次分析法(AHP)进行表型性状综合评价。【结果】在表型多样性方面,‘紫王子’海棠半同胞家系14个性状的H'R平均值比疑似父本群体提高了13%,其中地径生长量、分枝数、完全功能叶颜色的H'R和ER超过疑似父本群体23.8%~53.2%。在遗传改良方面,与疑似父本群体相比,半同胞家系中综合性状得分高的优株比例下降了37.0%,紫红叶优株和速生优株的比例分别下降了14.9%和37.5%,但枫叶型、分枝多、通直度高、耐热性强的优株比例分别增加了12.6%、48.3%、28.7%和18.5%;与母本相比,7.8%的优株综合性状得分超过母本;此外,有些单株综合得分略低于母本,但部分单一性状表现比较突出,如F113和F60优株的新叶为鲜见的亮红色,F35和F108优株的叶片为枫叶型,观赏效果佳。【结论】尽管自由授粉降低了优株比例,但有效扩大了子代群体的遗传基础,对部分性状的遗传改良具有明显的效果。
DOI:10.3969/j.issn.1000-2006.201708012URL [本文引用: 1]
【目的】基于海棠种质基因库优良品种资源丰富的优势,开展了以‘紫王子’海棠(Malus‘Purple Prince’)为母本的自由授粉子代试验研究,探索其半同胞家系表型特征及育种成效。【方法】运用Shannon-Wiener(H'R)及Pielou(ER)等指数进行14个性状的表型多样性比较,采用频率分布函数法进行亲子代间性状变异趋势及超亲性分析,通过层次分析法(AHP)进行表型性状综合评价。【结果】在表型多样性方面,‘紫王子’海棠半同胞家系14个性状的H'R平均值比疑似父本群体提高了13%,其中地径生长量、分枝数、完全功能叶颜色的H'R和ER超过疑似父本群体23.8%~53.2%。在遗传改良方面,与疑似父本群体相比,半同胞家系中综合性状得分高的优株比例下降了37.0%,紫红叶优株和速生优株的比例分别下降了14.9%和37.5%,但枫叶型、分枝多、通直度高、耐热性强的优株比例分别增加了12.6%、48.3%、28.7%和18.5%;与母本相比,7.8%的优株综合性状得分超过母本;此外,有些单株综合得分略低于母本,但部分单一性状表现比较突出,如F113和F60优株的新叶为鲜见的亮红色,F35和F108优株的叶片为枫叶型,观赏效果佳。【结论】尽管自由授粉降低了优株比例,但有效扩大了子代群体的遗传基础,对部分性状的遗传改良具有明显的效果。
DOI:10.11926/jtsb.3702URL [本文引用: 1]
为了解浙闽樱桃(Cerasus schneideriana)地理分布特征及与气候限制因子之间的关系,基于DIVA-GIS平台获取实际地理分布点的气候资料,利用BIOCLIM模型预测当下适生区范围以及预测其未来潜在分布。结果表明,浙闽樱桃实际分布区覆盖浙江、福建、安徽、江西、广西及湖南6省,浙闽交界的山区是自然分布的核心区,浙皖交界是其分布的北界。未来气候变化情境(CCM3)下,浙闽樱桃的潜在分布区概率将增大,且有北扩的趋势。主成分分析(PCA)表明,年降水量(bio12)、最湿季降雨量(bio16)、最暖季降雨量(bio18)、温度季节变化方差(bio4)是影响浙闽樱桃当下适生区的气候限制因子,频率直方图进一步确定他们的适宜范围分别为1503~2003mm、604~951mm、528~791mm和601~872(标准差*100)。ROC曲线检验表明BIOCLIM对浙闽樱桃分布预测的精度很高(AUC=0.998),结果是可信的。因此,浙闽樱桃适合生长在我国北亚热带温暖湿润区,水热因子是造成其分布格局的主导气候限制因子。
DOI:10.11926/jtsb.3702URL [本文引用: 1]
为了解浙闽樱桃(Cerasus schneideriana)地理分布特征及与气候限制因子之间的关系,基于DIVA-GIS平台获取实际地理分布点的气候资料,利用BIOCLIM模型预测当下适生区范围以及预测其未来潜在分布。结果表明,浙闽樱桃实际分布区覆盖浙江、福建、安徽、江西、广西及湖南6省,浙闽交界的山区是自然分布的核心区,浙皖交界是其分布的北界。未来气候变化情境(CCM3)下,浙闽樱桃的潜在分布区概率将增大,且有北扩的趋势。主成分分析(PCA)表明,年降水量(bio12)、最湿季降雨量(bio16)、最暖季降雨量(bio18)、温度季节变化方差(bio4)是影响浙闽樱桃当下适生区的气候限制因子,频率直方图进一步确定他们的适宜范围分别为1503~2003mm、604~951mm、528~791mm和601~872(标准差*100)。ROC曲线检验表明BIOCLIM对浙闽樱桃分布预测的精度很高(AUC=0.998),结果是可信的。因此,浙闽樱桃适合生长在我国北亚热带温暖湿润区,水热因子是造成其分布格局的主导气候限制因子。
Phenotypic characterization of Elaeagnus angustifolia using multivariate analysis
1
2018
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
基于SSR标记的四川野生中国樱桃遗传多样性和居群遗传结构分析
1
2013
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
基于SSR标记的四川野生中国樱桃遗传多样性和居群遗传结构分析
1
2013
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
用扫描仪及Image J软件精确测量叶片形态数量特征的方法
1
2009
... 将尾叶樱桃8个种群的200枚叶片利用佳能Lide 120扫描仪(Canon, B?c Ninh, Vietnam)逐一进行扫描, 参考
用扫描仪及Image J软件精确测量叶片形态数量特征的方法
1
2009
... 将尾叶樱桃8个种群的200枚叶片利用佳能Lide 120扫描仪(Canon, B?c Ninh, Vietnam)逐一进行扫描, 参考
基于种实性状的无患子天然群体表型多样性研究
1
2014
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
基于种实性状的无患子天然群体表型多样性研究
1
2014
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
浙江省蜡梅野生种群种实表型性状变异及其与土壤相关因子的关系
1
2018
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
浙江省蜡梅野生种群种实表型性状变异及其与土壤相关因子的关系
1
2018
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
1
2016
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
1
2016
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
用同工酶研究马尾松群体的遗传结构
2
1988
... 观测记载的数据在Excel 2007软件中进行整理汇总, 根据各性状25个样品的平均值($\bar{X}$)和标准偏差(SD)计算变异系数(CV =标准偏差/平均值); 采用SPSS 16.0软件中的单因素方差分析(显著性检验采用多重比较Duncan法), 研究叶表型性状在不同种群间的变异水平; 采用巢式方差设计(
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
用同工酶研究马尾松群体的遗传结构
2
1988
... 观测记载的数据在Excel 2007软件中进行整理汇总, 根据各性状25个样品的平均值($\bar{X}$)和标准偏差(SD)计算变异系数(CV =标准偏差/平均值); 采用SPSS 16.0软件中的单因素方差分析(显著性检验采用多重比较Duncan法), 研究叶表型性状在不同种群间的变异水平; 采用巢式方差设计(
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS
1
2001
... 本研究的气候数据来源于世界气候数据库WorldClim (http://www.worldclim.org/), 主要是根据1950-2000年世界各地气象站的气候观测记录, 采用插值法生成的空间分辨率为2.5° (约5 km2)的全球气候数据包; 各种群的经度、纬度、海拔等地理信息由手持GPS仪(UNI-T UT379B, 优利德, 广东东莞)野外测定.利用DIVA-GIS软件(
Heritable plant phenotypes track light and herbivory levels at fine spatial scales
1
2018
... 尾叶樱桃作为优良的国产樱属代表性野生植物, 颇具观赏价值, 其市场潜在价值近年来已经逐渐被人们所认识.然而当前尾叶樱桃天然种群正不断受到人类经济活动的影响(如城市化、旅游开发), 面临生境遭到破坏、资源减少的威胁; 就本研究而言, 8个尾叶樱桃代表种群均位于人为干扰较少的自然保护区或保护较好的景区, 其他地区发现天然种群的概率越来越低.由于种群内变异是尾叶樱桃种群变异的主要来源, 在开展种质资源收集、迁地或就地保存与遗传改良时, 可适当减少抽样种群个体数, 增加核心种群内的取样密度, 尤其是湖北、湖南、江西三省的种群, 以增加遗传变异的丰富性; 同时, 种群间又存在中等程度的叶表型分化, 不同层次的变异为优质种质资源和生物多样性保护提供了物质基础(
季风对亚洲热带植物分布格局的影响
1
2017
... 植物形态特征变异往往具有适应性意义, 广阔分布区、多样的生境与气候异质性, 为研究尾叶樱桃叶表型地理变异模式提供了可能.从种群生态学的角度看, 本研究选取的8个代表种群分布于我国南方长江流域, 跨度范围为23.50°-31.45° N, 103.37°-120.80° E, 平均海拔范围839-1 365 m, 其中最西缘峨眉山种群属于横断山脉东部边缘的邛崃山系, 平均海拔1 365 m, 在各种群中海拔分布最高; 而最东南缘的台湾种群, 与祖国大陆隔海相望, 所在生境受亚热带季风气候影响(
季风对亚洲热带植物分布格局的影响
1
2017
... 植物形态特征变异往往具有适应性意义, 广阔分布区、多样的生境与气候异质性, 为研究尾叶樱桃叶表型地理变异模式提供了可能.从种群生态学的角度看, 本研究选取的8个代表种群分布于我国南方长江流域, 跨度范围为23.50°-31.45° N, 103.37°-120.80° E, 平均海拔范围839-1 365 m, 其中最西缘峨眉山种群属于横断山脉东部边缘的邛崃山系, 平均海拔1 365 m, 在各种群中海拔分布最高; 而最东南缘的台湾种群, 与祖国大陆隔海相望, 所在生境受亚热带季风气候影响(
山桐子自然群体表型性状变异分析
1
2013
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
山桐子自然群体表型性状变异分析
1
2013
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
浙江柿天然群体表型多样性研究
1
2010
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
浙江柿天然群体表型多样性研究
1
2010
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
白皮松天然群体种实性状表型多样性研究
1
2002
... 观测记载的数据在Excel 2007软件中进行整理汇总, 根据各性状25个样品的平均值($\bar{X}$)和标准偏差(SD)计算变异系数(CV =标准偏差/平均值); 采用SPSS 16.0软件中的单因素方差分析(显著性检验采用多重比较Duncan法), 研究叶表型性状在不同种群间的变异水平; 采用巢式方差设计(
白皮松天然群体种实性状表型多样性研究
1
2002
... 观测记载的数据在Excel 2007软件中进行整理汇总, 根据各性状25个样品的平均值($\bar{X}$)和标准偏差(SD)计算变异系数(CV =标准偏差/平均值); 采用SPSS 16.0软件中的单因素方差分析(显著性检验采用多重比较Duncan法), 研究叶表型性状在不同种群间的变异水平; 采用巢式方差设计(
中国东部南北样带暖温带区栎属树种叶片形态性状对气候条件的响应
1
2013
... 片蒸腾作用的增强, 随降水的增加而增加, 以提高叶片的抗打击能力(
中国东部南北样带暖温带区栎属树种叶片形态性状对气候条件的响应
1
2013
... 片蒸腾作用的增强, 随降水的增加而增加, 以提高叶片的抗打击能力(
山西灵空山辽东栎种群叶性状表型变异研究
1
2005
... 将尾叶樱桃8个种群的200枚叶片利用佳能Lide 120扫描仪(Canon, B?c Ninh, Vietnam)逐一进行扫描, 参考
山西灵空山辽东栎种群叶性状表型变异研究
1
2005
... 将尾叶樱桃8个种群的200枚叶片利用佳能Lide 120扫描仪(Canon, B?c Ninh, Vietnam)逐一进行扫描, 参考
濒危植物水青树叶的表型性状变异
1
2016
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
濒危植物水青树叶的表型性状变异
1
2016
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
Phenotypic variation in Phoebe bournei populations preserved in the primary distribution area
1
2018
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
形态变化对叶片表面温度的影响
1
2018
... 植物形态特征变异往往具有适应性意义, 广阔分布区、多样的生境与气候异质性, 为研究尾叶樱桃叶表型地理变异模式提供了可能.从种群生态学的角度看, 本研究选取的8个代表种群分布于我国南方长江流域, 跨度范围为23.50°-31.45° N, 103.37°-120.80° E, 平均海拔范围839-1 365 m, 其中最西缘峨眉山种群属于横断山脉东部边缘的邛崃山系, 平均海拔1 365 m, 在各种群中海拔分布最高; 而最东南缘的台湾种群, 与祖国大陆隔海相望, 所在生境受亚热带季风气候影响(
形态变化对叶片表面温度的影响
1
2018
... 植物形态特征变异往往具有适应性意义, 广阔分布区、多样的生境与气候异质性, 为研究尾叶樱桃叶表型地理变异模式提供了可能.从种群生态学的角度看, 本研究选取的8个代表种群分布于我国南方长江流域, 跨度范围为23.50°-31.45° N, 103.37°-120.80° E, 平均海拔范围839-1 365 m, 其中最西缘峨眉山种群属于横断山脉东部边缘的邛崃山系, 平均海拔1 365 m, 在各种群中海拔分布最高; 而最东南缘的台湾种群, 与祖国大陆隔海相望, 所在生境受亚热带季风气候影响(
长柄扁桃天然种群表型变异
1
2017
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
长柄扁桃天然种群表型变异
1
2017
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
不同种源麻栎种子和苗木性状地理变异趋势面分析
1
2011
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
不同种源麻栎种子和苗木性状地理变异趋势面分析
1
2011
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
渐危植物浙江楠群落结构及叶片性状多样性
1
2018
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
渐危植物浙江楠群落结构及叶片性状多样性
1
2018
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
Leaf-size divergence along rainfall and soil-nutrient gradients: Is the method of size reduction common among clades?
1
2003
... 植物形态特征变异往往具有适应性意义, 广阔分布区、多样的生境与气候异质性, 为研究尾叶樱桃叶表型地理变异模式提供了可能.从种群生态学的角度看, 本研究选取的8个代表种群分布于我国南方长江流域, 跨度范围为23.50°-31.45° N, 103.37°-120.80° E, 平均海拔范围839-1 365 m, 其中最西缘峨眉山种群属于横断山脉东部边缘的邛崃山系, 平均海拔1 365 m, 在各种群中海拔分布最高; 而最东南缘的台湾种群, 与祖国大陆隔海相望, 所在生境受亚热带季风气候影响(
Geographical variation and the role of climate in leaf traits of a relict tree species across its distribution in China
2
2017
... 将尾叶樱桃8个种群的200枚叶片利用佳能Lide 120扫描仪(Canon, B?c Ninh, Vietnam)逐一进行扫描, 参考
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
Leaf size modifies support biomass distribution among stems, petioles and mid-ribs in temperate plants
1
2010
... 植物形态特征变异往往具有适应性意义, 广阔分布区、多样的生境与气候异质性, 为研究尾叶樱桃叶表型地理变异模式提供了可能.从种群生态学的角度看, 本研究选取的8个代表种群分布于我国南方长江流域, 跨度范围为23.50°-31.45° N, 103.37°-120.80° E, 平均海拔范围839-1 365 m, 其中最西缘峨眉山种群属于横断山脉东部边缘的邛崃山系, 平均海拔1 365 m, 在各种群中海拔分布最高; 而最东南缘的台湾种群, 与祖国大陆隔海相望, 所在生境受亚热带季风气候影响(
群体遗传多样性和数据分析
1
1995
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
群体遗传多样性和数据分析
1
1995
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
云南引种印楝实生种群的表型变异
1
2012
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
云南引种印楝实生种群的表型变异
1
2012
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
Longitudinal patterns of plant diversity in the North American boreal forest
1
1998
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
Phenotypic variation of the mangrove species Avicennia marina( Forssk.) Vierh. from seven provenances around Australia.
1
2018
... 植物表型多样性是遗传适应性和环境异质性长期共同作用的结果, 植物表型变异程度越丰富, 越能提高在不同环境下的适应程度(
青钱柳天然群体种子性状表型多样性
1
2009
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
青钱柳天然群体种子性状表型多样性
1
2009
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
Functional leaf trait diversity of 10 tree species in Congolese secondary tropical forest
1
2014
... 植物表型多样性是遗传适应性和环境异质性长期共同作用的结果, 植物表型变异程度越丰富, 越能提高在不同环境下的适应程度(
Constraints on the evolution of phenotypic plasticity in the clonal plant Hydrocotyle vulgaris
1
2018
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
1
2014
... 尾叶樱桃(Cerasus dielsiana)隶属蔷薇科樱属, 为我国特有落叶小乔木, 主要分布在我国亚热带季风区的各省山区, 分布海拔500-1 400 m; 因其树形优美、早春先花后叶、花色多样, 具有极高的观赏价值, 加上生态适应性广、抗逆性强, 市场开发前景十分广阔(
1
2014
... 尾叶樱桃(Cerasus dielsiana)隶属蔷薇科樱属, 为我国特有落叶小乔木, 主要分布在我国亚热带季风区的各省山区, 分布海拔500-1 400 m; 因其树形优美、早春先花后叶、花色多样, 具有极高的观赏价值, 加上生态适应性广、抗逆性强, 市场开发前景十分广阔(
不同居群野生早樱形态变异研究
1
2007
... 将尾叶樱桃8个种群的200枚叶片利用佳能Lide 120扫描仪(Canon, B?c Ninh, Vietnam)逐一进行扫描, 参考
不同居群野生早樱形态变异研究
1
2007
... 将尾叶樱桃8个种群的200枚叶片利用佳能Lide 120扫描仪(Canon, B?c Ninh, Vietnam)逐一进行扫描, 参考
湖北红椿天然居群表型变异地理趋势面研究
1
2018
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
湖北红椿天然居群表型变异地理趋势面研究
1
2018
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests
2
2007
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
华中五味子叶表型可塑性及环境因子对叶表型的影响
1
2016
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
华中五味子叶表型可塑性及环境因子对叶表型的影响
1
2016
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
暴马丁香不同种源种子和幼苗的表型性状变异及其与地理-气候因子的相关性
1
2016
... 尾叶樱桃作为优良的国产樱属代表性野生植物, 颇具观赏价值, 其市场潜在价值近年来已经逐渐被人们所认识.然而当前尾叶樱桃天然种群正不断受到人类经济活动的影响(如城市化、旅游开发), 面临生境遭到破坏、资源减少的威胁; 就本研究而言, 8个尾叶樱桃代表种群均位于人为干扰较少的自然保护区或保护较好的景区, 其他地区发现天然种群的概率越来越低.由于种群内变异是尾叶樱桃种群变异的主要来源, 在开展种质资源收集、迁地或就地保存与遗传改良时, 可适当减少抽样种群个体数, 增加核心种群内的取样密度, 尤其是湖北、湖南、江西三省的种群, 以增加遗传变异的丰富性; 同时, 种群间又存在中等程度的叶表型分化, 不同层次的变异为优质种质资源和生物多样性保护提供了物质基础(
暴马丁香不同种源种子和幼苗的表型性状变异及其与地理-气候因子的相关性
1
2016
... 尾叶樱桃作为优良的国产樱属代表性野生植物, 颇具观赏价值, 其市场潜在价值近年来已经逐渐被人们所认识.然而当前尾叶樱桃天然种群正不断受到人类经济活动的影响(如城市化、旅游开发), 面临生境遭到破坏、资源减少的威胁; 就本研究而言, 8个尾叶樱桃代表种群均位于人为干扰较少的自然保护区或保护较好的景区, 其他地区发现天然种群的概率越来越低.由于种群内变异是尾叶樱桃种群变异的主要来源, 在开展种质资源收集、迁地或就地保存与遗传改良时, 可适当减少抽样种群个体数, 增加核心种群内的取样密度, 尤其是湖北、湖南、江西三省的种群, 以增加遗传变异的丰富性; 同时, 种群间又存在中等程度的叶表型分化, 不同层次的变异为优质种质资源和生物多样性保护提供了物质基础(
湖南白檀居群形态多样性及与环境的相关性
1
2015
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
湖南白檀居群形态多样性及与环境的相关性
1
2015
... 在巢氏方差中各层次上的方差分量百分比是说明变异来源的一个重要指标, 也是计算表型分化系数的主要依据(
米老排天然群体的种实表型变异研究初报
1
2013
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
米老排天然群体的种实表型变异研究初报
1
2013
... 植物生态学研究的一个重要内容就是确定和量化各物种生态变量的主导维度, 并尝试阐释其机理(
“紫王子”海棠半同胞家系苗期表型特征分析
1
2017
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
“紫王子”海棠半同胞家系苗期表型特征分析
1
2017
... 植物物种多样性研究有多种标记方式, 形态学标记(表型)是其中的重要组成部分(
浙闽樱桃地理分布模拟及气候限制因子分析
1
2017
... 本研究的气候数据来源于世界气候数据库WorldClim (http://www.worldclim.org/), 主要是根据1950-2000年世界各地气象站的气候观测记录, 采用插值法生成的空间分辨率为2.5° (约5 km2)的全球气候数据包; 各种群的经度、纬度、海拔等地理信息由手持GPS仪(UNI-T UT379B, 优利德, 广东东莞)野外测定.利用DIVA-GIS软件(
浙闽樱桃地理分布模拟及气候限制因子分析
1
2017
... 本研究的气候数据来源于世界气候数据库WorldClim (http://www.worldclim.org/), 主要是根据1950-2000年世界各地气象站的气候观测记录, 采用插值法生成的空间分辨率为2.5° (约5 km2)的全球气候数据包; 各种群的经度、纬度、海拔等地理信息由手持GPS仪(UNI-T UT379B, 优利德, 广东东莞)野外测定.利用DIVA-GIS软件(

{kind=link}
{kind=link}