 ,1,*
,1,*Comparison of methods for detecting vulnerability of xylem embolism in Robinia pseudoacacia
AN Rui1, MENG Feng1, YIN Peng-Xian2, DU Guang-Yuan,1,*通讯作者:
收稿日期:2018-06-19接受日期:2018-10-19网络出版日期:2018-11-20
| 基金资助: |
Received:2018-06-19Accepted:2018-10-19Online:2018-11-20
| Fund supported: |

摘要
关键词:
Abstract
Methods Robinia pseudoacacia branches, with long xylem vessels, were used as plant materials for comparison of three different methods in establishing vulnerability curves, including bench top dehydration, Cochard Cavitron centrifugation and Sperry centrifugation. In the Sperry centrifugation method, rotors of two different sizes were used to test the ‘open vessel artifact’ hypothesis.
Important findings The vulnerability curve established by the bench top dehydration method displayed an “s” shape, while both the Cochard Cavitron centrifugation and Sperry centrifugation methods produced “r” shape curves. Vulnerability curves derived from the bench top dehydration method and the centrifugation methods were significantly different. Using the Sperry centrifugation method, the R. pseudoacacia branch samples in the 14.4 cm rotor had a higher proportion of open vessels, while the embolic vulnerability curves established on the 27.4 cm and 14.4 cm long stem segments were similar, indicating that the Sperry centrifugation method does not produce “open vessel artifact”.
Keywords:
PDF (1502KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
安瑞, 孟凤, 尹鹏先, 杜光源. 刺槐木质部栓塞脆弱性检测的方法比较. 植物生态学报[J], 2018, 42(11): 1113-1119 doi:10.17521/cjpe.2018.0145
AN Rui, MENG Feng, YIN Peng-Xian, DU Guang-Yuan.
近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(Allen et al., 2010)。在众多干旱相关的植物生理学因素中, 水力学失衡被认为是林木死亡的一个关键因素(董雷和李吉跃, 2013; 代永欣等, 2015; Adams et al., 2017)。根据内聚力-张力学说, 植物木质部水分因叶片蒸腾作用而处于“负压”状态, 即水力学压强低于大气压(Dixon & Joly, 1895; van den Honert, 1948)。因此, 植物水力学系统比较脆弱, 一旦发生干旱胁迫, 木质部的水力学压强将随之降低;当木质部压强降低至木质部结构能够承受的临界值以下, 则木质部水流将被“拉断”, 形成“空化”, 继而空气迅速进入该区域产生“栓塞” (Venturas et al., 2017)。由于栓塞化的木质部导管的压强与大气压相等, 且远远大于水力学功能正常的木质部导管中的“负压”, 栓塞化木质部的水力学功能的恢复是一个漫长而困难的过程(Sperry & Tyree, 1988)。较大程度的栓塞对于林木而言往往是灾难性的, 鉴于此, 客观描述植物木质部栓塞的脆弱性、准确测定植物木质部可承受的“负压”临界值, 对于深入理解植物抗旱生理机制以及准确判断森林生态受未来气候变化的影响, 具有重要意义。
木质部栓塞脆弱性可以用脆弱性曲线(VC)来描述, VC是木质部导水率损失的百分数与相对应水势的关系曲线(Sperry & Tyree, 1988; Maherali et al., 2004), 目前报道的VC主要有两种形态, 即“r”形和“s”形。VC呈“r”形的枝条在水势稍有降低时就会发生栓塞, 而VC呈“s”形的枝条则是在水势降低到一定程度后才会发生栓塞。根据Domec理论, 导水率损失50%时对应的枝条水势值(P50)可以很好地描述木质部的“负压”承受能力, 通常VC呈“r”形的枝条的P50远小于VC呈“s”形的枝条的P50 (Domec & Gartner, 2001)。
目前, 建立VC的方法有很多, 常用的方法有自然干燥法(Tyree et al., 1992)、压力套法(Melcher et al., 2003; Choat et al., 2005)、Cochard Cavitron离心机法(Cochard, 2002; Cochard et al., 2005)和Sperry离心机法(Sperry et al., 1988)。这些方法采用了不同原理来诱导木质部栓塞, 自然干燥法是直接将植物放在空气中进行脱水诱导栓塞; 压力套法则是通过压力套将压缩的空气注入导管中, 使其发生栓塞; 而Cochard Cavitron离心机法和Sperry离心机法则是利用不断增加的离心力诱导木质部栓塞。然而越来越多的研究发现, 在很多具有长导管的植物中, 用不同的方法建立的VC存在差异。一般自然干燥法建立的长导管植物的VC为“s”形, 而另外3种方法得到的VC则大多为“r”形(Cochard et al., 2013)。哪些方法建立的VC能够正确代表其物种的栓塞脆弱性?不同的****有不同的观点。Torres-Ruiz等(2015)以具有长导管的木犀榄(Olea europaea)为试验材料, 利用离心机法和自然干燥法进行比较, 发现当枝条样品的长度较短时, 离心机法建立的“r”形VC是错误的。Rockwell等(2014)在被子植物的研究中, 也得到相同的结论。但是, Hacke等(2015)对长导管橄榄(Canarium album)的研究中, 发现离心机法建立的曲线和自然干燥法建立的曲线都是“r”形。有****认为植物导管单元之间的纹孔膜和多孔板对植物茎流有重要的过滤作用, 在采用离心机方法建立VC时, 如果枝条导管长度大于转子半径, 则从枝条末端至转轴存在连通的单个导管单元, 水中的气泡或杂质将在离心作用下迅速聚集于枝条中段而产生“人为”栓塞, 即出现所谓的“开口导管假象” (Choat et al., 2010; Cochard et al., 2010)。Wang等(2014)对长导管树种刺槐进行了栓塞脆弱性分析, 发现自然干燥法测得VC的P50为3.57 MPa, 而Cochard Cavitron离心机法测得的P50为0.30 MPa。前者似乎更为合理, 因为在水分状况良好的情况下, 压力室法测得刺槐(Robinia pseudoacacia)的正午木质部水势约为-1.6 MPa。
压力套法使植物木质部处于“正压”状态, 与自然环境下的木质部导管的“负压”有显著差异; 而自然干燥法以及离心机法则更接近植物的自然状态。本研究选取一年生的长导管刺槐为对象, 用自然干燥法、Cochard Cavitron离心机法以及Sperry离心机法建立其栓塞脆弱性曲线, 并进行比较, 以探究这3种方法建立VC的有效性。此外, 本研究在使用Sperry离心机时根据刺槐的导管分布, 采用了不同内径的定制转子, 以检验是否存在“开口导管假象”, 以期为长导管植物栓塞脆弱性研究提供依据。
1 材料和方法
1.1 试验区的自然状况
试验在陕西杨陵区(34.27° N, 108.07° E, 海拔457 m)进行, 该地位于关中平原中部, 属于半湿润地区, 年平均气温12.9 ℃。一年中最热的月份(8月)平均气温为25.8 ℃, 而最冷的月份(2月)平均气温为-1.1 ℃, 最高气温能达到42 ℃, 最低气温能达到-12.9 ℃, 年降水量651 mm, 4-9月份为试验材料的生长季, 降水量占全年的79%。1.2 试验材料
试验材料选择陕西杨陵区渭河河堤正常生长的长导管树种刺槐。于2017年8-10月6:00-7:00在树冠中部剪取一年生枝条。所剪取的枝条的基部直径为7 mm左右, 长为1.4-2.0 m, 剪取后, 用水喷晒枝条叶子, 然后迅速放入黑色塑料袋中, 0.5 h内带回实验室。1.3 研究方法
1.3.1 木质部导管长度的测量选取来自不同个体树的6根枝条, 通过空气注入法测量木质部导管长度(Cohen et al., 2003)。将枝条顶端浸入水中, 通过空气压缩机从枝条基部注入空气(约0.12 MPa), 将枝条顶端流出的气泡收集在注满水的量筒中, 测定通过木质部导管的空气流量(Q)(Cohen et al., 2003)。依次从枝条上切割2 cm的茎段, 并且依次测量出不同茎段长度下的空气流量(Q)。根据Cohen理论:
其中, x是测量空气流量(Qx)时的枝条长度。枝条的平均导管长度L为k的绝对值的2倍。根据Cohen理论, 估计导管长度的概率分布函数(Px)。
1.3.2 栓塞脆弱性的测定
该试验诱导栓塞的方法主要是自然干燥法、Sperry离心法和Cochard Cavitron离心机法。
1.3.2.1 自然干燥法测定VC 将采集的枝条带回实验室, 置于实验台上进行自然脱水干燥。每个枝条上选区3个叶片, 用铝箔包覆1 h使其蒸腾降至最低, 然后采下叶片用压力室法测定其水势, 将3个叶片水势的平均值作为该枝条的水势。该枝条的导水率(Kh)用低压液流计测量, 从测过叶片水势的枝条部位分别取3段5 cm的茎段, 测出其Kh和最大导水率(Kmax), 则该茎段的木质部导水率损失百分比(PLC)为:
随着枝条脱水干燥, 木质部水势逐渐降低, 而PLC则随之逐渐增大。根据不同水势所对应的PLC, 进行威布尔曲线拟合, 得到枝条的VC。为了更好地进行曲线拟合, 采用了单威布尔累计分布函数(3)或双威布尔累计分布函数(4):
式中, T为张力, B和C为威布尔常量,α为双威布尔分布中第一组分所占的比例枝条PLC为50%的水势P50为:
选取6个来自不同植株的枝条进行测定, 其平均值即为木质部的P50。
1.3.2.2 Sperry离心机法测定VC 6:00-7:00采回实验室的枝条, 浸入装水的容器中, 在水下用刀片切掉枝条叶柄后, 从中部截取27.4 cm或14.4 cm的茎段, 用低压流量计(LPFM)法测其Kh; 在0.159 MPa的压力下用100 mmol·L-1除气KCl溶液冲洗4 min, 使枝条中的栓塞化导管恢复水力学功能, 用低压液流计测枝条的Kmax; 然后将枝段置入Sperry离心机中, 设定离心机转速, 使枝条中部(即转轴处)的张力(tension, T)为0.1 MPa, 离心15 min后从离心机取出, 用LPFM测其Kmax; 重复该步骤, 并逐渐增大离心机转速, 用LPFM测量不同T下诱导栓塞后的Kh。用公式(3)、(4)和(5)建立27.4 cm和14.4 cm茎段的VC。选取6个来自不同植株的枝条进行测定。
1.3.2.3 Cochard Cavitron离心机法测定VC 枝条采集和处理同1.3.2.2, 截取27.4 cm茎段放入Cochard Cavitron 离心机, T在0.1 MPa时离心30 min, 测得其Kmax, 之后逐渐增大T, 每个转速下等待2 min后测定其Kh。用公式(3)、(4)和(5)建立木质部栓塞脆弱性曲线和P50值。选取6个来自不同植株上的枝条进行测定。
1.3.2.4 测量比导率(Ks) 用Sperry离心机离心前, 测量枝条基部的直径(d), 计算该基部的面积(S), 本实验用其测量出的不同水势下的Kh和基部的面积来测量其相应水势下的Ks。选取6个来自不同植株上的枝条进行测定。
1.3.2.5 栓塞修复的测定 用Sperry离心机法测定VC后, 枝条PLC接近100%, 即木质部导管几乎完全栓塞。将枝条重新冲洗, 测量冲洗后的比导率, 并与Sperry离心机法离心之前的初始最大比导率作比较, 分析冲洗处理对枝条栓塞的修复效应。
2 结果和分析
2.1 刺槐木质部茎段端开口导管数
空气注入法测定出刺槐的导管长度为(20.020 ± 1.835) cm, 根据Cohen理论, 长14.4 cm的茎段所含开口导管的概率为(0.032 ± 0.002)%, 而长27.4 cm的茎段所含开口导管的概率为(0.019 ± 0.002)% (图1)。尽管27.4 cm和14.4 cm的刺槐茎段所含有的开口导管数上有显著性的差异(p < 0.05), 但采用Sperry离心机法在两种长度茎段上建立的VC无明显差异(图2), 它们的P50差异不显著(表1)。图1
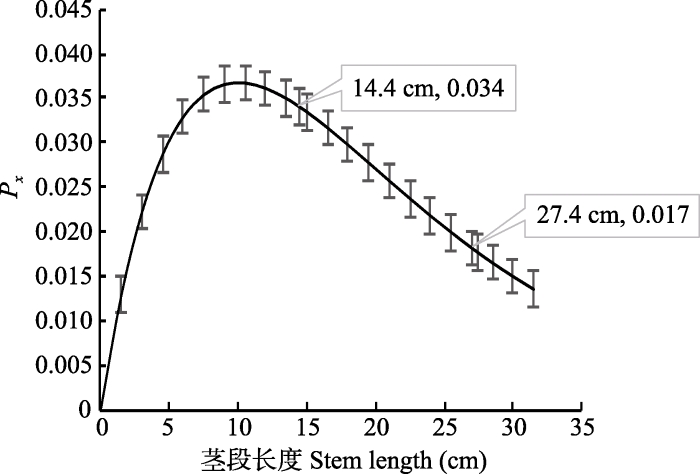 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1刺槐的导管在长度为x区间内的概率(Px)与茎段长度(x)建立的关系(平均值±标准偏差)。
Fig. 1The probability that the xylem conduit is within the length x interval (Px) and stem length (x) derived from the xylem conduit length distribution of Robinia pseudoacacia (mean ± SD).
Table 1
表1
表1基于不同方法计算出的刺槐导水率损失50%时的张力(P50)
Table 1
| 栓塞脆弱性曲线的建立方法 Methods in establishing embolism vulnerability curves | P50 (MPa) | 样品数 Number of samples | 茎段长度 Stem length (cm) | 来源 Source |
|---|---|---|---|---|
| Sperry离心机法 Sperry centrifugation method | -0.93 ± 0.021a | 6 | 27.4 | |
| -0.92 ± 0.058a | 6 | 14.4 | ||
| 自然干燥法 Bench top dehydration method | -2.91b | 31 | ||
| Cochard Cavitron 离心机法 Cochard Cavitron centrifugation method | -0.38 ± 0.044c | 6 | 27.4 | |
| -0.46 ± 0.030d | 6 | 27.4 | Dang et al., 2017 | |
| -0.22 ± 0.026e | 6 | 27.4 | Li et al., 2016 |
新窗口打开|下载CSV
图2
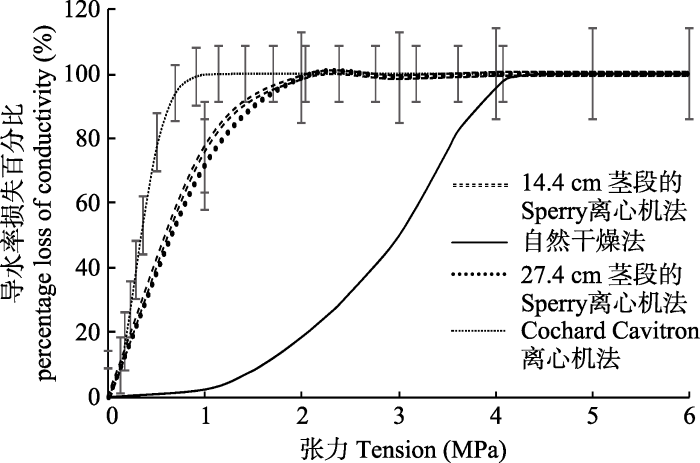 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2用不同方法建立的刺槐栓塞脆弱性曲线(平均值±标准偏差)。
Fig. 2Vulnerability curves of Robinia pseudoacacia established by different methods (mean ± SD).
2.2 刺槐木质部栓塞脆弱性
2.2.1 导水率对比3种方法在相同长度的茎段上建立的VC曲线, 其P50大小顺序为自然干燥法> Sperry离心机法> Cochard Cavitron离心机法(表1)。用自然干燥法测得的VC为“s”形, 水势低于-0.8 MPa后, PLC开始增加, 直到水势达到-4 MPa, PLC达到100% (图2)。Cochard Cavitron离心机法和Sperry离心机法建立的栓塞脆弱性曲线为“r”形(图2), 刺槐对木质部栓塞非常敏感, PLC随着水势降低而迅速增加, 直到水势大约等于-2 MPa时, PLC达到100%。因此, 作为诱导植物栓塞常用的3种方法, 使刺槐表现出的抗栓塞能力也不一样, 其中自然干燥法测得的刺槐VC表现出的抗栓塞能力最强, Sperry离心机法次之, Cochard Cavitron离心机法最小。对这3种方法进行差异显著性分析, 结果表明“s”形和“r”形VC的P50值差异极显著; 虽然Cochard Cavitron离心机法和Sperry离心机法均得到“r”形VC, 但它们的P50值也表现出极显著的差异(表1)。
2.2.2 比导率
两种不同长度的茎段的最大比导率(Ks,max)并不相同(图3), 且长27.4 cm的茎段的Ks,max显著大于长14.4 cm的茎段的Ks,max, 但是Ks随水势变化的规律相似, 均随水势的降低而减小(图3)。综合Sperry离心机法在两种不同长度茎段测得VC的对比, 以及比导率的比较, 可知开口导管数可能与栓塞性差异无关。
图3
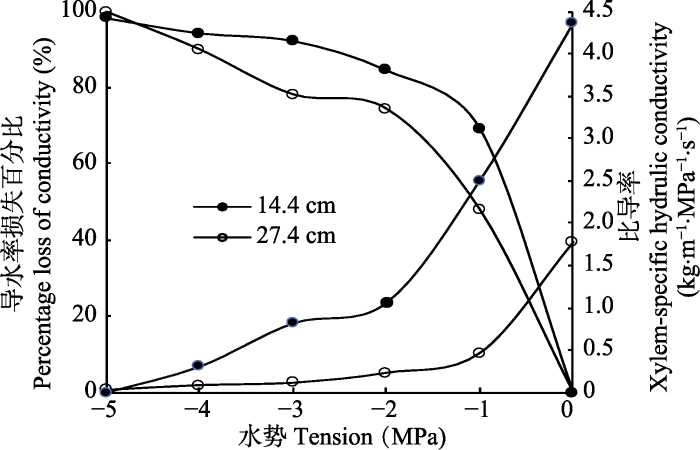 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3Sperry离心机法测得的不同长度刺槐枝条的栓塞脆弱性。
Fig. 3Vulnerability to cavitation as determined using a centrifuge-based method for Robinia pseudoacacia of different sample lengths.
2.3 刺槐木质部栓塞修复
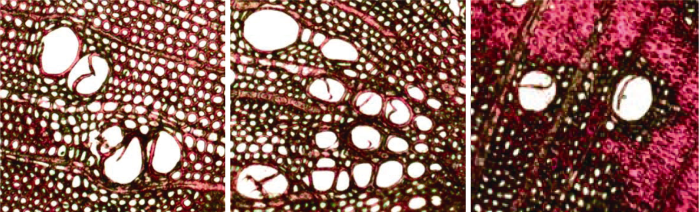
在-5 MPa下离心后, 长27.4 cm和14.4 cm的茎段的Ks均远小于离心前的Ks, 而通过冲洗对栓塞进行恢复后, 测得Ks与离心前的Ks差异不显著(图4)。两种不同长度的茎段的Ks,max存在差异, 27.4 cm茎段的Ks,max显著大于14.4 cm茎段的Ks,max (图4), 显微结构分析发现, 用于Sperry离心机法测量的14.4 m茎段中存在侵填体(图5), 这可能是导致两种不同长度样品Ks,max差异的原因之一。图4
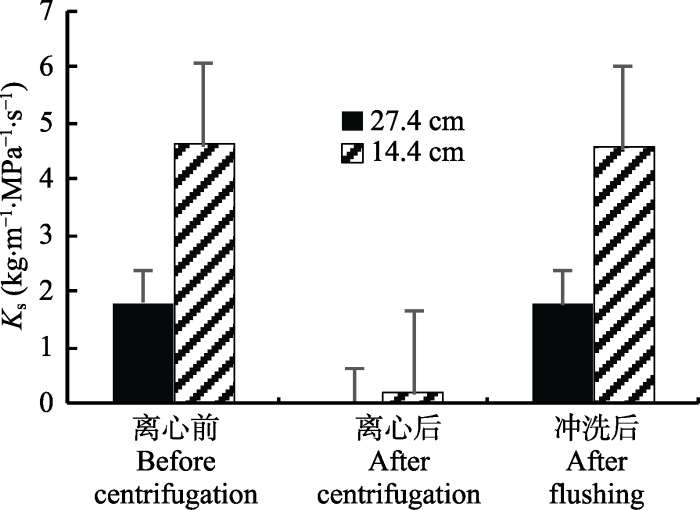 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4刺槐在离心前、离心后以及冲洗后的比导率(Ks)(平均值±标准偏差)。
Fig. 4Specific hydraulic conductivity (Ks) of Robinia pseudoacacia before centrifugation, after centrifugation, and after flushing (mean ± SD).
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5用荧光显微镜观察刺槐枝条横切面的三个视野。
Fig. 5Three fields of view for cross section of Robinia pseudoacacia stem under Fluorescence Microscopy.
3 讨论和结论
刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (Wang et al., 2014; 李荣等, 2016)的结果一致, 并且通过近几年报道的各物种的导管长度的数据来看, 刺槐的导管长度比大多数物种长(Jacobsen et al., 2014; 李荣等, 2016)。根据Cohen理论, 两种长度枝条的开口导管概率数分别为0.019%和0.032%, 如果Sperry离心机法在测定刺槐枝条时存在“开口导管假象” (Cochard et al., 2010; Choat et al., 2010), 那么两种不同长度枝条的VC应该有显著差异, 然而试验结果不支持该观点(图1)。近年来, 有研究在测定长导管植物VC时未发现“开口导管假象”(Jacobsen and Pratt, 2012; Sperry, et al., 2012)。例如, Sperry等(2012)用离心机法建立长导管植物Quercus gambelii和其他物种短茎段和长茎段的栓塞脆弱性曲线并没有差别。同时Jacobsen和Pratt (2012)用相同的方法建立的葡萄(Vitis vinifera)的VC, 发现其P50值也无明显差异。相比较而言,Cochard Cavitron离心机法可能更容易出现“开口导管假象” (Cochard et al., 2005)。本研究表明, 自然干燥法、Cochard Cavitron离心机法、Sperry离心机法建立刺槐的VC有较大差异。自然干燥建立的栓塞脆弱性曲线为“s”形, 而Cochard Cavitron离心机法和Sperry离心机法建立栓塞脆弱性曲线为“r”形, 这与Sperry等(2012)提出刺槐的栓塞脆弱性曲线为“r”形曲线相符, 这还与李荣等(2016)、党维等(2017)得到刺槐的脆弱性曲线一致。导致这一结果可能有两个原因:
一是自然干燥法、Sperry离心机法和Cochard Cavitron离心机法诱导栓塞的方式不同。自然干燥法是最简单和最直接诱导植物栓塞和空穴化的方法, 即通过离体枝条内水分的自然散失来诱导栓塞, 在不同栓塞程度下测定该枝条的导水率损失, 同时通过测量叶片或小枝的水势从而获取枝条木质部的水势。因此自然干燥法的栓塞形成过程比较漫长, 枝条各个区段处于相近的水势, 相互连通的导管之间保持水力学平衡。而离心机法建立VC时, 枝条两端位置的张力为0, 而中轴处张力最大, 即整个枝条的水势分布不均匀, 这一方面是枝条的中部更容易栓塞, 另一方面尤其在较高转速时, 气泡可能会因为水力学不平衡更容易进入导管。“气种”假说是一种植物发生栓塞的机理, 即木质部导管内栓塞的发生是外界空气或已经栓塞的导管内的空气泡, 通过导管间纹孔膜上的微孔扩散至相邻导管内, 水分蒸发导致连续水柱断裂, 随之空气泡的体积不断增加, 促使管道内的压力瞬时升高, 纹孔膜上的微孔被阻塞, 最终空气泡的体积充满整个导管, 使木质部输水能力下降(Zimmermann, 1983)。
二是与不同方法中的具体操作有关。用自然干燥法建立一个栓塞脆弱性曲线需要一个星期或者更长, 根据公式(2), PLC的确定需要冲洗枝条, 将冲洗后的导水率作为Kmax。但是本实验中自然干燥法所测的冲洗后的导水率即Kmax存在差异, 并且所测的Kmax值随着时间的推移, 出现下降的趋势。这可能是由于在自然干燥脱水过程中, 刺槐一部分栓塞化的木质部导管无法通过冲洗而得到恢复(Trifilò et al., 2015; 党维等, 2017)。同时随着干燥脱水时间的加长, 刺槐木质部导管中会产生侵填体, 这也是导致Kh下降的原因。故无法对冲洗后的Kmax进行准确测定, 导致其实际的PLC被低估。因此, 由自然干燥法建立刺槐的栓塞脆弱性曲线计算出的P50可能大于刺槐实际的P50。离心机法则是在很短时间内就能得到一条栓塞脆弱性曲线, 避免了刺槐样品随着检测时间的增加而出现的栓塞。但在离心力的不断加大的情况下, 其木质部的张力降低、木质部导管内的小气泡将不断向枝条中部富集(Wang et al., 2014), 这将低估其实际的导水率, 导致计算出的P50偏小。本研究涉及的两种离心机方法亦有所不同。Cochard Cavitron离心机法在离心的同时通过观察枝条两端的液面高度差测得Kh和PLC (Cochard 2002; Cochard et al., 2005)。但是随着离心机转速的增加, 枝条两端的液面位置变得不稳定, 这将导致测量出的导水率与实际有较大的偏差。这也是很多****提出Cochard Cavitron离心机法不适合长导管植物VC检测的原因(李荣等, 2015)。Sperry离心机法则是将栓塞诱导和Kh测定分为两步, 即先通过离心来诱导栓塞, 再用LPFM测量其导水率。在LPFM测量过程中枝条的顶端连接天平的乳胶管中有气泡产生, 说明部分栓塞化的导管在LPFM检测时得到了恢复, 这将会造成测量的导水率比实际的导水率要高。
综上所述, Cochard Cavitron离心机法在长导管植物的VC检测时存在“开口导管假象”, 而Sperry离心机法测得的结果更为可靠; 自然干燥法则可能低估了长导管植物的栓塞脆弱性。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1038/s41559-017-0248-xURLPMID:29046541 [本文引用: 1]

Abstract Widespread tree mortality associated with drought has been observed on all forested continents and global change is expected to exacerbate vegetation vulnerability. Forest mortality has implications for future biosphere-atmosphere interactions of carbon, water and energy balance, and is poorly represented in dynamic vegetation models. Reducing uncertainty requires improved mortality projections founded on robust physiological processes. However, the proposed mechanisms of drought-induced mortality, including hydraulic failure and carbon starvation, are unresolved. A growing number of empirical studies have investigated these mechanisms, but data have not been consistently analysed across species and biomes using a standardized physiological framework. Here, we show that xylem hydraulic failure was ubiquitous across multiple tree taxa at drought-induced mortality. All species assessed had 60% or higher loss of xylem hydraulic conductivity, consistent with proposed theoretical and modelled survival thresholds. We found diverse responses in non-structural carbohydrate reserves at mortality, indicating that evidence supporting carbon starvation was not universal. Reduced non-structural carbohydrates were more common for gymnosperms than angiosperms, associated with xylem hydraulic vulnerability, and may have a role in reducing hydraulic function. Our finding that hydraulic failure at drought-induced mortality was persistent across species indicates that substantial improvement in vegetation modelling can be achieved using thresholds in hydraulic function.
[本文引用: 1]
DOI:10.1111/j.1365-3040.2010.02160.xURLPMID:20444217 [本文引用: 2]
Among woody plants, grapevines are often described as highly vulnerable to water-stress induced cavitation with emboli forming at slight tensions. However, we found native embolism never exceeded 30% despite low xylem water potentials (Ψx) for stems of field grown vines. The discrepancy between native embolism measurements and those of previous reports led us to assess vulnerability curve generation using four separate methods and alterations (i.e. segment length and with/without flushing to remove embolism prior to measurement) of each. Centrifuge, dehydration and air-injection methods, which rely on measurement of percentage loss of hydraulic conductivity (PLC) in detached stems, were compared against non-invasive monitoring of xylem cavitation with nuclear magnetic resonance (NMR) imaging. Short segment air-injection and flushed centrifuge stems reached >90 PLC at Ψx of-0.5 and 611.5 MPa, respectively, whereas dehydration and long-segment air-injection measurements indicated no significant embolism at Ψx > 612.0 MPa. Observations from NMR agreed with the dehydration and long segment air-injection methods, showing the majority of vessels were still water-filled at Ψx > 611.5 MPa. Our findings show V. vinifera stems are far less vulnerable to water stress-induced cavitation than previously reported, and dehydration and long segment air-injection techniques are more appropriate for long-vesseled species and organs.
DOI:10.1111/j.1365-3040.2005.01336.xURL [本文引用: 1]
Air seeding threshold ( P a ) of xylem vessels from current year growth rings were measured along the vertical axis of mature sugar maple trees ( Acer saccharum Marsh.), with sampling points in primary leaf veins, petioles, 1-, 3-, and 7-year-old branches, large branches, the trunk and roots. The air seeding threshold was taken as the pressure required to force nitrogen gas through intervessel pit membranes. Although all measurements were made on wood produced in the same year, P a varied between different regions of A. saccharum , with distal organs such as leaves and petioles having lower P a than basal regions. Mean (SE) P a ranged from 1.0 ( 0.1) MPa in primary leaf veins to 4.8 ( 0.1) MPa in the main trunk. Roots exhibited a P a of 2.8 ( 0.2) MPa, lower than all other regions of the tree except leaf veins and petioles. Mean xylem vessel diameter increased basipetally, with the widest vessels occurring in the trunk and roots. Within the shoot, wider vessels had greater air seeding thresholds, contrasting with trends previously reported. However, further experimentation revealed that differences in P a between regions of the stem were driven by the presence of primary xylem conduits, rather than differences in vessel diameter. In 1-year-old branches, P a was significantly lower in primary xylem vessels than in adjacent secondary xylem vessels. This explained the lower values of P a measured in petioles and leaf veins, which possessed a greater ratio of primary xylem to secondary xylem than other regions. The difference in P a between primary and secondary xylem was attributed to the greater area of primary cell wall (pit membrane) exposed in primary xylem conduits with helical or annular thickening.
DOI:10.1046/j.1365-3040.2002.00863.xURL [本文引用: 2]
A technique for measuring hydraulic conductances of excised xylem segments exposed to high negative pressures is described. A centrifugal force is used to generate negative pressures ( P ) in the sample and to create a positive hydrostatic pressure difference (Δ P ) between its two ends. Δ P forces water through the sample at a flow rate ( F ) determined optically during centrifugation. The sample hydraulic conductance k is derived from F and Δ P . The sample vulnerability curve is given by the dependence of k on P . Results for Cedrus atlantica Manetti and Laurus nobilis L. shoots are given. The technique is appropriate for the analysis of xylem refilling under negative pressure.
DOI:10.1093/jxb/ert193URLPMID:23888067 [本文引用: 1]
Abstract Xylem cavitation resistance has profound implications for plant physiology and ecology. This process is characterized by a 'vulnerability curve' (VC) showing the variation of the percentage of cavitation as a function of xylem pressure potential. The shape of this VC varies from 'sigmoidal' to 'exponential'. This review provides a panorama of the techniques that have been used to generate such a curve. The techniques differ by (i) the way cavitation is induced (e.g. bench dehydration, centrifugation, or air injection), and (ii) the way cavitation is measured (e.g. percentage loss of conductivity (PLC) or acoustic emission), and a nomenclature is proposed based on these two methods. A survey of the literature of more than 1200 VCs was used to draw statistics on the usage of these methods and on their reliability and validity. Four methods accounted for more than 96% of all curves produced so far: bench dehydration-PLC, centrifugation-PLC, pressure sleeve-PLC, and Cavitron. How the shape of VCs varies across techniques and species xylem anatomy was also analysed. Strikingly, it was found that the vast majority of curves obtained with the reference bench dehydration-PLC method are 'sigmoidal'. 'Exponential' curves were more typical of the three other methods and were remarkably frequent for species having large xylem conduits (ring-porous), leading to a substantial overestimation of the vulnerability of cavitation for this functional group. We suspect that 'exponential' curves may reflect an open-vessel artefact and call for more precautions with the usage of the pressure sleeve and centrifugation techniques.
DOI:10.1111/j.1399-3054.2005.00526.xURL [本文引用: 3]
A new technique for generating xylem cavitation and vulnerability curves was evaluated. The centrifugal force was used to lower the negative pressure in a xylem segment and to induce a positive pressure difference between sample's ends. This enabled the determination of sample hydraulic conductance during centrifugation and, hence, its variation with decreasing xylem pressures. The centrifuge technique was compared with standard methods on a large number of species including conifers, diffuse-porous and ring-porous woody angiosperms. A very good agreement was found for coniferous and diffuse-porous species. However, the technique was not appropriate for ring-porous species, probably because many vessels were cut open in the centrifuged xylem segments. The main advantage of this technique is its rapidity, the vulnerability curve of a xylem segment being constructed typically in less than half an hour. This will greatly facilitate the study of xylem cavitation in ecological or genetic researches.
DOI:10.1111/j.1365-3040.2010.02163.xURLPMID:20444214 [本文引用: 2]
The Cavitron spinning technique is used to construct xylem embolism vulnerability curves (VCs), but its reliability has been questioned for species with long vessels. This technique generates two types of VC: sigmoid ‘s’-shaped and exponential, levelling-off ‘r’-shaped curves. We tested the hypothesis that ‘r’-shaped VCs were anomalous and caused by the presence of vessels cut open during sample preparation. A Cavitron apparatus was used to construct VCs from samples of different lengths in species with contrasting vessel lengths. The results were compared with VCs obtained using other independent techniques. When vessel length exceeded sample length, VCs were ‘r’-shaped and anomalous. Filling vessels cut open at both ends with air before measurement produced more typical ‘s’-shaped VCs. We also found that exposing segments of 11 woody species in a Cavitron at the pressure measured in planta before sampling considerably increased the degree of embolism above the native state level for species with long vessels. We concluded that open vessels were abnormally more vulnerable to cavitation than intact vessels. We recommend restricting this technique to species with short conduits. The relevance of our conclusions for other spinning techniques is discussed.
DOI:10.1093/jxb/erg202URLPMID:12815034 [本文引用: 2]
Studies showing that rootstock dwarfing potential is related to plant hydraulic conductance led to the hypothesis that xylem properties are also related. Vessel length distribution and other properties of apple wood from a series of varieties were measured using the 'air method' in order to test this hypothesis. Apparatus was built to measure and monitor conductivity to air of fresh wood segments of different lengths. Theory for determining vessel length distribution was improved to give a single parameter uni-modal vessel length probability density function.
URL [本文引用: 1]
随着全球气候变化,气温上升,干旱发生的可能性增加,干旱导致树木死亡现象的发生也越来越普遍,但干旱导致树木死亡确切的生理学机制还未完全解决,目前仍是研究的热点。近来提出的干旱导致树木死亡的假说主要有:水力学失败假说,认为干旱导致的低水势阻碍植物体内水分的长距离运输,使植物因组织脱水干燥而死亡;碳饥饿假说,认为干旱过程中发生的气孔关闭会限制碳摄取,导致植物体内可利用碳的耗竭;生物攻击假说,认为干旱中的植物易受生物因子(昆虫和病原体)的攻击而死亡。3种假说各自都能解释一定的现象,但又都存在缺陷,主要问题是不能解释所有干旱导致树木死亡的现象。近来的研究越来越关注这些机制之间的共同作用或相互作用,水力学失败、碳饥饿和生物因子之间会相互影响而加重干旱对植物的作用,尤其是水力学失败和碳饥饿相耦合已得到越来越多的认可。本文主要综述这几种致死机制的试验证据及存在的问题,同时综述当前研究存在的局限性,并讨论了未来可能的研究方向。
URL [本文引用: 1]
随着全球气候变化,气温上升,干旱发生的可能性增加,干旱导致树木死亡现象的发生也越来越普遍,但干旱导致树木死亡确切的生理学机制还未完全解决,目前仍是研究的热点。近来提出的干旱导致树木死亡的假说主要有:水力学失败假说,认为干旱导致的低水势阻碍植物体内水分的长距离运输,使植物因组织脱水干燥而死亡;碳饥饿假说,认为干旱过程中发生的气孔关闭会限制碳摄取,导致植物体内可利用碳的耗竭;生物攻击假说,认为干旱中的植物易受生物因子(昆虫和病原体)的攻击而死亡。3种假说各自都能解释一定的现象,但又都存在缺陷,主要问题是不能解释所有干旱导致树木死亡的现象。近来的研究越来越关注这些机制之间的共同作用或相互作用,水力学失败、碳饥饿和生物因子之间会相互影响而加重干旱对植物的作用,尤其是水力学失败和碳饥饿相耦合已得到越来越多的认可。本文主要综述这几种致死机制的试验证据及存在的问题,同时综述当前研究存在的局限性,并讨论了未来可能的研究方向。
DOI:10.11707/j.1001-7488.20170306URL [本文引用: 3]
【目的】植物木质部栓塞修复能力已成为植物抗旱性研究的热点。本研究探讨6个耐旱树种木质部水力特征及与栓塞修复能力之间的关系,以及决定栓塞修复能力的主要影响因子,为筛选抗旱树种提供理论依据。【方法】以西北农林科技大学博览园内生长良好的6个耐旱树种(刺槐、沙棘、榆树、元宝枫、旱柳、榛)的1年生枝为研究对象,采用Cochard Cavitron离心机诱导木质部栓塞,构建6个树种的木质部栓塞脆弱曲线,得到其栓塞脆弱性(P50)值,同时测定枝条的木质部水力直径(Dh)、黎明前木质部水势及自然状态下栓塞程度(NPLC),利用低压液流计及染色法测定已栓塞枝条不同时段(20,40,60,80,100 min)的栓塞修复程度(1-PLC),进而探究不同树种木质部栓塞修复能力与栓塞脆弱性、水力直径、黎明前水势和NPLC间的关系。【结果】1)6个树种栓塞脆弱性大小依次为刺槐榆树沙棘旱柳榛元宝枫,其中元宝枫的抗栓塞能力最强;水力直径依次为刺槐榆树榛沙棘旱柳元宝枫。2)用染色法测得的木质部栓塞修复程度与低压液流计测得的木质部栓塞修复程度相一致(回归方程是y=1.047x-7.567,R~2=0.863);低压液流计法测定的值完全可以反映在不同时间段栓塞枝条的修复状况。3)6个树种1年生枝木质部栓塞修复能力依次为刺槐榆树沙棘旱柳榛元宝枫,植物重新供水后,木质部栓塞修复能力强的树种能迅速恢复栓塞导管的输水功能;栓塞修复能力与栓塞脆弱性大小顺序完全一致,呈显著的正相关(R~2=0.980),即木质部栓塞脆弱性大的树种,其栓塞修复能力也越强;回归分析表明,6个树种木质部水力直径、黎明前水势和NPLC与栓塞修复能力有很强的相关性(R~2分别为0.868,0.740,0.741)。【结论】植物木质部水力特征与栓塞修复能力密切相关,特别是木质部栓塞脆弱性(P50)和水力直径与栓塞修复能力有很强烈的正相关,是决定栓塞修复能力的主要影响因子。
DOI:10.11707/j.1001-7488.20170306URL [本文引用: 3]
【目的】植物木质部栓塞修复能力已成为植物抗旱性研究的热点。本研究探讨6个耐旱树种木质部水力特征及与栓塞修复能力之间的关系,以及决定栓塞修复能力的主要影响因子,为筛选抗旱树种提供理论依据。【方法】以西北农林科技大学博览园内生长良好的6个耐旱树种(刺槐、沙棘、榆树、元宝枫、旱柳、榛)的1年生枝为研究对象,采用Cochard Cavitron离心机诱导木质部栓塞,构建6个树种的木质部栓塞脆弱曲线,得到其栓塞脆弱性(P50)值,同时测定枝条的木质部水力直径(Dh)、黎明前木质部水势及自然状态下栓塞程度(NPLC),利用低压液流计及染色法测定已栓塞枝条不同时段(20,40,60,80,100 min)的栓塞修复程度(1-PLC),进而探究不同树种木质部栓塞修复能力与栓塞脆弱性、水力直径、黎明前水势和NPLC间的关系。【结果】1)6个树种栓塞脆弱性大小依次为刺槐榆树沙棘旱柳榛元宝枫,其中元宝枫的抗栓塞能力最强;水力直径依次为刺槐榆树榛沙棘旱柳元宝枫。2)用染色法测得的木质部栓塞修复程度与低压液流计测得的木质部栓塞修复程度相一致(回归方程是y=1.047x-7.567,R~2=0.863);低压液流计法测定的值完全可以反映在不同时间段栓塞枝条的修复状况。3)6个树种1年生枝木质部栓塞修复能力依次为刺槐榆树沙棘旱柳榛元宝枫,植物重新供水后,木质部栓塞修复能力强的树种能迅速恢复栓塞导管的输水功能;栓塞修复能力与栓塞脆弱性大小顺序完全一致,呈显著的正相关(R~2=0.980),即木质部栓塞脆弱性大的树种,其栓塞修复能力也越强;回归分析表明,6个树种木质部水力直径、黎明前水势和NPLC与栓塞修复能力有很强的相关性(R~2分别为0.868,0.740,0.741)。【结论】植物木质部水力特征与栓塞修复能力密切相关,特别是木质部栓塞脆弱性(P50)和水力直径与栓塞修复能力有很强烈的正相关,是决定栓塞修复能力的主要影响因子。
DOI:10.1098/rstb.1895.0012URL [本文引用: 1]
DOI:10.1007/s004680100095URL [本文引用: 1]
DOI:10.5846/stxb201304270839URL [本文引用: 1]
植物在生长发育过程中受众多环境因子共同作用.随着全球气候变 化,气温升高、降水量下降等问题频繁出现.目前气象学家一致预测未来环境变暖会使干旱更加频繁剧烈,这一环境改变使植物死亡更加严重.植物在水分胁迫、特 别是干旱胁迫条件下,体内水分代谢与碳代谢会发生失衡现象:光合速率降低、蒸腾速率降低,带来生长降低;为维持植物新陈代谢,植物呼吸作用必然下调.在长 期干旱胁迫条件下植物体内碳水化合物储存发生失衡现象,这种失衡使植物陷入碳饥饿现象.另外,由于水分失衡而出现的木质部栓塞和空穴会进一步加剧水分运输 障碍,而修复空穴则需要大量非结构性碳水化合物(NSC),这使植物陷入两难选择.总结了植物干旱胁迫下,碳饥饿与水分代谢、植物死亡关系的相关研究,对 未来的研究方向和重点提出建议,以期对未来的植物死亡研究提供帮助.
DOI:10.5846/stxb201304270839URL [本文引用: 1]
植物在生长发育过程中受众多环境因子共同作用.随着全球气候变 化,气温升高、降水量下降等问题频繁出现.目前气象学家一致预测未来环境变暖会使干旱更加频繁剧烈,这一环境改变使植物死亡更加严重.植物在水分胁迫、特 别是干旱胁迫条件下,体内水分代谢与碳代谢会发生失衡现象:光合速率降低、蒸腾速率降低,带来生长降低;为维持植物新陈代谢,植物呼吸作用必然下调.在长 期干旱胁迫条件下植物体内碳水化合物储存发生失衡现象,这种失衡使植物陷入碳饥饿现象.另外,由于水分失衡而出现的木质部栓塞和空穴会进一步加剧水分运输 障碍,而修复空穴则需要大量非结构性碳水化合物(NSC),这使植物陷入两难选择.总结了植物干旱胁迫下,碳饥饿与水分代谢、植物死亡关系的相关研究,对 未来的研究方向和重点提出建议,以期对未来的植物死亡研究提供帮助.
DOI:10.1111/nph.13017URLPMID:25229841
Summary The standard centrifuge method has been frequently used to measure vulnerability to xylem cavitation. This method has recently been questioned. It was hypothesized that open vessels lead to exponential vulnerability curves, which were thought to be indicative of measurement artifact. We tested this hypothesis in stems of olive ( Olea europea ) because its long vessels were recently claimed to produce a centrifuge artifact. We evaluated three predictions that followed from the open vessel artifact hypothesis: shorter stems, with more open vessels, would be more vulnerable than longer stems; standard centrifuge-based curves would be more vulnerable than dehydration-based curves; and open vessels would cause an exponential shape of centrifuge-based curves. Experimental evidence did not support these predictions. Centrifuge curves did not vary when the proportion of open vessels was altered. Centrifuge and dehydration curves were similar. At highly negative xylem pressure, centrifuge-based curves slightly overestimated vulnerability compared to the dehydration curve. This divergence was eliminated by centrifuging each stem only once. The standard centrifuge method produced accurate curves of samples containing open vessels, supporting the validity of this technique and confirming its utility in understanding plant hydraulics. Seven recommendations for avoiding artefacts and standardizing vulnerability curve methodology are provided.
DOI:10.1111/j.1469-8137.2012.04118.xURLPMID:22448870 [本文引用: 2]
090004Vulnerability to cavitation curves are used to estimate xylem cavitation resistance and can be constructed using multiple techniques. It was recently suggested that a technique that relies on centrifugal force to generate negative xylem pressures may be susceptible to an open vessel artifact in long-vesselled species.090004Here, we used custom centrifuge rotors to measure different sample lengths of 1-yr-old stems of grapevine to examine the influence of open vessels on vulnerability curves, thus testing the hypothesized open vessel artifact. These curves were compared with a dehydration-based vulnerability curve.090004Although samples differed significantly in the number of open vessels, there was no difference in the vulnerability to cavitation measured on 0.14- and 0.271-m-long samples of Vitis vinifera. Dehydration and centrifuge-based curves showed a similar pattern of declining xylem-specific hydraulic conductivity (Ks) with declining water potential. The percentage loss in hydraulic conductivity (PLC) differed between dehydration and centrifuge curves and it was determined that grapevine is susceptible to errors in estimating maximum Ks during dehydration because of the development of vessel blockages.090004Our results from a long-vesselled liana do not support the open vessel artifact hypothesis.
DOI:10.3120/0024-9637-61.4.317URL [本文引用: 1]
DOI:10.17521/cjpe.2015.0260URL [本文引用: 4]
木质部栓塞脆弱性对干旱响应的研究已成为全球气候变化背景下的热点和重要内容。该文以6个耐旱树种刺槐(Robinia pseudoacacia)、沙棘(Hippophae rhamnoides)、榆树(Ulmus pumila)、元宝枫(Acer truncatum)、旱柳(Salix matsudana)、榛(Corylus heterophylla)为研究对象,采用Cochard Cavitron离心机技术建立木质部栓塞脆弱曲线,计算木质部栓塞脆弱性,利用染色法、硅胶注射法等测定木质部导管直径、导管内径跨度、导管连接度、导管密度、导管长度和木质部密度,探究木质部结构与栓塞脆弱性的关系,区分6个耐旱树种木质部结构在抗栓塞性上的差异,以期建立6个耐旱树种在木质部结构方面的抗栓塞性指标。结果表明:6个耐旱树种木质部栓塞脆弱性大小为刺槐榆树沙棘旱柳元宝枫榛,其中,刺槐、沙棘和榆树的栓塞脆弱曲线为"r"形,而元宝枫、旱柳和榛的栓塞脆弱曲线为"s"形,脆弱曲线为"r"形的树种与脆弱曲线为"s"形的树种栓塞脆弱性差异极显著(p0.01)。线性分析表明:木质部结构影响各树种的栓塞脆弱性,其中,木质部密度影响最大(t=0.702),导管直径次之(t=0.532),导管长度影响最小(t=0.010)。
DOI:10.17521/cjpe.2015.0260URL [本文引用: 4]
木质部栓塞脆弱性对干旱响应的研究已成为全球气候变化背景下的热点和重要内容。该文以6个耐旱树种刺槐(Robinia pseudoacacia)、沙棘(Hippophae rhamnoides)、榆树(Ulmus pumila)、元宝枫(Acer truncatum)、旱柳(Salix matsudana)、榛(Corylus heterophylla)为研究对象,采用Cochard Cavitron离心机技术建立木质部栓塞脆弱曲线,计算木质部栓塞脆弱性,利用染色法、硅胶注射法等测定木质部导管直径、导管内径跨度、导管连接度、导管密度、导管长度和木质部密度,探究木质部结构与栓塞脆弱性的关系,区分6个耐旱树种木质部结构在抗栓塞性上的差异,以期建立6个耐旱树种在木质部结构方面的抗栓塞性指标。结果表明:6个耐旱树种木质部栓塞脆弱性大小为刺槐榆树沙棘旱柳元宝枫榛,其中,刺槐、沙棘和榆树的栓塞脆弱曲线为"r"形,而元宝枫、旱柳和榛的栓塞脆弱曲线为"s"形,脆弱曲线为"r"形的树种与脆弱曲线为"s"形的树种栓塞脆弱性差异极显著(p0.01)。线性分析表明:木质部结构影响各树种的栓塞脆弱性,其中,木质部密度影响最大(t=0.702),导管直径次之(t=0.532),导管长度影响最小(t=0.010)。
DOI:10.17521/cjpe.2015.0080URL [本文引用: 1]
木质部空穴化和栓塞是木本植物在干旱等条件下遭受水分胁迫时产生的木质部输水功能障碍,在全球气候变化的大背景下,栓塞脆弱性对干旱响应的研究已成为热点和重要内容。近年来有关木质部栓塞脆弱性与植物输水结构和耐旱性的关系已有大量研究并取得一定成果,但是,不同****在不同地区对不同材料的研究结果存在很大不同。该文就近年来这一研究领域取得的成果及争议问题进行了概括和总结,主要涉及木质部栓塞脆弱性(P50)及脆弱曲线的建立方法、木质部栓塞脆弱性与木质部结构(导管直径、导管长度、纹孔膜、木质部密度、纤维及纤维管胞)间的关系和木质部栓塞脆弱性与耐旱性的关系,并对未来工作进行展望,提出在未来的工作中应对同一树种使用Cochard Cavitron离心机法、Sperry离心机技术与传统方法建立的脆弱曲线进行比较验证、计算P50值、分析植物个体器官水平差异(根、茎、叶)、测定树种生理生态指标,探索植物栓塞脆弱性与输水结构和耐旱性的关系,从而评估不同类型植物在未来气候变化下的耐旱能力。
DOI:10.17521/cjpe.2015.0080URL [本文引用: 1]
木质部空穴化和栓塞是木本植物在干旱等条件下遭受水分胁迫时产生的木质部输水功能障碍,在全球气候变化的大背景下,栓塞脆弱性对干旱响应的研究已成为热点和重要内容。近年来有关木质部栓塞脆弱性与植物输水结构和耐旱性的关系已有大量研究并取得一定成果,但是,不同****在不同地区对不同材料的研究结果存在很大不同。该文就近年来这一研究领域取得的成果及争议问题进行了概括和总结,主要涉及木质部栓塞脆弱性(P50)及脆弱曲线的建立方法、木质部栓塞脆弱性与木质部结构(导管直径、导管长度、纹孔膜、木质部密度、纤维及纤维管胞)间的关系和木质部栓塞脆弱性与耐旱性的关系,并对未来工作进行展望,提出在未来的工作中应对同一树种使用Cochard Cavitron离心机法、Sperry离心机技术与传统方法建立的脆弱曲线进行比较验证、计算P50值、分析植物个体器官水平差异(根、茎、叶)、测定树种生理生态指标,探索植物栓塞脆弱性与输水结构和耐旱性的关系,从而评估不同类型植物在未来气候变化下的耐旱能力。
DOI:10.1890/02-0538URL [本文引用: 1]
The ability of plants to supply water to their leaves is intimately associated with survival. Water supply to leaves depends on maintaining an intact water column in the xylem from the roots to shoots. Because this hydraulic pathway is under tension, it is vulnerable to breakage through the induction of air emboli (cavitation). Although the physiological benefit of resistance to water-stress-induced xylem cavitation for desiccation tolerance is clear, there is considerable interspecific variation within and across climates. To understand the adaptive significance of this variation and the potential trade-off with water transport, we compiled a database of 167 species from 50 seed plant families and examined relationships among resistance to xylem cavitation, water transport capacity (as determined by the specific conductivity of xylem [Ks]), and climate. Relationships were evaluated using standard cross-species correlations (r). Because inferences about the adaptive significance of these correlations can be biased by the potential similarity of closely related species, we also analyzed our data using phylogenetically independent contrast correlations (PIC) calculated over a range of alternate seed plant phylogenies. Resistance to cavitation, expressed as the xylem tension at which 50% of hydraulic conductivity was lost ( 50), ranged from -0.18 to -9.9 MPa for angiosperms and from -1.5 to -14.1 MPa for conifers. Conifers were most resistant to cavitation, with mean 5080% more negative than angiosperms. In contrast, Kswas 270% higher in angiosperms than conifers. Across all species, cavitation resistance increased with decreasing mean annual precipitation. However, significant phylogenetically independent contrast correlations between 50and annual precipitation were found within the evergreen angiosperms and conifers but not in the deciduous angiosperms. Thus, the adaptive significance of increased resistance to cavitation as a mechanism of drought tolerance may be of primary importance in evergreen angiosperms and conifers. In contrast, analysis of independent contrasts indicated that Ksincreased with decreasing rainfall in deciduous angiosperms, whereas there was no association between Ksand water availability for evergreen angiosperms and conifers. These results suggest that the evolution of increased Ksmay be a critical adaptation to water limitation in deciduous angiosperms. Although there was a significant cross-species correlation between 50and Ks, this relationship was not supported by the independent contrast correlation, suggesting that the evolutionary basis for a trade-off between cavitation resistance and water transport capacity is weak.
DOI:10.1104/pp.102.012856URL [本文引用: 1]
DOI:10.1104/pp.113.233817URLPMID:24501002 [本文引用: 1]
Cavitation has long been recognized as a key constraint on the structure and functional integrity of the xylem. Yet, recent results call into question how well we understand cavitation in plants. Here, we consider embolism formation in angiosperms at two scales. The first focuses on how air-seeding occurs at the level of pit membranes, raising the question of whether capillary failure is an appropriate physical model. The second addresses methodological uncertainties that affect our ability to infer the formation of embolism and its reversal in plant stems. Overall, our goal is to open up fresh perspectives on the structure-function relationships of xylem.
[本文引用: 3]
DOI:10.1111/j.1365-3040.1988.tb01774.xURL [本文引用: 1]
Abstract . Hydraulic conductivity of the xylem is computed as the quotient of mass flow rate and pressure gradient. Measurements on excised plant stems can be difficult to interpret because of time-dependent reductions in flow rate, and because of variable degrees of embolism. Using Acer saccharum Marsh. stems, we found that certain perfusing solutions including dilute fixatives (e.g. 0.05% formaldehyde) and acids with pH below 3 (e.g. 10 mol m 3 oxalic) prevent long-term decline in conductivity. Xylem embolism can be quantified by expressing the initial conductivity as a percentage of the maximum obtained after flow-impeding air emboli have been removed by repeated high-pressure (175 kPa) flushes. Correlation between microbial contamination and declining conductivity suggests that long-term (> 4h) declines are caused by microbial growth within the vessels. Unpredictable trends in short-term (< 4h) measurements may be caused by movements of air emboli in vessels and/or participate matter.
DOI:10.1104/pp.88.3.581URLPMID:16666352 [本文引用: 2]
We investigated the hypothesis that water stress-induced xylem embolism is caused by air aspirated into functional vessels from neighboring embolized ones (e.g. embolized by physical damage) via pores in intervessel pit membranes. The following experiments with sugar maple (Acer saccharum Marsh.) support the hypothesis. (a) Most vessels in dehydrating stem segments embolized at xylem pressures < -3 megapascals; at this point the pressure difference across intervessel pits between air-filled vessels at the segment's ends and internal water-filled vessels was >3 megapascals. This same pressure difference was found to be sufficient to force air across intervessel pits from air injection experiments of hydrated stem segments. This suggests air entry at pits is causing embolism in dehydrating stems. (b) Treatments that increased the permeability of intervessel pits to air injection also caused xylem to embolize at less negative xylem pressures. Permeability was increased either by perfusing stems with solutions of surface tension below that of water or by perfusion with a solution of oxalic acid and calcium. The mechanism of oxalic-calcium action on permeability is unknown, but may relate to the ability of oxalate to chelate calcium from the pectate fraction of the pit membrane. (c) Diameter of pores in pit membranes measured with the scanning electron microscope were within the range predicted by hypothesis ( 0.4 micrometer).
[本文引用: 1]
DOI:10.1104/pp.114.249706URLPMID:25378693 [本文引用: 1]
Embolism resistance is a critically important trait for evaluating the ability of plants to survive and recover from drought periods and predicting future drought-induced forest decline (<a id="xref-ref-3-1" class="xref-bibr" href="#ref-3">Choat et al., 2012). However, recent publications have provided evidence that some measurement techniques used to evaluate the hydraulic function and vulnerability to cavitation of plant organs may be prone to artifacts (<a id="xref-ref-16-1" class="xref-bibr" href="#ref-16">Sperry et al., 2012; <a id="xref-ref-5-1" class="xref-bibr" href="#ref-5">Cochard et al., 2013; <a id="xref-ref-17-1" class="xref-bibr" href="#ref-17">Torres-Ruiz et al., 2014; <a id="xref-ref-18-1" class="xref-bibr" href="#ref-18">Trifil et al., 2014). The discovery of these artifacts has raised questions regarding the reliability of some previously published plant hydraulics data, in particular data relating to the refilling of embolized xylem conduits while the xylem is under tension. In this context, <a id="xref-ref-20-1" class="xref-bibr" href="#ref-20">Wheeler et al. (2013) reported that sampling plant organs by cutting while the xylem is under tension can induce artificial increases in the degree of embolism at the moment of sample excision, even when cuts are made under water. The methodology applied by <a id="xref-ref-20-2" class="xref-bibr" href="#ref-20">Wheeler et al. (2013), however, did not allow the visualization of embolized or functional vessels, and native embolism levels could not be determined in intact plants before any cutting was done.
DOI:10.1093/treephys/tpv049URLPMID:26116926 [本文引用: 1]
Recent studies have reported correlations between stem sapwood capacitance (Cwood) and xylem vulnerability to embolism, but it is unclear how Cwood relates to the eventual ability of plants to reverse embolism. We investigated possible functional links between embolism reversal efficiency, Cwood, wood density (WD), vulnerability to xylem embolism and hydraulic safety margins in nine woody species native to dry sclerophyllous forests with different degrees of iso versus anisohydry. Substantial inter-specific differences in terms of seasonal/diurnal changes of xylem and leaf water potential, maximum diurnal values of transpiration rate and xylem vulnerability to embolism formation were recorded. Significant diurnal changes in percentage loss of hydraulic conductivity (PLC) were recorded for five species. Significant correlations were recorded between diurnal PLC changes and P50 and P88 values (i.e., xylem pressure inducing 50 and 88% PLC, respectively) as well as between diurnal PLC changes and safety margins referenced to P50 and P88. WD was linearly correlated with minimum diurnal leaf water potential, diurnal PLC changes and wood capacitance across all species. In contrast, significant relationships between P50, safety margin values referenced to P50 and WD were recorded only for the isohydric species. Functional links between diurnal changes in PLC, hydraulic strategies and WD and Cwood are discussed.
DOI:10.1039/df9480300146URL [本文引用: 1]
DOI:10.1111/jipb.12534URL [本文引用: 1]
Herein we review the current state-of-the-art of plant hydraulics in the context of plant physiology,ecology, and evolution, focusing on current and future research opportunities. We explain the physics of water transport in plants and the limits of this transport system,highlighting the relationships between xylem structure and function. We describe the great variety of techniques existing for evaluating xylem resistance to cavitation. We address several methodological issues and their connection with current debates on conduit refilling and exponentially shaped vulnerability curves. We analyze the trade-offs existing between water transport safety and efficiency. We also stress how little information is available on molecular biology of cavitation and the potential role of aquaporins in conduit refilling. Finally,we draw attention to how plant hydraulic traits can be used for modeling stomatal responses to environmental variables and climate change, including drought mortality.
[本文引用: 3]
[本文引用: 1]
A multi-species synthesis of physiological mechanisms in drought-induced tree mortality
1
2017
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests
1
2010
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
Measurement of vulnerability to water stress-induced cavitation in grapevine: A comparison of four techniques applied to long-vesseled species
2
2010
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
... 刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (
The spatial pattern of air seeding thresholds in mature sugar maple trees
1
2005
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
A technique for measuring xylem hydraulic conductance under high negative pressures
2
2002
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
... 二是与不同方法中的具体操作有关.用自然干燥法建立一个栓塞脆弱性曲线需要一个星期或者更长, 根据公式(2), PLC的确定需要冲洗枝条, 将冲洗后的导水率作为Kmax.但是本实验中自然干燥法所测的冲洗后的导水率即Kmax存在差异, 并且所测的Kmax值随着时间的推移, 出现下降的趋势.这可能是由于在自然干燥脱水过程中, 刺槐一部分栓塞化的木质部导管无法通过冲洗而得到恢复(
Methods for measuring plant vulnerability to cavitation: A critical review
1
2013
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
Evaluation of a new centrifuge technique for rapid generation of xylem vulnerability curves
3
2005
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
... 刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (
... 二是与不同方法中的具体操作有关.用自然干燥法建立一个栓塞脆弱性曲线需要一个星期或者更长, 根据公式(2), PLC的确定需要冲洗枝条, 将冲洗后的导水率作为Kmax.但是本实验中自然干燥法所测的冲洗后的导水率即Kmax存在差异, 并且所测的Kmax值随着时间的推移, 出现下降的趋势.这可能是由于在自然干燥脱水过程中, 刺槐一部分栓塞化的木质部导管无法通过冲洗而得到恢复(
Does sample length influence the shape of xylem embolism vulnerability curves? A test with the Cavitron spinning technique
2
2010
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
... 刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (
Air method measurements of apple vessel length distributions with improved apparatus and theory
2
2003
... 选取来自不同个体树的6根枝条, 通过空气注入法测量木质部导管长度(
... )(
干旱导致树木死亡机制研究进展
1
2015
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
干旱导致树木死亡机制研究进展
1
2015
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
6个树种1年生枝木质部的水力特征及与栓塞修复能力的关系
3
2017
... Tension at 50% hydraulic conductivity loss (P50) in branches of Robinia pseudoacacia calculated based on different methods
| 栓塞脆弱性曲线的建立方法 Methods in establishing embolism vulnerability curves | P50 (MPa) | 样品数 Number of samples | 茎段长度 Stem length (cm) | 来源 Source |
|---|---|---|---|---|
| Sperry离心机法 Sperry centrifugation method | -0.93 ± 0.021a | 6 | 27.4 | |
| -0.92 ± 0.058a | 6 | 14.4 | ||
| 自然干燥法 Bench top dehydration method | -2.91b | 31 | ||
| Cochard Cavitron 离心机法 Cochard Cavitron centrifugation method | -0.38 ± 0.044c | 6 | 27.4 | |
| -0.46 ± 0.030d | 6 | 27.4 | al., 2017 | |
| -0.22 ± 0.026e | 6 | 27.4 |
... 本研究表明, 自然干燥法、Cochard Cavitron离心机法、Sperry离心机法建立刺槐的VC有较大差异.自然干燥建立的栓塞脆弱性曲线为“s”形, 而Cochard Cavitron离心机法和Sperry离心机法建立栓塞脆弱性曲线为“r”形,
... 二是与不同方法中的具体操作有关.用自然干燥法建立一个栓塞脆弱性曲线需要一个星期或者更长, 根据公式(2), PLC的确定需要冲洗枝条, 将冲洗后的导水率作为Kmax.但是本实验中自然干燥法所测的冲洗后的导水率即Kmax存在差异, 并且所测的Kmax值随着时间的推移, 出现下降的趋势.这可能是由于在自然干燥脱水过程中, 刺槐一部分栓塞化的木质部导管无法通过冲洗而得到恢复(
6个树种1年生枝木质部的水力特征及与栓塞修复能力的关系
3
2017
... Tension at 50% hydraulic conductivity loss (P50) in branches of Robinia pseudoacacia calculated based on different methods
| 栓塞脆弱性曲线的建立方法 Methods in establishing embolism vulnerability curves | P50 (MPa) | 样品数 Number of samples | 茎段长度 Stem length (cm) | 来源 Source |
|---|---|---|---|---|
| Sperry离心机法 Sperry centrifugation method | -0.93 ± 0.021a | 6 | 27.4 | |
| -0.92 ± 0.058a | 6 | 14.4 | ||
| 自然干燥法 Bench top dehydration method | -2.91b | 31 | ||
| Cochard Cavitron 离心机法 Cochard Cavitron centrifugation method | -0.38 ± 0.044c | 6 | 27.4 | |
| -0.46 ± 0.030d | 6 | 27.4 | al., 2017 | |
| -0.22 ± 0.026e | 6 | 27.4 |
... 本研究表明, 自然干燥法、Cochard Cavitron离心机法、Sperry离心机法建立刺槐的VC有较大差异.自然干燥建立的栓塞脆弱性曲线为“s”形, 而Cochard Cavitron离心机法和Sperry离心机法建立栓塞脆弱性曲线为“r”形,
... 二是与不同方法中的具体操作有关.用自然干燥法建立一个栓塞脆弱性曲线需要一个星期或者更长, 根据公式(2), PLC的确定需要冲洗枝条, 将冲洗后的导水率作为Kmax.但是本实验中自然干燥法所测的冲洗后的导水率即Kmax存在差异, 并且所测的Kmax值随着时间的推移, 出现下降的趋势.这可能是由于在自然干燥脱水过程中, 刺槐一部分栓塞化的木质部导管无法通过冲洗而得到恢复(
On the ascent of sap
1
1895
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
Cavitation and water storage capacity in bole xylem segments of mature and young Douglas-fir trees
1
2001
... 木质部栓塞脆弱性可以用脆弱性曲线(VC)来描述, VC是木质部导水率损失的百分数与相对应水势的关系曲线(
植物干旱胁迫下水分代谢、碳饥饿与死亡机理
1
2013
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
植物干旱胁迫下水分代谢、碳饥饿与死亡机理
1
2013
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
The standard centrifuge method accurately measures vulnerability curves of long- vesselled olive stems
2015
No evidence for an open vessel effect in centrifuge-based vulnerability curves of a long-vesselled liana (Vitisvinifera)
2
2012
... 刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (
... 和其他物种短茎段和长茎段的栓塞脆弱性曲线并没有差别.同时
Geographic and seasonal variation in chaparral vulnerability to cavitation
1
2014
... 刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (
6个耐旱树种木质部结构与栓塞脆弱性的关系
4
2016
... Tension at 50% hydraulic conductivity loss (P50) in branches of Robinia pseudoacacia calculated based on different methods
| 栓塞脆弱性曲线的建立方法 Methods in establishing embolism vulnerability curves | P50 (MPa) | 样品数 Number of samples | 茎段长度 Stem length (cm) | 来源 Source |
|---|---|---|---|---|
| Sperry离心机法 Sperry centrifugation method | -0.93 ± 0.021a | 6 | 27.4 | |
| -0.92 ± 0.058a | 6 | 14.4 | ||
| 自然干燥法 Bench top dehydration method | -2.91b | 31 | ||
| Cochard Cavitron 离心机法 Cochard Cavitron centrifugation method | -0.38 ± 0.044c | 6 | 27.4 | |
| -0.46 ± 0.030d | 6 | 27.4 | al., 2017 | |
| -0.22 ± 0.026e | 6 | 27.4 |
... 刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (
... ;
... 本研究表明, 自然干燥法、Cochard Cavitron离心机法、Sperry离心机法建立刺槐的VC有较大差异.自然干燥建立的栓塞脆弱性曲线为“s”形, 而Cochard Cavitron离心机法和Sperry离心机法建立栓塞脆弱性曲线为“r”形,
6个耐旱树种木质部结构与栓塞脆弱性的关系
4
2016
... Tension at 50% hydraulic conductivity loss (P50) in branches of Robinia pseudoacacia calculated based on different methods
| 栓塞脆弱性曲线的建立方法 Methods in establishing embolism vulnerability curves | P50 (MPa) | 样品数 Number of samples | 茎段长度 Stem length (cm) | 来源 Source |
|---|---|---|---|---|
| Sperry离心机法 Sperry centrifugation method | -0.93 ± 0.021a | 6 | 27.4 | |
| -0.92 ± 0.058a | 6 | 14.4 | ||
| 自然干燥法 Bench top dehydration method | -2.91b | 31 | ||
| Cochard Cavitron 离心机法 Cochard Cavitron centrifugation method | -0.38 ± 0.044c | 6 | 27.4 | |
| -0.46 ± 0.030d | 6 | 27.4 | al., 2017 | |
| -0.22 ± 0.026e | 6 | 27.4 |
... 刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (
... ;
... 本研究表明, 自然干燥法、Cochard Cavitron离心机法、Sperry离心机法建立刺槐的VC有较大差异.自然干燥建立的栓塞脆弱性曲线为“s”形, 而Cochard Cavitron离心机法和Sperry离心机法建立栓塞脆弱性曲线为“r”形,
本木植物木质部栓塞脆弱性研究新进展
1
2015
... 二是与不同方法中的具体操作有关.用自然干燥法建立一个栓塞脆弱性曲线需要一个星期或者更长, 根据公式(2), PLC的确定需要冲洗枝条, 将冲洗后的导水率作为Kmax.但是本实验中自然干燥法所测的冲洗后的导水率即Kmax存在差异, 并且所测的Kmax值随着时间的推移, 出现下降的趋势.这可能是由于在自然干燥脱水过程中, 刺槐一部分栓塞化的木质部导管无法通过冲洗而得到恢复(
本木植物木质部栓塞脆弱性研究新进展
1
2015
... 二是与不同方法中的具体操作有关.用自然干燥法建立一个栓塞脆弱性曲线需要一个星期或者更长, 根据公式(2), PLC的确定需要冲洗枝条, 将冲洗后的导水率作为Kmax.但是本实验中自然干燥法所测的冲洗后的导水率即Kmax存在差异, 并且所测的Kmax值随着时间的推移, 出现下降的趋势.这可能是由于在自然干燥脱水过程中, 刺槐一部分栓塞化的木质部导管无法通过冲洗而得到恢复(
Adaptive variation in the vulnerability of woody plants to xylem cavitation
1
2004
... 木质部栓塞脆弱性可以用脆弱性曲线(VC)来描述, VC是木质部导水率损失的百分数与相对应水势的关系曲线(
Vulnerability of xylem vessels to cavitation in sugar maple. Scaling from individual vessels to whole branches
1
2003
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
Cavitation and its discontents: Opportunities for resolving current controversies
1
2014
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
Vulnerability curves by centrifugation: Is there an open vessel artifact, and are “r” shaped curves necessarily invalid?
3
2012
... 刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (
... ).例如,
... 本研究表明, 自然干燥法、Cochard Cavitron离心机法、Sperry离心机法建立刺槐的VC有较大差异.自然干燥建立的栓塞脆弱性曲线为“s”形, 而Cochard Cavitron离心机法和Sperry离心机法建立栓塞脆弱性曲线为“r”形,
A method for measuring hydraulic conductivity and embolism in xylem
1
1988
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
Mechanism of water stress-induced xylem embolism
2
1988
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
... 木质部栓塞脆弱性可以用脆弱性曲线(VC)来描述, VC是木质部导水率损失的百分数与相对应水势的关系曲线(
Loss of hydraulic conductivity due to water stress in intact juveniles of Quercus rubra and Populus deltoides
1
1992
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
Direct micro-CT observation confirms the induction of embolism upon xylem cutting under tension
1
2015
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
Diurnal changes in embolism rate in nine dry forest trees: Relationships with species-specific xylem vulnerability, hydraulic strategy and wood traits
1
2015
... 二是与不同方法中的具体操作有关.用自然干燥法建立一个栓塞脆弱性曲线需要一个星期或者更长, 根据公式(2), PLC的确定需要冲洗枝条, 将冲洗后的导水率作为Kmax.但是本实验中自然干燥法所测的冲洗后的导水率即Kmax存在差异, 并且所测的Kmax值随着时间的推移, 出现下降的趋势.这可能是由于在自然干燥脱水过程中, 刺槐一部分栓塞化的木质部导管无法通过冲洗而得到恢复(
Water transport in plants as a catenary process
1
1948
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
Plant xylem hydraulics: What we understand, current research, and future challenges
1
2017
... 近年来, 随着全球变暖的加剧, 区域性林木死亡率不断升高(
Water relations of Robinia pseudoacacia L.: Do vessels cavitate and refill diurnally or are R-shaped curves invalid in Robinia?
3
2014
... 目前, 建立VC的方法有很多, 常用的方法有自然干燥法(
... 刺槐是一种长导管物种, 本研究得到刺槐的平均导管长度为20.02 cm, 这与最近的一些文献报道的刺槐的平均导管长度为20.55 cm (
... 二是与不同方法中的具体操作有关.用自然干燥法建立一个栓塞脆弱性曲线需要一个星期或者更长, 根据公式(2), PLC的确定需要冲洗枝条, 将冲洗后的导水率作为Kmax.但是本实验中自然干燥法所测的冲洗后的导水率即Kmax存在差异, 并且所测的Kmax值随着时间的推移, 出现下降的趋势.这可能是由于在自然干燥脱水过程中, 刺槐一部分栓塞化的木质部导管无法通过冲洗而得到恢复(
Xylem Structure and the Ascent of Sap
1
1983
... 一是自然干燥法、Sperry离心机法和Cochard Cavitron离心机法诱导栓塞的方式不同.自然干燥法是最简单和最直接诱导植物栓塞和空穴化的方法, 即通过离体枝条内水分的自然散失来诱导栓塞, 在不同栓塞程度下测定该枝条的导水率损失, 同时通过测量叶片或小枝的水势从而获取枝条木质部的水势.因此自然干燥法的栓塞形成过程比较漫长, 枝条各个区段处于相近的水势, 相互连通的导管之间保持水力学平衡.而离心机法建立VC时, 枝条两端位置的张力为0, 而中轴处张力最大, 即整个枝条的水势分布不均匀, 这一方面是枝条的中部更容易栓塞, 另一方面尤其在较高转速时, 气泡可能会因为水力学不平衡更容易进入导管.“气种”假说是一种植物发生栓塞的机理, 即木质部导管内栓塞的发生是外界空气或已经栓塞的导管内的空气泡, 通过导管间纹孔膜上的微孔扩散至相邻导管内, 水分蒸发导致连续水柱断裂, 随之空气泡的体积不断增加, 促使管道内的压力瞬时升高, 纹孔膜上的微孔被阻塞, 最终空气泡的体积充满整个导管, 使木质部输水能力下降(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}