 ,*北京大学城市与环境学院, 北京大学地表过程分析与模拟教育部重点实验室, 北京 100871
,*北京大学城市与环境学院, 北京大学地表过程分析与模拟教育部重点实验室, 北京 100871Ecosystem carbon stock and within-system distribution in successional Fagus lucida forests in Mt. Yueliang, Guizhou, China
ZHOU Xu-Li, CAI Qiong, XIONG Xin-Yu, FANG Wen-Jing, ZHU Jian-Xiao, ZHU Jiang-Ling, FANG Jing-Yun, JI Cheng-Jun,*College of Urban and Environmental Sciences, and Key Laboratory for Earth Surface Processes of the Ministry of Education, Peking University, Beijing 100871, China通讯作者:
编委: 李胜功
责任编辑: 李敏
| 基金资助: |
Online:2018-07-20
| Supported by: | SupportedbytheNationalKeyResearchandDevelopmentProgramofChina( |

摘要
关键词:
Abstract
Keywords:
PDF (1271KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
周序力, 蔡琼, 熊心雨, 方文静, 朱剑霄, 朱江玲, 方精云, 吉成均. 贵州月亮山不同演替阶段亮叶水青冈林碳储量及其分配格局. 植物生态学报, 2018, 42(7): 703-712 doi:10.17521/cjpe.2018.0064
ZHOU Xu-Li, CAI Qiong, XIONG Xin-Yu, FANG Wen-Jing, ZHU Jian-Xiao, ZHU Jiang-Ling, FANG Jing-Yun, JI Cheng-Jun.
全球森林碳储量达到861 Pg (1 Pg = 1015 g), 并以每年2.4 Pg的速度增长, 在缓解大气CO2浓度升高方面具有重要作用(Pan et al., 2011)。未受人类活动干扰的森林生态系统碳库包括植被、木质残体、凋落物和土壤碳库(IPCC, 2013)。这些碳库受到诸多环境因子的影响, 如森林类型(张全智和王传宽, 2010)、林龄(Pregitzer & Euskirchen, 2004)、温度与降水(Aplet & Vitousek, 1994)等。其中, 林龄对各组分碳储量的影响至关重要(Gower et al., 1997; Pregitzer & Euskirchen, 2004; Zhu et al., 2017a)。
已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化。研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(Zhu et al., 2017b), 广西7-32年格木(Erythrophleum fordii)林(明安刚等, 2014), 韩国庆尚北道10-71年赤松(Pinus densiflora)林(Noh et al., 2010)和加拿大安大略南部2-65年北美乔松(Pinus strobus)林(Peichl & Arain, 2006)等。也有全球尺度的研究, 如Pregitzer和Euskirchen (2004)整合分析了全球200年以下针叶林、温带与热带森林碳储量随林龄变化的情况。以往的研究表明, 在一定年龄范围内, 随林龄增加, 森林生态系统碳储量和植被碳储量通常呈增加趋势, 凋落物和木质残体也会随之变化(Hooker & Compton, 2003; Pregitzer & Euskirchen, 2004; Peichl & Arain, 2006; Li et al., 2011; Zhu et al., 2017b)。而对于土壤碳储量, 随林龄增加通常也表现为碳积累过程(Hooker & Compton, 2003; Pregitzer & Euskirchen, 2004; Zhu et al., 2017b), 然而林龄对土壤碳积累的作用可能受到土地利用历史、气候和植被类型等多种因素的影响, 故随演替进行, 土壤碳储量也可能不变或呈现先减少后增加的趋势(Peichl & Arain, 2006; Nave et al., 2010; Li et al., 2011)。在碳储量的分配方面, 随演替进行, 通常植被碳储量贡献增加, 而土壤碳储量贡献减小(Pregitzer & Euskirchen, 2004; Peichl & Arain, 2006; Zhu et al., 2017a)。但这些研究多针对人工林或中高纬度地区的针叶林, 有关亚热带天然阔叶林生态系统碳储量及其分配格局在不同演替阶段的变化目前仍鲜有报道(宫超等, 2011)。
亚热带森林在陆地生态系统碳循环中起着重要作用。有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (Yu et al., 2014)。水青冈属(Fagus)植物是中国暖温带与亚热带地区广泛分布的落叶阔叶乔木, 也是欧洲、北美和日本落叶阔叶林的代表性树种, 水青冈林是北半球重要的森林类型(Wang & Fujiwara, 2003; Fang & Lechowicz, 2006)。对于水青冈林的碳储量和生产力, 国外已有大量研究(Kakubari, 1991; Mund, 2004), 而在我国, 水青冈林碳储量的相关研究几乎未见报道。在贵州月亮山, 集中分布着大片处于不同演替阶段的亮叶水青冈(Fagus lucida)天然林(杨业勤, 1994), 这在我国水青冈其他分布区较为少见, 为研究不同演替阶段水青冈林碳库结构提供了理想的对象, 同时对了解我国亚热带森林结构与功能随林龄的变化也具有一定的参考意义。因此, 我们选择月亮山3个不同演替阶段(33年、82年和208年)的9个亮叶水青冈林为调查对象, 对样方内植被、土壤、木质残体和凋落物4个组分的碳储量进行了测定与估算, 旨在探讨不同演替阶段亮叶水青冈林生态系统的碳储量及其分配格局的变化。
1 材料和方法
1.1 样地信息
研究地点位于贵州省黔东南州榕江县月亮山省级自然保护区(108.22°-108.32° E, 25.57°-25.65° N)。月亮山地处贵州省东南部, 位于华南褶皱带, 地质构造属新华夏体系, 基岩多为浅变质黏土岩类, 地貌类型为强切割侵蚀构造中低山。该区域属于亚热带山地季风气候, 年平均气温12.1-15.7 ℃, 年降水量1 030-1 688 mm。地带性植被为常绿阔叶林, 1 100 m以上主要为常绿落叶阔叶混交林, 土壤多为山地黄棕壤。月亮山低山地区受砍伐和放牧等人为活动干扰较大, 而中山常绿阔叶混交林保存相对完好。亮叶水青冈林是该区域常绿阔叶混交林中主要的森林类型(杨业勤, 1994; 吴开岑等, 2013; 张亚冰等, 2016)。2017年6月, 我们在海拔1 400-1 474 m范围内, 选取了3个不同演替阶段(平均林龄分别为33年、82年和208年)的亮叶水青冈林样地, 每个样地设置3个重复样方(20 m × 30 m)。9个亮叶水青冈样方中, 3个33年亮叶水青冈样方为天然次生林, 20世纪80年代砍伐后天然更新, 其余样方均为原始林。该区域土壤深度约为30 cm。在本研究中, 33年林乔木层主要树种有亮叶水青冈、贵州桤叶树(Clethra kaipoensis)和中华枫(Acer sinense)等, 88年林主要树种有亮叶水青冈、多脉青冈(Cyclobalanopsis multinervis)和尖叶四照花(Dendrobenthamia angustata)等, 208年林主要树种有亮叶水青冈、多脉青冈和杜鹃(Rhododendron sp.)等。样方基本信息见表1。
Table 1
表1
表1贵州月亮山水青冈林9个样方样地信息表
Table 1
| 样地编号 Stand No. | 33年林 33 a forest | 82年林 82 a forest | 208年林 208 a forest | ||||||
|---|---|---|---|---|---|---|---|---|---|
| YLS 1 | YLS 2 | YLS 3 | YLS 4 | YLS 5 | YLS 6 | YLS 7 | YLS 8 | YLS 9 | |
| 海拔 Altitude (m) | 1 422 | 1 400 | 1 405 | 1 457 | 1 451 | 1 439 | 1 474 | 1 471 | 1 469 |
| 森林起源 Forest origin | 次生林 Secondary forest | 次生林 Secondary forest | 次生林 Secondary forest | 原始林 Primeval forest | 原始林 Primeval forest | 原始林 Primeval forest | 原始林 Primeval forest | 原始林 Primeval forest | 原始林 Primeval forest |
| 林龄 Stand age (a) | 30 | 36 | 34 | 71 | 84 | 92 | 203 | 215 | 207 |
| 坡向 Aspect | 南偏东80度 80° SE | 南偏东25度 25° SE | 南偏东25度 25° SE | 北偏西80度 80° NW | 正西 W | 北偏西60度 60° NW | 南偏东55度 55° SE | 正北 N | 南偏西70度 70° SW |
| 坡度 Slope (°) | 35 | 45 | 47 | 35 | 16 | 37 | 32 | 36 | 32 |
| 平均胸径 Mean DBH (cm) | 8.9 | 9.2 | 8.9 | 17.0 | 17.4 | 14.6 | 16.1 | 19.2 | 18.9 |
| 最大胸径 DBHmax (cm) | 25.9 | 24.5 | 26.1 | 42.7 | 75.8 | 44.6 | 90.7 | 74.5 | 71.6 |
| 平均树高 Mean Height (m) | 7.7 | 7.7 | 7.5 | 10.6 | 13.7 | 9.9 | 8.1 | 11.7 | 9.9 |
| 最大树高 Heightmax (m) | 12 | 12 | 13 | 16 | 25 | 21 | 17 | 23 | 24 |
| 总胸高断面积 TBA (m2·hm-2) | 24.7 | 25.1 | 27.5 | 41.4 | 65.0 | 52.5 | 79.0 | 49.3 | 63.2 |
| 密度 Stand density (No.· hm-2) | 3 200 | 3 167 | 3 483 | 1 500 | 1 850 | 2 300 | 1 800 | 1 000 | 1 400 |
| 乔木种数 Number of trees | 21 | 19 | 23 | 17 | 19 | 24 | 18 | 21 | 19 |
| 灌木种数 Number of shrubs | 48 | 52 | 44 | 49 | 70 | 70 | 47 | 52 | 55 |
| 草本种数 Number of herbs | 27 | 20 | 24 | 22 | 23 | 25 | 15 | 22 | 24 |
新窗口打开|下载CSV
1.2 野外调查及数据获取
记录每个样方的经纬度、海拔、坡向、坡度等基本信息, 对样方内植被(包括乔木层、灌木层和草本层)、木质残体、凋落物和土壤的碳密度(Mg·hm-2)分别进行调查和估算。1.2.1 林龄测定
在样方内选取10株胸径最大的亮叶水青冈, 在1.3米处利用生长锥从两个垂直的方向进行树芯采集。样品带回实验室, 粘贴打磨, 用交叉定年法确定树木的年龄, 取两个方向的平均值作为每株树的年龄。将每个样方内亮叶水青冈年龄的最大值作为该样方水青冈林的林龄。
1.2.2 植被碳储量
测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名。利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量。由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(邱学忠等, 1984; 邓仕坚等, 2000; 姚迎九等, 2003; 王向雨等, 2007; 明安刚等, 2012)。以0.5作为碳转换系数将树木生物量换算为生物量碳(Myneni et al., 2001; Zhu et al., 2015)对样地内不同树种的生物量进行估算, 具体见表2。将样方内所有乔木生物量碳加和后, 根据样方面积计算单位面积的碳储量(碳密度, Mg·hm-2)。
Table 2
表2
表2乔木层树种生物量方程
Table 2
| 物种 Species | 地上生物量 AGB (kg) | 地下生物量 BGB (kg) | 本文主要对应树种 Species in this study | 参考文献 References |
|---|---|---|---|---|
| 锥属 Castanopsis | AGB = 0.0177(D2H)1.0168 + 0.0364(D2H)0.6530 + 0.1533(D2H)0.2948 | BGB = 0.00911(D2H)0.933951 | 锥属 Castanopsis | Qiu et al., 1984 |
| 水青冈属 Fagus | AGB = 0.0125(D2H)1.05 + 0.000933(D 2H)1.23 + 0.000294(D 2H)1.20 | BGB = 0.00322(D2H)1.13 | 亮叶水青冈 Fagus lucida | Wang et al., 2007 |
| 樟科 Lauraceae | AGB = 0.055603(D2H)0.850193 + 0.014757(D2H)0.808395 + 0.006652(D2H)1.051841 + 0.059871(D2H)0.574327 | BGB = 0.184736(D2H)0.616421 | 木姜子属 Litsea | Yao et al., 2003 |
| 枫香 Liquidambar formosana | AGB = 0.174(D2H)0.7661 + 3 × 10-8(D2H)2 + 0.001(D2H) + 9.7883 + 0.0002(D2H)1.2696 + 0.0002D3.2304 + 3 × 10-7(D2H)1.5626 | BGB = 0.0094(D2H)0.9538 | 枫香树 Liquidambar formosana | Ming et al., 2012 |
| 柯属 Lithocarpus | AGB = 0.0347(D2H)0.9470 + 0.0084(D2H)0.9112 + 0.0072(D2H)0.6893 | BGB = 0.01534(D2H)0.95121 | 柯属 Lithocarpus | Qiu et al., 1984 |
| 木兰科 Magnoliaceae | AGB = 0.502921(D2H)0.56821 + 0.007183(D2H)0.92191 + 0.02252(D2H)0.62601 | BGB = 0.0364(D2H)0.79111 | 木兰属 Magnolia | Qiu et al., 1984 |
| 落叶阔叶树 Deciduous broad-leaved trees | AGB = 0.0650(D2H)0.84 + 1.59(D2H)0.38 + 0.218(D2H)0.34 | BGB = 0.291(D2H)0.55 | 鹅耳枥属 Carpinus, 槭属 Acer | Wang et al., 2007 |
| 常绿阔叶树 Evergreen broad-leaved trees | AGB = 0.17686(D2H)0.75995 + 0.11499(D2H)0.69997 + 0.107513(D2H)0.53231 | BGB = 0.095827(D2H)0.7165 | 青冈属 Cyclobalanopsis, 冬青属 Ilex | Deng et al., 2000 |
| 落叶小乔木 Small deciduous trees | AGB = 0.0434(D2H)0.91 + 0.000902(D2H)1.31 + 0.000790(D2H)1.05 | BGB = 0.000781(D2H)1.05 | 尖叶四照花 Dendrobenthamia angustata, 贵州桤叶树 Clethra kaipoensis | Wang et al., 2007 |
| 常绿小乔木 Small evergreen trees | AGB = 0.190(D2H)0.663 + 0.123(D2H)1.023 + 0.00728(D2H)0.548 | BGB = 0.0557(D2H)0.622 | 川桂 Cinnamomum wilsonii, 杜鹃属 Rhododendron | Wang et al., 2007 |
新窗口打开|下载CSV
在样方内随机选取3个2 m × 2 m的小样方, 连根收割小样方内所有灌木和草本, 测定鲜质量, 并取样带回实验室, 在65 ℃烘干至恒质量后, 测定含水率, 分别计算其地上生物量和地下生物量。灌木和草本层样品经研磨后用元素分析仪(2400IICHNS/O, Perkin-Elmer, Boston, USA)分别测定地上与地下部分的碳含量, 结合生物量计算灌木层和草本层的碳密度。
1.2.3 木质残体和凋落物碳储量
将木质残体区分为细木质残体(大头直径2-10 cm; FWD)和粗木质残体(大头直径≥10 cm; CWD) (Harmon et al., 1986)。测量样方内所有枯立木的胸径和高度; 对倒木记录其两头和中间的直径、长度, 以及腐朽程度(1-4, Zhu et al., 2017b)。倒木和枯立木体积按以下公式计算:
Volume = πd2L/4 (1)
其中d为枯立木胸径或倒木两端和中心的平均直径, L为倒木的长度(m)或枯立木的高(m)。采集不同腐朽程度的枯倒木样品(每株枯倒木取3段木段), 带回实验室烘干至恒质量后测量干质量, 研磨过筛后测定碳含量。计算采样枯倒木体积与干质量的关系, 由此推算样地内枯倒木的干质量与碳密度。
对样方内所有FWD进行收集并称质量。在样方内随机设置3个1 m × 1 m小样方, 收集小样方内所有凋落物(包括落叶、树皮、果实、直径< 2 cm的枯枝以及地表其他植物残体)并称质量。每个600 m2样方内, FWD与凋落物各取3份样品, 带回实验室烘干(65 ℃)至恒质量后测定含水率, 并换算其干质量, 研磨过0.15 mm筛后测定碳含量(Zhu et al., 2017a, 2017b)。
1.2.4 土壤碳储量
在样方内随机挖取3个土壤剖面, 以10 cm为间隔分层, 取到实际土壤深度(约30 cm, 即0-10、10-20和20-30 cm分3层取样)。每层取两份土壤样品。其中一份使用环刀(100 cm3, 直径50.5 mm, 高50 mm)取样, 采用105 ℃烘干48 h后测定各层土壤容重; 另一份取约300 g鲜土, 在室温(约25 ℃, 两周)自然风干, 去除杂质后研磨过筛(0.15 mm), 用元素分析仪测定土壤碳含量。
1.3 数据分析
采用单因素方差分析和多重比较(最小显著差异法)探讨不同林龄亮叶水青冈林生态系统各组分碳储量的差异。2 结果
2.1 不同林龄水青冈林生态系统各组分碳储量
2.1.1 植被碳储量不同演替阶段亮叶水青冈林的植被碳密度分别为(60.2 ± 7.5) Mg·hm-2 (33年林)、(211.1 ± 87.5) Mg·hm-2 (82年林)和(396.6 ± 150.8) Mg·hm-2 (208年林), 其中乔木层占据植被碳密度最主要的部分((58.0 ± 7.0) - (394.0 ± 149.6) Mg·hm-2, 96.3%- 99.3%), 其次为灌木层((2.0 ± 0.9) - (2.5 ± 1.4) Mg·hm-2, 0.6%-3.5%), 草本层所占比例最小((0.01 ± 0.005) - (0.06 ± 0.02) Mg·hm-2, <0.1%)(图1A)。从33年林到208年林, 亮叶水青冈林生物量碳密度显著增加(F = 19.3, p = 0.003), 其中乔木层碳储量随林龄的增加而显著增加(F =19.5, p = 0.003), 而灌木层和草本层碳储量无显著变化(图1A)。在生物量碳储量的分配上, 地上部分碳储量占总储量的主要部分((47.8 ± 6.0) - (301.9 ± 109.9) Mg·hm-2, 76.1%- 79.4%))。随着林龄的增加, 植被地上(F = 20.6, p = 0.003)与地下(F = 15.9, p = 0.005)部分碳储量都显著增加(图1B)。
图1
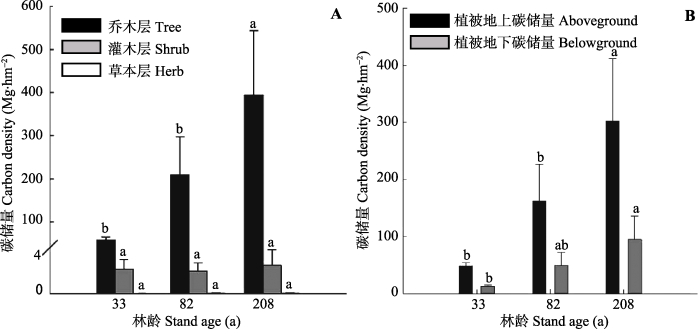 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同演替阶段月亮山亮叶水青冈林不同植被组成的碳储量(平均值+标准误差)。A, 不同生活型碳储量。B, 植被地上碳储量和地下碳储量。
Fig. 1Changes in vegetation carbon density of the nine successional Fagus lucida forests in Mt. Yueliang (mean + SE). A, Carbon density in different life forms (trees, shrubs and herbs). B, Above- and below-ground carbon density.
2.1.2 凋落物和木质残体碳储量
不同演替阶段亮叶水青冈林的植物残体(凋落物+木质残体)碳储量为(1.4 ± 0.2) - (4.8 ± 2.5) Mg·hm-2, 其中木质残体碳储量为(0.5 ± 0.1) - (3.2 ± 2.6) Mg·hm-2, 凋落物碳储量为(0.9 ± 0.3) - (1.6 ± 0.2) Mg·hm-2。随着演替的进行, 植物残体的碳储量显著增加(F = 8.7, p = 0.021; 图2A), 其中凋落物(F = 8.0, p < 0.05)和木质残体(F = 5.74, p = 0.05)的碳储量在208年亮叶水青冈林中最高(分别为(1.6 ± 0.2) Mg·hm-2和(3.2 ± 2.6) Mg·hm-2), 而在33年林中最低(分别为(0.9 ± 0.3) Mg·hm-2和(0.5 ± 0.1) Mg·hm-2) (图2A)。木质残体中, 粗木质残体(F = 5.7, p < 0.05)碳储量在208年林中最高((2.87 ± 2.61) Mg·hm-2), 在33年林中最低((0.07 ± 0.04) Mg·hm-2), 而细木质残体碳储量无明显变化(图2A)。随着演替的进行, 植物残体中凋落物所占比例逐渐降低; 而木质残体比例逐渐增加, 且主要由粗木质残体贡献(图2B)。
图2
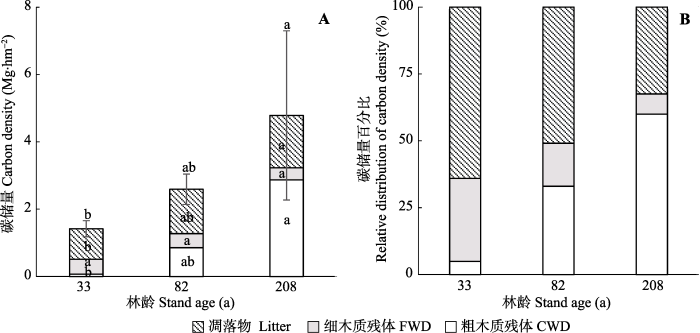 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同演替阶段亮叶水青冈林凋落物和木质残体的碳储量。A, 植物残体碳储量绝对值(平均值+标准误差)。B, 植物残体碳储量相对值。
Fig. 2Absolute (mean + SE) (A) and relative distribution of carbon density (B) of plant debris in the nine successional Fagus lucida forests in Mt. Yueliang. CWD, coarse woody debris; FWD, fine woody debris.
2.1.3 土壤碳储量
不同演替阶段亮叶水青冈林的土壤碳储量分别为(125.3 ± 39.0) Mg·hm-2 (33年林)、(52.1 ± 6.6) Mg·hm-2 (82年林)和(113.8 ± 32.5) Mg·hm-2 (208年林)(图3A)。在土壤碳储量的垂直分配上, 表层(0-10 cm)土壤碳储量所占比重最大(43.5-44.3%; 图3B); 在33年和82年林中, 表层土壤碳储量皆显著大于下层土壤(p < 0.05)。在不同演替阶段, 土壤总碳储量及10-20 cm和20-30 cm土壤碳储量皆无显著变化; 而表层土壤碳储量在33年林中最高, 82年林中最低(图3A)。随演替进行, 表层土壤碳储量比重基本保持不变(林龄从低到高依次为44.3%、44.0%和43.5%), 10-20 cm土壤碳储量所占比重略有降低(依次从34.4%降至27.8%、23.9%), 而20-30 cm土壤碳储量比重略有增加(依次从21.3%增加至28.2%、32.6%)? (图3B)。
图3
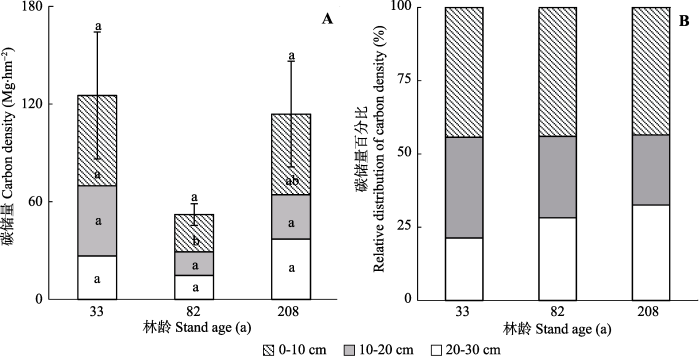 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同演替阶段亮叶水青冈林土壤碳储量。A, 土壤碳储量绝对值(平均值+标准误差)。B, 土壤碳储量相对值。
Fig. 3Absolute (mean + SE) (A) and relative distribution of soil carbon density (B) by soil depth in the nine successional Fagus lucida forests of Mt. Yueliang.
2.2 生态系统碳储量分配格局
不同演替阶段亮叶水青冈林生态系统碳储量总量分别为(186.9 ± 46.0) Mg·hm-2 (33年林)、(211.1 ± 87.5) Mg·hm-2 (82年林)和(515.1 ± 176.4) Mg·hm-2 (208年林), 主要由植被碳储量(占32.2%-79.4%)和土壤碳储量(占19.6-67.0%)贡献(图4)。凋落物与木质残体碳储量的贡献几乎可以忽略不计(<1%)(图4)。生态系统碳储量总量在208年亮叶水青冈林中最高, 而在33年林中最低(F = 10.7, p < 0.05; 图4A); 其分配格局也随林龄的变化而改变。33年林中, 生态系统碳储量主要由土壤贡献(占67.0%), 而82年和208年亮叶水青冈林中生态系统碳储量主要由植被贡献(分别占79.4%和78.8%)(图4B)。凋落物和木质残体碳储量的比例变化较小(图4B)。图4
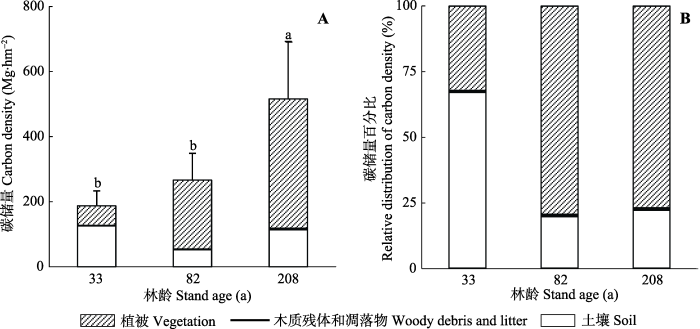 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同演替阶段亮叶水青冈林生态系统碳储量。A, 生态系统碳储量绝对值(平均值+标准误差)。B, 生态系统碳储量相对值。
Fig. 4Absolute (mean + SE) (A) and relative distribution of ecosystem carbon density (B) in the nine successional Fagus lucida forests of Mt. Yueliang.
3 讨论
3.1 植被碳储量
林龄是影响森林生态系统碳储量的重要因素(Chapin et al., 2002; Pregitzer & Euskirchen, 2004)。植被碳储量随演替进行而增加的现象在针叶林、温带和热带地区的天然林与人工林中普遍存在(Pregitzer & Euskirchen, 2004; 唐旭利和周国逸, 2005; Noh et al., 2010; Li et al., 2011; 付威波等, 2014; Zhu et al., 2017b)。本研究中, 月亮山亮叶水青冈林植被碳储量随林龄增长亦显著增加; 其中乔木层是亮叶水青冈林植被碳储量的主要贡献者, 且其比例随林龄增长而增加, 这与以往研究的结果(Peichl & Arain, 2006; Bradford & Kastendick, 2010)也相似。而不同演替阶段亮叶水青冈林的灌木层与草本层碳储量皆无显著差异。有关林下生物量随林龄增长的变化趋势目前并无一致的结论。一些研究发现, 在幼龄林阶段, 林下生物量随林龄增长而降低; 而此后林龄进一步增长时(约40年以后), 林下层生物量基本无明显变化趋势, 如美国明尼苏达州北部7-160年Pinus resinosa林和6-133年杨树(Populus tremuloides和P. grandidentata)林(Bradford & Kastendick, 2010)。也有研究发现, 一定林龄范围内, 灌木层生物量随林龄增加而增长, 而草本层相反, 如2-65年北美乔松林(P. strobus)(Peichl & Arain, 2006), 韩国8-51年红松林(P. koraiensis)(Li et al., 2011)等。而在7-32年格木林中, 林下生物量沿林龄序列先增加后减少(明安刚等, 2014)。林下生物量与乔木层郁闭度密切相关, 通常随郁闭度增加而减小(Sigurdsson et al., 2005)。本研究中, 不同演替阶段亮叶水青冈林郁闭度接近, 故灌木层与草本层碳储量无显著差异。此外, 林下生物量还受到土壤养分状况、干扰和管理方式等多种因素的影响(Peichl & Arain, 2006)。
3.2 凋落物与木质残体碳储量
随林龄增长, 木质残体碳储量呈增加趋势, 这与我国森林整体(Zhu et al., 2017b)以及大兴安岭落叶松林(Zhu et al., 2017a)的研究结果一致。而以往的研究多报道木质残体碳储量随林龄增加呈U型变化(Spies et al., 1988; Sturtevant et al., 1997; Pregitzer & Euskirchen, 2004), 主要是因为调查的森林演替多起源于火烧等重大干扰, 因而幼龄林中残留有较多的木质残体(Spies et al., 1988; Pregitzer & Euskirchen, 2004; Zhu et al., 2017b)。有关凋落物随林龄的变化趋势, 不同研究的结果不尽相同。如全球200年以下温带森林(Pregitzer & Euskirchen, 2004)与北美7-160年Pinus resinosa林(Bradford & Kastendick, 2010)地表有机质层碳储量都随林龄增长呈增加趋势。研究者认为这可能是由于幼龄林中凋落物被收获或干扰较大从而加速了凋落物分解(Jandl et al., 2007; Bradford & Kastendick, 2010)。也有研究报道凋落物碳储量与林龄无关, 如北美6-133年杨树林(Bradford & Kastendick, 2010)和大兴安岭15-138年落叶松林(Zhu et al., 2017a)等, 其原因可能是凋落物的分解速率相对较快, 因而能较快地达到输入与输出之间的平衡(Peichl & Arain, 2006; Bradford & Kastendick, 2010)。此外, 凋落物碳储量随林龄的变化与森林类型也有关(Bradford & Kastendick, 2010)。植物残体碳储量表征植物残体产量与其分解量的动态平衡(Zhu et al., 2017b)。本研究中, 随演替的进行, 植被碳储量显著增加, 导致凋落物和木质残体的产量也随之增加; 而由于33年林为砍伐后次生, 木质残体与凋落物层受人为干扰和破坏较大, 增加了其流失和分解的速率(Bradford & Kastendick, 2010)。因而33年林中木质残体与凋落物的碳储量都低于82年和208年亮叶水青冈林。
3.3 土壤碳储量
在一定林龄范围内, 随林龄增加, 森林土壤通常表现为碳积累过程, 即土壤碳储量一般呈增加趋势(Hooker & Compton, 2003; Pregitzer & Euskirchen, 2004; Zhu et al., 2017b)。然而林龄对土壤碳积累的作用可能受到多种因素的影响, 其中最重要的影响因子包括土地利用历史、气候和植被类型(Paul et al., 2002; Peichl & Arain, 2006)。因而, 也有研究报道在农田或弃耕地上造林的森林土壤碳储量与林龄无显著关系(Peltoniemi et al., 2004), 或随林龄增长先降低后增加, 其中0-10 cm土壤碳储量约30年后才可以恢复到造林前的水平(Paul et al., 2002)。本研究中, 不同演替阶段亮叶水青冈林土壤总碳储量无显著差异; 而0-10 cm土壤碳储量在33年林最高, 82年林最低。这种变化格局可能与研究样地的土地利用历史有关(Paul et al., 2002; Pregitzer & Euskirchen, 2004; Peichl & Arain, 2006)。33年亮叶水青冈林为砍伐后次生, 此前该样地原生植被林龄可能较大, 有较多的碳积累, 故其土壤碳储量值较高。尽管砍伐等干扰通常会使土壤碳储量减少, 尤其是表层土, 但一般在一定时间后可以恢复(Paul et al., 2002; Nave et al., 2010)。所以本研究中, 林龄较小的次生林的土壤碳储量与林龄较大的原始林相当, 甚至表层土壤碳储量高于林龄较大的样地, 主要取决于33年林土壤中原有的碳储量较高。此外, 相比于深层的矿质土壤, 表层土壤更容易受到干扰等因素的影响, 故其变异也更大(Paul et al., 2002; Peichl & Arain, 2006; Nave et al., 2010)。因此, 在讨论森林土壤碳储量随林龄的变化格局时, 原有的土地利用方式是一个不可忽视的影响因子。
3.4 生态系统碳储量及其分配
本研究中, 不同演替阶段亮叶水青冈林生态系统碳储量从33年林的186.9 Mg·hm-2增加到208年林的515.1 Mg·hm-2。贵州梵净山44-185年水青冈(F. longipetiolata和F. lucida)林的生态系统碳储量在190.7-503.9 Mg·hm-2之间(本课题组未发表数据); 德国Thuringia中部30-171年欧洲水青冈(F. sylvatica)林的碳储量为126-413 Mg·hm-2 (Mund, 2004); 而全球不同林龄(≤200 year)温带森林的生态系统碳储量平均值为121-537 Mg·hm-2 (Pregitzer & Euskirchen, 2004), 本研究的结果与这些研究的结果较为接近。亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低。这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(明安刚等, 2014)、川中丘陵桤木(Alnus cremastogyne)-柏木(Cupressus funebris)混交林(吴鹏飞等, 2008)、大兴安岭兴安落叶松林(Zhu et al., 2017a)以及韩国红松林(Li et al., 2011)和北美乔松林(Peichl & Arain, 2006)等。亮叶水青冈林生态系统碳储量的分配比例与相近林龄范围的欧洲水青冈林和我国一些温带与亚热带森林的分配格局也较为接近。如德国30-171年欧洲水青冈林植被碳储量贡献为34.4%-75.5%, 土壤碳储量占比为23.6%- 62.0%, 凋落物碳储量比例不到4% (Mund, 2004)。
3.5 不确定性
本研究调查了贵州榕江县月亮山不同演替阶段的9个亮叶水青冈林生态系统的全组分碳储量及各组分的相对贡献。可能会给结果带来不确定性的因素主要有两方面: 一是由于保护区禁止采伐树木, 本研究在估算乔木生物量时利用的是文献中已有的生物量方程(Zhu et al., 2017a), 尽管尽可能采用相似树种和相近气候条件下的生物量方程, 但仍可能给结果带来一定的不确定性。其二, 对乔木层生物量碳储量, 本研究采用通用的0.5作为碳转换系数(Myneni et al., 2001; Zhu et al., 2015), 这可能与乔木实际的碳含量有一定的偏差(Li et al., 2011; Ma et al., 2018)。致谢 感谢北京大学城市与环境学院生态学系的肖剑、倪晓凤在野外调查中的帮助, 以及贵州榕江县林业局工作人员的协助。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.2307/2261393URL [本文引用: 1]
DOI:10.1139/X10-002URL [本文引用: 8]

Forest managers are seeking strategies to create stands that can adapt to new climatic conditions and simultane- ously help mitigate increases in atmospheric CO2. Adaptation strategi
[本文引用: 1]
DOI:10.1007/s11769-000-0054-1URL [本文引用: 1]
Based on the mean DBH measurements of sample trees,the bioproductivity of subtropical )). The results showed that the total biomass of the forest was 451.02 t m, and the component biomass of tree, shrub, liana and litter layer was 426.76,17.76,1.80 and 4.70t m, respectively. The net production and average production of tree layer were 34.46 and 13.32 t m r,respectively.
DOI:10.1007/s11769-000-0054-1URL [本文引用: 1]
Based on the mean DBH measurements of sample trees,the bioproductivity of subtropical )). The results showed that the total biomass of the forest was 451.02 t m, and the component biomass of tree, shrub, liana and litter layer was 426.76,17.76,1.80 and 4.70t m, respectively. The net production and average production of tree layer were 34.46 and 13.32 t m r,respectively.
DOI:10.1111/j.1365-2699.2006.01533.xURL [本文引用: 1]
Aim Beech ( Fagus L., Fagaceae) species are representative trees of temperate deciduous broadleaf forests in the Northern Hemisphere. We focus on the distributional limits of beech species, in particular on identifying climatic factors associated with their present range limits.
DOI:10.5846/stxb201405090930URL [本文引用: 1]
根据1,2,3,5,8a共5个不同年龄的15块1000 m2尾巨桉样地(3次重复)调查资料,利用18株不同年龄和径阶的样木数据,建立以胸径(D)为单变量的生物量回归方程.采用样木回归分析法(乔木层)和 样方收获法(灌木层、草本层、地上凋落物)获取不同林龄尾巨桉人工林的生物量,分析了其组成、分配及不同林龄生物量的变化趋势.结果表明:林分总生物量随 林龄而增加,1,2,3,5年生和8年生尾巨桉人工林生物量分别为12.49,47.75,64.51,105.77和137.51 t/hm2,其中活体植物占85.60%-97.61%,地上凋落物占2.39%-14.40%;层次分配方面乔木层占绝对优势,占 54.80%-91.56%,且随林龄的增加而增大,其次为凋落物,灌木层和草本层生物量较小,分别占1.02%-6.47%和 0.28%-24.33%,均随林龄的增加呈递减趋势;乔木层以干所占比例最高,占51.07%-98.48%,且随林龄而增加,枝、叶、根分别占 5.76%-11.80%,2.17%-21.01%和6.72%-14.87%,均随林龄而下降;灌木层以枝所占比例最高,为 37.89%-56.79%,叶和根分别占16.35%-34.24%和19.52%-39.52%,随林龄的变化均不大;草本层分配1-5年生以地上所 占比例较大,8年生地下所占比例高达63.87%;尾巨桉人工林乔木层各器官、地上凋落物及总生物量具有良好的优化增长模型,其总生物量的增长模型为 Y=-1.693× 104+ 3.337× 104X-1.761X2;8年生尾巨桉人工林总生物量与30年生的木莲人工林持平,低于热带雨林,但其年均净生产量高达17.19 t/hm2,是一个光合效率高、固碳潜力大的速生丰产优良造林树种.
DOI:10.5846/stxb201405090930URL [本文引用: 1]
根据1,2,3,5,8a共5个不同年龄的15块1000 m2尾巨桉样地(3次重复)调查资料,利用18株不同年龄和径阶的样木数据,建立以胸径(D)为单变量的生物量回归方程.采用样木回归分析法(乔木层)和 样方收获法(灌木层、草本层、地上凋落物)获取不同林龄尾巨桉人工林的生物量,分析了其组成、分配及不同林龄生物量的变化趋势.结果表明:林分总生物量随 林龄而增加,1,2,3,5年生和8年生尾巨桉人工林生物量分别为12.49,47.75,64.51,105.77和137.51 t/hm2,其中活体植物占85.60%-97.61%,地上凋落物占2.39%-14.40%;层次分配方面乔木层占绝对优势,占 54.80%-91.56%,且随林龄的增加而增大,其次为凋落物,灌木层和草本层生物量较小,分别占1.02%-6.47%和 0.28%-24.33%,均随林龄的增加呈递减趋势;乔木层以干所占比例最高,占51.07%-98.48%,且随林龄而增加,枝、叶、根分别占 5.76%-11.80%,2.17%-21.01%和6.72%-14.87%,均随林龄而下降;灌木层以枝所占比例最高,为 37.89%-56.79%,叶和根分别占16.35%-34.24%和19.52%-39.52%,随林龄的变化均不大;草本层分配1-5年生以地上所 占比例较大,8年生地下所占比例高达63.87%;尾巨桉人工林乔木层各器官、地上凋落物及总生物量具有良好的优化增长模型,其总生物量的增长模型为 Y=-1.693× 104+ 3.337× 104X-1.761X2;8年生尾巨桉人工林总生物量与30年生的木莲人工林持平,低于热带雨林,但其年均净生产量高达17.19 t/hm2,是一个光合效率高、固碳潜力大的速生丰产优良造林树种.
URL [本文引用: 1]
研究了湖南鹰嘴界自然保护区内马尾松(Pinus massoniana)林、马尾松阔叶树混交林和常绿阔叶林这3种处于不同演替阶段森林类型的碳储量及时空分布格局.结果表明:3种类型森林生态系统碳储量分别为182.86、179.84和229.12 Mg C·hm-2,其中乔木层占59.57% -67.88%,随森林进展演替增加,乔木层是生态系统碳储量主要贡献者,且各林分均以树干占乔术层碳储量比例最大;土壤层占31.05%~ 36.55%,碳储量随演替进展而增加,但对森林生态系统碳储量贡献率减小;林下植被和凋落物层分别占0.41% ~3.04%和0.65% ~2.53%,均随演替进展而减少,对生态系统碳储量贡献微弱;演替过程中生物量碳与土壤碳储量比为1.96、1.69和2.20,生物量碳在生态系统碳储量中所占比例呈增加趋势.可见在常绿阔叶林分布区,实施近自然林经营,模拟常绿阔叶林结构,是提升该区域森林碳汇能力的重要途径.
URL [本文引用: 1]
研究了湖南鹰嘴界自然保护区内马尾松(Pinus massoniana)林、马尾松阔叶树混交林和常绿阔叶林这3种处于不同演替阶段森林类型的碳储量及时空分布格局.结果表明:3种类型森林生态系统碳储量分别为182.86、179.84和229.12 Mg C·hm-2,其中乔木层占59.57% -67.88%,随森林进展演替增加,乔木层是生态系统碳储量主要贡献者,且各林分均以树干占乔术层碳储量比例最大;土壤层占31.05%~ 36.55%,碳储量随演替进展而增加,但对森林生态系统碳储量贡献率减小;林下植被和凋落物层分别占0.41% ~3.04%和0.65% ~2.53%,均随演替进展而减少,对生态系统碳储量贡献微弱;演替过程中生物量碳与土壤碳储量比为1.96、1.69和2.20,生物量碳在生态系统碳储量中所占比例呈增加趋势.可见在常绿阔叶林分布区,实施近自然林经营,模拟常绿阔叶林结构,是提升该区域森林碳汇能力的重要途径.
DOI:10.1029/97JD02317URL [本文引用: 1]
DOI:10.1016/S0065-2504(08)60121-XURL [本文引用: 1]
DOI:10.1890/1051-0761(2003)013[0299:FECANA]2.0.CO;2URL [本文引用: 3]
Forests of the northeastern United States are expected to serve as a substantial sink for carbon (C) and nitrogen (N) as they recover from extensive clearing and agriculture. However, questions remain concerning the rate, distribution, and duration of this potential sink. We used a chronosequence approach to determine changes in ecosystem C and N during the first 115 years of forest development after agricultural abandonment in Rhode Island, USA. All sites had similar soils, climate, land-use history, and dominant overstory vegetation (Pinus strobus), but differed in time since agricultural abandonment. Total ecosystem C increased linearly across the chronosequence at a mean rate of $2.10\>Mg\>C\!\cdot\!ha^{-1}\!\cdot\!yr^{-1}$. Most of the C was sequestered into plant biomass (73%) with less stored in the forest floor (17%) and deep mineral soil (6%; 20-70 cm depth). Total ecosystem N did not change over time; instead N accumulated in the forest floor $(11.6\>kg\>N\!\cdot\!ha^{-1}\!\cdot\!yr^{-1})$ and plant biomass $(4.1\>kg\>N\!\cdot\!ha^{-1}\!\cdot\!yr^{-1})$, and these increases were roughly balanced by a significant decrease in N content of the mineral soil $(-12.4\>kg\>N\!\cdot\!ha^{-1}\!\cdot\!yr^{-1})$. Over 90% of the storage of C and N in plant biomass occurred in the bole wood (C:N 350). Since nearly two-thirds of the forests in Rhode Island have established since 1900, net ecosystem C storage will likely continue for the next century if these forests remain undisturbed. Our results show that wood biomass is an important sink for both C and N in regrowing forests for at least one century after agricultural abandonment.
[本文引用: 2]
DOI:10.1016/j.geoderma.2006.09.003URL [本文引用: 1]
We reviewed the experimental evidence for long-term carbon (C) sequestration in soils as consequence of specific forest management strategies. Utilization of terrestrial C sinks alleviates the burden of countries which are committed to reducing their greenhouse gas emissions. Land-use changes such as those which result from afforestation and management of fast-growing tree species, have an immediate effect on the regional rate of C sequestration by incorporating carbon dioxide (CO 2) in plant biomass. The potential for such practices is limited in Europe by environmental and political constraints. The management of existing forests can also increase C sequestration, but earlier reviews found conflicting evidence regarding the effects of forest management on soil C pools. We analyzed the effects of harvesting, thinning, fertilization application, drainage, tree species selection, and control of natural disturbances on soil C dynamics. We focused on factors that affect the C input to the soil and the C release via decomposition of soil organic matter (SOM). The differentiation of SOM into labile and stable soil C fractions is important. There is ample evidence about the effects of management on the amount of C in the organic layers of the forest floor, but much less information about measurable effects of management on stable C pools in the mineral soil. The C storage capacity of the stable pool can be enhanced by increasing the productivity of the forest and thereby increasing the C input to the soil. Minimizing the disturbances in the stand structure and soil reduces the risk of unintended C losses. The establishment of mixed species forests increases the stability of the forest and can avoid high rates of SOM decomposition. The rate of C accumulation and its distribution within the soil profile differs between tree species. Differences in the stability of SOM as a direct species effect have not yet been reported.
DOI:10.1562/2005-06-08-RA-568 [本文引用: 3]
ABSTRACT textversion:publisher
DOI:10.1007/s12374-010-9140-9URL [本文引用: 4]
This study examined the biomass and carbon pools of the main ecosystem components in an age sequence of five Korean pine plantation forest stands in central Korea. The C contents in the tree and ground vegetation biomass, coarse woody debris, forest floor, and mineral soil were estimated by analyzing the C concentration of each component. The aboveground and total tree biomass increased with increasing stand age. The highest C concentration across this chronosequence was found in the tree branch while the lowest C concentration was found in the ground vegetation. The observed C contents for tree components, ground vegetation, and coarse woody debris were generally lower than the predicted C contents estimated from a biomass C factor of 0.5. Forest floor C content was age-independent. Total mineral soil C content appeared to decline initially after establishing Korean pine plantations and recover by the stand age of 35 years. Although aboveground tree biomass C content showed considerable accumulation with increasing age, the relative contribution of below ground C to total ecosystem C content varied substantially. These results suggest that successional development as temporal factor has a key role in estimating the C storage in Korean pine plantation forests.
DOI:10.5194/bg-15-1-2018URL [本文引用: 1]
URL [本文引用: 3]
应用相对生长法对桂西南28年生米老排人工林生物量及其分配特征进行了研究.结果表明:28年生米老排人 工林生物量为281.47 t·hm-2,生态系统生物量分配格局为乔木层(97.89%)>凋落物层(1.87%)>灌木层(0.16%)>草本层(0.08%);其中,乔木层生 物量为275.54 t· hm-2,其生物量在各器官的分配规律为树干(63.01%)>树根(21.01%)>树枝(9.64%)>树皮(4.38%)>树叶(1.72%)>果 实(0.25%);乔木生物量的径级分布接近正态分布,生物量主要集中在径级为25~29 cm的林木,占乔木层生物量总量的48.15%;28年生米老排人工林林分年均净生产力为15.61 t· hm-2·a-1,各组分净生产力大小顺序为乔木层(81.50%)>凋落物层(16.82%)>灌木层(0.98%)>草本层(0.70%);乔木层年 均净生产力为12.72 t·hm-2·a-1,各器官净生产力大小顺序为树干(48.76%)>树叶(18.64%)>树根(16.26%)>树枝(7.46%)>果实 (5.50%)>树皮(3.39%).
URL [本文引用: 3]
应用相对生长法对桂西南28年生米老排人工林生物量及其分配特征进行了研究.结果表明:28年生米老排人 工林生物量为281.47 t·hm-2,生态系统生物量分配格局为乔木层(97.89%)>凋落物层(1.87%)>灌木层(0.16%)>草本层(0.08%);其中,乔木层生 物量为275.54 t· hm-2,其生物量在各器官的分配规律为树干(63.01%)>树根(21.01%)>树枝(9.64%)>树皮(4.38%)>树叶(1.72%)>果 实(0.25%);乔木生物量的径级分布接近正态分布,生物量主要集中在径级为25~29 cm的林木,占乔木层生物量总量的48.15%;28年生米老排人工林林分年均净生产力为15.61 t· hm-2·a-1,各组分净生产力大小顺序为乔木层(81.50%)>凋落物层(16.82%)>灌木层(0.98%)>草本层(0.70%);乔木层年 均净生产力为12.72 t·hm-2·a-1,各器官净生产力大小顺序为树干(48.76%)>树叶(18.64%)>树根(16.26%)>树枝(7.46%)>果实 (5.50%)>树皮(3.39%).
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
DOI:10.1073/pnas.261555198URLPMID:11742094 [本文引用: 3]
The terrestrial carbon sink, as of yet unidentified, represents 15-30% of annual global emissions of carbon from fossil fuels and industrial activities. Some of the missing carbon is sequestered in vegetation biomass and, under the Kyoto Protocol of the United Nations Framework Convention on Climate Change, industrialized nations can use certain forest biomass sinks to meet their greenhouse gas emissions reduction commitments. Therefore, we analyzed 19 years of data from remote-sensing spacecraft and forest inventories to identify the size and location of such sinks. The results, which cover the years 1981-1999, reveal a picture of biomass carbon gains in Eurasian boreal and North American temperate forests and losses in some Canadian boreal forests. For the 1.42 billion hectares of Northern forests, roughly above the 30th parallel, we estimate the biomass sink to be 0.68 卤 0.34 billion tons carbon per year, of which nearly 70% is in Eurasia, in proportion to its forest area and in disproportion to its biomass carbon pool. The relatively high spatial resolution of these estimates permits direct validation with ground data and contributes to a monitoring program of forest biomass sinks under the Kyoto protocol.
DOI:10.1016/j.foreco.2009.12.009URL [本文引用: 1]
Forest soil carbon (C) storage is a significant component of the global C cycle, and is important for sustaining forest productivity. Although forest management may have substantial impacts on soil C storage, experimental data from forest harvesting studies have not been synthesized recently. To quantify the effects of harvesting on soil C, and to identify sources of variation in soil C responses to harvest, we used meta-analysis to test a database of 432 soil C response ratios drawn from temperate forest harvest studies around the world. Harvesting reduced soil C by an average of 8 ± 3% (95% CI), although numerous sources of variation mediated this significant, overall effect. In particular, we found that C concentrations and C pool sizes responded differently to harvesting, and forest floors were more likely to lose C than mineral soils. Harvesting caused forest floor C storage to decline by a remarkably consistent 30 ± 6%, but losses were significantly smaller in coniferous/mixed stands (6120%) than hardwoods (6136%). Mineral soils showed no significant, overall change in C storage due to harvest, and variation among mineral soils was best explained by soil taxonomy. Alfisols and Spodosols exhibited no significant changes, and Inceptisols and Ultisols lost mineral soil C (6113% and 617%, respectively). However, these C losses were neither permanent nor unavoidable. Controls on variation within orders were not consistent, but included species composition, time, and sampling depth. Temporal patterns and soil C budgets suggest that forest floor C losses probably have a lesser impact on total soil C storage on Alfisols, Inceptisols, and Ultisols than on Spodosols, which store proportionately large amounts of C in forest floors with long C recovery times (50–70 years). Mineral soil C losses on Inceptisols and Ultisols indicate that these orders are vulnerable to significant harvest-induced changes in total soil C storage, but alternative residue management and site preparation techniques, and the passage of time, may mitigate or negate these losses. Key findings of this analysis, including the dependence of forest floor and mineral soil C storage changes on species composition and soil taxonomic order, suggest that further primary research may make it possible to create predictive maps of forest harvesting effects on soil C storage.
[本文引用: 1]
DOI:10.1126/science.1201609URLPMID:174893267101550638 [本文引用: 1]
The terrestrial carbon sink has been large in recent decades, but its size and location remain uncertain. Using forest inventory data and long-term ecosystem carbon studies, we estimate a total forest sink of 2.4 ± 0.4 petagrams of carbon per year (Pg C year
DOI:10.1016/S0378-1127(01)00740-XURL [本文引用: 17]
Quantifying changes in soil C may be an important consideration under large-scale afforestation or reforestation. We reviewed global data on changes in soil C following afforestation, available from 43 published or unpublished studies, encompassing 204 sites. Data were highly variable, with soil C either increasing or decreasing, particularly in young (<10-year) forest stands. Because studies varied in the number of years since forest establishment and the initial soil C content, we calculated change in soil C as a weighted-average (i.e. sum of C change divided by sum of years since forest establishment) relative to the soil C content under previous agricultural systems at <10, >10 and <30 cm sampling depths. On average, soil C in the <10 cm (or <30 cm) layers generally decreased by 3.46% per year (or 0.63% per year) relative to the initial soil C content during the first 5 years of afforestation, followed by a decrease in the rate of decline and eventually recovery to C contents found in agricultural soils at about age 30. In plantations older than 30 years, C content was similar to that under the previous agricultural systems within the surface 10 cm of soil, yet at other sampling depths, soil C had increased by between 0.50 and 0.86% per year. Amounts of C lost or gained by soil are generally small compared with accumulation of C in tree biomass. The most important factors affecting change in soil C were previous land use, climate and the type of forest established. Results suggest that most soil C was lost when softwoods, particularly Pinus radiata plantations, were established on ex-improved pastoral land in temperate regions. Accumulation of soil C was greatest when deciduous hardwoods, or N 2-fixing species (either as an understorey or as a plantation), were established on ex-cropped land in tropical or subtropical regions. Long-term management regimes (e.g. stocking, weed control, thinning, fertiliser application and fire management) may also influence accumulation of soil C. Accumulation is maximised by maintaining longer (20 50 years) forest rotations. Furthermore, inclusion of litter in calculations reversed the observed average decrease in soil C, so that amount of C in soil and litter layer was greater than under preceding pasture.
DOI:10.1016/j.agrformet.2006.08.004URL [本文引用: 1]
We assessed the successional development of above- and belowground ecosystem biomass and carbon (C) pools in an age-sequence of four White pine ( Pinus strobus L.) plantation stands (2-, 15-, 30-, and 65-years-old) in Southern Ontario, Canada. Biomass and C stocks of above- and belowground live and dead tree biomass, understorey and forest ground vegetation, forest floor C (LFH-layer), and woody debris were determined from plot-level inventories and destructive tree sampling. Small root biomass (<5 mm) and mineral soil C stocks were estimated from soil cores. Aboveground tree biomass became the major ecosystem C pool with increasing age, reaching 0.5, 66, 92, and 176 t ha 611 in the 2-, 15-, 30-, and 65-year-old stands, respectively. Tree root biomass increased from 0.1 to 10, 18, 38 t ha 611 in the 2-, 15-, 30-, and 65-year-old stands, respectively, contributing considerably to the total ecosystem C in the three older stands. Forest floor C was 0.8, 7.5, 5.4, and 12.1 t C ha 611 in the 2-, 15-, 30-, and 65-year-old stands, respectively, indicating an increase during the first two decades, but no further age-effect during the later growth phase. Mineral soil C was age-independent with 37.2, 33.9, 39.1, and 36.7 t C ha 611 in the 2-, 15-, 30-, and 65-year-old stands, respectively. Aboveground ecosystem C increased with age from 3 to 40, 52, and 100 t C ha 611 in the 2-, 15-, 30-, and 65-year-old stands, respectively, due to an increase in aboveground tree biomass. Belowground ecosystem C remained similiar in the early decades after establishment with 37, 39, and 39 t C ha 611 in the 2-, 15-, and 30-year-old stands, but increased to 56 t C ha 611 in the 65-year-old stand due to an increase in root biomass. The difference in total ecosystem C between the 2- and 65-year-old stand was 116 t C ha 611. Our results highlight the importance of considering the successional development of forest ecosystem C pools, when estimating C sink potentials over their complete life cycle.
DOI:10.1111/j.1365-2486.2004.00881.x [本文引用: 13]
Forest soils store a substantial amount of carbon, often more than the forest vegetation does. Estimates of the amount of soil carbon, and in particular estimates of changes in these amounts are still inaccurate. Measuring soil carbon is laborious, and measurements taken at a few statistically unrepresentative sites are difficult to scale to larger areas.
DOI:10.1111/gcb.2004.10.issue-12URL [本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.1051/forest:2005079URL [本文引用: 2]
DOI:10.2307/1941147URL [本文引用: 1]
DOI:10.1890/1051-0761(1997)007[0702:CWDAAF]2.0.CO;2URL [本文引用: 1]
We examined the structural attributes of coarse woody debris (CWD) in balsam fir and mixed balsam fir-black spruce forests in western and central Newfoundland. The purpose of the study as to document CWD abundance and structure within various stages of stand development, and to generate simple models to predict CWD availability as habitat for forest wildlife. Our objectives were threefold: (1) determine the temporal patterns of CWD following clear-cut disturbance, and compare these second-growth stands to old sites ($>$80 yr) or natural origin; (2) relate the temporal patterns of CWD to stand development; and (3) demonstrate how factors such as site conditions and disturbance may influence these temporal patterns. Our chronosequence included 19 second-growth stands with a mean tree age ranging from 33 to 80 yr, and 7 old-growth stands ranging from 87 to 110 yr. The volume of coarse woody debris (CWD) here defined as downed wood, was relatively low to intermediate early in the chronosequence (e.g., 32.3 m$^3$/ha in a 36-yr-old stand), lowest in a 58-yr-old stand (15.2 m$^3$/ha), and highest in an 80-yr-old stand (78.1 m$^3$/ha). Results indicated that CWD volume followed the general "U-shaped" temporal trend observed in other forest systems. The presence of CWD early in the sequence was strongly influenced by residual hardwoods (i.e., birch) left by the original logging operation. The highest observed volume of CWD corresponded with stand senescence and appeared to be maintained within old-growth stands. Snag (standing dead wood) density was low within the youngest stands ($<$200 snags/ha), and peaked within mature second growth. Defoliation disturbance increased both the volume and the structural diversity of CWD within silviculturallly mature second-growth stands. Results from our study indicate that CWD in stands $>$50-60 yr of age is not residual but generated primarily from regenerating tree structure. Factors affecting the rate of stem growth, e.g., site quality and initial stocking levels, should influence the accumulation and overall abundance of CWD within later stages of forests development. We therefore applied yield-density relationships as a method of examining stand-level dynamics of CWD.
DOI:10.17521/cjpe.2005.0075URL [本文引用: 1]
森林生态系统中的粗死木质残体(Coarse woody debris,CWD)不仅能够为其它生物提供生境,维持森林结构,而且对生物地球化学循环起着不可忽视的作用,CWD作为森林生态系统中重要的结构和功 能元素,已经引起广泛关注.然而,华南地区典型亚热带森林生态系统中CWD的结构和功能方面的研究很少.该文报道了鼎湖山自然保护区内典型南亚热带森林生 态系统中CWD的贮量及其特征,所选择的森林包括马尾松(Pinus massoniana)林、针阔叶混交林和季风常绿阔叶林,它们分别代表该气候区域内处于森林演替早期、中期和后期3个阶段的森林类型.其中马尾松林和针 阔叶混交林都起源于20世纪30年代人工种植的马尾松纯林,由于长期受到包括收割松针、CWD和林下层植物等在内的人为活动的干扰,到2003年调查时马 尾松林仍属于针叶林;而混交林样地自种植之后就未受到人为活动的干扰,自然过渡为针阔叶混交林类型.人为干扰对马尾松人工林的结构和功能产生了巨大的影 响,马尾松林的生物量仅为针阔叶混交林生物量的35%.组成马尾松林、针阔叶混交林和季风常绿阔叶林CWD的树种数量分别为7、18和29;马尾松林中几 乎没有CWD存在(贮量仅为0.1 MgC·hm.),针阔叶混交林CWD的贮量为8.7 Mg C·hm-2,季风常绿阔叶林CWD的贮量为13.2 Mg C·hm-2,分别占地上部分生物量的9.1%和11.3%;针阔叶混交林和季风常绿阔叶林中只有将近10%的CWD以枯立的方式存在.该区域内CWD的 分解速率较快,在区域碳循环中将扮演重要角色,保留林地中的CWD是维持本区域森林生产力和森林可持续管理的重要举措.
DOI:10.17521/cjpe.2005.0075URL [本文引用: 1]
森林生态系统中的粗死木质残体(Coarse woody debris,CWD)不仅能够为其它生物提供生境,维持森林结构,而且对生物地球化学循环起着不可忽视的作用,CWD作为森林生态系统中重要的结构和功 能元素,已经引起广泛关注.然而,华南地区典型亚热带森林生态系统中CWD的结构和功能方面的研究很少.该文报道了鼎湖山自然保护区内典型南亚热带森林生 态系统中CWD的贮量及其特征,所选择的森林包括马尾松(Pinus massoniana)林、针阔叶混交林和季风常绿阔叶林,它们分别代表该气候区域内处于森林演替早期、中期和后期3个阶段的森林类型.其中马尾松林和针 阔叶混交林都起源于20世纪30年代人工种植的马尾松纯林,由于长期受到包括收割松针、CWD和林下层植物等在内的人为活动的干扰,到2003年调查时马 尾松林仍属于针叶林;而混交林样地自种植之后就未受到人为活动的干扰,自然过渡为针阔叶混交林类型.人为干扰对马尾松人工林的结构和功能产生了巨大的影 响,马尾松林的生物量仅为针阔叶混交林生物量的35%.组成马尾松林、针阔叶混交林和季风常绿阔叶林CWD的树种数量分别为7、18和29;马尾松林中几 乎没有CWD存在(贮量仅为0.1 MgC·hm.),针阔叶混交林CWD的贮量为8.7 Mg C·hm-2,季风常绿阔叶林CWD的贮量为13.2 Mg C·hm-2,分别占地上部分生物量的9.1%和11.3%;针阔叶混交林和季风常绿阔叶林中只有将近10%的CWD以枯立的方式存在.该区域内CWD的 分解速率较快,在区域碳循环中将扮演重要角色,保留林地中的CWD是维持本区域森林生产力和森林可持续管理的重要举措.
DOI:10.3969/j.issn.1004-9398.2007.02.014URL [本文引用: 1]
通过对神农架地区广泛分布的米心水青冈林和锐齿槲栎林的生物量特征的调查研究表明:神农架地区102-130 a成熟米心水青冈林的生物量在251.30~358.63 T·hm-2之间,平均为288.70±48.30 T·hm-2,20~60 a锐齿槲栎林群落生物量在134.85~301.20 T·hm-2,平均为231.60±78.10 T·hm-2.两种类型森林生物量的95%以上集中在乔木层.在米心水青冈林生物量从大到小的序列中,前5种植物分别占乔木层总生物量 58.67%~96.37%不等,同样锐齿槲栎林中,前5种植物占群落生物量的68.13%~95.26%.常绿植物占乔木层生物量的比例变化较大,米心 水青冈林中占2.85~18.70%,锐齿槲栎林中一般常绿植物占0.8%~9.98%,只有一个锐齿槲栎林样地常绿植物(主要是粉白杜鹃)占乔木层生物 量的44.04%.米心水青冈生物量根冠比为0.27±0.05,锐齿槲栎林为0.21±0.06.神农架地区米心水青冈林的生物量,在成熟的欧洲水青冈 林和日本的水青冈林生物量范围之内,而锐齿槲栎林的生物量远远大于我国温带落叶栎林的生物量.
DOI:10.3969/j.issn.1004-9398.2007.02.014URL [本文引用: 1]
通过对神农架地区广泛分布的米心水青冈林和锐齿槲栎林的生物量特征的调查研究表明:神农架地区102-130 a成熟米心水青冈林的生物量在251.30~358.63 T·hm-2之间,平均为288.70±48.30 T·hm-2,20~60 a锐齿槲栎林群落生物量在134.85~301.20 T·hm-2,平均为231.60±78.10 T·hm-2.两种类型森林生物量的95%以上集中在乔木层.在米心水青冈林生物量从大到小的序列中,前5种植物分别占乔木层总生物量 58.67%~96.37%不等,同样锐齿槲栎林中,前5种植物占群落生物量的68.13%~95.26%.常绿植物占乔木层生物量的比例变化较大,米心 水青冈林中占2.85~18.70%,锐齿槲栎林中一般常绿植物占0.8%~9.98%,只有一个锐齿槲栎林样地常绿植物(主要是粉白杜鹃)占乔木层生物 量的44.04%.米心水青冈生物量根冠比为0.27±0.05,锐齿槲栎林为0.21±0.06.神农架地区米心水青冈林的生物量,在成熟的欧洲水青冈 林和日本的水青冈林生物量范围之内,而锐齿槲栎林的生物量远远大于我国温带落叶栎林的生物量.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.3969/j.issn.1673-923X.2003.01.001URL [本文引用: 1]
为了合理开发利用樟树人工林资源,为进一步研究樟树人工林生态系统提供基础数据,对18年生樟树人工林生物量的结构与分布特点进行了研究.结果表明:18 年生樟树人工林分生物量为111.08 t/hm2,年平均净生产力为6.17 t*hm-2a-1,生态系统的生物量分配格局为乔木层>枯枝落叶层>下木层>草本层,其中乔木层生物量为91.97 t*hm-2,净生产力为5.11 t*hm-2a-1,其生物量分配格局为树干>树枝>树根>树皮>树叶;在林分产量结构方面,5 m以下树干生物量占其总量的80%,树枝的生物量主要分布在4~11 m,占其总量的86%,树叶的生物量主要分布在8~12 m,占树叶总生物量的78%,地下根系在离地面40 cm深土层内的生物量占总根量的84.74%.目前,该林分群落结构不合理,应调整群落产量结构,以提高其综合效益.
DOI:10.3969/j.issn.1673-923X.2003.01.001URL [本文引用: 1]
为了合理开发利用樟树人工林资源,为进一步研究樟树人工林生态系统提供基础数据,对18年生樟树人工林生物量的结构与分布特点进行了研究.结果表明:18 年生樟树人工林分生物量为111.08 t/hm2,年平均净生产力为6.17 t*hm-2a-1,生态系统的生物量分配格局为乔木层>枯枝落叶层>下木层>草本层,其中乔木层生物量为91.97 t*hm-2,净生产力为5.11 t*hm-2a-1,其生物量分配格局为树干>树枝>树根>树皮>树叶;在林分产量结构方面,5 m以下树干生物量占其总量的80%,树枝的生物量主要分布在4~11 m,占其总量的86%,树叶的生物量主要分布在8~12 m,占树叶总生物量的78%,地下根系在离地面40 cm深土层内的生物量占总根量的84.74%.目前,该林分群落结构不合理,应调整群落产量结构,以提高其综合效益.
DOI:10.1073/pnas.1317065111URL [本文引用: 1]
Temperate- and high-latitude forests have been shown to contribute a carbon sink in the Northern Hemisphere, but fewer studies have addressed the carbon balance of the subtropical forests. In the present study, we integrated eddy covariance observations established in the 1990s and 2000s to show that East Asian monsoon subtropical forests between 20 degrees N and 40 degrees N represent an average net ecosystem productivity (NEP) of 362 +/- 39 g C m(-2) yr(-1) (mean +/- 1 SE). This average forest NEP value is higher than that of Asian tropical and temperate forests and is also higher than that of forests at the same latitudes in Europe-Africa and North America. East Asian monsoon subtropical forests have comparable NEP to that of subtropical forests of the southeastern United States and intensively managed Western European forests. The total NEP of East Asian monsoon subtropical forests was estimated to be 0.72 +/- 0.08 Pg C yr(-1), which accounts for 8% of the global forest NEP. This result indicates that the role of subtropical forests in the current global carbon cycle cannot be ignored and that the regional distributions of the Northern Hemisphere's terrestrial carbon sinks are needed to be reevaluated. The young stand ages and high nitrogen deposition, coupled with sufficient and synchronous water and heat availability, may be the primary reasons for the high NEP of this region, and further studies are needed to quantify the contribution of each underlying factor.
URL [本文引用: 1]
量化森林碳储量及其分配格局是森林碳循环和陆地生态系统模型的重要研究内容.采用样地清查和异速生长方程法测定了相同气候条件下林龄相近(42~59年生)的6种典型温带森林类型(杨桦林、硬阔叶林、红松林、兴安落叶松林、杂木林和蒙古栎林)的碳密度和碳分配格局.结果表明,所处的立地条件和植被组成不同的6种温带森林,其生态系统组分碳密度(除碎屑碳库外)差异不显著,但用林分胸高断面积标准化之后则存在显著差异.6种温带森林的总碳密度在186.9~349.2tC/hm2之间波动;其中,植被碳密度、碎屑碳密度和土壤碳密度分别在86.3~122.7,6.5~10.5和93.7~220.1tC/hm2之间波动,占总碳密度的(39.7±7.1)%(均值±标准差),(3.3±1.1)%和(57.0±7.9)%.在植被碳库中,乔木层占99%以上.叶生物量、中细根(直径5mm)生物量、根冠比、中细根生物量与叶生物量之比分别在2.08~4.72tC/hm2,0.95~3.24tC/hm2,22.0%~28.3%,34.5%~122.2%之间波动.6种森林中,红松林的叶生产效率(总生物量与叶生物量之比)最低(22.6g/g),兴安落叶松林的中细根生产效率(总生物量与中细根生物量之比)最高(124.7g/g).除蒙古栎林外,其他5种森林的中细根碳密度(包括死活中细根量)均随土壤层次加深而下降;而蒙古栎林的中细根碳密度的垂直分布却有下移趋势.两种人工林(红松林和兴安落叶松林)的粗木质残体碳密度显著低于4种天然林.本研究指出,特定森林碳分配格局的分异主要受植被类型、经营历史、局域土壤的水分和养分有效性等因素的共同作用,同时也为温带森林碳循环模型提供了重要的构建和校验参数.
URL [本文引用: 1]
量化森林碳储量及其分配格局是森林碳循环和陆地生态系统模型的重要研究内容.采用样地清查和异速生长方程法测定了相同气候条件下林龄相近(42~59年生)的6种典型温带森林类型(杨桦林、硬阔叶林、红松林、兴安落叶松林、杂木林和蒙古栎林)的碳密度和碳分配格局.结果表明,所处的立地条件和植被组成不同的6种温带森林,其生态系统组分碳密度(除碎屑碳库外)差异不显著,但用林分胸高断面积标准化之后则存在显著差异.6种温带森林的总碳密度在186.9~349.2tC/hm2之间波动;其中,植被碳密度、碎屑碳密度和土壤碳密度分别在86.3~122.7,6.5~10.5和93.7~220.1tC/hm2之间波动,占总碳密度的(39.7±7.1)%(均值±标准差),(3.3±1.1)%和(57.0±7.9)%.在植被碳库中,乔木层占99%以上.叶生物量、中细根(直径5mm)生物量、根冠比、中细根生物量与叶生物量之比分别在2.08~4.72tC/hm2,0.95~3.24tC/hm2,22.0%~28.3%,34.5%~122.2%之间波动.6种森林中,红松林的叶生产效率(总生物量与叶生物量之比)最低(22.6g/g),兴安落叶松林的中细根生产效率(总生物量与中细根生物量之比)最高(124.7g/g).除蒙古栎林外,其他5种森林的中细根碳密度(包括死活中细根量)均随土壤层次加深而下降;而蒙古栎林的中细根碳密度的垂直分布却有下移趋势.两种人工林(红松林和兴安落叶松林)的粗木质残体碳密度显著低于4种天然林.本研究指出,特定森林碳分配格局的分异主要受植被类型、经营历史、局域土壤的水分和养分有效性等因素的共同作用,同时也为温带森林碳循环模型提供了重要的构建和校验参数.
DOI:10.11926/j.issn.1005-3395.2016.06.004URL [本文引用: 1]
为了解贵州省月亮山森林土壤的化学计量特征,分层采集5种森林类型的土壤,测定C、N、P含量并分析化学计量特征。结果表明,5种森林类型0~20 cm土壤中变异最大的是P,20~100 cm土壤中变异最大的是C,最小的是N。5种森林类型0~10 cm土壤的C、N含量远高于中国陆地0~10 cm土壤的平均含量,而P含量远低于其平均含量。硬阔和软阔自然林土壤C、N、P含量存在明显差异,C含量为硬阔自然林软阔自然林,N和P含量基本呈现为软阔自然林硬阔自然林。不同森林类型土壤的C∶N为硬阔自然林软阔自然林,针阔混交林杉木和马尾松纯林。硬阔和软阔自然林、针阔混交林的C∶P、N∶P均随土层深度的增加而下降,而杉木和马尾松纯林的C∶P、N∶P随土层深度增加呈先下降后升高再下降的趋势,且均在30~50 cm处出现累积峰。相关分析表明0~100 cm土壤的C、N、P含量呈极显著正相关关系,土壤C含量、N含量与C∶N、C∶P、N∶P均呈极显著的正相关关系,P含量与C∶N呈极显著正相关关系(P0.001)。因此,贵州月亮山5种森林类型和不同土层的C、N、P含量及其化学计量特征存在显著差异。
DOI:10.11926/j.issn.1005-3395.2016.06.004URL [本文引用: 1]
为了解贵州省月亮山森林土壤的化学计量特征,分层采集5种森林类型的土壤,测定C、N、P含量并分析化学计量特征。结果表明,5种森林类型0~20 cm土壤中变异最大的是P,20~100 cm土壤中变异最大的是C,最小的是N。5种森林类型0~10 cm土壤的C、N含量远高于中国陆地0~10 cm土壤的平均含量,而P含量远低于其平均含量。硬阔和软阔自然林土壤C、N、P含量存在明显差异,C含量为硬阔自然林软阔自然林,N和P含量基本呈现为软阔自然林硬阔自然林。不同森林类型土壤的C∶N为硬阔自然林软阔自然林,针阔混交林杉木和马尾松纯林。硬阔和软阔自然林、针阔混交林的C∶P、N∶P均随土层深度的增加而下降,而杉木和马尾松纯林的C∶P、N∶P随土层深度增加呈先下降后升高再下降的趋势,且均在30~50 cm处出现累积峰。相关分析表明0~100 cm土壤的C、N、P含量呈极显著正相关关系,土壤C含量、N含量与C∶N、C∶P、N∶P均呈极显著的正相关关系,P含量与C∶N呈极显著正相关关系(P0.001)。因此,贵州月亮山5种森林类型和不同土层的C、N、P含量及其化学计量特征存在显著差异。
DOI:10.1007/s11427-015-4935-zURLPMID:26501378 [本文引用: 2]
a, respectively. Corresponding net ecosystem production was 0.12, 0.43, and 3.53 Mg C ha a. The C stocks and fluxes in 2011-2014 were significantly larger than those in 1992-1994 in which the biomass C densities in birch, oak, and pine plots increased from 50.0, 37.7, and 54.0 Mg C ha-1 to 214.8, 241.7, and 238.4 Mg C ha; and soil heterotrophic respiration increased from 2.78, 3.49, and 1.81 Mg C ha a to 5.20, 4.10, and 3.20 Mg C ha a. These results suggest that the mountainous temperate forest ecosystems in Beijing have served as a carbon sink in the last two decades. These observations of C stocks and fluxes provided field-based data for a long-term study of C cycling in temperate forest ecosystems.
DOI:10.1038/s41467-017-00207-1URLPMID:5532249 [本文引用: 7]
Forests play an important role in global carbon cycles. However, the lack of available information on carbon stocks in dead organic matter, including woody debris and litter, reduces the reliability of assessing the carbon cycles in entire forest ecosystems. Here we estimate that the national DOM carbon stock in the period of 2004–2008 is 92565±655465Tg, with an average density of 5.9565±650.3565Mg C65ha611. Over the past two decades from periods of 1984611988 to 2004612008, the national dead organic matter carbon stock has increased by 6.765±652.265Tg carbon per year, primarily due to increasing forest area. Temperature and precipitation increase the carbon density of woody debris, but decrease that of litter. Additionally, the woody debris increases significantly with above ground biomass and forest age. Our results can improve estimates of the carbon budget in China's forests and for better understanding of effects of climate and stand characteristics on dead organic matter distribution. Reliable estimates of the total forest carbon (C) pool are lacking due to insufficient information on dead organic matter (DOM). Here, the authors estimate that the current DOM C stock in China is 92565±655465Tg and that it grew by 6.765±652.265Tg C/yr over the past two decades primarily due to increasing forest area
[本文引用: 10]
An age-altitude matrix analysis of Hawaiian rain-forest succession
1
1994
... 全球森林碳储量达到861 Pg (1 Pg = 1015 g), 并以每年2.4 Pg的速度增长, 在缓解大气CO2浓度升高方面具有重要作用(
Age-related patterns of forest complexity and carbon storage in pine and aspen-?birch ecosystems of northern Minnesota, USA
8
2010
... 林龄是影响森林生态系统碳储量的重要因素(
... 而不同演替阶段亮叶水青冈林的灌木层与草本层碳储量皆无显著差异.有关林下生物量随林龄增长的变化趋势目前并无一致的结论.一些研究发现, 在幼龄林阶段, 林下生物量随林龄增长而降低; 而此后林龄进一步增长时(约40年以后), 林下层生物量基本无明显变化趋势, 如美国明尼苏达州北部7-160年Pinus resinosa林和6-133年杨树(Populus tremuloides和P. grandidentata)林(
... 有关凋落物随林龄的变化趋势, 不同研究的结果不尽相同.如全球200年以下温带森林(
... ;
... ).也有研究报道凋落物碳储量与林龄无关, 如北美6-133年杨树林(
... ;
... ).此外, 凋落物碳储量随林龄的变化与森林类型也有关(
... ).本研究中, 随演替的进行, 植被碳储量显著增加, 导致凋落物和木质残体的产量也随之增加; 而由于33年林为砍伐后次生, 木质残体与凋落物层受人为干扰和破坏较大, 增加了其流失和分解的速率(
Principles of Terrestrial Ecosystem Ecology. Springer-Verlag
1
2002
... 林龄是影响森林生态系统碳储量的重要因素(
湖南会同红栲-青冈-刨花楠群落生物生产力的研究
1
2000
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
湖南会同红栲-青冈-刨花楠群落生物生产力的研究
1
2000
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
Climatic limits for the present distribution of beech ( Fagus L.) species in the world
1
2006
... 亚热带森林在陆地生态系统碳循环中起着重要作用.有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (
不同林龄尾巨桉人工林的生物量及其分配特征
1
2014
... 林龄是影响森林生态系统碳储量的重要因素(
不同林龄尾巨桉人工林的生物量及其分配特征
1
2014
... 林龄是影响森林生态系统碳储量的重要因素(
中亚热带常绿阔叶林不同演替阶段碳储量与格局特征
1
2011
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
中亚热带常绿阔叶林不同演替阶段碳储量与格局特征
1
2011
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
Carbon distribution and aboveground net primary production in aspen, jack pine, and black spruce stands in Saskatchewan and Manitoba, Canada
1
1997
... 全球森林碳储量达到861 Pg (1 Pg = 1015 g), 并以每年2.4 Pg的速度增长, 在缓解大气CO2浓度升高方面具有重要作用(
Ecology of coarse woody debris in temperate ecosystems
1
1986
... 将木质残体区分为细木质残体(大头直径2-10 cm; FWD)和粗木质残体(大头直径≥10 cm; CWD) (
Forest ecosystem carbon and nitrogen accumulation during the first century after agricultural abandonment
3
2003
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... ).而对于土壤碳储量, 随林龄增加通常也表现为碳积累过程(
... 在一定林龄范围内, 随林龄增加, 森林土壤通常表现为碳积累过程, 即土壤碳储量一般呈增加趋势(
2
2013
... 全球森林碳储量达到861 Pg (1 Pg = 1015 g), 并以每年2.4 Pg的速度增长, 在缓解大气CO2浓度升高方面具有重要作用(
... 有关凋落物随林龄的变化趋势, 不同研究的结果不尽相同.如全球200年以下温带森林(
How strongly can forest management influence soil carbon sequestration?
1
2007
... 亚热带森林在陆地生态系统碳循环中起着重要作用.有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (
Primary productivity changes for a fifteen-?year period in a natural beech ( Fagus crenata) forest in the Naeba mountains
3
1991
... 林龄是影响森林生态系统碳储量的重要因素(
... 亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低.这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(
... 本研究调查了贵州榕江县月亮山不同演替阶段的9个亮叶水青冈林生态系统的全组分碳储量及各组分的相对贡献.可能会给结果带来不确定性的因素主要有两方面: 一是由于保护区禁止采伐树木, 本研究在估算乔木生物量时利用的是文献中已有的生物量方程(
Biomass and carbon storage in an age-sequence of Korean pine (Pinus koraiensis) plantation forests in central Korea
4
2011
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... ;
... 而不同演替阶段亮叶水青冈林的灌木层与草本层碳储量皆无显著差异.有关林下生物量随林龄增长的变化趋势目前并无一致的结论.一些研究发现, 在幼龄林阶段, 林下生物量随林龄增长而降低; 而此后林龄进一步增长时(约40年以后), 林下层生物量基本无明显变化趋势, 如美国明尼苏达州北部7-160年Pinus resinosa林和6-133年杨树(Populus tremuloides和P. grandidentata)林(
... 本研究调查了贵州榕江县月亮山不同演替阶段的9个亮叶水青冈林生态系统的全组分碳储量及各组分的相对贡献.可能会给结果带来不确定性的因素主要有两方面: 一是由于保护区禁止采伐树木, 本研究在估算乔木生物量时利用的是文献中已有的生物量方程(
Variations and determinants of carbon content in plants: A global synthesis
1
2018
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
桂西南28年生米老排人工林生物量及其分配特征
3
2012
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... 而不同演替阶段亮叶水青冈林的灌木层与草本层碳储量皆无显著差异.有关林下生物量随林龄增长的变化趋势目前并无一致的结论.一些研究发现, 在幼龄林阶段, 林下生物量随林龄增长而降低; 而此后林龄进一步增长时(约40年以后), 林下层生物量基本无明显变化趋势, 如美国明尼苏达州北部7-160年Pinus resinosa林和6-133年杨树(Populus tremuloides和P. grandidentata)林(
... 亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低.这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(
桂西南28年生米老排人工林生物量及其分配特征
3
2012
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... 而不同演替阶段亮叶水青冈林的灌木层与草本层碳储量皆无显著差异.有关林下生物量随林龄增长的变化趋势目前并无一致的结论.一些研究发现, 在幼龄林阶段, 林下生物量随林龄增长而降低; 而此后林龄进一步增长时(约40年以后), 林下层生物量基本无明显变化趋势, 如美国明尼苏达州北部7-160年Pinus resinosa林和6-133年杨树(Populus tremuloides和P. grandidentata)林(
... 亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低.这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(
不同林龄格木人工林碳储量及其分配特征
3
2014
... 亚热带森林在陆地生态系统碳循环中起着重要作用.有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (
... 本研究中, 不同演替阶段亮叶水青冈林生态系统碳储量从33年林的186.9 Mg·hm-2增加到208年林的515.1 Mg·hm-2.贵州梵净山44-185年水青冈(F. longipetiolata和F. lucida)林的生态系统碳储量在190.7-503.9 Mg·hm-2之间(本课题组未发表数据); 德国Thuringia中部30-171年欧洲水青冈(F. sylvatica)林的碳储量为126-413 Mg·hm-2 (
... 亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低.这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(
不同林龄格木人工林碳储量及其分配特征
3
2014
... 亚热带森林在陆地生态系统碳循环中起着重要作用.有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (
... 本研究中, 不同演替阶段亮叶水青冈林生态系统碳储量从33年林的186.9 Mg·hm-2增加到208年林的515.1 Mg·hm-2.贵州梵净山44-185年水青冈(F. longipetiolata和F. lucida)林的生态系统碳储量在190.7-503.9 Mg·hm-2之间(本课题组未发表数据); 德国Thuringia中部30-171年欧洲水青冈(F. sylvatica)林的碳储量为126-413 Mg·hm-2 (
... 亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低.这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(
Carbon Pools of European Beech Forests (Fagus sylvatica) under Different Silvicultural Management
2
2004
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
... 本研究调查了贵州榕江县月亮山不同演替阶段的9个亮叶水青冈林生态系统的全组分碳储量及各组分的相对贡献.可能会给结果带来不确定性的因素主要有两方面: 一是由于保护区禁止采伐树木, 本研究在估算乔木生物量时利用的是文献中已有的生物量方程(
A large carbon sink in the woody biomass of northern forests
3
2001
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... 本研究中, 不同演替阶段亮叶水青冈林土壤总碳储量无显著差异; 而0-10 cm土壤碳储量在33年林最高, 82年林最低.这种变化格局可能与研究样地的土地利用历史有关(
... ;
Harvest impacts on soil carbon storage in temperate forests
1
2010
... 林龄是影响森林生态系统碳储量的重要因素(
Carbon and nitrogen storage in an age-sequence of Pinus densiflora stands in Korea
1
2010
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
A large and persistent carbon sink in the world’s forests
1
2011
... 全球森林碳储量达到861 Pg (1 Pg = 1015 g), 并以每年2.4 Pg的速度增长, 在缓解大气CO2浓度升高方面具有重要作用(
Change in soil carbon following afforestation
17
2002
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... ;
... ), 然而林龄对土壤碳积累的作用可能受到土地利用历史、气候和植被类型等多种因素的影响, 故随演替进行, 土壤碳储量也可能不变或呈现先减少后增加的趋势(
... ;
... 林龄是影响森林生态系统碳储量的重要因素(
... 而不同演替阶段亮叶水青冈林的灌木层与草本层碳储量皆无显著差异.有关林下生物量随林龄增长的变化趋势目前并无一致的结论.一些研究发现, 在幼龄林阶段, 林下生物量随林龄增长而降低; 而此后林龄进一步增长时(约40年以后), 林下层生物量基本无明显变化趋势, 如美国明尼苏达州北部7-160年Pinus resinosa林和6-133年杨树(Populus tremuloides和P. grandidentata)林(
... ).本研究中, 不同演替阶段亮叶水青冈林郁闭度接近, 故灌木层与草本层碳储量无显著差异.此外, 林下生物量还受到土壤养分状况、干扰和管理方式等多种因素的影响(
... 有关凋落物随林龄的变化趋势, 不同研究的结果不尽相同.如全球200年以下温带森林(
... 在一定林龄范围内, 随林龄增加, 森林土壤通常表现为碳积累过程, 即土壤碳储量一般呈增加趋势(
... ;
... ), 或随林龄增长先降低后增加, 其中0-10 cm土壤碳储量约30年后才可以恢复到造林前的水平(
... 本研究中, 不同演替阶段亮叶水青冈林土壤总碳储量无显著差异; 而0-10 cm土壤碳储量在33年林最高, 82年林最低.这种变化格局可能与研究样地的土地利用历史有关(
... ;
... ).33年亮叶水青冈林为砍伐后次生, 此前该样地原生植被林龄可能较大, 有较多的碳积累, 故其土壤碳储量值较高.尽管砍伐等干扰通常会使土壤碳储量减少, 尤其是表层土, 但一般在一定时间后可以恢复(
... ).所以本研究中, 林龄较小的次生林的土壤碳储量与林龄较大的原始林相当, 甚至表层土壤碳储量高于林龄较大的样地, 主要取决于33年林土壤中原有的碳储量较高.此外, 相比于深层的矿质土壤, 表层土壤更容易受到干扰等因素的影响, 故其变异也更大(
... ;
... 亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低.这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(
Above- and belowground ecosystem biomass and carbon pools in an age-sequence of temperate pine plantation forests
1
2006
... 在一定林龄范围内, 随林龄增加, 森林土壤通常表现为碳积累过程, 即土壤碳储量一般呈增加趋势(
Changes in soil carbon with stand age—An evaluation of a modelling method with empirical data
13
2004
... 全球森林碳储量达到861 Pg (1 Pg = 1015 g), 并以每年2.4 Pg的速度增长, 在缓解大气CO2浓度升高方面具有重要作用(
... ;
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... ;
... .在碳储量的分配方面, 随演替进行, 通常植被碳储量贡献增加, 而土壤碳储量贡献减小(
... 林龄是影响森林生态系统碳储量的重要因素(
... ).植被碳储量随演替进行而增加的现象在针叶林、温带和热带地区的天然林与人工林中普遍存在(
... 随林龄增长, 木质残体碳储量呈增加趋势, 这与我国森林整体(
... ;
... 有关凋落物随林龄的变化趋势, 不同研究的结果不尽相同.如全球200年以下温带森林(
... 在一定林龄范围内, 随林龄增加, 森林土壤通常表现为碳积累过程, 即土壤碳储量一般呈增加趋势(
... 本研究中, 不同演替阶段亮叶水青冈林土壤总碳储量无显著差异; 而0-10 cm土壤碳储量在33年林最高, 82年林最低.这种变化格局可能与研究样地的土地利用历史有关(
... 本研究中, 不同演替阶段亮叶水青冈林生态系统碳储量从33年林的186.9 Mg·hm-2增加到208年林的515.1 Mg·hm-2.贵州梵净山44-185年水青冈(F. longipetiolata和F. lucida)林的生态系统碳储量在190.7-503.9 Mg·hm-2之间(本课题组未发表数据); 德国Thuringia中部30-171年欧洲水青冈(F. sylvatica)林的碳储量为126-413 Mg·hm-2 (
Carbon cycling and storage in world forests: Biome patterns related to forest age
2
2004
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
云南哀牢山徐家坝地区木果石栎林生物量的初步研究
1
1984
... 而不同演替阶段亮叶水青冈林的灌木层与草本层碳储量皆无显著差异.有关林下生物量随林龄增长的变化趋势目前并无一致的结论.一些研究发现, 在幼龄林阶段, 林下生物量随林龄增长而降低; 而此后林龄进一步增长时(约40年以后), 林下层生物量基本无明显变化趋势, 如美国明尼苏达州北部7-160年Pinus resinosa林和6-133年杨树(Populus tremuloides和P. grandidentata)林(
云南哀牢山徐家坝地区木果石栎林生物量的初步研究
1
1984
... 而不同演替阶段亮叶水青冈林的灌木层与草本层碳储量皆无显著差异.有关林下生物量随林龄增长的变化趋势目前并无一致的结论.一些研究发现, 在幼龄林阶段, 林下生物量随林龄增长而降低; 而此后林龄进一步增长时(约40年以后), 林下层生物量基本无明显变化趋势, 如美国明尼苏达州北部7-160年Pinus resinosa林和6-133年杨树(Populus tremuloides和P. grandidentata)林(
Biomass and composition of understory vegetation and the forest floor carbon stock across Siberian larch and mountain birch chronosequences in Iceland
2
2005
... 随林龄增长, 木质残体碳储量呈增加趋势, 这与我国森林整体(
... ), 主要是因为调查的森林演替多起源于火烧等重大干扰, 因而幼龄林中残留有较多的木质残体(
Coarse woody debris in Douglas-fir forests of western Oregon and Washington
1
1988
... 随林龄增长, 木质残体碳储量呈增加趋势, 这与我国森林整体(
Coarse woody debris as a function of age, stand structure, and disturbance in boreal Newfoundland
1
1997
... 林龄是影响森林生态系统碳储量的重要因素(
南亚热带典型森林演替类型粗死木质残体贮量及其对碳循环的潜在影响
1
2005
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
南亚热带典型森林演替类型粗死木质残体贮量及其对碳循环的潜在影响
1
2005
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
神农架地区米心水青冈林和锐齿槲栎林生物量的研究
1
2007
... 亚热带森林在陆地生态系统碳循环中起着重要作用.有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (
神农架地区米心水青冈林和锐齿槲栎林生物量的研究
1
2007
... 亚热带森林在陆地生态系统碳循环中起着重要作用.有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (
A preliminary vegetation study of Fagus forests in central China: Species composition, structure and ecotypes
1
2003
... Allometric equations for calculating aboveground biomass (AGB) and belowground biomass (BGB) of dominant tree species in this study
| 物种 Species | 地上生物量 AGB (kg) | 地下生物量 BGB (kg) | 本文主要对应树种 Species in this study | 参考文献 References |
|---|---|---|---|---|
| 锥属 Castanopsis | AGB = 0.0177(D2H)1.0168 + 0.0364(D2H)0.6530 + 0.1533(D2H)0.2948 | BGB = 0.00911(D2H)0.933951 | 锥属 Castanopsis | Qiu et al., 1984 |
| 水青冈属 Fagus | AGB = 0.0125(D2H)1.05 + 0.000933(D 2H)1.23 + 0.000294(D 2H)1.20 | BGB = 0.00322(D2H)1.13 | 亮叶水青冈 Fagus lucida | |
| 樟科 Lauraceae | AGB = 0.055603(D2H)0.850193 + 0.014757(D2H)0.808395 + 0.006652(D2H)1.051841 + 0.059871(D2H)0.574327 | BGB = 0.184736(D2H)0.616421 | 木姜子属 Litsea | Yao et al., 2003 |
| 枫香 Liquidambar formosana | AGB = 0.174(D2H)0.7661 + 3 × 10-8(D2H)2 + 0.001(D2H) + 9.7883 + 0.0002(D2H)1.2696 + 0.0002D3.2304 + 3 × 10-7(D2H)1.5626 | BGB = 0.0094(D2H)0.9538 | 枫香树 Liquidambar formosana | Ming et al., 2012 |
| 柯属 Lithocarpus | AGB = 0.0347(D2H)0.9470 + 0.0084(D2H)0.9112 + 0.0072(D2H)0.6893 | BGB = 0.01534(D2H)0.95121 | 柯属 Lithocarpus | Qiu et al., 1984 |
| 木兰科 Magnoliaceae | AGB = 0.502921(D2H)0.56821 + 0.007183(D2H)0.92191 + 0.02252(D2H)0.62601 | BGB = 0.0364(D2H)0.79111 | 木兰属 Magnolia | Qiu et al., 1984 |
| 落叶阔叶树 Deciduous broad-leaved trees | AGB = 0.0650(D2H)0.84 + 1.59(D2H)0.38 + 0.218(D2H)0.34 | BGB = 0.291(D2H)0.55 | 鹅耳枥属 Carpinus, 槭属 Acer | Wang et al., 2007 |
| 常绿阔叶树 Evergreen broad-leaved trees | AGB = 0.17686(D2H)0.75995 + 0.11499(D2H)0.69997 + 0.107513(D2H)0.53231 | BGB = 0.095827(D2H)0.7165 | 青冈属 Cyclobalanopsis, 冬青属 Ilex | Deng et al., 2000 |
| 落叶小乔木 Small deciduous trees | AGB = 0.0434(D2H)0.91 + 0.000902(D2H)1.31 + 0.000790(D2H)1.05 | BGB = 0.000781(D2H)1.05 | 尖叶四照花 Dendrobenthamia angustata, 贵州桤叶树 Clethra kaipoensis | Wang et al., 2007 |
| 常绿小乔木 Small evergreen trees | AGB = 0.190(D2H)0.663 + 0.123(D2H)1.023 + 0.00728(D2H)0.548 | BGB = 0.0557(D2H)0.622 | 川桂 Cinnamomum wilsonii, 杜鹃属 Rhododendron | Wang et al., 2007 |
榕江月亮山植物群落的特征及多样性
1
2013
... 研究地点位于贵州省黔东南州榕江县月亮山省级自然保护区(108.22°-108.32° E, 25.57°-25.65° N).月亮山地处贵州省东南部, 位于华南褶皱带, 地质构造属新华夏体系, 基岩多为浅变质黏土岩类, 地貌类型为强切割侵蚀构造中低山.该区域属于亚热带山地季风气候, 年平均气温12.1-15.7 ℃, 年降水量1 030-1 688 mm.地带性植被为常绿阔叶林, 1 100 m以上主要为常绿落叶阔叶混交林, 土壤多为山地黄棕壤.月亮山低山地区受砍伐和放牧等人为活动干扰较大, 而中山常绿阔叶混交林保存相对完好.亮叶水青冈林是该区域常绿阔叶混交林中主要的森林类型(
榕江月亮山植物群落的特征及多样性
1
2013
... 研究地点位于贵州省黔东南州榕江县月亮山省级自然保护区(108.22°-108.32° E, 25.57°-25.65° N).月亮山地处贵州省东南部, 位于华南褶皱带, 地质构造属新华夏体系, 基岩多为浅变质黏土岩类, 地貌类型为强切割侵蚀构造中低山.该区域属于亚热带山地季风气候, 年平均气温12.1-15.7 ℃, 年降水量1 030-1 688 mm.地带性植被为常绿阔叶林, 1 100 m以上主要为常绿落叶阔叶混交林, 土壤多为山地黄棕壤.月亮山低山地区受砍伐和放牧等人为活动干扰较大, 而中山常绿阔叶混交林保存相对完好.亮叶水青冈林是该区域常绿阔叶混交林中主要的森林类型(
不同林龄桤-柏混交林生态系统的碳储量及其分配
1
2008
... 亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低.这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(
不同林龄桤-柏混交林生态系统的碳储量及其分配
1
2008
... 亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低.这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(
2
1994
... 亚热带森林在陆地生态系统碳循环中起着重要作用.有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (
... 研究地点位于贵州省黔东南州榕江县月亮山省级自然保护区(108.22°-108.32° E, 25.57°-25.65° N).月亮山地处贵州省东南部, 位于华南褶皱带, 地质构造属新华夏体系, 基岩多为浅变质黏土岩类, 地貌类型为强切割侵蚀构造中低山.该区域属于亚热带山地季风气候, 年平均气温12.1-15.7 ℃, 年降水量1 030-1 688 mm.地带性植被为常绿阔叶林, 1 100 m以上主要为常绿落叶阔叶混交林, 土壤多为山地黄棕壤.月亮山低山地区受砍伐和放牧等人为活动干扰较大, 而中山常绿阔叶混交林保存相对完好.亮叶水青冈林是该区域常绿阔叶混交林中主要的森林类型(
2
1994
... 亚热带森林在陆地生态系统碳循环中起着重要作用.有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (
... 研究地点位于贵州省黔东南州榕江县月亮山省级自然保护区(108.22°-108.32° E, 25.57°-25.65° N).月亮山地处贵州省东南部, 位于华南褶皱带, 地质构造属新华夏体系, 基岩多为浅变质黏土岩类, 地貌类型为强切割侵蚀构造中低山.该区域属于亚热带山地季风气候, 年平均气温12.1-15.7 ℃, 年降水量1 030-1 688 mm.地带性植被为常绿阔叶林, 1 100 m以上主要为常绿落叶阔叶混交林, 土壤多为山地黄棕壤.月亮山低山地区受砍伐和放牧等人为活动干扰较大, 而中山常绿阔叶混交林保存相对完好.亮叶水青冈林是该区域常绿阔叶混交林中主要的森林类型(
18年生樟树人工林生物量的结构与分布
1
2003
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
18年生樟树人工林生物量的结构与分布
1
2003
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
High carbon dioxide uptake by subtropical forest ecosystems in the East Asian monsoon region
1
2014
... 亚热带森林在陆地生态系统碳循环中起着重要作用.有研究表明, 仅东亚季风区亚热带森林(占全球森林面积的5%)的净生态系统生产力(NEP)就占全球森林NEP的8% (
6种温带森林碳密度与碳分配
1
2010
... 全球森林碳储量达到861 Pg (1 Pg = 1015 g), 并以每年2.4 Pg的速度增长, 在缓解大气CO2浓度升高方面具有重要作用(
6种温带森林碳密度与碳分配
1
2010
... 全球森林碳储量达到861 Pg (1 Pg = 1015 g), 并以每年2.4 Pg的速度增长, 在缓解大气CO2浓度升高方面具有重要作用(
贵州月亮山5种森林类型土壤生态化学计量特征研究
1
2016
... 研究地点位于贵州省黔东南州榕江县月亮山省级自然保护区(108.22°-108.32° E, 25.57°-25.65° N).月亮山地处贵州省东南部, 位于华南褶皱带, 地质构造属新华夏体系, 基岩多为浅变质黏土岩类, 地貌类型为强切割侵蚀构造中低山.该区域属于亚热带山地季风气候, 年平均气温12.1-15.7 ℃, 年降水量1 030-1 688 mm.地带性植被为常绿阔叶林, 1 100 m以上主要为常绿落叶阔叶混交林, 土壤多为山地黄棕壤.月亮山低山地区受砍伐和放牧等人为活动干扰较大, 而中山常绿阔叶混交林保存相对完好.亮叶水青冈林是该区域常绿阔叶混交林中主要的森林类型(
贵州月亮山5种森林类型土壤生态化学计量特征研究
1
2016
... 研究地点位于贵州省黔东南州榕江县月亮山省级自然保护区(108.22°-108.32° E, 25.57°-25.65° N).月亮山地处贵州省东南部, 位于华南褶皱带, 地质构造属新华夏体系, 基岩多为浅变质黏土岩类, 地貌类型为强切割侵蚀构造中低山.该区域属于亚热带山地季风气候, 年平均气温12.1-15.7 ℃, 年降水量1 030-1 688 mm.地带性植被为常绿阔叶林, 1 100 m以上主要为常绿落叶阔叶混交林, 土壤多为山地黄棕壤.月亮山低山地区受砍伐和放牧等人为活动干扰较大, 而中山常绿阔叶混交林保存相对完好.亮叶水青冈林是该区域常绿阔叶混交林中主要的森林类型(
A significant carbon sink in temperate forests in Beijing: Based on 20-year field measurements in three stands
2
2015
... 测量样方内所有胸径≥3 cm活立木的胸径和树高, 并记录物种名.利用相关生长方程计算每株树的生物量, 其中乔木层地上生物量为乔木茎、枝和叶生物量的总和, 乔木层地下生物量为乔木根生物量.由于保护区内禁止砍伐, 无法对样地内树种进行生物量方程构建, 故通过文献检索, 利用邻近地区或气候条件相近地区对应树种的生物量方程(
... 本研究调查了贵州榕江县月亮山不同演替阶段的9个亮叶水青冈林生态系统的全组分碳储量及各组分的相对贡献.可能会给结果带来不确定性的因素主要有两方面: 一是由于保护区禁止采伐树木, 本研究在估算乔木生物量时利用的是文献中已有的生物量方程(
Carbon stocks and changes of dead organic matter in China’s forests
7
2017a
... 全球森林碳储量达到861 Pg (1 Pg = 1015 g), 并以每年2.4 Pg的速度增长, 在缓解大气CO2浓度升高方面具有重要作用(
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... 对样方内所有FWD进行收集并称质量.在样方内随机设置3个1 m × 1 m小样方, 收集小样方内所有凋落物(包括落叶、树皮、果实、直径< 2 cm的枯枝以及地表其他植物残体)并称质量.每个600 m2样方内, FWD与凋落物各取3份样品, 带回实验室烘干(65 ℃)至恒质量后测定含水率, 并换算其干质量, 研磨过0.15 mm筛后测定碳含量(
... 随林龄增长, 木质残体碳储量呈增加趋势, 这与我国森林整体(
... 有关凋落物随林龄的变化趋势, 不同研究的结果不尽相同.如全球200年以下温带森林(
... 亮叶水青冈林生态系统总碳储量随林龄的增长主要由植被碳储量贡献, 故随林龄增长, 亮叶水青冈林植被碳储量比重增加, 而土壤碳储量比重降低.这与国内外许多亚热带和温带森林的研究结果一致, 如广西格木林(
... 本研究调查了贵州榕江县月亮山不同演替阶段的9个亮叶水青冈林生态系统的全组分碳储量及各组分的相对贡献.可能会给结果带来不确定性的因素主要有两方面: 一是由于保护区禁止采伐树木, 本研究在估算乔木生物量时利用的是文献中已有的生物量方程(
Plant debris and its contribution to ecosystem carbon storage in successional Larix gmelinii forests in northeastern China
10
2017b
... 已有不少研究探讨不同林龄森林的碳储量及其分配格局的变化.研究对象多为局域某一类型森林的林龄序列, 如大兴安岭15-138年落叶松(Larix gmelinii)林(
... ;
... ;
... 将木质残体区分为细木质残体(大头直径2-10 cm; FWD)和粗木质残体(大头直径≥10 cm; CWD) (
... 对样方内所有FWD进行收集并称质量.在样方内随机设置3个1 m × 1 m小样方, 收集小样方内所有凋落物(包括落叶、树皮、果实、直径< 2 cm的枯枝以及地表其他植物残体)并称质量.每个600 m2样方内, FWD与凋落物各取3份样品, 带回实验室烘干(65 ℃)至恒质量后测定含水率, 并换算其干质量, 研磨过0.15 mm筛后测定碳含量(
... 林龄是影响森林生态系统碳储量的重要因素(
... 随林龄增长, 木质残体碳储量呈增加趋势, 这与我国森林整体(
... ;
... 有关凋落物随林龄的变化趋势, 不同研究的结果不尽相同.如全球200年以下温带森林(
... 在一定林龄范围内, 随林龄增加, 森林土壤通常表现为碳积累过程, 即土壤碳储量一般呈增加趋势(
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}