Drivers of composition and density pattern of tree seedlings in a secondary mixed conifer and broad-leaved forest, Jiaohe, Jilin, China
MENG Ling-Jun, YAO Jie, QIN Jiang-Huan, FAN Chun-Yu, ZHANG Chun-Yu, ZHAO Xiu-Hai,*Research Center of Forest Management Engineering of State Forestry and Grassland Administration, Beijing Forestry University, Beijing 100083, China
Supported by the Key Project of National Key Research and Development Plan.2017YFC0504005 the Program of National Natural Science Foundation of China.31670643
Abstract Aims Our aim is to explore the composition and temporal dynamics of tree seedlings and to further understand the impacts and relative importance of biotic neighborhood and habitat heterogeneity on seedling density patterns in a secondary mixed conifer and broad-leaved forest in Jiaohe, Jilin Province, North-eastern China.Methods We established 209 seedling census stations in the mixed conifer and broad-leaved forest. Based on two seedling censuses conducted in 2016 and 2017, we analyzed species composition and quantitative dynamics of seedlings. Generalized mixed linear models were used to test the relative effects of biotic neighborhood and habitat heterogeneity on seedling density patterns.Important findings (1) A total of 4 245 seedlings were investigated in all seedling census stations of this plot, belonging to 12 genera, 10 families and 18 species. The number of new-born seedlings varied greatly among species and census years. Fraxinus mandschurica and Tilia amurensis seedlings occurred and died in large numbers. (2) At the community level, the sum of conspecific adult basal area, the soil total phosphorus and available potassium exhibited significant positive effects on seedling density. The habitat suitable for trees was also suitable for seedlings. (3) The factors influencing seedling density of Fraxinus mandschurica were consistent with those of seedling density at the community level. Pinus koraiensis had significant habitat preference, it had higher density in the habitat with lower moisture, soil total phosphorus, available nitrogen, and available phosphorus. Conspecific adult neighbor effects on annual seedling density was more significant than on perennial seedling density. This study suggests that dispersal limitation and habitat filtering together affect seedling density pattern, and the relative importance of biotic neighbors and habitat heterogeneity varies among seedling species and age classes. Keywords:secondary mixed conifer and broad-leaved forest;generalized linear mixed-effect model;tree seedling density;biotic neighbor;habitat heterogeneity
PDF (1102KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 孟令君, 姚杰, 秦江环, 范春雨, 张春雨, 赵秀海. 吉林蛟河针阔混交林乔木幼苗组成及其密度格局影响因素. 植物生态学报[J], 2018, 42(6): 653-662 doi:10.17521/cjpe.2018.0025 MENG Ling-Jun. Drivers of composition and density pattern of tree seedlings in a secondary mixed conifer and broad-leaved forest, Jiaohe, Jilin, China. Chinese Journal of Plant Ecology[J], 2018, 42(6): 653-662 doi:10.17521/cjpe.2018.0025
Table 4 表4 表4个体水平幼苗密度模型的赤池信息量准则(AIC)和ΔAIC值 Table 4Akaike’s information criteria (AIC) and ΔAIC values of the models for individual-level seedling density
Fig. 4Estimates of the coefficients (± 2 × SE) of the best models for seedling density over two years at the community level. Solid symbols indicate parameters significantly different from zero (p < 0.05), and hollow symbols indicate no significant difference from zero (p > 0.05). See Table 3 for variable abbreviations.
Fig. 5Estimates of the coefficients (± 2 × SE) of the best models for seedling density at different age classes. Solid symbols indicate parameters significantly different from zero (p < 0.05), and hollow symbols indicate no significant difference from zero (p > 0.05). See Table 3 for variable abbreviations.
Fig. 6Estimates of the coefficients (± 2 × SE) of the best models for the seedling density of different species. Solid symbols indicate parameters significantly different from zero (p < 0.05), and hollow symbols indicate no significant difference from zero (p > 0.05). See Table 3 for variable abbreviations.
BaiX, QueenboroughSA, WangX, ZhangJ, LiB, YuanZ, XingD, LinF, YeJ, HaoZ ( 2012). Effects of local biotic neighbors and habitat heterogeneity on tree and shrub seedling survival in an old-growth temperate forest Oecologia, 170, 755-765. DOI:10.1007/s00442-012-2348-2URL [本文引用: 6]
BinY, YeWH, CaoHL, HuangZL, LianJY ( 2011). Seedlings distribution in a subtropical evergreen broad-leaved forest plot in the Dinghu Mountain Biodiversity Science, 19, 127-133. [本文引用: 1]
BurnhamKP, AndersonDR ( 2002). Model Selection and Inference: A Practical Information-Theoretic Approach. Springer Science & Business Media New York. [本文引用: 1]
CanhamCD, LePagePT, CoatesKD ( 2004). A neighborhood analysis of canopy tree competition: Effects of shading versus crowding Canadian Journal of Forest Research, 34, 778-787. DOI:10.1139/x03-232URL [本文引用: 1]
ChenL, MiX, ComitaLS, ZhangL, RenH ( 2010). Community-?level consequences of density dependence and habitat association in a subtropical broad-leaved forest Ecology Letters, 13, 695-704. DOI:10.1111/j.1461-0248.2010.01468.xURL [本文引用: 2]
ComitaLS, HubbellSP ( 2009). Local neighborhood and species’ shade tolerance influence survival in a diverse seedling bank Ecology, 90, 328-334. DOI:10.1890/08-0451.1URL [本文引用: 2]
ComitaLS, QueenboroughSA, MurphySJ, EckJL, XuK, KrishnadasM, BeckmanN, ZhuY ( 2014). Testing predictions of the Janzen-Connell hypothesis: A meta-analysis of experimental evidence for distance- and density-dependent seed and seedling survival Journal of Ecology, 102, 845-856. DOI:10.1111/1365-2745.12232URL [本文引用: 2]
ConnellJH ( 1971). On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In: den Boer PJ, Gradwell GR eds. Dynamics of Population Center for Agricultural Publishing and Documentation, Wageningen, The Netherlands.289-312. [本文引用: 1]
ConnellJH, GreenPT ( 2000). Seedling dynamics over thirty-two years in a tropical rain forest tree Ecology, 568-584. DOI:10.2307/177449URL [本文引用: 1] We examined patterns of seedling recruitment and their underlying mechanisms in a population of Chrysophyllum sp. nov. (Sapotaceae), a shade-tolerant canopy species in mature tropical rainforest in Queensland, Australia. We considered spatial scales ranging from 1 m2to 1 ha, and temporal scales ranging from 1 to 32 yr. Over the 32-yr study period there were six episodes of very high seedling recruitment (masts) at intervals ranging from 4 to 10 yr. Less than 2% of new recruits were found in the nine censuses in other years. We found no significant correlations between the numbers of seedlings per census and either seasonal or annual rainfall, number of dry months per year, or El Nino Southern Oscillation (ENSO) events but found two correlations with mean seasonal air temperatures in the years preceding flowering. There were long-term changes in abundance in both time and space. In 1965 there were 163 seedling-sized plants in two dense patches, whereas in 1996, there were 4000 in 15 patches. Once a new patch became established, seedlings recruited there in each succeeding mast episode. Adult trees varied in their production of seedlings. Only 25% of the trees in the sample analyzed participated in all mast years; others had few seedlings for up to 28 yr before beginning to produce many seedlings. Seedlings were shade tolerant. They grew extremely slowly in the shaded understory, mean height only doubling in 27 yr. They were also quite long-lived; 6% of seedlings recruited in 1969 were still alive 27 yr later. There was little effect of natural enemies on seeds or seedlings. We found weak positive and negative effects of density on seed germination, seedling mortality, and growth. Mortality rates of Chrysophyllum seedlings did not show any trends with time, nor with distance from conspecific adults. These results suggest that abundance of older stages is determined by recruitment as well as subsequent growth and mortality. Mortality rates of seedlings of species other than Chrysophyllum decreased with distance from adult Chrysophyllum trees. Under present conditions, the Chrysophyllum population may be increasing in relation to that of other species, perhaps leading to a reduction in tree diversity in this tropical rainforest.
FanHB, ZangRG, LiDZ ( 1996). Natural regeneration of Mongolian oak population Chinese Journal of Ecology, 15(3), 15-20. [本文引用: 1]
JanzenDH ( 1970). Herbivores and the number of tree species in tropical forests The American Society of Naturalists, 104, 501-528. DOI:10.1086/282687URL [本文引用: 1] A high number of tree species, low density of adults of each species, and long distances between conspecific adults are characteristic of many low-land tropical forest habitats. I propose that these three traits, in large part, are the result of the action of predators on seeds and seedlings. A model is presented that allows detailed examination of the effect of different predators, dispersal agents, seed-crop sizes, etc. on these three traits. In short, any event that increases the efficiency of the predators at eating seeds and seedlings of a given tree species may lead to a reduction in population density of the adults of that species and/or to increased distance between new adults and their parents. Either event will lead to more space in the habitat for other species of trees, and therefore higher total number of tree species, provided seed sources are available over evolutionary time. As one moves from the wet lowland tropics to the dry tropics or temperate zones, the seed and seedling predators in a habitat are hypothesized to be progressively less efficient at keeping one or a few tree species from monopolizing the habitat through competitive superiority. This lowered efficiency of the predators is brought about by the increased severity and unpredictability of the physical environment, which in turn leads to regular or erratic escape of large seed or seedling cohorts from the predators.
JohnsonDJ, BeaulieuWT, BeverJD, ClayK ( 2012). Conspecific negative density dependence and forest diversity Science, 336, 904-907. DOI:10.1126/science.1220269URL [本文引用: 2]
JohnsonDJ, BourgNA, HoweR, McSheaWJ, WolfA, ClayK ( 2014). Conspecific negative density-dependent mortality and the structure of temperate forests Ecology, 95, 2493-2503. DOI:10.1890/13-2098.1URL [本文引用: 1]
KammesheidtL ( 1998). The role of tree sprouts in the restoration of stand structure and species diversity in tropical moist forest after slash-and-burn agriculture in Eastern Paraguay Plant Ecology, 139, 155-165. DOI:10.1023/A:1009763402998URL [本文引用: 1]
KitajimaK, AugspurgerCK ( 1989). Seed and seedling ecology of a monocarpic tropical tree Ecology, 70, 1102-1114. DOI:10.2307/1941379URL [本文引用: 1] Tachigalia versicolor is a monocarpic canopy tree species on Barro Colorado Island, Panama, This 2-yr demographic and physiological study examined whether seed and seedling survival of this monocarpic species is high compared to nonmonocarpic species previously studied, as life history theory predicts. For the weighted average of two trees, the percentage of offspring estimated to survive from dispersed seed to germination, 1 yr, and 2 yr was 30, 7.2, and 4.3%, respectively. The large seeds (500-600 mg) suffered greater mortality in absolute numbers than did the seedlings. Major mortality agents of seeds were bruchid beetles (Amblycerus tachygaliae) and terrestrial vertebrates, while vertebrate herbivores and damping-off fungi killed seedlings. Seedlings survival and growth were enhanced under canopies of dying T. versicolor adults, relative to under canopies of living conspecific and nonconspecific adults. The majority of 2-yr-old seedlings occurred within 0.40 m from the parent trees, reflecting the original peak of the distribution of wind-dispersed seeds. Few seedlings persisted beyond 40 m. Seedling survival in the first 2 mo was higher in shaded understory than in light-gaps, due to higher vertebrate herbivory in sun than shade. Seedling growth and survival beyond 2 mo was higher in light-gaps than in shaded understory. T. versicolor seedlings exhibited a wide range in maximum net photosynthetic rate when grown in contrasting light conditions, and were able to acclimate to a sudden increase in light level by means of morphological and physiological adjustments of newly developed leaves. These results indicate that T. versicolor can utilize a wide range of light environments in the forest during seedling establishment. Relative to nomonocarpic wind-dispersed tree species previously studied in the same community. T. versicolor ranks near the top in its seed mass, dispersal capability, photosynthetic flexibility to contrasting light conditions, and probability of seedling survival in the shaded understory through 1 yr. These characteristics enable T. versicolor to establish large numbers of seedlings. The mean total number of offspring of two trees estimated to survive through 2 yr was 4395 out of 103 055 dispersed seeds and 30 529 germinated seeds. In comparison to nonmonocarpic species, juvenile survival of T. versicolor is high as expected for this monocarpic tree species.
LiSF, LiuWD, SuJR, ZhangZJ, LiuQY ( 2008). Woody seedling regeneration in secondary succession of monsoon broad-leaved evergreen forest in Puer, Yunnan, Southwest China Acta Ecologica Sinica, 18, 5653-5662. [本文引用: 1]
LiSG, BaiB, LuCH ( 2010). Seed rain and seed bank of Chinese yew ( Taxus chinensis var. mairei) population in Tianmu Mountain. Acta Ecologica Sinica, 30, 276-279. [本文引用: 1]
LiXL, WangH, ZhengZ, LinLX, DengXB, CaoM ( 2009). Composition, spatial distribution and survival during the dry season of tree seedlings in a tropical forest in Xishuang-Banna, SW China Chinese Journal of Plant Ecology, 33, 658-671. [本文引用: 1]
LinF, ComitaLS, WangXG, BaiXJ, YuanZQ, XingDL, HaoZQ ( 2014). The contribution of understory light availability and biotic neighborhood to seedling survival in secondary versus old-growth temperate forest Plant Ecology, 215, 795-807. DOI:10.1007/s11258-014-0332-0URL [本文引用: 1]
LiuS, JinGZ ( 2008). Spatiotemporal dynamics of seed rain in a broadleaved-Korean pine mixed forest in Xiaoxing’an Mountains, China Acta Ecologica Sinica, 28, 5731-5740. [本文引用: 1]
LiuS, LiaoJX, XiaoC, FanXH ( 2016). Effects of biotic neighbors and habitat heterogeneity on tree seedling survival in a secondary mixed conifer and broad-leaved forest in Changbai Mountain Chinese Journal of Plant Ecology, 40, 711-722. DOI:10.17521/cjpe.2015.0366URL [本文引用: 4] 基于长白山次生针阔混交林样地,以520个1 m×1 m幼苗样方中胸径小于1 cm的乔木幼苗为研究对象,选取2013年和2014年的幼苗调查数据,运用广义线性混合模型(GLMM)分析了生物邻体和生境异质性对幼苗存活的影响,探讨了次生针阔混交林幼苗存活影响因素及物种共存机制。结果表明:(1)适宜大树生长的局域生境同样也适宜幼苗的存活,幼苗存活率与土壤含水量和有效氮等土壤养分显著正相关。(2)幼苗个体之间存在明显的竞争,较多的幼苗邻体显著降低幼苗的存活率。同种大树邻体和同种幼苗邻体与幼苗存活显著负相关,表明存在负密度制约效应。(3)随着幼苗年龄的增加,生境异质性对幼苗存活的影响逐渐增大。该研究证实了密度制约效应和生境异质性对幼苗存活有着重要影响,其相对重要性随着幼苗年龄级、功能群以及物种种类而变化。 [ 刘帅, 廖嘉星, 肖翠, 范秀华 ( 2016). 长白山次生针阔混交林乔木幼苗存活的影响因素分析 植物生态学报, 40, 711-722.] DOI:10.17521/cjpe.2015.0366URL [本文引用: 4] 基于长白山次生针阔混交林样地,以520个1 m×1 m幼苗样方中胸径小于1 cm的乔木幼苗为研究对象,选取2013年和2014年的幼苗调查数据,运用广义线性混合模型(GLMM)分析了生物邻体和生境异质性对幼苗存活的影响,探讨了次生针阔混交林幼苗存活影响因素及物种共存机制。结果表明:(1)适宜大树生长的局域生境同样也适宜幼苗的存活,幼苗存活率与土壤含水量和有效氮等土壤养分显著正相关。(2)幼苗个体之间存在明显的竞争,较多的幼苗邻体显著降低幼苗的存活率。同种大树邻体和同种幼苗邻体与幼苗存活显著负相关,表明存在负密度制约效应。(3)随着幼苗年龄的增加,生境异质性对幼苗存活的影响逐渐增大。该研究证实了密度制约效应和生境异质性对幼苗存活有着重要影响,其相对重要性随着幼苗年龄级、功能群以及物种种类而变化。
LuoYQ, YuMS, YuJJ, ZhengSL, LiuJJ, YuMJ ( 2017). Effects of plant traits and the relative abundance of common woody species on seedling herbivory in the Thousand Island Lake region Chinese Journal of Plant Ecology, 41, 1033-1040. [本文引用: 1]
MakanaJ, ThomasSC ( 2004). Dispersal limits natural recruitment of African mahoganies Oikos, 106, 67-72. DOI:10.1111/j.0030-1299.2004.13084.xURL [本文引用: 3] The abundance of forest tree species may be locally limited by the inability of species to disperse to all sites suitable for germination and establishment. This phenomenon of "dispersal limitation" has been suggested to promote the maintenance of high species diversity in tropical forests by slowing down competitive exclusion. We present the first direct experimental evaluation of dispersal limitation in tropical forests, and of its importance relative to other factors affecting recruitment. Seed addition, litter removal and light availability effects on seedling establishment were evaluated in a full factorial experiment on two African mahogany species, Entandrophragma utile and Khaya anthotheca . Seedling recruitment after 18 months, was 22 times greater in seed addition treatments (with addition rates corresponding to the tail of seed shadows of large reproductive trees) compared to controls. This seed addition effect was an order of magnitude greater than either gap or litter removal effects, with similar results observed in both logged and primary forest locations. We conclude that dispersal limitation strongly limits seedling establishment for the two species studied. An important "applied corollary" of this result is that seed supplementation may substantially increase local abundance of valuable or rare species in disturbed or managed tropical forests.
MasakiT, NakashizukaT ( 2002). Seedling demography of Swida controversa: Effect of light and distance to conspecifics. Ecology, 83, 3497-3507. [本文引用: 1]
MontgomeryRA, ChazdonRL ( 2002). Light gradient partitioning by tropical tree seedlings in the absence of canopy gaps Oecologia, 131, 165-174. DOI:10.1007/s00442-002-0872-1URL [本文引用: 3]
NordenN, ChazdonRL, ChaoA, JiangYH, Vílchez-AlvaradoB ( 2009). Resilience of tropical rain forests: Tree community reassembly in secondary forests Ecology Letters, 12, 385-394. DOI:10.1111/j.1461-0248.2009.01292.xURLPMID:19379133 [本文引用: 1] Understanding the recovery dynamics of ecosystems presents a major challenge in the human-impacted tropics. We tested whether secondary forests follow equilibrium or non-equilibrium dynamics by evaluating community reassembly over time, across different successional stages, and among multiple life stages. Based on long-term and static data from six 1-ha plots in NE Costa Rica, we show that secondary forests are undergoing reassembly of canopy tree and palm species composition through the successful recruitment of seedlings, saplings, and young trees of mature forest species. Such patterns were observed over time within sites and across successional stages. Floristic reassembly in secondary forests showed a clear convergence with mature forest community composition, supporting an equilibrium model. This resilience stems from three key factors co-occurring locally: high abundance of generalist species in the regional flora, high levels of seed dispersal, and local presence of old-growth forest remnants.
NorghauerJM, NewberyDM ( 2014). Herbivores differentially limit the seedling growth and sapling recruitment of two dominant rain forest trees Oecologia, 174, 459-469. DOI:10.1007/s00442-013-2769-6URL [本文引用: 1]
OshimaC, TokumotoY, NakagawaM ( 2015). Biotic and abiotic drivers of dipterocarp seedling survival following mast fruiting in Malaysian Borneo Journal of Tropical Ecology, 31, 129-137. DOI:10.1017/S026646741400073XURL [本文引用: 2]
PaineCT, HarmsKE ( 2009). Quantifying the effects of seed arrival and environmental conditions on tropical seedling community structure Oecologia, 160, 139-150. DOI:10.1007/s00442-008-1269-6URL [本文引用: 1]
PasquiniSC, SantiagoLS ( 2012). Nutrients limit photosynthesis in seedlings of a lowland tropical forest tree species Oecologia, 168, 311-319 DOI:10.1007/s00442-011-2099-5URLPMID:21837408 [本文引用: 2] We investigated how photosynthesis by understory seedlings of the lowland tropical tree species Alseis blackiana responded to 10 years of soil nutrient fertilization with N, P and K. We ask whether nutrients are limiting to light and CO2 acquisition in a low light understory environment.We measured foliar nutrient concentrations of N, P and K, isotopic composition of carbon (δ0161 C) and nitrogen (δ0163 N), and light response curves of photosynthesis and chlorophyll fluorescence. Canopy openness was measured above each study seedling and included in statistical analyses to account for variation in light availability. Foliar N concentration increased by 20% with N addition. Foliar P concentration increased by 78% with P addition and decreased by 14% with N addition. Foliar K increased by 8% with addition. Foliar δ 0161 C showed no significant responses, and foliar 07 I5 N decreased strongly with N addition, matching the low < 5 15 N values of applied fertilizer. Canopy openness ranged from 0.01 to 6.71% with a mean of 1.76 ± 0.14 (± 1SE). Maximum photosynthetic CO 2 assimilation rate increased by 9% with N addition. Stomatal conductance increased with N addition and with 05 and in combination. Chlorophyll fluorescence measurements revealed that quantum yield of photosystem II increased with K addition, maximum electron transport rate trended 9% greater with N addition (p = 0.07), and saturating photosynthetically active radiation increased with N addition. The results demonstrate that nutrient addition can enhance photosynthetic processes, even under low light availability.
PoorterL, RoseSA ( 2005). Light-dependent changes in the relationship between seed mass and seedling traits: A meta-analysis for rain forest tree species Oecologia, 142, 378-387. DOI:10.1007/s00442-004-1732-yURL [本文引用: 1]
QianH ( 2009). Beta diversity in relation to dispersal ability for vascular plants in North America Global Ecology and Biogeography, 18, 327-332. DOI:10.1111/geb.2009.18.issue-3URL [本文引用: 1]
RecordS, KobeRK, VriesendorpCF, FinleyAO ( 2016). Seedling survival responses to conspecific density, soil nutrients, and irradiance vary with age in a tropical forest Ecology, 97, 2406-2415. DOI:10.1002/ecy.1458URL [本文引用: 1]
ShiLL, LuoZR, XiaJT, ZhaoWJ, WuYG, DingBY ( 2014). Woody seedling dynamics and the correlation between habitat and regeneration/mortality in a subtropical evergreen broad-leaved forest in China Acta Ecologica Sinica, 34, 6510-6518. DOI:10.5846/stxb201302150268URL [本文引用: 4] 以百山祖自然保护区5 hm2永久样地150个幼苗监测站木本植物幼苗为研究对象,2008-2011年定期调查样方中幼苗的种类、数量、萌发、死亡等,探究亚热带中山常绿阔叶 林幼苗种类组成、数量动态及其与生境的相关性.结果表明:1)百山祖样地在2008年至2011年出现的幼苗属于26科,40属,共53个物种,不同物种 萌发时段有异;2)2009年样地幼苗存活比率为7.7%,2010年为-20.8%,2011年则是-0.3%,幼苗存活比率不高,种类和数量呈减少趋 势;3)存活幼苗中有明显的优势物种,分别为光亮山矾(Symplocos lucida)、尖连蕊茶(Camellia cuspidata)、浙闽新木姜子(Neolitsea aurata var.undulatula)、尖叶菝葜(Smilax arisanensis)和短尾柯(Lithocarpus brevicaudatus),5个物种之和占幼苗总数比例>50%;4)种子的萌发与生境有极显著的相关性,且与生境因子中水分关联最大;5)存活幼苗 数与样站坡位、水分、落叶层厚度呈现显著相关性,水和光照是影响幼苗存活的主要因素. [ 施璐璐, 骆争荣, 夏家天, 赵文娟, 吴友贵, 丁炳扬 ( 2014). 亚热带中山常绿阔叶林木本植物幼苗数量动态及其与生境的相关性 生态学报, 2014,34, 6510-6518.] DOI:10.5846/stxb201302150268URL [本文引用: 4] 以百山祖自然保护区5 hm2永久样地150个幼苗监测站木本植物幼苗为研究对象,2008-2011年定期调查样方中幼苗的种类、数量、萌发、死亡等,探究亚热带中山常绿阔叶 林幼苗种类组成、数量动态及其与生境的相关性.结果表明:1)百山祖样地在2008年至2011年出现的幼苗属于26科,40属,共53个物种,不同物种 萌发时段有异;2)2009年样地幼苗存活比率为7.7%,2010年为-20.8%,2011年则是-0.3%,幼苗存活比率不高,种类和数量呈减少趋 势;3)存活幼苗中有明显的优势物种,分别为光亮山矾(Symplocos lucida)、尖连蕊茶(Camellia cuspidata)、浙闽新木姜子(Neolitsea aurata var.undulatula)、尖叶菝葜(Smilax arisanensis)和短尾柯(Lithocarpus brevicaudatus),5个物种之和占幼苗总数比例>50%;4)种子的萌发与生境有极显著的相关性,且与生境因子中水分关联最大;5)存活幼苗 数与样站坡位、水分、落叶层厚度呈现显著相关性,水和光照是影响幼苗存活的主要因素.
SongB, St?cklinJ, GaoYQ, ZhangZQ, YangY, LiZM, SunH ( 2013). Habitat-specific responses of seed germination and seedling establishment to soil water condition in two Rheum species in the high Sino-Himalayas. Ecological research, 28, 643-651. [本文引用: 1]
SuY, JiaoJY, WangZJ ( 2014). Characteristics of seedling survival in habitats of hill and gully slopes in hill-gully Loess Plateau region of northern Shaanxi Chinese Journal of Plant Ecology, 38, 694-709. DOI:10.3724/SP.J.1258.2014.00065URL [本文引用: 1] 幼苗是植被恢复演替的基础,在生态系统中具有非常重要的作用。为了探索黄土丘陵沟壑区坡沟立地环境下幼苗的存活特征,在陕西省安塞县宋家沟和纸坊沟流域设置15个固定样地进行定点动态监测,运用单因素方差分析法和系统聚类分析法对不同立地环境下的幼苗密度和幼苗存活率进行了分析。结果表明:1)不同坡沟立地环境下,幼苗存活特征差异明显,整体表现为阳坡幼苗密度较小而存活率较高,阴坡幼苗密度较大但存活率较低。2)细裂叶莲蒿(Artemisia gmelinii)、菊叶委陵菜(Potentilla tanacetifolia)、阿尔泰狗娃花(Heteropappus altaicus)等物种具有较高的幼苗存活率,在生存竞争中幼苗是以"质"取胜;而猪毛蒿(Artemisia scoparia)、中华苦荬菜(Ixeris chinensis)、阴行草(Siphonostegia chinensis)等物种具有较高的幼苗密度,在生存竞争中是以"量"取胜。3)同一物种在坡沟立地环境下表现出不同的存活曲线,不同物种在不同立地环境下也可呈现同一存活曲线;细裂叶莲蒿、菊叶委陵菜、长芒草(Stipa bungeana)、阿尔泰狗娃花、兴安胡枝子(Lespedeza davurica)等物种,幼苗不仅在雨季存活能力强且具有较强的越冬能力;猪毛蒿、香青兰(Dracocephalum moldavica)、臭蒿(Artemisia hedinii)等物种在雨季前幼苗存活能力较强,但冬季存活能力弱。4)坡面不同微环境下幼苗存活特征也存在差异,即微环境的改变在一定程度上影响着幼苗的生长、存活和越冬。因此,在黄土丘陵沟壑区应根据物种幼苗存活特性对幼苗进行适当的抚育管理,这将有利于幼苗的建植和植被更新。 [ 苏嫄, 焦菊英, 王志杰 ( 2014). 陕北黄土丘陵沟壑区坡沟立地环境下幼苗的存活特征 植物生态学报, 38, 694-709.] DOI:10.3724/SP.J.1258.2014.00065URL [本文引用: 1] 幼苗是植被恢复演替的基础,在生态系统中具有非常重要的作用。为了探索黄土丘陵沟壑区坡沟立地环境下幼苗的存活特征,在陕西省安塞县宋家沟和纸坊沟流域设置15个固定样地进行定点动态监测,运用单因素方差分析法和系统聚类分析法对不同立地环境下的幼苗密度和幼苗存活率进行了分析。结果表明:1)不同坡沟立地环境下,幼苗存活特征差异明显,整体表现为阳坡幼苗密度较小而存活率较高,阴坡幼苗密度较大但存活率较低。2)细裂叶莲蒿(Artemisia gmelinii)、菊叶委陵菜(Potentilla tanacetifolia)、阿尔泰狗娃花(Heteropappus altaicus)等物种具有较高的幼苗存活率,在生存竞争中幼苗是以"质"取胜;而猪毛蒿(Artemisia scoparia)、中华苦荬菜(Ixeris chinensis)、阴行草(Siphonostegia chinensis)等物种具有较高的幼苗密度,在生存竞争中是以"量"取胜。3)同一物种在坡沟立地环境下表现出不同的存活曲线,不同物种在不同立地环境下也可呈现同一存活曲线;细裂叶莲蒿、菊叶委陵菜、长芒草(Stipa bungeana)、阿尔泰狗娃花、兴安胡枝子(Lespedeza davurica)等物种,幼苗不仅在雨季存活能力强且具有较强的越冬能力;猪毛蒿、香青兰(Dracocephalum moldavica)、臭蒿(Artemisia hedinii)等物种在雨季前幼苗存活能力较强,但冬季存活能力弱。4)坡面不同微环境下幼苗存活特征也存在差异,即微环境的改变在一定程度上影响着幼苗的生长、存活和越冬。因此,在黄土丘陵沟壑区应根据物种幼苗存活特性对幼苗进行适当的抚育管理,这将有利于幼苗的建植和植被更新。
WangCH, LiJQ, ChenFQ, YangY ( 2011). Factors affecting seedling regeneration of Liquidambar formosana in the L. formosana forests in hilly regions of Southeast Hubei, China. Chinese Journal of Plant Ecology, 35, 187-194. [本文引用: 2]
WangX, WiegandT, HaoZ, LiB, YeJ, LinF ( 2010). Species associations in an old-growth temperate forest in north-eastern China Journal of Ecology, 98, 674-686. DOI:10.1111/j.1365-2745.2010.01644.xURL [本文引用: 1] 1. Studying the spatial pattern of plants may provide significant insights into processes and mechanisms that maintain species richness. We used data from a fully mapped 25-ha temperate forest plot at Changbaishan (CBS), north-eastern China, to conduct a community-wide assessment of the type and frequency of intra- and interspecific spatial association patterns. We analysed complex scale effects in the patterning of large trees of 15 common species. First, we tested for overall spatial patterning at 6, 30 and 50 m neighbourhoods and classified the types of bivariate association patterns at these spatial scales (analysis 1). We then explored small-scale (0 20 m) association patterns conditioning on the larger-scale pattern (analysis 2) and tested for positive large-scale (50 250 m) association patterns (analysis 3).2. Analysis 1 provided ample evidence for non-random spatial patterning, and the type and frequency of spatial association patterns changed with scale. Trees of most species pairs co-occurred less than expected by chance and positive associations were rare in local neighbourhoods. Analysis 2 revealed a separation of scales in which significant small-scale interactions faded away at distances of 10 15 m. One third of all species pairs showed significant and mostly negative bivariate small-scale association, which occurred more often than expected by chance between species sharing attributes such as family, fruit type and habitat association. This suggests the occurrence of competitive interactions. Analysis 3 showed that only 8% of all species pairs co-occurred at large scales.3. Comparison of our results with an analogous study conducted in the species-rich tropical forest at Sinharaja, Sri Lanka, revealed several structural similarities including the dominance of segregation and partial overlap in the overall patterning (analysis 1) and the separation of scales (analysis 2). However, species pairs at CBS showed considerably more significant negative small-scale associations (31% vs. 6% at Sinharaja).4. Synthesis. The techniques presented here allow for a detailed analysis of the complex spatial associations in species-rich forests and have the potential to reveal indicative patterns that may allow researchers to discriminate among competing hypotheses of community assemblage and dynamics. However, this will require comparative studies involving a large number of plots.
WrightSJ ( 2002). Plant diversity in tropical forests: A review of mechanisms of species coexistence Oecologia, 130, 1-14. DOI:10.1007/s004420100809URLPMID:28547014 [本文引用: 1] Evidence concerning mechanisms hypothesized to explain species coexistence in hyper-diverse communities is reviewed for tropical forest plants. Three hypotheses receive strong support. Niche differences are evident from non-random spatial distributions along micro-topographic gradients and from a survivorship-growth tradeoff during regeneration. Host-specific pests reduce recruitment near reproductive adults (the Janzen-Connell effect), and, negative density dependence occurs over larger spatial scales among the more abundant species and may regulate their populations. A fourth hypothesis, that suppressed understory plants rarely come into competition with one another, has not been considered before and has profound implications for species coexistence. These hypotheses are mutually compatible. Infrequent competition among suppressed understory plants, niche differences, and Janzen-Connell effects may facilitate the coexistence of the many rare plant species found in tropical forests while negative density dependence regulates the few most successful and abundant species.
WrightSJ, YavittJB, WurzburgerN, TurnerBL, TannerEV, SayerEJ, SantiagoLS, KaspariM, HedinLO, HarmsKE, GarciaMN, CorreMD ( 2011). Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest Ecology, 92, 1616-1625. DOI:10.1890/10-1558.1URL [本文引用: 1]
YanXF, CaoM ( 2007). Effects of light intensity on seed germination and seedling early growth of Shorea wantianshuea. Chinese Journal of Applied Ecology, 18, 23-29. [本文引用: 4]
YanY, ZhangC, WangY, ZhaoX, von GadowK ( 2015). Drivers of seedling survival in a temperate forest and their relative importance at three stages of succession Ecology and Evolution, 5, 4287-4299. DOI:10.1002/ece3.2015.5.issue-19URL [本文引用: 4]
YanY, ZhangXN, YaoJ, ZhangCY, ZhaoXH ( 2016). Composition and temporal dynamics of tree seedlings at different successional stages of conifer and broad-leaved mixed forests in Jiaohe, Jilin Province, China Chinese Journal of Plant Ecology, 40, 127-139. DOI:10.17521/cjpe.2015.0365URL [本文引用: 4] 为了解蛟河地区木本植物幼苗的数量特征及其年际动态,以及影响幼苗数量的生态学过程,该文以不同演替阶段针阔混交林监测样地为基础,在样地内共设置了451个种子雨-幼苗观测样站。通过2012–2014年连续3年的幼苗监测调查,对木本植物幼苗物种组成、数量特征、高度级结构及其年际动态进行了分析,并用多元回归的方法分析了幼苗数量与林分类型、同种成体胸高断面积之和及林冠开阔度的关系。结果表明:(1)林下幼苗层主要木本植物的组成在不同演替阶段林分中差异不大,并与样地内主要树种组成保持着一定相似性,但幼苗重要值排序在年际间和不同演替阶段稍有波动。(2)3个样地内幼苗个体数量和物种数都主要集中在5–20 cm高度级之间,之后随高度级增加而逐渐减少,但幼苗个体数量减少更快。表明与种间竞争相比,物种内部的相互作用是导致幼苗死亡的主要原因,证实了负密度制约效应在幼苗更新过程中的作用。(3)5个主要树种的幼苗数量都与同种成体胸高断面积之和呈显著的正相关关系,还呈现出明显的生境偏好,表明扩散限制和生态位过程也会影响幼苗的数量和分布。该文间接证实了负密度制约效应、扩散限制和生态位过程对温带地区不同演替阶段森林群落中幼苗数量组成的影响。 [ 闫琰, 张新娜, 姚杰, 张春雨, 赵秀海 ( 2016). 吉林蛟河不同演替阶段针阔混交林乔木幼苗数量组成及其时间动态 植物生态学报, 40, 127-139.] DOI:10.17521/cjpe.2015.0365URL [本文引用: 4] 为了解蛟河地区木本植物幼苗的数量特征及其年际动态,以及影响幼苗数量的生态学过程,该文以不同演替阶段针阔混交林监测样地为基础,在样地内共设置了451个种子雨-幼苗观测样站。通过2012–2014年连续3年的幼苗监测调查,对木本植物幼苗物种组成、数量特征、高度级结构及其年际动态进行了分析,并用多元回归的方法分析了幼苗数量与林分类型、同种成体胸高断面积之和及林冠开阔度的关系。结果表明:(1)林下幼苗层主要木本植物的组成在不同演替阶段林分中差异不大,并与样地内主要树种组成保持着一定相似性,但幼苗重要值排序在年际间和不同演替阶段稍有波动。(2)3个样地内幼苗个体数量和物种数都主要集中在5–20 cm高度级之间,之后随高度级增加而逐渐减少,但幼苗个体数量减少更快。表明与种间竞争相比,物种内部的相互作用是导致幼苗死亡的主要原因,证实了负密度制约效应在幼苗更新过程中的作用。(3)5个主要树种的幼苗数量都与同种成体胸高断面积之和呈显著的正相关关系,还呈现出明显的生境偏好,表明扩散限制和生态位过程也会影响幼苗的数量和分布。该文间接证实了负密度制约效应、扩散限制和生态位过程对温带地区不同演替阶段森林群落中幼苗数量组成的影响。
YaoJ, YanY, ZhangCY, PiTH, ZhaoXH ( 2015). Composition and monthly dynamics of tree seedlings in a coniferous and broad-leaved mixed forest in Jiaohe, Jilin Province, China Chinese Journal of Plant Ecology, 39, 717-725. DOI:10.17521/cjpe.2015.0068URL [本文引用: 2] 以吉林省蛟河42hm^2针阔混交林动态监测样地为平台,在样地内选取224个1m×1m幼苗监测小样方,基于2013年6至8月每月中旬的幼苗监测数据,对乔木幼苗物种组成、数量特征、月际动态及幼苗与同种大树之间的关系进行分析。结果表明:(1)该群落共调查到乔木树种幼苗16种。物种组成在6月和7月间变化不大,但与8月差异较大。物种组成在各样方间表现出极大的差异性,不同种幼苗出现的样方数为1-159。(2)6月至8月,幼苗个体从1722株减少到1214株,降幅达29.5%。不同种幼苗在个体数量上变异较大,个体数超过100的幼苗有色木槭(Acer mono)、水曲柳(Fraxinus mandschurica)、紫椴(Tilia amurensis)和东北槭(Acer mandshuricum),四者个体数之和占所有物种个体总数的84.26%;不同种幼苗密度变异性较大,只有色木槭、水曲柳、紫椴和东北槭密度大于1株·m^-2,其他种幼苗密度均较低。(3)新生幼苗更新、死亡格局表现出明显的种间差异与月际问差异,7月新生幼苗总体死亡率(48.9%)显著高于8月(28.3%);整个调查季(6-8月),新生幼苗总体死亡率(40.56%)明显高于多年生幼苗(7.34%)。就幼苗更新情况而言,该研究群落一年中更新主要集中在6月。(4)色木槭、紫椴、杉松(Abies hoZophylla)幼苗的密度与同种成年个体的胸高断面积之和之间存在显著的正相关关系p=O.006、0.013、0.037),表明三者幼苗密度随周围母树多度的增加而增加,其他物种幼苗密度与同种个体数目或同种胸高断面积之和未表现出显著的相关关系。 [ 姚杰, 闫琰, 张春雨, 邳田辉, 赵秀海 ( 2015). 吉林蛟河针阔混交林乔木幼苗组成与月际动态 植物生态学报, 39, 717-725.] DOI:10.17521/cjpe.2015.0068URL [本文引用: 2] 以吉林省蛟河42hm^2针阔混交林动态监测样地为平台,在样地内选取224个1m×1m幼苗监测小样方,基于2013年6至8月每月中旬的幼苗监测数据,对乔木幼苗物种组成、数量特征、月际动态及幼苗与同种大树之间的关系进行分析。结果表明:(1)该群落共调查到乔木树种幼苗16种。物种组成在6月和7月间变化不大,但与8月差异较大。物种组成在各样方间表现出极大的差异性,不同种幼苗出现的样方数为1-159。(2)6月至8月,幼苗个体从1722株减少到1214株,降幅达29.5%。不同种幼苗在个体数量上变异较大,个体数超过100的幼苗有色木槭(Acer mono)、水曲柳(Fraxinus mandschurica)、紫椴(Tilia amurensis)和东北槭(Acer mandshuricum),四者个体数之和占所有物种个体总数的84.26%;不同种幼苗密度变异性较大,只有色木槭、水曲柳、紫椴和东北槭密度大于1株·m^-2,其他种幼苗密度均较低。(3)新生幼苗更新、死亡格局表现出明显的种间差异与月际问差异,7月新生幼苗总体死亡率(48.9%)显著高于8月(28.3%);整个调查季(6-8月),新生幼苗总体死亡率(40.56%)明显高于多年生幼苗(7.34%)。就幼苗更新情况而言,该研究群落一年中更新主要集中在6月。(4)色木槭、紫椴、杉松(Abies hoZophylla)幼苗的密度与同种成年个体的胸高断面积之和之间存在显著的正相关关系p=O.006、0.013、0.037),表明三者幼苗密度随周围母树多度的增加而增加,其他物种幼苗密度与同种个体数目或同种胸高断面积之和未表现出显著的相关关系。
YinHJ, ChengXY, LaiT, LinB, LiuQ ( 2011). Seed rain, soil seed bank and seedling regeneration in a 65-year Picea asperata plantation in subalpine coniferous, western Sichuan, China. Chinese Journal of Plant Ecology, 35, 35-44. DOI:10.3724/SP.J.1258.2011.00035URL [本文引用: 3] 川西亚高山人工针叶林已成为亚高山森林的重要组成部分,它们是否具有持续的自然更新能力,是 决定川西亚高山针叶林群落演替方向和维持该区针叶林大面积存在的基础。以川西米亚罗亚高山人工云杉(Piceaasperata)林(65a)为研究对 象,对种子雨量年际变化、土壤种子库动态、种子萌发和幼苗定居等更新过程的关键环节进行了连续7年(2002-2008年)的野外观测,以研究人工云杉林 更新潜力及影响其更新的限制因素。结果表明:该区云杉林种子雨一般从每年的10月初开始下落,一直到翌年的1月底或2月初结束;云杉种子散落存在明显的大 小年现象,种子散落周期为4年,且大小年之间种子产量差异极大:云杉种子从下落到土壤到种子完全失去活力不到1年时间,属于Thompson和Grime 定义的第1I类土壤种子库类型。腐烂死亡和动物取食是土壤种子库损耗的主要因素,而种子通过萌发真正转化为幼苗的比例非常低,仅占2002年下落种子总量 的3.6%。种子萌发后,环境筛的作用导致云杉幼苗大量死亡,尤其是在种子萌发后的一个生长季节内,其死亡率高达78%。凋落物和苔藓是构成人工云杉林下 地表的两种主要地被物类型,二者占所有调查幼苗数量的93%左右;两种地被物类型上肚2cm层幼苗存活率最高,分别占存活幼苗总数的76.07%和 86.72%,随地被物厚度增加,幼苗存活率呈明显下降趋势,而幼苗死亡率呈明显升高的趋势,表明林下地被物厚度也是影响云杉幼苗定居的重要因素。两种地 被物对幼苗生长的影响不同,除株高之外,分布在苔藓上的云杉幼苗生长参数(地径、分枝数、干重以及干重年增长率)明显高于分布于凋落物上的幼苗,表明苔藓 地被物更有利于云杉幼苗定居。尽管该区大量云杉种子下落,但由于种子的高损耗率、幼苗的 [ 尹华军, 程新颖, 赖挺, 林波, 刘庆 ( 2011). 川西亚高山65年人工云杉林种子雨、种子库和幼苗定居研究 植物生态学报, 35, 35-44.] DOI:10.3724/SP.J.1258.2011.00035URL [本文引用: 3] 川西亚高山人工针叶林已成为亚高山森林的重要组成部分,它们是否具有持续的自然更新能力,是 决定川西亚高山针叶林群落演替方向和维持该区针叶林大面积存在的基础。以川西米亚罗亚高山人工云杉(Piceaasperata)林(65a)为研究对 象,对种子雨量年际变化、土壤种子库动态、种子萌发和幼苗定居等更新过程的关键环节进行了连续7年(2002-2008年)的野外观测,以研究人工云杉林 更新潜力及影响其更新的限制因素。结果表明:该区云杉林种子雨一般从每年的10月初开始下落,一直到翌年的1月底或2月初结束;云杉种子散落存在明显的大 小年现象,种子散落周期为4年,且大小年之间种子产量差异极大:云杉种子从下落到土壤到种子完全失去活力不到1年时间,属于Thompson和Grime 定义的第1I类土壤种子库类型。腐烂死亡和动物取食是土壤种子库损耗的主要因素,而种子通过萌发真正转化为幼苗的比例非常低,仅占2002年下落种子总量 的3.6%。种子萌发后,环境筛的作用导致云杉幼苗大量死亡,尤其是在种子萌发后的一个生长季节内,其死亡率高达78%。凋落物和苔藓是构成人工云杉林下 地表的两种主要地被物类型,二者占所有调查幼苗数量的93%左右;两种地被物类型上肚2cm层幼苗存活率最高,分别占存活幼苗总数的76.07%和 86.72%,随地被物厚度增加,幼苗存活率呈明显下降趋势,而幼苗死亡率呈明显升高的趋势,表明林下地被物厚度也是影响云杉幼苗定居的重要因素。两种地 被物对幼苗生长的影响不同,除株高之外,分布在苔藓上的云杉幼苗生长参数(地径、分枝数、干重以及干重年增长率)明显高于分布于凋落物上的幼苗,表明苔藓 地被物更有利于云杉幼苗定居。尽管该区大量云杉种子下落,但由于种子的高损耗率、幼苗的
YuF, ShiXX, ChenLL, HuangQP, SongB ( 2013). Species composition and regeneration characteristics of main woody plant seedlings in a pine-oak mixed forest in Qinling Mountains Acta Botanica Boreali-Occidentalia Sinica, 33, 592-598. [本文引用: 1]
Seed dispersal near and far: Patterns across temperate and tropical forests 1 1999
... 幼苗的存活与更新受到多种因素的影响, 主要分为生物和非生物因素两大类.生物因素包括植物自身的属性、生物邻体、植食性动物以及病原体等(Mangan et al., 2010; Johnson et al., 2012; Norghauer & Newbery, 2014; 苏嫄等, 2014; 骆杨青等, 2017); 非生物因素包括光照、水分、土壤等(Kammesheidt, 1998; Connell & Green, 2000; Poorter & Rose, 2005; 王传华等, 2011; 刘帅等, 2016).植物种子的产量、扩散距离、传播方式等直接影响幼苗更新密度格局(Clark et al., 1999).幼苗更新空间中包含大量邻体, 基于J-C假说的研究发现同种邻体间可能通过传播专一宿主的自然天敌和食草动物的取食抑制同种幼苗的存活, 而异种邻体的聚集可能会阻碍特异性天敌的传播, 从而有利于同种幼苗的存活(Janzen 1970; Connell 1971; Makana & Thomas, 2004; Johnson et al., 2012; Comita et al., 2014).此外, Johnson等(2014)发现当幼苗周围同种成体密度较大时, 幼苗密度较高, 与母树多度相关的植物种子的扩散限制可能是决定乔木幼苗密度格局的一个重要因素. ...
Local neighborhood and species’ shade tolerance influence survival in a diverse seedling bank 2 2009
... 为了检验生物邻体和生境异质性对幼苗密度格局的相对重要性, 研究划分了4个模型: (1)零模型(以样站和物种作为随机效应).(2)生物模型(在零模型基础上, 以大树邻体作为固定效应).大树效应是指以幼苗样方为中心一定半径范围内的同种大树胸高断面积之和与异种大树胸高断面积之和(Comita & Hubbell, 2009).大树邻体效应随距离(半径)增大而逐渐减弱, 有关温带森林的研究发现距离超过20 m后树种间的相互作用变得极小(Wang et al., 2010), 故选取半径20 m研究大树邻体的影响(Canham et al., 2004; Bai et al., 2012).(3)生境模型(在零模型基础上, 以林冠开阔度、土壤含水量、pH值和土壤养分作为固定效应).(4)全模型(在零模型基础上, 生物邻体和生境因子作为固定效应).模型的比较选择运用赤池信息量准则(AIC), AIC值越小, 代表模型拟合越好, 模型之间AIC值的差值小于2时, 这两个模型被视为是等同的(Burnham & Anderson, 2002).我们在3个水平下检验了影响幼苗密度的生物与生境因素: (1)群落水平, 包括所有物种幼苗; (2)龄级水平, 当年生和多年生幼苗; (3)物种水平, 以2016年存活的幼苗个体中重要值排名前5位的物种进行分析.为考虑群落空间自相关, 所有模型将样站作为一个随机效应(Bai et al., 2012), 不同物种间幼苗密度差异较大, 因此将物种作为另一个随机效应加入到群落水平和年龄级的模型中(Chen et al., 2010).本文中所有数据分析与绘图均采用Microsoft Excel 2016和国际通用软件R 3.4.2 (http://www.r-project. org), 其中GLMM分析采用‘lme4’软件包完成. ...
... 扩散限制和生境过滤共同影响幼苗密度格局.本研究中, 群落水平上的幼苗密度与同种成年邻体和土壤PC1、PC2呈显著正相关, 生物邻体与生境异质性对幼苗密度都有显著影响, Bai等(2012)和Oshima等(2015)得到了一致的结论.适宜大树生长的局域环境同样适宜幼苗的更新(Comita & Hubbell, 2009; Comita et al., 2014), 局域同种成体密度越高, 群落的幼苗密度越高, 这与闫琰等(2016)对吉林蛟河针阔叶混交林幼苗更新的研究结果一致, 一定程度上证明了种子的扩散限制是决定树木幼苗种群大小的一个重要因素(Makana & Thomas, 2004; Norden et al., 2009). ...
Testing predictions of the Janzen-Connell hypothesis: A meta-analysis of experimental evidence for distance- and density-dependent seed and seedling survival 2 2014
... 幼苗的存活与更新受到多种因素的影响, 主要分为生物和非生物因素两大类.生物因素包括植物自身的属性、生物邻体、植食性动物以及病原体等(Mangan et al., 2010; Johnson et al., 2012; Norghauer & Newbery, 2014; 苏嫄等, 2014; 骆杨青等, 2017); 非生物因素包括光照、水分、土壤等(Kammesheidt, 1998; Connell & Green, 2000; Poorter & Rose, 2005; 王传华等, 2011; 刘帅等, 2016).植物种子的产量、扩散距离、传播方式等直接影响幼苗更新密度格局(Clark et al., 1999).幼苗更新空间中包含大量邻体, 基于J-C假说的研究发现同种邻体间可能通过传播专一宿主的自然天敌和食草动物的取食抑制同种幼苗的存活, 而异种邻体的聚集可能会阻碍特异性天敌的传播, 从而有利于同种幼苗的存活(Janzen 1970; Connell 1971; Makana & Thomas, 2004; Johnson et al., 2012; Comita et al., 2014).此外, Johnson等(2014)发现当幼苗周围同种成体密度较大时, 幼苗密度较高, 与母树多度相关的植物种子的扩散限制可能是决定乔木幼苗密度格局的一个重要因素. ...
... 扩散限制和生境过滤共同影响幼苗密度格局.本研究中, 群落水平上的幼苗密度与同种成年邻体和土壤PC1、PC2呈显著正相关, 生物邻体与生境异质性对幼苗密度都有显著影响, Bai等(2012)和Oshima等(2015)得到了一致的结论.适宜大树生长的局域环境同样适宜幼苗的更新(Comita & Hubbell, 2009; Comita et al., 2014), 局域同种成体密度越高, 群落的幼苗密度越高, 这与闫琰等(2016)对吉林蛟河针阔叶混交林幼苗更新的研究结果一致, 一定程度上证明了种子的扩散限制是决定树木幼苗种群大小的一个重要因素(Makana & Thomas, 2004; Norden et al., 2009). ...
On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In: den Boer PJ, Gradwell GR eds. Dynamics of Population 1 1971
... 幼苗的存活与更新受到多种因素的影响, 主要分为生物和非生物因素两大类.生物因素包括植物自身的属性、生物邻体、植食性动物以及病原体等(Mangan et al., 2010; Johnson et al., 2012; Norghauer & Newbery, 2014; 苏嫄等, 2014; 骆杨青等, 2017); 非生物因素包括光照、水分、土壤等(Kammesheidt, 1998; Connell & Green, 2000; Poorter & Rose, 2005; 王传华等, 2011; 刘帅等, 2016).植物种子的产量、扩散距离、传播方式等直接影响幼苗更新密度格局(Clark et al., 1999).幼苗更新空间中包含大量邻体, 基于J-C假说的研究发现同种邻体间可能通过传播专一宿主的自然天敌和食草动物的取食抑制同种幼苗的存活, 而异种邻体的聚集可能会阻碍特异性天敌的传播, 从而有利于同种幼苗的存活(Janzen 1970; Connell 1971; Makana & Thomas, 2004; Johnson et al., 2012; Comita et al., 2014).此外, Johnson等(2014)发现当幼苗周围同种成体密度较大时, 幼苗密度较高, 与母树多度相关的植物种子的扩散限制可能是决定乔木幼苗密度格局的一个重要因素. ...
Seedling dynamics over thirty-two years in a tropical rain forest tree 1 2000
... 幼苗的存活与更新受到多种因素的影响, 主要分为生物和非生物因素两大类.生物因素包括植物自身的属性、生物邻体、植食性动物以及病原体等(Mangan et al., 2010; Johnson et al., 2012; Norghauer & Newbery, 2014; 苏嫄等, 2014; 骆杨青等, 2017); 非生物因素包括光照、水分、土壤等(Kammesheidt, 1998; Connell & Green, 2000; Poorter & Rose, 2005; 王传华等, 2011; 刘帅等, 2016).植物种子的产量、扩散距离、传播方式等直接影响幼苗更新密度格局(Clark et al., 1999).幼苗更新空间中包含大量邻体, 基于J-C假说的研究发现同种邻体间可能通过传播专一宿主的自然天敌和食草动物的取食抑制同种幼苗的存活, 而异种邻体的聚集可能会阻碍特异性天敌的传播, 从而有利于同种幼苗的存活(Janzen 1970; Connell 1971; Makana & Thomas, 2004; Johnson et al., 2012; Comita et al., 2014).此外, Johnson等(2014)发现当幼苗周围同种成体密度较大时, 幼苗密度较高, 与母树多度相关的植物种子的扩散限制可能是决定乔木幼苗密度格局的一个重要因素. ...
Herbivores and the number of tree species in tropical forests 1 1970
... 幼苗的存活与更新受到多种因素的影响, 主要分为生物和非生物因素两大类.生物因素包括植物自身的属性、生物邻体、植食性动物以及病原体等(Mangan et al., 2010; Johnson et al., 2012; Norghauer & Newbery, 2014; 苏嫄等, 2014; 骆杨青等, 2017); 非生物因素包括光照、水分、土壤等(Kammesheidt, 1998; Connell & Green, 2000; Poorter & Rose, 2005; 王传华等, 2011; 刘帅等, 2016).植物种子的产量、扩散距离、传播方式等直接影响幼苗更新密度格局(Clark et al., 1999).幼苗更新空间中包含大量邻体, 基于J-C假说的研究发现同种邻体间可能通过传播专一宿主的自然天敌和食草动物的取食抑制同种幼苗的存活, 而异种邻体的聚集可能会阻碍特异性天敌的传播, 从而有利于同种幼苗的存活(Janzen 1970; Connell 1971; Makana & Thomas, 2004; Johnson et al., 2012; Comita et al., 2014).此外, Johnson等(2014)发现当幼苗周围同种成体密度较大时, 幼苗密度较高, 与母树多度相关的植物种子的扩散限制可能是决定乔木幼苗密度格局的一个重要因素. ...
Conspecific negative density dependence and forest diversity 2 2012
... 幼苗的存活与更新受到多种因素的影响, 主要分为生物和非生物因素两大类.生物因素包括植物自身的属性、生物邻体、植食性动物以及病原体等(Mangan et al., 2010; Johnson et al., 2012; Norghauer & Newbery, 2014; 苏嫄等, 2014; 骆杨青等, 2017); 非生物因素包括光照、水分、土壤等(Kammesheidt, 1998; Connell & Green, 2000; Poorter & Rose, 2005; 王传华等, 2011; 刘帅等, 2016).植物种子的产量、扩散距离、传播方式等直接影响幼苗更新密度格局(Clark et al., 1999).幼苗更新空间中包含大量邻体, 基于J-C假说的研究发现同种邻体间可能通过传播专一宿主的自然天敌和食草动物的取食抑制同种幼苗的存活, 而异种邻体的聚集可能会阻碍特异性天敌的传播, 从而有利于同种幼苗的存活(Janzen 1970; Connell 1971; Makana & Thomas, 2004; Johnson et al., 2012; Comita et al., 2014).此外, Johnson等(2014)发现当幼苗周围同种成体密度较大时, 幼苗密度较高, 与母树多度相关的植物种子的扩散限制可能是决定乔木幼苗密度格局的一个重要因素. ...
... ; Johnson et al., 2012; Comita et al., 2014).此外, Johnson等(2014)发现当幼苗周围同种成体密度较大时, 幼苗密度较高, 与母树多度相关的植物种子的扩散限制可能是决定乔木幼苗密度格局的一个重要因素. ...
Conspecific negative density-dependent mortality and the structure of temperate forests 1 2014
... 幼苗的存活与更新受到多种因素的影响, 主要分为生物和非生物因素两大类.生物因素包括植物自身的属性、生物邻体、植食性动物以及病原体等(Mangan et al., 2010; Johnson et al., 2012; Norghauer & Newbery, 2014; 苏嫄等, 2014; 骆杨青等, 2017); 非生物因素包括光照、水分、土壤等(Kammesheidt, 1998; Connell & Green, 2000; Poorter & Rose, 2005; 王传华等, 2011; 刘帅等, 2016).植物种子的产量、扩散距离、传播方式等直接影响幼苗更新密度格局(Clark et al., 1999).幼苗更新空间中包含大量邻体, 基于J-C假说的研究发现同种邻体间可能通过传播专一宿主的自然天敌和食草动物的取食抑制同种幼苗的存活, 而异种邻体的聚集可能会阻碍特异性天敌的传播, 从而有利于同种幼苗的存活(Janzen 1970; Connell 1971; Makana & Thomas, 2004; Johnson et al., 2012; Comita et al., 2014).此外, Johnson等(2014)发现当幼苗周围同种成体密度较大时, 幼苗密度较高, 与母树多度相关的植物种子的扩散限制可能是决定乔木幼苗密度格局的一个重要因素. ...
The role of tree sprouts in the restoration of stand structure and species diversity in tropical moist forest after slash-and-burn agriculture in Eastern Paraguay 1 1998
... 幼苗的存活与更新受到多种因素的影响, 主要分为生物和非生物因素两大类.生物因素包括植物自身的属性、生物邻体、植食性动物以及病原体等(Mangan et al., 2010; Johnson et al., 2012; Norghauer & Newbery, 2014; 苏嫄等, 2014; 骆杨青等, 2017); 非生物因素包括光照、水分、土壤等(Kammesheidt, 1998; Connell & Green, 2000; Poorter & Rose, 2005; 王传华等, 2011; 刘帅等, 2016).植物种子的产量、扩散距离、传播方式等直接影响幼苗更新密度格局(Clark et al., 1999).幼苗更新空间中包含大量邻体, 基于J-C假说的研究发现同种邻体间可能通过传播专一宿主的自然天敌和食草动物的取食抑制同种幼苗的存活, 而异种邻体的聚集可能会阻碍特异性天敌的传播, 从而有利于同种幼苗的存活(Janzen 1970; Connell 1971; Makana & Thomas, 2004; Johnson et al., 2012; Comita et al., 2014).此外, Johnson等(2014)发现当幼苗周围同种成体密度较大时, 幼苗密度较高, 与母树多度相关的植物种子的扩散限制可能是决定乔木幼苗密度格局的一个重要因素. ...
Seed and seedling ecology of a monocarpic tropical tree 1 1989
... 处在林下环境的植物幼苗往往受光照、水分、土壤养分等环境因子的限制(闫兴富和曹敏, 2007; Pasquini & Santiago, 2012; 施璐璐等, 2014).在环境资源竞争中部分幼苗占据优势得以生存, 而处于劣势的幼苗生长受到抑制甚至死亡(祝燕等, 2009).近年来, 探讨幼苗更新影响因素的研究有很多, 研究的出发点有以下两种: (1)单一环境因素对幼苗更新的影响(Montgomery & Chazdon, 2002; 闫兴富和曹敏, 2007; Song et al., 2013).如闫兴富和曹敏(2007)分析了光照强度对望天树(Parashorea chinensis)幼苗更新的影响, 发现中等程度的遮阴有利于种子萌发, 强光照和深度遮阴均对种子萌发不利, 幼苗株高、基径等生长指标均在部分遮阴处理条件下最大.李晓亮等(2009)发现幼苗会因水分胁迫发生大量的死亡, 且这种死亡格局有明显的种间差异.Wright等(2011)通过土壤养分元素(N、P、K)连续11年的添加实验发现土壤养分对植物幼苗生长有强烈的影响.(2)多个环境因素对幼苗更新的综合影响(王传华等, 2011; 尹华军等, 2011; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... ; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... ).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... 处在林下环境的植物幼苗往往受光照、水分、土壤养分等环境因子的限制(闫兴富和曹敏, 2007; Pasquini & Santiago, 2012; 施璐璐等, 2014).在环境资源竞争中部分幼苗占据优势得以生存, 而处于劣势的幼苗生长受到抑制甚至死亡(祝燕等, 2009).近年来, 探讨幼苗更新影响因素的研究有很多, 研究的出发点有以下两种: (1)单一环境因素对幼苗更新的影响(Montgomery & Chazdon, 2002; 闫兴富和曹敏, 2007; Song et al., 2013).如闫兴富和曹敏(2007)分析了光照强度对望天树(Parashorea chinensis)幼苗更新的影响, 发现中等程度的遮阴有利于种子萌发, 强光照和深度遮阴均对种子萌发不利, 幼苗株高、基径等生长指标均在部分遮阴处理条件下最大.李晓亮等(2009)发现幼苗会因水分胁迫发生大量的死亡, 且这种死亡格局有明显的种间差异.Wright等(2011)通过土壤养分元素(N、P、K)连续11年的添加实验发现土壤养分对植物幼苗生长有强烈的影响.(2)多个环境因素对幼苗更新的综合影响(王传华等, 2011; 尹华军等, 2011; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... ; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... ).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest 1 2011
... 处在林下环境的植物幼苗往往受光照、水分、土壤养分等环境因子的限制(闫兴富和曹敏, 2007; Pasquini & Santiago, 2012; 施璐璐等, 2014).在环境资源竞争中部分幼苗占据优势得以生存, 而处于劣势的幼苗生长受到抑制甚至死亡(祝燕等, 2009).近年来, 探讨幼苗更新影响因素的研究有很多, 研究的出发点有以下两种: (1)单一环境因素对幼苗更新的影响(Montgomery & Chazdon, 2002; 闫兴富和曹敏, 2007; Song et al., 2013).如闫兴富和曹敏(2007)分析了光照强度对望天树(Parashorea chinensis)幼苗更新的影响, 发现中等程度的遮阴有利于种子萌发, 强光照和深度遮阴均对种子萌发不利, 幼苗株高、基径等生长指标均在部分遮阴处理条件下最大.李晓亮等(2009)发现幼苗会因水分胁迫发生大量的死亡, 且这种死亡格局有明显的种间差异.Wright等(2011)通过土壤养分元素(N、P、K)连续11年的添加实验发现土壤养分对植物幼苗生长有强烈的影响.(2)多个环境因素对幼苗更新的综合影响(王传华等, 2011; 尹华军等, 2011; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
不同光照对望天树种子萌发和幼苗早期生长的影响 4 2007
... 处在林下环境的植物幼苗往往受光照、水分、土壤养分等环境因子的限制(闫兴富和曹敏, 2007; Pasquini & Santiago, 2012; 施璐璐等, 2014).在环境资源竞争中部分幼苗占据优势得以生存, 而处于劣势的幼苗生长受到抑制甚至死亡(祝燕等, 2009).近年来, 探讨幼苗更新影响因素的研究有很多, 研究的出发点有以下两种: (1)单一环境因素对幼苗更新的影响(Montgomery & Chazdon, 2002; 闫兴富和曹敏, 2007; Song et al., 2013).如闫兴富和曹敏(2007)分析了光照强度对望天树(Parashorea chinensis)幼苗更新的影响, 发现中等程度的遮阴有利于种子萌发, 强光照和深度遮阴均对种子萌发不利, 幼苗株高、基径等生长指标均在部分遮阴处理条件下最大.李晓亮等(2009)发现幼苗会因水分胁迫发生大量的死亡, 且这种死亡格局有明显的种间差异.Wright等(2011)通过土壤养分元素(N、P、K)连续11年的添加实验发现土壤养分对植物幼苗生长有强烈的影响.(2)多个环境因素对幼苗更新的综合影响(王传华等, 2011; 尹华军等, 2011; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... ; 闫兴富和曹敏, 2007; Song et al., 2013).如闫兴富和曹敏(2007)分析了光照强度对望天树(Parashorea chinensis)幼苗更新的影响, 发现中等程度的遮阴有利于种子萌发, 强光照和深度遮阴均对种子萌发不利, 幼苗株高、基径等生长指标均在部分遮阴处理条件下最大.李晓亮等(2009)发现幼苗会因水分胁迫发生大量的死亡, 且这种死亡格局有明显的种间差异.Wright等(2011)通过土壤养分元素(N、P、K)连续11年的添加实验发现土壤养分对植物幼苗生长有强烈的影响.(2)多个环境因素对幼苗更新的综合影响(王传华等, 2011; 尹华军等, 2011; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... ).如闫兴富和曹敏(2007)分析了光照强度对望天树(Parashorea chinensis)幼苗更新的影响, 发现中等程度的遮阴有利于种子萌发, 强光照和深度遮阴均对种子萌发不利, 幼苗株高、基径等生长指标均在部分遮阴处理条件下最大.李晓亮等(2009)发现幼苗会因水分胁迫发生大量的死亡, 且这种死亡格局有明显的种间差异.Wright等(2011)通过土壤养分元素(N、P、K)连续11年的添加实验发现土壤养分对植物幼苗生长有强烈的影响.(2)多个环境因素对幼苗更新的综合影响(王传华等, 2011; 尹华军等, 2011; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... 处在林下环境的植物幼苗往往受光照、水分、土壤养分等环境因子的限制(闫兴富和曹敏, 2007; Pasquini & Santiago, 2012; 施璐璐等, 2014).在环境资源竞争中部分幼苗占据优势得以生存, 而处于劣势的幼苗生长受到抑制甚至死亡(祝燕等, 2009).近年来, 探讨幼苗更新影响因素的研究有很多, 研究的出发点有以下两种: (1)单一环境因素对幼苗更新的影响(Montgomery & Chazdon, 2002; 闫兴富和曹敏, 2007; Song et al., 2013).如闫兴富和曹敏(2007)分析了光照强度对望天树(Parashorea chinensis)幼苗更新的影响, 发现中等程度的遮阴有利于种子萌发, 强光照和深度遮阴均对种子萌发不利, 幼苗株高、基径等生长指标均在部分遮阴处理条件下最大.李晓亮等(2009)发现幼苗会因水分胁迫发生大量的死亡, 且这种死亡格局有明显的种间差异.Wright等(2011)通过土壤养分元素(N、P、K)连续11年的添加实验发现土壤养分对植物幼苗生长有强烈的影响.(2)多个环境因素对幼苗更新的综合影响(王传华等, 2011; 尹华军等, 2011; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... ; 闫兴富和曹敏, 2007; Song et al., 2013).如闫兴富和曹敏(2007)分析了光照强度对望天树(Parashorea chinensis)幼苗更新的影响, 发现中等程度的遮阴有利于种子萌发, 强光照和深度遮阴均对种子萌发不利, 幼苗株高、基径等生长指标均在部分遮阴处理条件下最大.李晓亮等(2009)发现幼苗会因水分胁迫发生大量的死亡, 且这种死亡格局有明显的种间差异.Wright等(2011)通过土壤养分元素(N、P、K)连续11年的添加实验发现土壤养分对植物幼苗生长有强烈的影响.(2)多个环境因素对幼苗更新的综合影响(王传华等, 2011; 尹华军等, 2011; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
... ).如闫兴富和曹敏(2007)分析了光照强度对望天树(Parashorea chinensis)幼苗更新的影响, 发现中等程度的遮阴有利于种子萌发, 强光照和深度遮阴均对种子萌发不利, 幼苗株高、基径等生长指标均在部分遮阴处理条件下最大.李晓亮等(2009)发现幼苗会因水分胁迫发生大量的死亡, 且这种死亡格局有明显的种间差异.Wright等(2011)通过土壤养分元素(N、P、K)连续11年的添加实验发现土壤养分对植物幼苗生长有强烈的影响.(2)多个环境因素对幼苗更新的综合影响(王传华等, 2011; 尹华军等, 2011; 施璐璐等, 2014; Oshima et al., 2015).如施璐璐等(2014)发现幼苗的种子萌发、存活数量与光照和水分都有极显著的相关性, 且与水分关联最大.Oshima等(2015)发现南亚热带雨林幼苗存活与光照和土壤水分等生境因子显著相关.刘帅等(2016)对长白山次生针阔混交林研究发现幼苗存活率与土壤含水量和土壤有效氮等养分含量显著正相关.有关温带森林幼苗存活机制的研究已有报道(张健等, 2009; Bai et al., 2012; Lin et al., 2014; Yan et al., 2015), 而幼苗密度格局影响因素的研究比较有限. ...
 ,*北京林业大学国家林业和草原局森林经营工程技术研究中心, 北京 100083
,*北京林业大学国家林业和草原局森林经营工程技术研究中心, 北京 100083
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT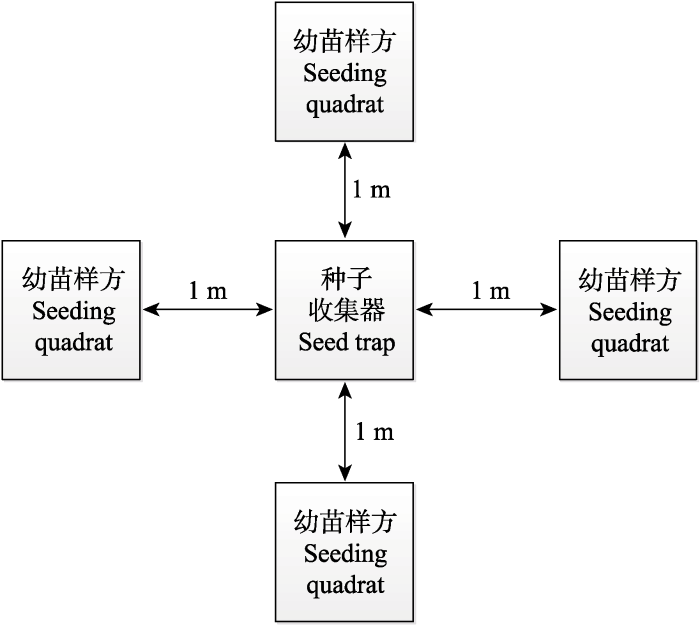 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT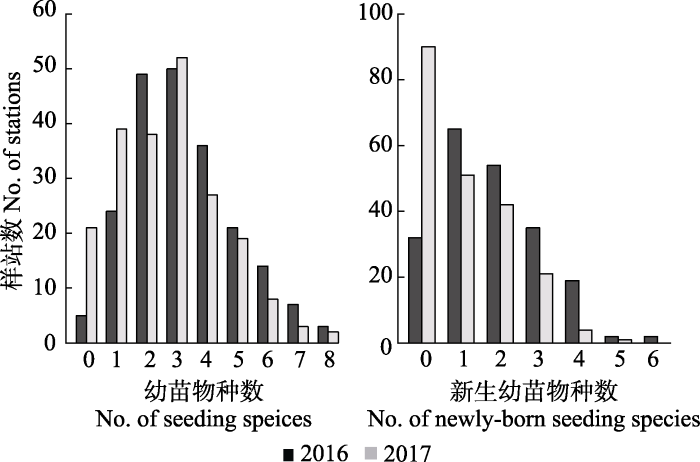 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT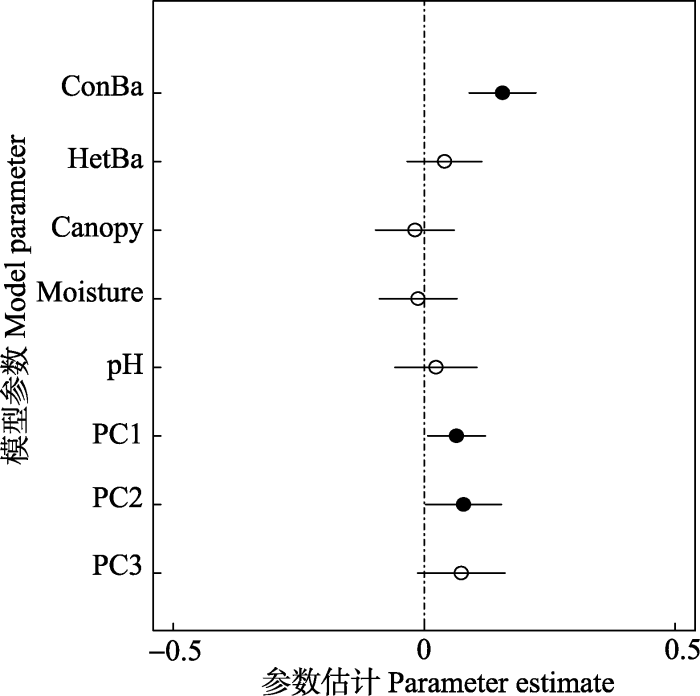 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT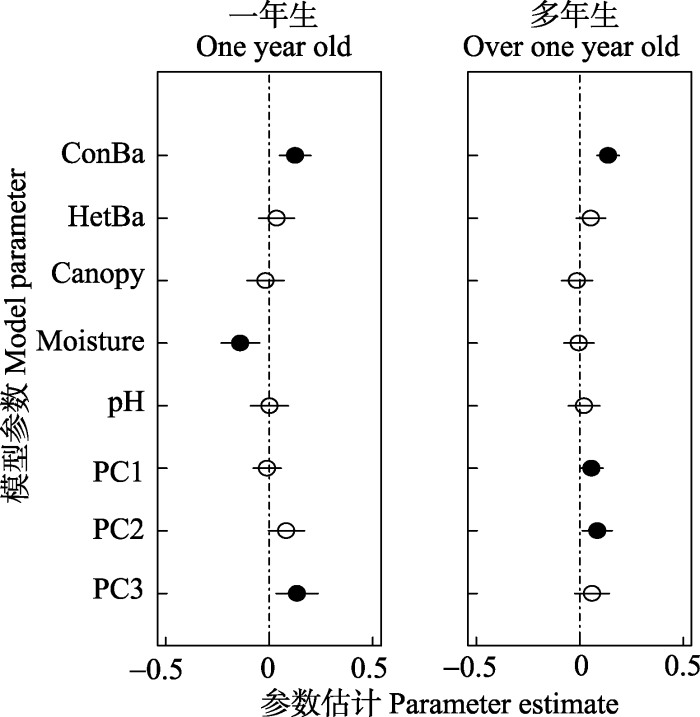 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT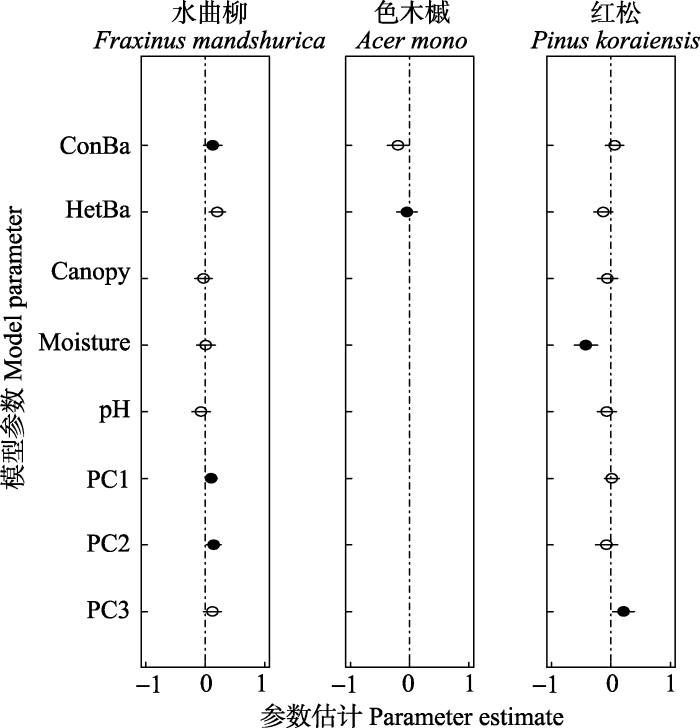 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}