 ,*中国热带农业科学院环境与植物保护研究所/农业部儋州农业环境科学观测实验站, 海口 571101
,*中国热带农业科学院环境与植物保护研究所/农业部儋州农业环境科学观测实验站, 海口 571101Seasonal changes of photosynthetic characteristics of Alpinia oxyphylla growing under Hevea brasiliensis
CHENG Han-Ting, LI Qin-Fen, LIU Jing-Kun, YAN Ting-Liang, ZHANG Qiao-Yan, WANG Jin-Chuang,*Environment and Plant Protection Institute, Chinese Academy of Tropical Agricultural Sciences/Danzhou Scientific Observing and Experimental Station of Agro-Environment, Ministry of Agriculture of the People’s Republic of China, Haikou 571101, China通讯作者: jinchuangwang@yahoo.com
编委: 王根轩
责任编辑: 李敏
收稿日期:2018-01-12修回日期:2018-04-08网络出版日期:2018-05-20
Received:2018-01-12Revised:2018-04-08Online:2018-05-20

摘要
利用农林复合模式发展生态农业可提高资源利用效率, 橡胶(Hevea brasiliensis)-益智(Alpinia oxyphylla)间作模式是橡胶园最主要的农林复合模式。该研究通过野外原位定位实验, 研究不同季节橡胶林下环境因子对益智光合作用的影响, 并进一步分析益智光合作用与主要环境因子的关系。结果表明: (1) 3月益智净光合速率日变化为“V”形曲线, 14:00降到最低值; 而6月、9月和12月益智净光合速率日变化趋势为10:00达到最大值, 随后缓慢降低; 在雨季(6月和9月)蒸腾速率的日平均值和日最高值均显著高于旱季(3月和12月)。表明林下益智在不同季节均能维持植株正常生长, 且表现出了较强的适应能力。3月土壤水分亏缺造成益智叶片气孔导度降低, 使其净光合速率维持在较低的水平。(2)通过光响应曲线修正模型计算出益智叶片各光合响应参数, 发现3月最大净光合效率和光饱和点显著低于6月、9月和12月; 而光补偿点和暗呼吸速率却显著高于6月、9月和12月, 表明3月土壤水分亏缺导致益智光合酶活性降低, 而表现出光抑制现象, 同时呼吸强度加剧, 光合能力显著下降。(3)采取相关分析发现, 3月气温与净光合速率显著负相关, 空气湿度与净光合速率显著正相关, 高温和低湿度共同限制了益智的光合作用; 而9月和12月, 林下光合有效辐射是益智光合作用的限制因子。
关键词:
Abstract
Keywords:
PDF (1214KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
程汉亭, 李勤奋, 刘景坤, 严廷良, 张俏燕, 王进闯. 橡胶林下益智光合特性的季节动态变化. 植物生态学报[J], 2018, 42(5): 585-594 DOI:10.17521/cjpe.2018.0016
CHENG Han-Ting, LI Qin-Fen, LIU Jing-Kun, YAN Ting-Liang, ZHANG Qiao-Yan, WANG Jin-Chuang.
利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(张斌和张桃林, 1997; Nair & Graetz, 2004; Mcneely & Schroth, 2006; Dossa et al., 2008)。橡胶(Hevea brasiliensis)-益智(Alpinia oxyphylla)间作模式是中国海南推广和种植面积最大的农林复合模式, 也是橡胶种植户脱贫致富的有效方法(Ogwuche et al., 2012), 具有较好的经济效益和生态效益(程汉亭等, 2015)。益智为姜科山姜属(Alpinia)多年生常绿草本植物, 主要分布在中国的海南、广东、广西, 近年来云南、福建亦有少量试种; 生于或栽培于林下阴湿处(程汉亭等, 2017), 是我国重点发展的“四大南药”之一, 也是海南特有的药用植物。
光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(Tuittila et al., 2004)。植物的光合速率不仅受环境因子的影响, 还与植物本身的生理生态特性密切相关。对气体交换的研究有利于深入了解林下植物的光合活性和生产力, 特别是旱季土壤水分胁迫对植物碳同化和水分利用效率的影响(Muthuri et al., 2009)。同时, 了解气体交换的季节变化特征有助于改进对林下植物水分需求的有效管理。热带地区雨/旱季分明, 橡胶林土壤水分(祁栋灵等, 2017)、林下光合有效辐射(PAR)(吴志祥等, 2013)、橡胶树物候期(陈小敏等, 2016)呈季节性变化, 这些因素必然会影响林下植物的生长和光合作用。目前国内外****已经开展了一些环境因子对橡胶林下植物生长和光合特性的研究。沈守艮和郑征(2008)研究了西双版纳橡胶-茶农林复合系统中耐阴植物普洱茶(Camellia assamica)的光合特性, 发现林下PAR是引起林下茶树光合差异的主要因素, 橡胶林下PAR的增强提高了茶树光合产物的积累。在橡胶-小粒咖啡(Coffea arabica)农林复合系统里, Righi等(2007, 2008)分别研究了不同光照和水分梯度下咖啡树的生长状况, 获得了林下咖啡树生长的最佳光照和水分条件。Righi等(2008)发现橡胶-菜豆(Phaseolus vulgaris)农林复合系统林下可利用PAR量是菜豆维持正常生长和产量的必要保障。然而, 环境因子对橡胶林下益智光合作用的研究却鲜有报道, 特别是益智光合作用的季节特征如何?不同时间尺度下其光合作用主要受到哪些环境因子的制约?对这些问题尚缺乏系统深入的认识。
本研究通过野外定位实验, 分析不同季节橡胶林下PAR、温度、湿度和CO2浓度等因子对益智光合作用的影响, 研究益智叶片光合碳同化过程对林下环境季节变化的生理响应机制, 揭示益智光合碳同化对橡胶林下小气候季节性变化的适应策略, 为进一步提出橡胶-益智农林复合系统的科学管理措施提供理论依据。
1 材料和方法
1.1 样地概况
实验选择在海南省白沙黎族自治县阜龙乡天堂村人工种植的橡胶林(109.47° E, 19.38° N, 海拔780 m)。所选的实验样地中橡胶树树龄13年, 平均胸径18.3 cm, 树冠平均高度约11 m, 平均冠幅7 m, 盖度80%-85%, 橡胶树株行距3 m × 6 m; 橡胶树6 m的宽行里间作1行益智, 益智株距1.5 m, 种植时间约6年, 株高1.5-1.8 m。实验样地所在区域属于典型的热带季风气候, 终年高温, 雨/旱季明显, 降水集中在雨季, 且降水量大(图1)。从2016年1月至2016年12月, 海南白沙黎族自治县天堂村实验样地气温变幅为4.7-40.9 ℃, 最低气温出现在2016年的1月, 最高气温出现在2016年的3月和4月; 月平均最低气温出现在2月, 最高气温出现在6月(图1A)。实验样地PAR季节变化如图1B所示, PAR季节性差异明显, 1月最低, 平均为621 μmol·m-2·s-1, 7月最高, 平均值为1 436 μmol·m-2·s-1。全年降水量表现为1-4月、11和12月的月降水量均低于54 mm, 其中2月、11月和12月降水量最低, 分别为15.8、11.6和12.0 mm; 5-10月降水充沛, 月降水量均高于100 mm, 其中8月降水量高达1201.2 mm (图1C)。土壤含水量月变化趋势与降水量的趋势一致, 1-3月土壤含水量低, 分别为11.98%、11.97%、12.00%, 此时土壤属于轻度干旱状态; 6-9月土壤潮湿, 其含水量均高于16.35%, 其中8月土壤含水量最高, 为19.32% (图1C)。
图1
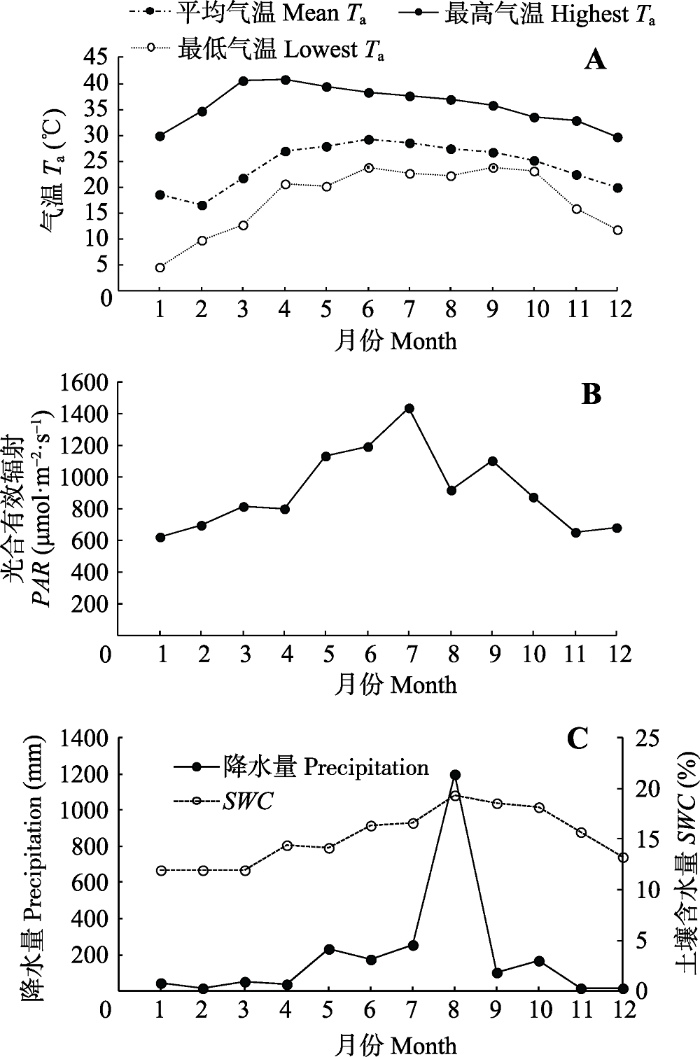 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1气温、光合有效辐射、降水和橡胶林下土壤含水量的季节变化。
Fig. 1Seasonal changes of air temperature (Ta), photosynthetic active radiation (PAR), precipitation and soil water content (SWC) under the Hevea brasiliensis forest.
1.2 研究方法
在自然条件下, 选择长势均匀的益智植株, 并选取生长良好无病虫害的益智功能叶片(枝条顶部向下数第3或第4片发育完全的新生叶片)进行光合参数的测定。在2016年3月、6月、9月和12月中旬选择连续晴天无云的天气, 用LI-6400便携式光合测量系统(LI-COR, Lincoln, USA)测量益智叶片的光合日变化、光响应曲线, 每个参数测定3片功能叶, 即3次重复。1.2.1 光合日变化测定
每间隔2 h (在8:00、10:00、12:00、14:00、16:00、18:00)测定益智功能叶片的气体交换参数, 测定参数包括净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、叶片PAR、蒸腾速率(Tr)、环境CO2浓度(Ca)、空气湿度(RH)、气温(Ta)等, 并据此计算气孔限制值(Ls) = 1 - Ci/Ca、瞬时水分利用效率(WUE) = Pn/Tr。叶室温度(Tleaf)和叶室内空气湿度(RH)为环境温湿度, 叶室光强为林下光合有效辐射(PAR), 气体流速500 μmol·s-1, 叶室CO2浓度为环境CO2浓度(Ca) (380-420 μmol·mol-1)。
1.2.2 光响应曲线测定
在9:00-11:00进行光响应曲线的测定。光响应曲线的测定用LI-6400-02B红蓝光源设定叶室中PAR强度分别为: 1500、1200、1000、800、600、400、200、150、120、90、60、30、0 μmol·m-2·s-1, 叶室温度和湿度为环境温湿度, 气体流速500 μmol·s-1, 测定前对叶片在饱和光强800 μmol·m-2·s-1下诱导30 min。每个光强下测定时间为3 min, 整个程序需要约40 min。以Pn为纵坐标、PAR为横坐标做曲线, 并根据叶子飘的直角双曲线修正模型(Ye, 2007)拟合数据得到最大净光合速率(Pnmax)、光饱和点(Is)、光补偿点(Ic)、暗呼吸速率(Rd)和表观量子效率(AQE)。
1.2.3 叶绿素组分测定
摘取测量完光合作用的功能叶片(枝条顶部向下数第3或第4片发育完全的新生叶片)用锡箔纸密封并带回实验室液氮低温保存(-80 ℃)。在室温(25 ℃)条件下进行叶绿素提取实验。为保证益智叶片叶绿素活性, 实验前将叶片从低温下取出, 室温条件下放置半小时。用直径1.5 cm的打孔器在益智叶片上避开主叶脉打下10个叶圆片, 按Lichtenthaler和Wellburn (1983)的方法测定叶绿素和类胡萝卜素的含量。
1.2.4 土壤含水量测定
在距离益智植株50 cm处, 利用土钻取0-10 cm土样, 共选3个样本, 每个样本3个重复。利用烘干法(105 ℃)测定土壤含水量。
1.2.5 比叶质量测定
用直径1.5 cm的打孔器在益智功能叶片(枝条顶部向下数第3或第4片发育完全的新生叶片)上避开主叶脉打下30个叶圆片, 在70 ℃条件下烘24 h至恒质量, 用电子天平(精度为0.0001 g)称质量, 然后计算比叶质量(LMA) = 叶片干质量/叶面积(g·m-2)。
1.3 数据处理
获得的数据采用Excel 2007进行处理和图表绘制, 采用DPS 7.55软件进行单因素方差分析, 多重比较用最小显著差异法。相关性分析采用Pearson双侧显著检验。2 结果和分析
2.1 不同季节环境因子日变化特征
橡胶林下PAR、Ta和RH日变化见图2。图2
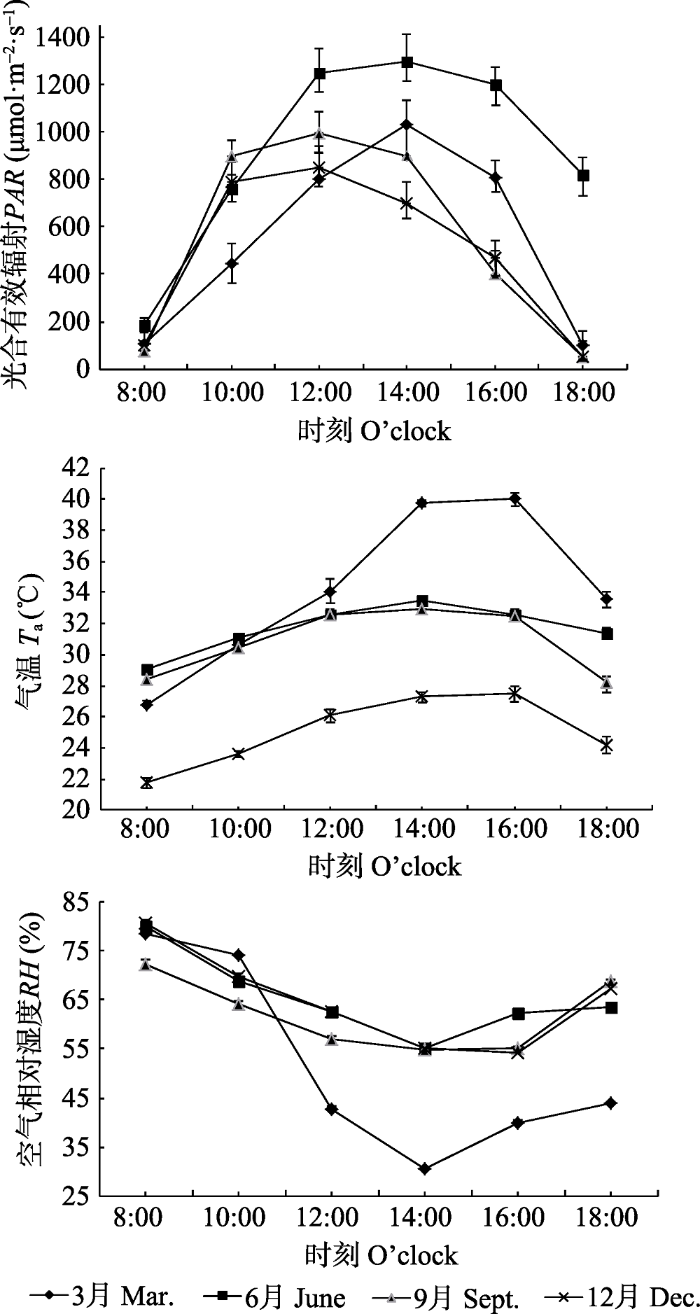 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2橡胶林下光合有效辐射、气温和空气相对湿度日变化(平均值±标准偏差)。
Fig. 2Daily variations of photosynthetic active radiation (PAR), air temperature (Ta), air humidity (RH) under the Hevea brasiliensis forest (mean ± SD).
不同季节橡胶林下环境因子PAR均呈单峰型曲线, 最大值出现在12:00-14:00, 最小值均出现在监测时段的8:00和18:00 (图2A), 这与太阳辐射一致。因为橡胶林冠的遮挡, 林下PAR还与橡胶树叶物候期的生长发育和太阳年辐射规律密切相关。
不同季节Ta随着PAR的增强而逐渐升高, 在14:00达到最高值, 而在16:00以后随着PAR的减小而下降(图2B)。其中6月和9月Ta日变化趋势一致, 10:00-16:00时段Ta无显著差异; 3月在14:00-16:00时段Ta峰值显著高于6月、9月和12月, Ta高达40.06 ℃, 这是由于3月出现了短暂的极端高温天气造成的; 12月的一天中各时段的Ta均低于6月、9月和12月, 最高气温仅为27.53 ℃。
不同季节RH均表现为早晨较高, 随着PAR和Ta的增高而下降, 14:00降到最低值, 随后又缓慢升高。6月、9月和12月的RH日变化趋势比较相似; 3月RH在10:00-14:00降幅显著大于6月、9月和12月, 14:00 RH仅为30.57%, 降幅达58.66% (图2C)。
2.2 不同季节益智气体交换参数日变化特征
不同季节中随着一天中PAR、Ta和RH的变化, 益智叶片气体交换参数Pn、Ci、Gs、LS、Tr和WUE也呈规律性的变化(图3)。图3
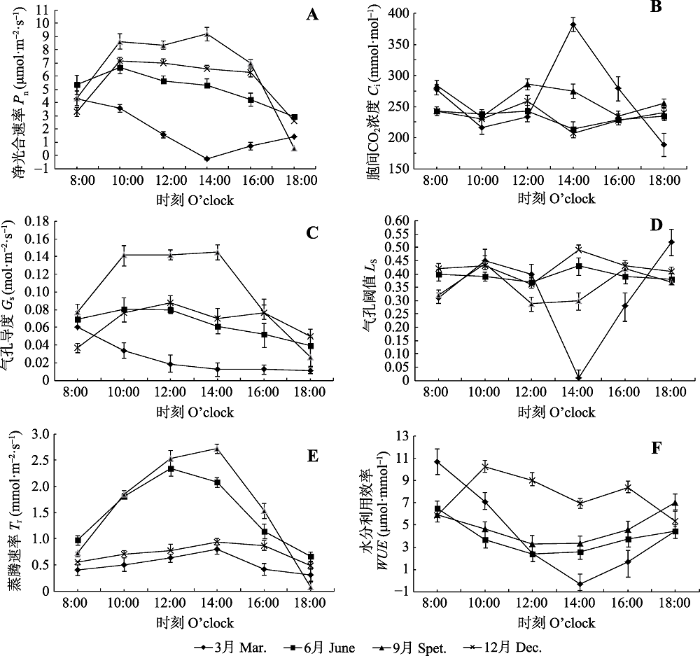 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同月份益智光合特性的日变化(平均值±标准偏差)。
Fig. 3Diurnal changes of photosynthetic characteristics of Alpinia oxyphylla in different months (mean ± SD). Ci, intercellular CO2 concentration; Gs, stomatal conductance; Ls, stomatal limitation; Pn, the net photosynthetic rate; Tr, transpiration rate; WUE, water use efficiency.
2.2.1 净光合速率和胞间CO2浓度
图3表明益智Pn在不同季节日变化趋势也不同, 6月和12月Pn在10:00达到最大值, 随后缓慢降低, 而9月Pn在10:00和14:00出现双峰现象; 最高值出现在14:00, 为9.19 μmol·m-2·s-1, 其峰值显著高于3月、6月和12月(p < 0. 05); 3月益智的Pn呈“V”形曲线, 8:00 Pn开始急剧下降, 14:00降到最低值, 为-0.27 μmol·m-2·s-1, 表明在旱季(3月) 14:00, 益智光合作用受到高温胁迫, 使得Rubisco活化酶活性降低, 造成叶肉光合能力下降。
胞间CO2浓度(Ci)是光合生理研究中最重要的一个参数。由图3可知, 不同季节Ci随着一天中环境因子的变化, 均出现上下波动的现象; 6月、9月和12月Ci呈“W”形曲线, 分别在10:00和14:00出现波谷; 而3月Ci在14:00急剧上升, 达到381.41 mmol·mol-1, 这主要是益智Pn受阻, 造成细胞内CO2积累的结果。
2.2.2 气孔导度和气孔阀值
气孔导度(Gs)与Pn密切相关, 6月、9月和12月益智Gs日变化与Pn日变化趋势基本一致; 而3月Gs在8:00-14:00缓慢下降到最低值, 为0.01 mol·m-2·s-1, 14:00-18:00变化不显著, 这主要是因为3月干旱少雨, 土壤含水量为全年最低水平, 呈现半干旱状态, 空气湿度小, 水分胁迫下降低了益智叶片的气孔开度以降低蒸腾作用, 减少水分流失。
气孔阀值(Ls)和Ci都是分析气孔限制和非气孔限制对光合速率影响的判断依据。由图3所示, 不同季节Ls的日变化与Ci呈相反的趋势。
2.2.3 蒸腾速率和水分利用率
不同季节益智叶片的蒸腾速率(Tr)日变化均呈单峰曲线, 早晨和晚上Tr较低, 12:00-14:00最高(图3)。雨季(6月和9月) Tr的日平均值和日最高值均显著高于旱季(3月和12月), 其中9月日平均值和日最高值分别为1.57和2.72 mmol·m-2·s-1, 分别是旱季(12月)日平均值和日最高值的2.18倍和2.89倍。
雨季(6月和9月)和旱季的3月, 益智叶片的水分利用效率(WUE)呈“V”形曲线, 早晨和傍晚WUE最高, 12:00-14:00最低, 与雨季(6月和9月)相比, 旱季3月WUE下降幅度更大; 3月8:00的WUE为10.69 μmol·mmol-1, 比6月高4.17 μmol·mmol-1, 3月14:00 WUE降到-0.33 μmol·mmol-1, 此时又比6月低2.89 μmol·mmol-1, 这可能是3月益智遭受土壤水分胁迫和极端高温影响, 气体交换速率减缓, 蒸腾作用和光合作用均受到抑制的结果。其中在3月14:00强光照和低土壤水分环境下, 能观察到益智叶片短暂缺水而蜷曲的现象, 这一形态变化与其WUE急剧下降为负值的实验结果相吻合。
2.3 益智叶片叶绿素含量、比叶质量和叶片含水量季节变化特征
从叶绿素各组分的季节变化(表1)中可看出, 3月益智叶片的总叶绿素、叶绿素b、类胡萝卜素含量均显著高于6月、9月和12月, 而叶绿素a/b显著低于6月、9月和12月(p < 0.05); 叶绿素a的含量在不同季节没有表现出显著差异(p > 0.05)。3月益智叶片LMA显著高于6月、9月和12月, 分别高了16.00%、4.77%和11.74%。益智叶片含水量季节变化趋势与叶片LMA变化趋势相反。Table 1
表1
表1益智叶片光和色素含量、比叶质量和叶片含水量
Table 1
| 叶绿素a Chl a (mg·cm-2) | 叶绿素b Chl b (mg·cm-2) | 类胡萝卜素 Car (mg·cm-2) | 总叶绿素 Chl (mg·cm-2) | 叶绿素a/b Chl a/b | 比叶质量 LMA (g·m-2) | 叶片含水量 Leaf water content (%) | |
|---|---|---|---|---|---|---|---|
| 3月 Mar. | 2.60a | 1.64a | 3.15a | 7.39a | 1.59a | 51.26a | 65.17c |
| 6月 June | 2.60a | 1.28b | 2.67c | 6.55b | 2.02b | 43.06c | 74.31a |
| 9月 Sept. | 2.62a | 1.33b | 2.60c | 6.55b | 1.97b | 48.82ab | 75.95a |
| 12月 Dec. | 2.52a | 1.31b | 2.76b | 6.48b | 1.93b | 45.25bc | 68.64b |
新窗口打开|下载CSV
2.4 光响应曲线季节变化特征
植物光响应曲线能反映净光合速率随光强变化的特征, 如图4所示, 与旱季相比较, 随着PAR的增加, 雨季益智叶片Pn显著增加。当PAR < 90 μmol·m-2·s-1时, 4个季节的益智叶片Pn线性递增, 光响应曲线变化规律差异不显著; PAR > 90 μmol·m-2·s-1时, 3月益智叶片光响应曲线拐点显著低于6月、9月和12月; 当PAR > 800 μmol·m-2·s-1时, 各季节的Pn趋于饱和状态。图4
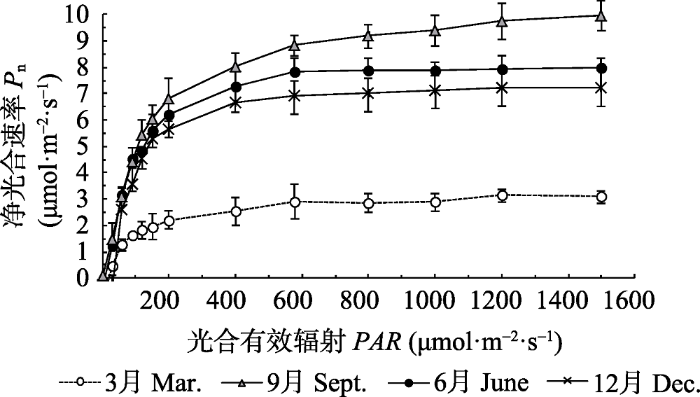 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同月份益智叶片的光响应曲线(平均值±标准偏差)。
Fig. 4Light response curves of net photosynthetic rate (Pn) in Alpinia oxyphylla in different months (mean ± SD).
由Ye (2007)的直角双曲线修正模型计算出益智叶片各光合响应参数, 如表2所示: 不同季节益智叶片Pnmax差异显著, 3月Pnmax最小, 仅为3.213 μmol·m-2·s-1, 仅是9月Pnmax的30%; 4个季节叶片的AQE大小顺序为3月< 12月< 9月< 6月; 3月份叶片的Ic和Rd均显著高于6月、9月和12月。表明旱季转入雨季后益智叶片线粒体呼吸速率下降, 导致Ic降低, 进一步提升了益智植株的光合能力。
Table 2
表2
表2不同月份益智叶片光响应曲线特征参数
Table 2
| 月份 Month | 表观量子效率 AQE | 光补偿点 Ic (μmol·m-2·s-1) | 暗呼吸速率 Rd (μmol·mol-1) | 最大净光合速率 Pnmax (μmol·m-2·s-1) | 光饱和点 Is (μmol·m-2·s-1) |
|---|---|---|---|---|---|
| 3月 Mar. | 0.068c | 16.144a | 0.865a | 3.213d | 522.968b |
| 6月 June | 0.100a | 5.813b | 0.550b | 8.006c | 1010.264a |
| 9月 Sept. | 0.095a | 3.514c | 0.324c | 10.648a | 1021.726a |
| 12月 Dec. | 0.086b | 3.906c | 0.326c | 8.783b | 964.900a |
新窗口打开|下载CSV
2.5 不同季节益智净光合速率与主要生理生态因子的关系
由表3可知, 不同季节影响益智植株Pn的主导因子是不同的。3月Ta和RH均与Pn呈极显著相关关系(p < 0.01), Ca和Gs与Pn呈显著相关关系(p < 0.05); 6月和9月Gs与Pn呈极显著相关关系(p < 0.01), 9月Ta和PAR与Pn呈显著相关关系(p < 0.05); 12月PAR与Pn呈极显著相关关系(p < 0.01), Gs与Pn呈显著相关关系(p < 0.05)。Table 3
表3
表3不同月份净光合速率(Pn)与主要生理生态因子的相关系数
Table 3
| 月份 Month | 生理生态因子 Physio-ecological factors | Pn | Ca | PAR | Ta | RH | Gs | Ci |
|---|---|---|---|---|---|---|---|---|
| 3月 Mar. | Pn | 1.000 | ||||||
| Ca | 0.883* | 1.000 | ||||||
| PAR | -0.678 | -0.664 | 1.000 | |||||
| Ta | -0.947** | -0.889* | 0.735 | 1.000 | ||||
| RH | 0.985** | 0.847* | -0.669 | -0.902* | 1.000 | |||
| Gs | 0.891* | 0.946** | -0.539 | -0.852* | 0.898* | 1.000 | ||
| Ci | -0.464 | -0.234 | 0.554 | 0.488 | -0.372 | -0.044 | 1.000 | |
| 6月 June | Pn | 1.000 | ||||||
| Ca | 0.692 | 1.000 | ||||||
| PAR | -0.349 | -0.868* | 1.000 | |||||
| Ta | -0.265 | -0.793 | 0.937** | 1.000 | ||||
| RH | 0.449 | 0.909* | -0.904* | -0.950** | 1.000 | |||
| Gs | 0.921** | 0.507 | -0.108 | -0.099 | 0.279 | 1.000 | ||
| Ci | 0.283 | 0.576 | -0.494 | -0.702 | 0.729 | 0.403 | 1.000 | |
| 9月 Sept. | Pn | 1.000 | ||||||
| Ca | -0.416 | 1.000 | ||||||
| PAR | 0.908* | -0.472 | 1.000 | |||||
| Ta | 0.821* | -0.665 | 0.733 | 1.000 | ||||
| RH | -0.716 | 0.837* | -0.678 | -0.960** | 1.000 | |||
| Gs | 0.940** | -0.310 | 0.941** | 0.665 | -0.549 | 1.000 | ||
| Ci | -0.009 | -0.037 | -0.055 | -0.067 | 0.112 | 0.174 | 1.000 | |
| 12月 Dec. | Pn | 1.000 | ||||||
| Ca | -0.536 | 1.000 | ||||||
| PAR | 0.969** | -0.470 | 1.000 | |||||
| Ta | 0.579 | -0.878* | 0.507 | 1.000 | ||||
| RH | -0.561 | 0.928** | -0.483 | -0.989** | 1.000 | |||
| Gs | 0.908* | -0.737 | 0.902* | 0.706 | -0.693 | 1.000 | ||
| Ci | -0.232 | 0.350 | -0.133 | -0.389 | 0.459 | -0.022 | 1.000 |
新窗口打开|下载CSV
3 讨论和结论
光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(Lewis et al., 2002)。随着季节更替, 植物光合作用的影响因素如PAR、空气温湿度及植物生理和结构因素等各方面也在不断发生变化, 这些变化必然引起植物光合特征随季节更替而变化。阴生植物一般光合速率偏低(Araus & Serret, 1986), 4个季节益智植株Pn的日平均值来看, 全年益智植株Pn较低, 具有喜阴性特点。除了3月Pn日平均值仅为1.86 μmol·m-2·s-1, 6月、9月和12月的Pn日平均值在5.18-6.33 μmol·m-2·s-1之间, 表明在雨季旱季更替下, 益智光合生理受到环境因子的显著影响。其中3月降水量极少, 造成土壤水分亏缺和空气湿度较低, 水分胁迫造成Gs降低, 导致3月全天益智叶片Pn处于极低的水平。在全天大部分时间里, 水分胁迫引起的Gs降低是低Pn的主导因子(许大全, 1997); 水分胁迫下除了气孔因素导致光合作用受抑制, 高温也是光合抑制的非气孔因素之一, 当环境温度高于35 ℃时, CO2同化、O2释放和光合磷酸化均遭受抑制(Lewis et al., 1999), 其中Rubisco活化酶是叶片的光合潜力主要限制酶(Crafts- Brandner & Salvucc, 2000), 3月, 14:00-16:00气温高达40.6 ℃, 受高温影响Pn降为负值, Ci却显著增高并达到全天的最大值(图3), 根据非气孔限制理论及其判别依据(郑义新等, 1995), 高温导致叶片叶肉细胞光合相关酶活性降低, 此时高温胁迫是光合碳同化受抑制的主导因子。6月、9月和12月的PAR、Pn和Gs日变化趋势基本一致(图2, 图3), 相关性分析结果显示9月PAR与Pn呈显著相关关系, 12月为极显著相关关系(表3), 即9月和12月PAR是益智光合作用的限制因子。为什么旱季的12月水分不是益智光合作用的限制因子呢?这主要是因为虽然旱季的12月降水量小, 土壤含水量偏低(轻度干旱)(图1), 但是12月有数量可观的雾降水补充(庞家平等, 2009), 缓解了土壤水分的不足, 因此益智生长和光合作用未受到水分的胁迫, 所以12月水分没有成为益智光合作用的限制因子。6月为旱季向雨季的过渡期, 这个时期的环境因子与Pn无相关性, 而Gs与Pn极显著相关(表3), 这个现象的机理有待进一步探讨。结合林下PAR季节变化(图1)发现, 旱季(3月)林下PAR较低, 促使益智增加叶绿素b的含量, 降低叶绿素a/b比值, 增强耐阴性, 以适应季节变化引起的低光环境。植物叶绿素各组分含量差异可用来表征叶片吸收太阳辐射的变化, 各种功能色素组分比值的变化, 也反映植物适应不同光环境的光合生理策略(Richardson et al., 2002)。叶绿素a含量决定植物的光合效率和能力, 叶绿素b在调控光合机构天线大小、维持其稳定性和提高其适应性中都起着重要作用。由表1可知, 3月益智叶片叶绿素b含量高于6月、9月和12月, 结合林下PAR季节变化(图1)发现, 旱季(3月)林下PAR较低, 促使益智增加叶绿素b的含量, 调控光合机构天线的大小, 提高光合机构对林下PAR的敏感性, 增强耐阴能力, 以适应季节变化引起的低光环境(Satoh et al., 2001)。LMA是表征植物生长过程中碳同化策略的关键叶性状指标(Wright et al., 2002)。大多数研究发现环境中光照越弱、叶片越薄、单位面积干质量越小, 比叶面积越大(左端阳等, 2014)。本研究中3月的LMA显著高于6月和9月, 这主要与橡胶树叶片物候期密切相关。3月橡胶树处于落叶期, 橡胶林叶面积指数较低、林下透光率高(吴志祥等, 2013); 同时3月极端高温天气和土壤水分亏缺, 在逆境环境下植物通过提高LMA减少光抑制、增加水分利用效率(徐飞等, 2010)。
光响应曲线中的AQE、Ic、Is、Rd等参数能反映环境条件及植物自身生理状况对光合作用的影响(Ogren, 1993)。植物Ic和Is的高低直接反映了植物对弱光的利用能力, 是植物耐阴性评价的重要指标(王建华等, 2011)。6-12月益智叶片的Is为968-1023 μmol·m-2·s-1, 而大多数C3阳生植物饱和光强在1000 μmol·m-2·s-1左右, 即益智表现出阳生植物的特性; 6月、9月和12月益智Ic为3.5-5.8 μmol·m-2·s-1, 具有极强的弱光利用能力而表现出极强的耐阴性(朱延姝等, 2010)。由此可知益智具有较强的利用弱光(漫射光、散射光、透射光)的能力, 其耐阴能力较强且光环境适应幅度较宽。3月时, 受到土壤水分亏缺的影响, AQE、Pnmax显著低于其他3个月份(表2), 有研究表明干旱胁迫或者盐胁迫条件会导致植物AQY的降低(Xia et al., 2011; Lin et al., 2015), 光能利用效率降低。其机理是植物体水分亏缺导致光合相关酶活性降低, 引起光合作用产生非竞争性抑制(Lin et al., 2015), 使Pn下降并随PAR的变化而光响应曲线拐点前移(图4), 从而导致AQE、Pnmax降低, 表现出光抑制现象。
综上所述, 益智属于典型的耐阴植物, 且能适应透光率较高的强光环境, 益智在不同季节环境因子变化下表现出不同的光能利用策略。在雨季的9月和旱季的12月, PAR是益智光合作用的主要影响因子; 在旱季的3月降水量低造成土壤水分含量不足是益智光合作用的主要限制生态因子, 中午高温和低空气湿度的环境因子是益智光合日变化出现“午休现象”的主要诱导因素。因此在生产实践中, 旱季时尤其是3月橡胶-益智间作体系应适当开展设施灌溉, 提高土壤含水率, 同时对土壤表面实施盖草措施以减少土壤水分的蒸发, 缓解土壤干旱状态, 保障益智植株正常的生长和光合作用。

致谢
感谢中国热带农业科学院基本科研业务费专项资金(17CXTD-04、NO.1630042017028、NO.2017hzs1J021)资助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]

Leaf structure, photosynthetic characteristics and related physiological parameters have been studied in three ornamental shade species: Fatsia japonica, Cissus rhombifolia (relatively light-tolerant plants), and Philodendron scandens (obligate shade plant). Species were grown in a shadehouse. Maximum photosynthetic photon flux density was 470 mol m-2s-1. Net rate of CO2uptake at light saturation (maximum Pn) in Fatsia was 6.90 1.27 mol m-2s-1. In Cissus and Philodendron values were about 30% and 63% less respectively, than those measured in Fatsia. The nitrogen content, relative dry wt, specific leaf dry wt (SLDW), chlorophyll a/b ratio, and nitrogen to chlorophyll ratio were lower in Philodendron. However, leaf thickness in Philodendron (296 17 m) was about 54% and 160% higher, respectively, than in Fatsia and Cissus, and the ratio between mesophyll cell area and leaf surface area (Ames/A) was nearly similar in the three species. However Philodendron exhibited a percentage of palisade parenchyma about three times lower than that observed in the two other species. The chloroplast number per mm of cell wall in transverse sections (chloroplast density) in the palisade parenchyma was fairly constant (about 65), irrespective of species. The "chloroplast density" in the spongy parenchyma of Philodendron was about 53% and 63%, respectively, of Fatsia and Cissus values. In Fatsia and Cissus chloroplast ultrastructure seems to change gradually and continuously from sun to shade type with the depth from the adaxial to abaxial surface. Special emphasis was given in order to determine the structural parameters best correlated with maximum Pn between the different species. In this way chloroplast number in transverse sections (chloroplast number) and the ratio between chloroplast area and leaf surface area (Achl/A) were the parameters best correlated with maximum Pn, and stomatal frequency was also a good determinant of maximum Pn. However, leaf thickness, SLDW, and even Ames/A ratio were weakly correlated with maximum Pn.
DOIURL [本文引用: 1]
橡胶林第一蓬叶的展叶期、稳定期和老化速度,极大影响橡胶树叶片的数量和质量,从而影响干胶产量,因此,监测橡胶林春季叶片生长情况对防治病虫害、指导割胶作业和林间水肥管理具有积极意义。本研究基于2001-2014年1-5月每16d的遥感增强型植被指数(EVI)分布图,通过计算相邻时段EVI的变化量,分析橡胶林MODIS/EVI指数的时空分布及变化特征,确定判断橡胶叶片春季物候期的EVI指标。结果表明:第一蓬叶展叶期橡胶林平均EVI值为0.4,稳定期平均EVI值为0.5,以此作为这两个生育期的判断标准,对2001-2014年逐年橡胶林第一蓬叶的展叶期和稳定期进行反演,结果表明,海南岛橡胶林叶片平均展叶期出现在3月中旬前期,稳定期主要出现在5月上旬,叶片老化平均历时50d,与历年人工观测结果相比有一定差异,平均偏晚8.2d。该方法简便易行,可以用来监测区域天然橡胶物候的变化态势。
DOIURL [本文引用: 1]
橡胶林第一蓬叶的展叶期、稳定期和老化速度,极大影响橡胶树叶片的数量和质量,从而影响干胶产量,因此,监测橡胶林春季叶片生长情况对防治病虫害、指导割胶作业和林间水肥管理具有积极意义。本研究基于2001-2014年1-5月每16d的遥感增强型植被指数(EVI)分布图,通过计算相邻时段EVI的变化量,分析橡胶林MODIS/EVI指数的时空分布及变化特征,确定判断橡胶叶片春季物候期的EVI指标。结果表明:第一蓬叶展叶期橡胶林平均EVI值为0.4,稳定期平均EVI值为0.5,以此作为这两个生育期的判断标准,对2001-2014年逐年橡胶林第一蓬叶的展叶期和稳定期进行反演,结果表明,海南岛橡胶林叶片平均展叶期出现在3月中旬前期,稳定期主要出现在5月上旬,叶片老化平均历时50d,与历年人工观测结果相比有一定差异,平均偏晚8.2d。该方法简便易行,可以用来监测区域天然橡胶物候的变化态势。
URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
分析近两年橡胶价格持续低迷背景下,海南省昌江县民营橡胶发展现状及发展中存在的问题,并提出相应的措施和对策,为昌江县民营橡胶发展决策提供参考。
URL [本文引用: 1]
分析近两年橡胶价格持续低迷背景下,海南省昌江县民营橡胶发展现状及发展中存在的问题,并提出相应的措施和对策,为昌江县民营橡胶发展决策提供参考。
DOIURLPMID [本文引用: 1]
Abstract Net photosynthesis (Pn) is inhibited by moderate heat stress. To elucidate the mechanism of inhibition, we examined the effects of temperature on gas exchange and ribulose 1,5-bisphosphate carboxylase/oxygenase (Rubisco) activation in cotton and tobacco leaves and compared the responses to those of the isolated enzymes. Depending on the CO(2) concentration, Pn decreased when temperatures exceeded 35-40 degrees C. This response was inconsistent with the response predicted from the properties of fully activated Rubisco. Rubisco deactivated in leaves when temperature was increased and also in response to high CO(2) or low O(2). The decrease in Rubisco activation occurred when leaf temperatures exceeded 35 degrees C, whereas the activities of isolated activase and Rubisco were highest at 42 degrees C and >50 degrees C, respectively. In the absence of activase, isolated Rubisco deactivated under catalytic conditions and the rate of deactivation increased with temperature but not with CO(2). The ability of activase to maintain or promote Rubisco activation in vitro also decreased with temperature but was not affected by CO(2). Increasing the activase/Rubisco ratio reduced Rubisco deactivation at higher temperatures. The results indicate that, as temperature increases, the rate of Rubisco deactivation exceeds the capacity of activase to promote activation. The decrease in Rubisco activation that occurred in leaves at high CO(2) was not caused by a faster rate of deactivation, but by reduced activase activity possibly in response to unfavorable ATP/ADP ratios. When adjustments were made for changes in activation state, the kinetic properties of Rubisco predicted the response of Pn at high temperature and CO(2).
DOIURL [本文引用: 1]
Coffee ( Coffea canephora var robusta ) is grown in Southwestern Togo under shade of native Albizia adianthifolia as a low input cropping system. However, there is no information on carbon and nutrient cycling in these shaded coffee systems. Hence, a study was conducted in a mature coffee plantation in Southwestern Togo to determine carbon and nutrient stocks in shaded versus open-grown coffee systems. Biomass of Albizia trees was predicted by allometry, whereas biomass of coffee bushes was estimated through destructive sampling. Above- and belowground biomass estimates were respectively, 14002Mg02ha 611 and 3202Mg02ha 611 in the coffee– Albizia association, and 29.702Mg02ha 611 and 18.702Mg02ha 611 in the open-grown system. Albizia trees contributed 87% of total aboveground biomass and 55% of total root biomass in the shaded coffee system. Individual coffee bushes consistently had higher biomass in the open-grown than in the shaded coffee system. Total C stock was 8102Mg02ha 611 in the shaded coffee system and only 22.902Mg02ha 611 for coffee grown in the open. Apart from P and Mg, considerable amounts of major nutrients were stored in the shade tree biomass in non-easily recyclable fractions. Plant tissues in the shaded coffee system had higher N concentration, suggesting possible N fixation. Given the potential for competition between the shade trees and coffee for nutrients, particularly in low soil fertility conditions, it is suggested that the shade trees be periodically pruned in order to increase organic matter addition and nutrient return to the soil.
DOIURL [本文引用: 1]
Two major components of climate change, increasing atmospheric [CO2] and increasing temperature, may substantially alter the effects of water availability to plants through effects on the rate of water loss from leaves. We examined the interactive effects of elevated [CO2] and temperature on seasonal patterns of stomatal conductance (g(s)), transpiration (E) and instantaneous transpiration efficiency (ITE) in Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) seedlings. Seedlings were grown in sunlit chambers at either ambient CO2 (AC) or ambient + 180 mum mol mol(-1) CO2 (EC), and at ambient temperature (AT) or ambient + 3.5 degreesC (ET) in a full-factorial design. Needle gas exchange at the target growth conditions was measured approximately monthly over 21 months. Across the study period and across temperature treatments, growth in elevated [CO2] decreased E by an average of 12% and increased ITE by an average of 46%. The absolute reduction of E associated with elevated [CO2] significantly increased with seasonal increases in the needle-to-air vapour pressure deficit (D). Across CO2 treatments, growth in elevated temperature increased E an average of 37%, and did not affect ITE. Combined, growth in elevated [CO2] and elevated temperature increased E an average of 19% compared with the ACAT treatment. The CO2 supply and growth temperature did not significantly affect stomatal sensitivity to D or the relationship between gs and net photosynthetic rates. This study suggests that elevated [CO2] may not completely ameliorate the effect of elevated temperature on E, and that climate change may substantially alter needle-level water loss and water use efficiency of Douglas-fir seedlings.
DOIURLPMID [本文引用: 1]
Abstract Increases in atmospheric CO(2) concentration and temperature are predicted to increase the light response of photosynthesis by increasing light-saturated photosynthetic rates and apparent quantum yields. We examined the interactive effects of elevated atmospheric CO(2) concentration and temperature on the light response of photosynthesis in Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) seedlings. Seedlings were grown in sunlit chambers controlled to track either ambient (~400 ppm) CO(2) or ambient + 200 ppm CO(2), at ambient temperature or ambient + 4 degrees C. Photosynthetic light response curves were measured over an 18-month period beginning 32 months after treatments were initiated. Light-response curves were measured at the growth CO(2) concentration, and were used to calculate the light-saturated rate of photosynthesis, light compensation point, quantum yield and respiration rate. Elevated CO(2) increased apparent quantum yields during two of five measurement periods, but did not significantly affect light-saturated net photosynthetic rates, light compensation points or respiration rates. Elevated temperature increased all parameters. There were no significant interactions between CO(2) concentration and temperature. We conclude that down-regulation of photosynthesis occurred in the elevated CO(2) treatments such that carbon uptake at a given irradiance was similar across CO(2) treatments. In contrast, increasing temperature may substantially increase carbon uptake rates in Douglas-fir, assuming other environmental factors do not limit photosynthesis; however, it is not clear whether the increased carbon uptake will increase growth rates or be offset by increased carbon efflux through respiration.
DOIURL [本文引用: 1]
DOIURLPMID [本文引用: 2]
Photosynthesis-irradiance (PI) curves are extensively used in field and laboratory research to evaluate the photon-use efficiency of plants. However, most existing models for PI curves focus on the relationship between the photosynthetic rate (Pn) and photosynthetically active radiation (PAR), and do not take account of the influence of environmental factors on the curve. In the present study, we used a new non-competitive inhibited Michaelis-Menten model (NIMM) to predict the co-variation of Pn, PAR, and the relative pollution index (I). We then evaluated the model with published data and our own experimental data. The results indicate that the Pn of plants decreased with increasingIin the environment and, as predicted, were all fitted well by the NIMM model. Therefore, our model provides a robust basis to evaluate and understand the influence of environmental pollution on plant photosynthesis.
DOIURL [本文引用: 1]
The environmental services that agroforestry practices can provide, and especially their potential contribution to the conservation of biodiversity, have only recently attracted wider attention among agroforestry and conservation scientists. This new view is consistent with the ecosystem approach to natural resource management advocated by the Convention on Biological Diversity. This collection of six papers, which is based on a Workshop held in June uly 2004, brings together studies of biodiversity impacts of traditional agroforestry practices from Central and South America, Africa and Asia. The contributions highlight the considerable potential of traditional agroforestry practices to support biodiversity conservation, but also show their limits. These include the importance of sufficient areas of natural habitat and of appropriate hunting regulations for maintaining high levels of biodiversity in agroforestry land use mosaics, as well as the critical role of markets for tree products and of a favourable policy environment for agroforestry land uses. In combination the case studies suggest that maintaining diversity in approaches to management of agroforestry systems, along with a pragmatic, undogmatic view on natural resource management, will provide the widest range of options for adapting to changing land use conditions.
DOIURL [本文引用: 1]
Improved understanding of the impact of leafing phenology and soil water supplies on the gas exchange and water use efficiency ( WUE ) of trees in agroforestry systems in the semi-arid tropics is essential to optimise tree/crop interactions and the use of scarce natural resources, particularly in view of recent initiatives to use carbon credits to reward subsistence farmers who plant and manage trees effectively. Understanding of the relationships between photosynthetic and transpiration rates and chlorophyll content may help to explain variation in growth rate, productivity and WUE between species and growth environments. Such information is lacking for the species examined here, the evergreen Grevillea robusta (A. Cunn), semi-deciduous Alnus acuminata (HBK) and deciduous Paulownia fortunei (Hemsl.). Although differences in leafing phenology influence the extent of competition or complementarity between trees and crops in agroforestry systems, and hence crop yield, farmers may deliberately plant fast-growing, competitive tree species if they provide attractive economic returns or carbon credit subsidies. The objectives were to characterise: (1) the extent of variation in foliar gas exchange and instantaneous water use efficiency ( WUE i ) between trees with contrasting leafing phenologies; (2) the influence of leaf age and available soil moisture on gas exchange and WUE i of trees; and (3) chlorophyll and foliar nitrogen (N) concentrations and gas exchange parameters for maize in agroforestry and sole crop systems. Foliar chlorophyll and N concentrations, assimilation ( A ) and transpiration rate ( E ) were much greater in P. fortunei than in A. acuminata or G. robusta . A , E and WUE i were greatest in young mature leaves of all three species but declined as they senesced. WUE i increased as soil moisture content declined and was similar in all species, suggesting their differing leafing phenologies were responsible for observed differences in long-term water use and growth. A and WUE i were greater in maize than in tree species, consistent with its C4 photosynthetic pathway. The implications for tree and crop growth, yield and acceptability to subsistence farmers in the semi-arid tropics are discussed.
DOIURL [本文引用: 1]
Nutrient buildup in the soil caused by increased animal manure and fertilizer use in agricultural and forestry practices may increase the potential for their loss from the soil, leading to groundwater contamination and nonpoint source pollution. Studies in the tropics have suggested that agroforestry practices can reduce such nutrient (especially nitrogen) losses because of enhanced nutrient uptake by tree and crop roots from varying soil depths, compared to more localized and shallow rooting depths of sole crop stands. In temperate systems, such benefits have been well documented for riparian forest buffer practices. Currently, other temperate agroforestry practices are also being considered for their potential to reduce runoff and leaching of chemicals and thereby improve environmental quality within the agricultural landscape. In this regard, the lorida P-Index, which considers both phosphorus transport characteristics and management practices, may be a useful tool in the evaluation of nutrient management practices and environmental benefits of agroforestry. Preliminary results from an alleycropping site and a silvopastoral site on two different soil types in Florida suggest that both of these agroforestry practices will likely reduce nutrient loss compared to conventional agricultural practices. The primary aspects of P-Index include consideration of transport factors such as soil erosion, soil runoff class, leaching potential, and distance from a water body along with management factors such as soil test P, P application method, and source and rate of P application. P-Index evaluation of these studies indicates that both agroforestry sites can be on a nitrogen-based nutrient management program. The relevance of some management practices such as application of manure vs. inorganic fertilizer is also discussed in light of the P-Index and the two agroforestry practices.
DOIURLPMID [本文引用: 1]
Abstract Photosynthesis in the intermediate light range is most efficient when the convexity of the photosynthetic light-response curve is high. Factors determining the convexity were examined for intact leaves using Salix sp. and for a plant cell culture using the green microalga Coccomyxa sp. It was found that the leaf had lower convexity than diluted plant cells because the light gradient through the leaf was not fully matched by a corresponding gradient in photosynthetic capacity. The degree to which the leaf gradients were matched was quantified by measuring photosynthesis at both leaf surfaces using modulated fluorescence. Two principal growth conditions were identified as those causing mismatch of leaf gradients and lowering of the convexity relative to cells. The first was growth under low light, where leaves did not develop any noteworthy gradient in photosynthetic capacity. This led to decreased convexity, particularly in old leaves with high chlorophyll content and, hence, steep light gradients. Second and less conspicuous was growth under high light conditions when light was given bilaterally rather than unilaterally, which yielded leaves of high photosynthetic capacity at both surfaces. Two situations were also identified that caused the convexity to decrease at the chloroplast level: (a) increased light during growth, for both leaves and cells, and (b) increased CO2 concentration during measurement of high-light-grown leaves. These changes of the intrinsic convexity were interpreted to indicate that the convexity declines with increased capacity of ribulose-1,5-bisphosphate carboxylase/oxygenase relative to the capacity of electron transport.
DOIURL [本文引用: 1]
Byline: P. OGWUCHE, H.Y. UMAR, T.U. ESEKHADE AND SUNDAY Y. FRANCIS ABSTRACTS The study revealed a three years intercropping experiment of rubber with some arable crops at Rubber Research Institute of Nigeria (RRIN), Iyanomo, Benin City, Nigeria. A stratified intercropping method was adopted on a hectare of plot with three years old rubber plantation while another one hectare was used as control plot. Cost and returns from the intercrops were determined using descriptive statistics, such as means and budgetary technique. A straight line depreciation method was used to estimate the fixed cost value of the farming tools used during the study. The results indicated that a total amount of - 193,310 was spent in the three years as cost of the intercropping, while income of N 763,330 was obtained from sales of total quantity, 26,078 kg produced in the three years from six different types of crops intercropped with rubber. These gave a gross margin and net profit of N 715,360 and N 640,020, respectively. The result also indicated a higher annual increase in girth of rubber intercropped compared to the sole rubber plantation. (c) 2012 Friends Science Publishers Key Words: Intercrop; Income flow; Budgetary techniques INTRODUCTION Natural rubber (Hevea brasiliensis) is a tropical perennial crop. It grows well on a well drain soil with temperature ranges from 21 to 30oC and rainfall of 2000 mm and above per annum (Umar et al., 2011). The plant is essentially grown for its latex content, which is converted to many elastic substances such as coagula, rubber smoked sheet, (RSS) etc used in the manufacture of some industrial products such as tires, hand grooves, condoms, tubes, shoes, etc., (Kpolo, 1999; RRIS, 2002; Rubber Asia, 2006). Natural rubber production in Nigeria is believed to have begun around the year 1876 with the exploitation of the local variety, Funtumia elastica (Umar and Ugwa, 2006). Funtumia elastica has poor yield (about 340 kg/ha/yr) and bark regeneration after tapping. These qualities caused discouragement of its production among rubber farmers. The discovery of Knuth Mull (Hevea brasiliensis) which arrived Nigeria from Kew garden in England in about 1895, which has better yield (3600 kg/ha/yr) and easy/quick bark regeneration after tapping (Ogowewo, 1989), renewed farmer's interest in the cultivation of natural rubber to the extent that before the discovery of crude oil in commercial quantity in Nigeria, natural rubber production ranked sixth among the economic cash crops in the country. The production trend however, experienced a staggered movement over the past 40 years. The situation became worse between 1980s and 1990s when Federal government of Nigeria shifted her emphasis for the nation's economic drive from agriculture to crude oil (Oyedipe, 2001). The repercussion of the government decision was not palatable to the country's economy and the general populace as poverty and social vices rose higher than ever due to many people became jobless; agriculture was given lesser priority and was generally unprofitable as government rather encouraged the system of importation of food items at far cheaper rates than their...
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
DOIURL [本文引用: 1]
Summary 61 68Over the last decade, technological developments have made it possible to quickly and nondestructively assess, in situ , the chlorophyll (Chl) status of plants. We evaluated the performance of these optical methods, which are based on the absorbance or reflectance of certain wavelengths of light by intact leaves. 61 68As our benchmark, we used standard extraction techniques to measure Chl a , Chl b , and total Chl content of paper birch ( Betula papyrifera ) leaves. These values were compared with the nominal Chl index values obtained with two hand-held Chl absorbance meters and several reflectance indices correlated with foliar Chl. 61 68The noninvasive optical methods all provided reliable estimates of relative leaf Chl. However, across the range of Chl contents studied (0.0004–0.0455mgcm 612 ), some reflectance indices consistently out-performed the hand-held meters. Most importantly, the reflectance indices that performed best were not those most commonly used in the literature. 61 68We report equations to convert from index values to actual Chl content, but caution that differences in leaf structure may necessitate species-specific calibration equations.
DOIURL [本文引用: 1]
Solar radiation is an important factor for plant growth, being its availability to understory crops strongly modified by trees in an Agroforestry System (AFS). Coffee trees (Coffea arabica - cv. Obat00 IAC 1669-20) were planted at a 3.4 x 0.9 m spacing inside and aside rows of monocrops of 12 year-old rubber trees (Hevea spp.), in Piracicaba-SP, Brazil (22024230 S, 47023800 W - altitude: 546m). One-year-old coffee plants exposed to 25; 30; 35; 40; 45; 80; 90; 95 and 100% of the total solar radiation were evaluated according to its biophysical parameters of solar radiation interception and capture. The Goudriaan (1977) adapted by Bernardes et al. (1998) model for radiation attenuation fit well to the measured data. Coffee plants tolerate a decrease in solar radiation availability to 50% without undergoing a reduction on growth and LAI, which was approximately 2m05.m-2 under this condition. Further reductions on the availability of solar radiation caused a reduction in LAI (1.5m05.m-2), thus poor land cover and solar radiation interception, resulting in growth reduction.brA radia0400o solar é um fator importante para o crescimento das plantas, sendo sua disponibilidade fortemente modificada pelas árvores em sistemas agroflorestais (SAFs). Foram estudadas plantas de café (Coffea arabica - cv. Obat00 IAC 1669-20) espa04adas 3,4 x 0,9 m em fileiras, dentro e ao lado de uma planta0400o de seringueira (Hevea spp.) de 12 anos de idade, em Piracicaba, SP, Brasil (22024230 S, 47023800 W - altitude: 546 m). Cafeeiros de 1 ano de idade expostos aos níveis de 25, 30, 35, 40, 45, 80, 90, 95 e 100% de radia0400o solar foram avaliados quanto aos seus par09metros biofísicos de intercepta0400o e captura de luz. O modelo de Goudriaan (1977) adaptado por Bernardes et al. (1998) para estimativa da radia0400o solar disponível ajustou-se bem aos dados medidos. Os cafeeiros toleraram diminui0400o de 50% na disponibilidade de radia0400o solar sem sofrer redu0401es no crescimento e IAF, sendo este de aproximadamente 2 m05.m-2, nessas condi0401es. Redu0401es na disponibilidade de luz acima desse valor levaram a uma diminui0400o do IAF (1,5 m05.m-2), resultando em menor cobertura do solo pelos cafeeiros e menor intercepta0400o da radia0400o solar, com conseqüentes diminui0401es no crescimento.
DOIURL [本文引用: 2]
Water uptake and use by plants are essentially energy processes that can be largely modified by percentage of soil cover, plant type; foliage area and its distribution; phenological stage and several environmental factors. Coffee trees (Coffea arabica - cv. Obat00 IAC 1669-20) in Agrforestry System (AFS) spaced 3.4x0.9m apart, were planted inside and along rows of 12- year-old rubber trees (Hevea spp.) in Piracicaba-SP, Brazil (22 4230 S, 47 3800 W - altitude: 546m). Sap flow of one-year-old coffee plants exposed to 35; 45; 80; 95 and 100% of total solar radiation was estimated by the heat balance technique (Dynamax Inc.). Coffee plants under shade showed greater water loss per unit of incident irradiance. On the other hand, plants in monocrop (full sun) had the least water loss per unit of incident irradiance. For the evaluated positions average water use was (gH2O.m-2Leaf area.MJ-1): 64.71; 67.75; 25.89; 33.54; 27.11 in Dec./2002 and 97.14; 72.50; 40.70; 32.78; 26.13 in Feb./2003. This fact may be attributed to the higher stomata sensitivity of the coffee plants under more illuminated conditions, thus plants under full sun presented the highest water use efficiency. Express transpiration by leaf mass can be a means to access plant adaptation to the various environments, which is inaccessible when the approach is made by leaf area.brA absor0400o e uso da água pelas plantas s00o processos essencialmente energéticos que podem ser grandemente modificados pela porcentagem de cobertura do solo, tipo de plantas, área foliar e sua distribui0400o, estágio fenológico e diversos fatores ambientais. Cafeeiros (Coffea arabica - cv. Obat00 IAC 1669-20) em Sistema Agroflorestal (AFS) espa04ados de 3.4 x 0.9 m foram plantados dentro e ao lado de um seringal de 12 anos de idade (Hevea spp.) em Piracicaba, SP, Brasil (22 4230S, 47 3800 W - altitude: 546 m). O fluxo de seiva dos cafeeiros com 1 ano de idade expostos a 35, 45, 80, 95 e 100% da radia0400o solar foi estimado pela técnica do balan04o de calor (Dynamax Inc.). Cafeeiros sombreados sofreram a maior perda de água por unidade de radia0400o incidente. No entanto, as plantas em monocultivo (pleno sol) apresentaram a menor perda de água por unidade de radia0400o incidente. Nas dist09ncias avaliadas, o uso médio de água foi (gH2O.m-2área foliar.MJ-1): 64,71; 67,75; 25,89; 33,54; e 27,11 em dez./2002 e 97,14; 72,50; 40,70; 32,78; e 26,13 em fev./2003. Tal fato pode ser atribuído à maior sensibilidade estomatal dos cafeeiros sob condi0401es de maior ilumina0400o, o que faz que plantas a pleno sol apresentem eficiência de uso da água mais elevada. Expressar a transpira0400o por massa de folha pode ser um meio de verificar a adapta0400o das plantas aos diversos ambientes, a qual é inacessível quando realizado por área foliar.
DOIURLPMID [本文引用: 1]
Photosynthetic pigments bind to their specific proteins to form pigment-protein complexes. To investigate the pigment-binding activities of the proteins, chlorophyll b was for introduced the first time to a cyanobacterium that did not synthesize that pigment, and expression of its function in the native pigment-protein complex of cyanobacterium was confirmed by energy transfer. Arabidopsis CAO (chlorophyll a oxygenase) cDNA was introduced into the genome of Synechocystis sp. PCC6803. The transformant cells accumulated chlorophyll b, with the chlorophyll b content being in the range of 1.4 to 10.6% of the total chlorophyll depending on the growth phase. Polyacrylamide gel electrophoresis analysis of the chlorophyll-protein complexes of transformant cells showed that chlorophyll b was
.
DOIURL
胶-茶群落在西双版纳地区有大面积分布,研究该群落中茶树的光合特性及其影响因子,可以为胶茶间作和茶树种植的实践提供理论依据.在自然条件下使用Li-6400便携式光合系统测定了西双版纳低(570m)、高(870m)两个不同海拔高度的胶-茶群落中茶树连体叶片的光合日进程,并对比研究了群落和纯林中茶树叶片的光响应.结果表明,两个群落中茶树叶片的最大净光合速率、暗呼吸速率、光补偿点和光饱和点均显著低于茶树纯林.两个群落中茶树的净光合速率日进程曲线均为“单峰”型,高海拔茶树的光量子通量密度、净光合速率、蒸腾速率和气孔导度均显著高于低海拔茶树的.相关性分析表明,光量子通量密度是引起群落中茶树光合差异的主要因素.群落林下光照的增强提高了茶树的净光合积累.在高海拔地区采用以茶树为主的胶茶种植结构将促进经济效益的提高.图3表3参24
DOIURL
胶-茶群落在西双版纳地区有大面积分布,研究该群落中茶树的光合特性及其影响因子,可以为胶茶间作和茶树种植的实践提供理论依据.在自然条件下使用Li-6400便携式光合系统测定了西双版纳低(570m)、高(870m)两个不同海拔高度的胶-茶群落中茶树连体叶片的光合日进程,并对比研究了群落和纯林中茶树叶片的光响应.结果表明,两个群落中茶树叶片的最大净光合速率、暗呼吸速率、光补偿点和光饱和点均显著低于茶树纯林.两个群落中茶树的净光合速率日进程曲线均为“单峰”型,高海拔茶树的光量子通量密度、净光合速率、蒸腾速率和气孔导度均显著高于低海拔茶树的.相关性分析表明,光量子通量密度是引起群落中茶树光合差异的主要因素.群落林下光照的增强提高了茶树的净光合积累.在高海拔地区采用以茶树为主的胶茶种植结构将促进经济效益的提高.图3表3参24
[本文引用: 1]
URL [本文引用: 1]
系统研究了不同程度的遮荫处理(0%、43%、70%、97%)对连翘叶片光合特性和叶绿素a荧光参数的影响。结果表明:随着遮荫程度增加,最大净光合速率、光补偿点、光饱和点、暗呼吸速率降低;净光合速率日变化均呈单峰型,峰值和光能利用率增加;叶绿素a+b、叶绿素a、叶绿素b含量增加,叶绿素a/b降低;叶绿素a荧光参数Fv/Fm和Fv/Fo日变化呈单谷曲线,值均高于全光照的,且随着遮荫程度的提高其值均依次增加。这说明,连翘是一种耐荫性很强的植物,遮荫可使其降低光补偿点、光饱和点、净光合速率、暗呼吸速率以及叶绿素a/b,增加总叶绿素、叶绿素a、叶绿素b含量、光能利用率以及PSⅡ原初光能转化效率和潜在活性,从而增强其在弱光条件下的生长发育能力。
URL [本文引用: 1]
系统研究了不同程度的遮荫处理(0%、43%、70%、97%)对连翘叶片光合特性和叶绿素a荧光参数的影响。结果表明:随着遮荫程度增加,最大净光合速率、光补偿点、光饱和点、暗呼吸速率降低;净光合速率日变化均呈单峰型,峰值和光能利用率增加;叶绿素a+b、叶绿素a、叶绿素b含量增加,叶绿素a/b降低;叶绿素a荧光参数Fv/Fm和Fv/Fo日变化呈单谷曲线,值均高于全光照的,且随着遮荫程度的提高其值均依次增加。这说明,连翘是一种耐荫性很强的植物,遮荫可使其降低光补偿点、光饱和点、净光合速率、暗呼吸速率以及叶绿素a/b,增加总叶绿素、叶绿素a、叶绿素b含量、光能利用率以及PSⅡ原初光能转化效率和潜在活性,从而增强其在弱光条件下的生长发育能力。
DOIURL [本文引用: 1]
DOIURL [本文引用: 2]
橡胶林生态系统光合有效辐射 (PAR)作为生态系统所有生命活动、有机物质合成和产量形成的最基本能量来源,是影响生态系统能量转化和物质生产的最重要生态因子,对其时空变异的研 究,有助于理解系统内生理生态因子变化和光合行为,为充分开发利用林下资源提供理论依据。本研究分析2010年海南岛橡胶林生态系统PAR的时空分布格局 及相互关系。结果表明,1)在日尺度上,橡胶林不同高度PAR日变化均呈现为相对比较规则的单峰型,最大值出现在正午前后;PAR从林冠顶层向下逐渐减 弱,平均日总量全光照条件下(16m高度)最大,为28.89mol.m-2,到12、8、4m和2m处PAR平均日总量分别占85.22%、 37.49%、17.48%和13.26%。2)在月尺度上,一年中橡胶林各层PAR月总量在7月达到最大值,在1月或者12月达到最小值。一年中PAR 雨季高于旱季,但表现略有差异,橡胶林林冠顶层(16m)和林冠层(12m)白昼时段均表现为雨季高于旱季,而冠下3层(8、4m和2m)只有正午前后几 小时雨季高于旱季,早晚时段则相反。3)橡胶林PAR年总量全光照条件下(16m)最大为10 543.75mol.m-2,往下逐渐减弱,12、8、4m和2m高度的光合有效辐射年总量分别为8 984.92、3 954.61、1 844.86mol.m-2和1 396.41mol.m-2,分别占到总量的85.21%、37.51%、17.50%和13.24%。4)橡胶林光合有效辐射透射率也因林冠等的遮挡, 年内变化明显,各垂直高度也表现不同。
DOIURL [本文引用: 2]
橡胶林生态系统光合有效辐射 (PAR)作为生态系统所有生命活动、有机物质合成和产量形成的最基本能量来源,是影响生态系统能量转化和物质生产的最重要生态因子,对其时空变异的研 究,有助于理解系统内生理生态因子变化和光合行为,为充分开发利用林下资源提供理论依据。本研究分析2010年海南岛橡胶林生态系统PAR的时空分布格局 及相互关系。结果表明,1)在日尺度上,橡胶林不同高度PAR日变化均呈现为相对比较规则的单峰型,最大值出现在正午前后;PAR从林冠顶层向下逐渐减 弱,平均日总量全光照条件下(16m高度)最大,为28.89mol.m-2,到12、8、4m和2m处PAR平均日总量分别占85.22%、 37.49%、17.48%和13.26%。2)在月尺度上,一年中橡胶林各层PAR月总量在7月达到最大值,在1月或者12月达到最小值。一年中PAR 雨季高于旱季,但表现略有差异,橡胶林林冠顶层(16m)和林冠层(12m)白昼时段均表现为雨季高于旱季,而冠下3层(8、4m和2m)只有正午前后几 小时雨季高于旱季,早晚时段则相反。3)橡胶林PAR年总量全光照条件下(16m)最大为10 543.75mol.m-2,往下逐渐减弱,12、8、4m和2m高度的光合有效辐射年总量分别为8 984.92、3 954.61、1 844.86mol.m-2和1 396.41mol.m-2,分别占到总量的85.21%、37.51%、17.50%和13.24%。4)橡胶林光合有效辐射透射率也因林冠等的遮挡, 年内变化明显,各垂直高度也表现不同。
DOIURL [本文引用: 1]
Campsis radicans (L.) Seem is one of the main forestation tree species in semi-arid loess hilly region. Using the CIRAS-2 portable photosynthesis system, the light-response of the photosynthetic efficiency parameters of three-year-old C .radicansleaves, such as net photosynthetic rate (PN), transpiration rate (Tr), water use efficiency (WUE), and light use efficiency (LUE), were studied under different soil water conditions in order to explore the effects of soil water stress on photosynthesis and the suitable soil water content for water-saving irrigation of this liana. Soil water and light intensity needed by the growth and development of C .radicans were analyzed. The results show that PN, Tr, WUE, and LUE of the leaves had threshold value to soil water and photosynthetically-active radiation (PAR). The non-rectangular hyperbola model was used to simulate light-response curve and the convexity was about 1. With the increase of soil relative water content of field capacity (Wr, ranged from 20.1% to 71.1%), the light compensation point declined while light saturation point, the maximum PN and apparent quantum yield increased. When Wr was about 71.7%, the light compensation point was at the minimum (21.61 mol/m2/s) and the light saturation point was at the maximum (1400 mol/m2/s). In order to maintain the normal plant growth and have higher PN, LUE, and WUE synchronously, the range of Wr was from 49.5 to 71.1%. The optimum Wr was 71.1%, and the minimum Wr was 28.2% for the normal growth of C .radicans. The high PN and WUE were recorded when PAR ranged from 800 to 1600 mol/m2/s and the light saturation points ranged from 800 to 1400 mol/m2/s. The peak value of LUE was found when PAR ranged from 100 to 300 mol/m2/s, indicating that the C. radicans had high adaptability to light conditions. Key words: Campsis radicans, soil water content, photosynthetically-active radiation, photosynthetic efficiency, water use efficiency
[本文引用: 1]
简要分析评论了光合作用气孔限制分析中的一些似是而非的观念及其作法的不可靠性。
[本文引用: 1]
简要分析评论了光合作用气孔限制分析中的一些似是而非的观念及其作法的不可靠性。
DOIURL [本文引用: 1]
通过搭建遮阳棚设置3个光照强度,模拟森林幼苗生长的林缘、林窗和林下光环境,研究麻栎和刺槐幼苗形态结构、光合特性、生物量积累及其分配对不同光照强度的响应。结果表明,极度弱光环境限制了幼苗的株高、基径、总叶面积、冠面积、叶面积指数和总叶数等形态指标的增长,与全光环境相比,适度遮荫有利于幼苗的形态生长,光照对麻栎形态的异速生长曲线影响较小,刺槐较麻栎的曲线变化更明显一些,弱光降低了异速生长指数。随遮荫程度的增加,麻栎的净光合速率、蒸腾速率、气孔导度逐渐降低,刺槐的净光合速率则在适度遮荫下最大,两者的光能利用效率均显著增大,水分利用效率先升后降。随光照强度的减弱,麻栎各器官的生物量积累降低,叶面积比率的增加和根冠比的减少有利于增加光能的捕获,而刺槐的生物量积累和分配则在适度遮荫下达到最优;各叶绿素含量均显著上升,而叶绿素a/b则逐渐下降。麻栎和刺槐显示出不同的生存策略和光利用策略,在一定程度上可以揭示它们在阔叶林群落中不同的演替地位。
DOIURL [本文引用: 1]
通过搭建遮阳棚设置3个光照强度,模拟森林幼苗生长的林缘、林窗和林下光环境,研究麻栎和刺槐幼苗形态结构、光合特性、生物量积累及其分配对不同光照强度的响应。结果表明,极度弱光环境限制了幼苗的株高、基径、总叶面积、冠面积、叶面积指数和总叶数等形态指标的增长,与全光环境相比,适度遮荫有利于幼苗的形态生长,光照对麻栎形态的异速生长曲线影响较小,刺槐较麻栎的曲线变化更明显一些,弱光降低了异速生长指数。随遮荫程度的增加,麻栎的净光合速率、蒸腾速率、气孔导度逐渐降低,刺槐的净光合速率则在适度遮荫下最大,两者的光能利用效率均显著增大,水分利用效率先升后降。随光照强度的减弱,麻栎各器官的生物量积累降低,叶面积比率的增加和根冠比的减少有利于增加光能的捕获,而刺槐的生物量积累和分配则在适度遮荫下达到最优;各叶绿素含量均显著上升,而叶绿素a/b则逐渐下降。麻栎和刺槐显示出不同的生存策略和光利用策略,在一定程度上可以揭示它们在阔叶林群落中不同的演替地位。
DOIURL [本文引用: 2]
The calculated maximum net photosynthetic rate ( P N ) at saturation irradiance ( I m ) of 1 314.13 08mol m 612 s 611 was 25.49 08mol(CO 2 ) m 612 s 611 , and intrinsic quantum yield at zero irradiance was 0.103. The results fitted by nonrectangular hyperbolic model, rectangular hyperbolic method, binomial regression method, and the new model were compared. The maximum P N values calculated by nonrectangular hyperbolic model and rectangular hyperbolic model were higher than the measured values, and the I m calculated by nonrectangular hyperbolic model and rectangular hyperbolic model were less than measured values. Results fitted by new model showed that the response curve of P N to I was nonlinear at low I for Oryza sativa , P N increased nonlinearly with I below saturation value. Above this value, P N decreased nonlinearly with I .
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Relationships between photosynthetic capacity and leaf structure in several shade plants
1
1986
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
海南岛天然橡胶林春季物候期的遥感监测
1
2016
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
海南岛天然橡胶林春季物候期的遥感监测
1
2016
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
不同采收期对药用植物——益智种子质量的影响研究
1
2017
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
不同采收期对药用植物——益智种子质量的影响研究
1
2017
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
胶价低迷背景下昌江县民营橡胶产业的发展现状和对策
1
2015
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
胶价低迷背景下昌江县民营橡胶产业的发展现状和对策
1
2015
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
Rubisco activase constrains the photosynthetic potential of leaves at high temperature and CO2
1
2000
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
Above- and belowground biomass, nutrient and carbon stocks contrasting an open-grown and a shaded coffee plantation
1
2008
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
Seasonal patterns of photosynthesis in Douglas fir seedlings during the third and fourth year of exposure to elevated CO2 and temperature
1
2002
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
Seasonal patterns of photosynthetic light response in Douglas-fir seedlings subjected to elevated atmospheric CO2 and temperature
1
1999
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
Determination of total carotenoids and chlorophylls a and b of leaf in different solvents
1
1983
... 摘取测量完光合作用的功能叶片(枝条顶部向下数第3或第4片发育完全的新生叶片)用锡箔纸密封并带回实验室液氮低温保存(-80 ℃).在室温(25 ℃)条件下进行叶绿素提取实验.为保证益智叶片叶绿素活性, 实验前将叶片从低温下取出, 室温条件下放置半小时.用直径1.5 cm的打孔器在益智叶片上避开主叶脉打下10个叶圆片, 按
Plant photosynthesis-irradiance curve responses to pollution show non-competitive inhibited Michaelis kinetics
2
2015
... 光响应曲线中的AQE、Ic、Is、Rd等参数能反映环境条件及植物自身生理状况对光合作用的影响(
... ), 光能利用效率降低.其机理是植物体水分亏缺导致光合相关酶活性降低, 引起光合作用产生非竞争性抑制(
Agroforestry and biodiversity conservation-traditional practices, present dynamics, and lessons for the future
1
2006
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
Gas exchange and water use efficiency of trees and maize in agroforestry systems in semi-arid Kenya
1
2009
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
Agroforestry as an approach to minimizing nutrient loss from heavily fertilized soils: The Florida experience
1
2004
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
Convexity of the photosynthetic light-response curve in relation to intensity and direction of light during growth
1
1993
... 光响应曲线中的AQE、Ic、Is、Rd等参数能反映环境条件及植物自身生理状况对光合作用的影响(
Economies of intercropping natural rubber with arable crops: A panacea for poverty alleviation of rubber farmers
1
2012
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
橡胶-大叶千斤拔复合生态系统中的植物生长与土壤水分养分动态
1
2009
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
橡胶-大叶千斤拔复合生态系统中的植物生长与土壤水分养分动态
1
2009
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
海南西部低割龄橡胶林土壤水分季节变化特征及其对气象因子响应研究初报
1
2017
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
海南西部低割龄橡胶林土壤水分季节变化特征及其对气象因子响应研究初报
1
2017
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
An evaluation of noninvasive methods to estimate foliar chlorophyll content
1
2002
... 结合林下PAR季节变化(
Measurement and simulation of solar radiation availability in relation to the growth of coffee plants in an agroforestry system with rubber trees
1
2007
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
Coffee water use in agroforestry system with rubber trees
2
2008
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
... )分别研究了不同光照和水分梯度下咖啡树的生长状况, 获得了林下咖啡树生长的最佳光照和水分条件.
Chlorophyll b expressed in cyanobacteria functions as a light-harvesting antenna in photosystem I through flexibility of the proteins
1
2001
... 结合林下PAR季节变化(
西双版纳胶-茶群落中茶树的光合特性及其影响因子
2008
西双版纳胶-茶群落中茶树的光合特性及其影响因子
2008
Sensitivity of C sequestration in reintroduced Sphagnum to water-level variation in a cutaway peatland
1
2004
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
遮荫对连翘光合特性和叶绿素荧光参数的影响
1
2011
... 光响应曲线中的AQE、Ic、Is、Rd等参数能反映环境条件及植物自身生理状况对光合作用的影响(
遮荫对连翘光合特性和叶绿素荧光参数的影响
1
2011
... 光响应曲线中的AQE、Ic、Is、Rd等参数能反映环境条件及植物自身生理状况对光合作用的影响(
Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span
1
2002
... 结合林下PAR季节变化(
海南岛橡胶林光合有效辐射的时空分布
2
2013
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
... 结合林下PAR季节变化(
海南岛橡胶林光合有效辐射的时空分布
2
2013
... 光合作用固定CO2是植物物质生产的基础, 也是全球碳循环的重要环节(
... 结合林下PAR季节变化(
Critical responses of photosynthetic efficiency in Campsis radicans( L.) Seem to soil water and light intensities
1
2011
... 光响应曲线中的AQE、Ic、Is、Rd等参数能反映环境条件及植物自身生理状况对光合作用的影响(
光合作用气孔限制分析中的一些问题
1
1997
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
光合作用气孔限制分析中的一些问题
1
1997
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
不同光环境对麻栎和刺槐幼苗生长和光合特征的影响
1
2010
... 结合林下PAR季节变化(
不同光环境对麻栎和刺槐幼苗生长和光合特征的影响
1
2010
... 结合林下PAR季节变化(
A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa
2
2007
... 在9:00-11:00进行光响应曲线的测定.光响应曲线的测定用LI-6400-02B红蓝光源设定叶室中PAR强度分别为: 1500、1200、1000、800、600、400、200、150、120、90、60、30、0 μmol·m-2·s-1, 叶室温度和湿度为环境温湿度, 气体流速500 μmol·s-1, 测定前对叶片在饱和光强800 μmol·m-2·s-1下诱导30 min.每个光强下测定时间为3 min, 整个程序需要约40 min.以Pn为纵坐标、PAR为横坐标做曲线, 并根据叶子飘的直角双曲线修正模型(
...
低丘红壤区农林间作系统的水分生态特征及生产力
1
1997
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
低丘红壤区农林间作系统的水分生态特征及生产力
1
1997
... 利用农林复合模式发展生态农业不仅可提高养分吸收利用效率、减缓水土流失、维持局部小气候稳定性、提高生物多样性、增加碳储量, 而且能充分利用水肥光热资源、提高系统生产力(
水分胁迫下植物叶片光合的气孔和非气孔限制
1
1995
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
水分胁迫下植物叶片光合的气孔和非气孔限制
1
1995
... 光合作用是植物生长发育中非常重要的生理过程, 受环境条件的长期影响, 植物从生理到形态形成了一系列应对环境变化的适应对策(
弱光胁迫对不同生育期番茄光合特性的影响
1
2010
... 光响应曲线中的AQE、Ic、Is、Rd等参数能反映环境条件及植物自身生理状况对光合作用的影响(
弱光胁迫对不同生育期番茄光合特性的影响
1
2010
... 光响应曲线中的AQE、Ic、Is、Rd等参数能反映环境条件及植物自身生理状况对光合作用的影响(
三七(Panax notoginseng)对不同光照强度的生理生态适应性研究
1
2014
... 结合林下PAR季节变化(
三七(Panax notoginseng)对不同光照强度的生理生态适应性研究
1
2014
... 结合林下PAR季节变化(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}